INFOR
chilonis
DALAM
RMASI DA
Ishii (HYM
M KAITAN
DEPAR
INS
ASAR PAR
MENOPT
NNYA DE
MIFTARTEMEN
FAKULT
STITUT P
RASITOID
TERA: TR
NGAN PE
AHUL JANPROTEK
TAS PERT
ERTANIA
BOGOR
2010
D TELUR
RICHOGR
ENGENDA
NNAHKSI TANA
TANIAN
AN BOGO
R Trichogr
RAMMAT
ALIAN H
AMAN
OR
ramma
TIDAE)
HAYATI
ABSTRAK
MIFTAHUL JANNAH. Informasi Dasar Parasitoid Telur Trichogramma chilonis Ishii (Hymenoptera: Trichogrammatidae) dalam Kaitannya dengan Pengendalian Hayati. Dibimbing oleh DAMAYANTI BUCHORI.
Hama penggerek batang merupakan hama yang paling penting pada tanaman tebu. Musuh alami yang biasa digunakan untuk mengendalikan hama ini adalah parasitoid telur Trichogramma chilonis. Informasi mengenai kebugaran parasitoid ini diperlukan untuk mengetahui kemampuan memarasit hama di lapangan, namun selama ini belum dilakukan pengujian kebugaran T. chilonis di laboratorium. Penelitian ini dilakukan untuk mempelajari kebugaran T. chilonis, kisaran inang dan preferensinya. Parameter kebugaran yang dimaksud diantaranya tingkat parasitisasi, produksi telur, lama hidup, nisbah kelamin, lama perkembangan hidup, persentase laju pemunculan imago dan kematian pradewasa. T. chilonis yang akan diuji berasal dari Lampung. Jenis serangga inang yang digunakan adalah Corcyra cephalonica, Chilo auricilius, Crocidolomia binotalis, Spodoptera litura dan Helicoverpa armigera. Telur inang ditempelkan pada pias dan diletakkan dalam tabung reaksi bersama T. Chilonis untuk diparasit. Pada uji preferensi lama pemaparan parasitoid pada inang ditentukan, yaitu 30 menit dan 24 jam. Penelitian ini menggunakan rancangan acak lengkap (RAL), data dianalisis dengan analisis ragam dan dilakukan uji Tukey pada taraf nyata 5%. Hasil penelitian menunjukkan bahwa kebugaran T. chilonis lebih baik pada inang C. cephalonica daripada C. auricilius. T. chilonis yang bersifat polifag terbukti mampu memarasit kelima jenis inang meskipun dengan tingkat parasitisasi yang bervariasi. Preferensi tertinggi terjadi pada inang H. armigera, disebabkan oleh ukuran telur inang yang relatif besar dan kualitas telur yang lebih baik daripada empat jenis inang lainnya.
INFOR
chilonis
DALAM
sebagaiRMASI DA
Ishii (HYM
M KAITAN
i salah satu sDEPAR
INS
ASAR PAR
MENOPT
NNYA DE
MIFTA syarat untuk DepartemeRTEMEN
FAKULT
STITUT P
RASITOID
TERA: TR
NGAN PE
AHUL JAN Skripsi memperoleh en Proteksi TPROTEK
TAS PERT
ERTANIA
BOGOR
2010
D TELUR
RICHOGR
ENGENDA
NNAH h gelar Sarja TanamanKSI TANA
TANIAN
AN BOGO
R Trichogr
RAMMAT
ALIAN H
ana PertanianAMAN
OR
ramma
TIDAE)
HAYATI
n padaJudul Penelitian : Informasi Dasar Parasitoid Telur Trichogramma chilonis Ishii (Hymenoptera: Trichogrammatidae) dalam Kaitannya dengan Pengendalian Hayati
Nama Mahasiswa : Miftahul Jannah NRP : A34061055
Disetujui Dosen Pembimbing
Dr. Ir. Damayanti Buchori, MSc NIP. 19601028198603 2 002
Diketahui Plh. Ketua Departemen
Dr. Ir. Abdjad Asih Nawangsih, MSi NIP. 19650621198910 2 001
RIWAYAT HIDUP
Penulis dilahirkan di Way Galih, Lampung pada tanggal 26 Juni 1989. Penulis adalah anak pertama dari tiga bersaudara dari pasangan Mulyanto dan Marsidah. Setelah lulus dari SMA Negeri 2 Bandar Lampung, penulis diterima sebagai mahasiswa Institut Pertanian Bogor melalui jalur USMI (Undangan Seleksi Masuk IPB). Pada tingkat dua, penulis diterima di Mayor Departemen Proteksi Tanaman, Fakultas Pertanian dengan Minor Ekonomi Pertanian, Departemen Ekonomi Sumberdaya dan Lingkungan, Fakultas Ekonomi dan Manajemen, Institut Pertanian Bogor.
Selama mengikuti pendidikan di Institut Pertanian Bogor, penulis aktif di UKM (Unit Kegiatan Mahasiswa) Gentra Kaheman dan berbagai kepanitian seperti MPKMB (Masa Perkenalan Kampus Mahasiswa Baru), OH DPT (Open House Departemen Proteksi Tanaman), MPD (Masa Perkenalan Departemen) dan Green Competition. Selain itu penulis juga aktif mengikuti berbagai seminar yang diadakan di lingkungan kampus dan pernah menjadi asisten dosen pada mata kuliah Dasar-dasar Proteksi Tanaman, Hama dan Penyakit Tanaman Setahun serta Pemanfaatan dan Pengelolaan Pestisida. Penulis juga memiliki pengalaman magang selama satu bulan di PT. Sweet Indolampung yang bergerak di bidang industri gula.
PRAKATA
Puji syukur penulis panjatkan ke Hadirat Allah SWT, karena berkat rahmat dan hidayah-Nya penulis dapat menyelesaikan skripsi yang berjudul “Informasi Dasar Parasitoid Telur Trichogramma chilonis Ishii (Hymenoptera: Trichogrammatidae) dalam Kaitannya dengan Pengendalian Hayati” dengan baik.
Penulisan skripsi ini bertujuan memenuhi syarat untuk memperoleh gelar Sarjana Pertanian. Penelitian dilakukan dari bulan Februari hingga Juni 2010 di Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Institut Pertanian Bogor. Dana penelitian bersumber dari dana yang diberikan oleh PT. Sweet Indolampung yang tergabung dalam Sugar Group Companies. Penulisan skripsi ini banyak dibantu oleh berbagai pihak baik secara moril maupun materiil. Oleh karena itu penulis mengucapkan terimakasih kepada:
1. Dr. Ir. Damayanti Buchori, MSc. selaku Dosen Pembimbing yang telah banyak meluangkan waktunya untuk memberikan arahan, masukan, bimbingan, saran serta motivasi selama penulisan skripsi.
2. Ibu Adha Sari yang telah membantu dalam mencari pustaka rujukan dan memberi masukan serta saran.
3. Orang tua dan adik-adik yang terus memberikan motivasi.
4. Teman-teman, sahabat serta orang terdekat yang senantiasa memberikan dukungan moril.
5. Serta semua pihak yang telah membantu dalam proses penyelesaian skripsi ini.
Penulis menyadari masih terdapat banyak kekurangan dalam penulisan skripsi ini, untuk itu diharapkan kritik dan saran yang dapat membangun penulis agar lebih baik dalam penulisan-penulisan selanjutnya. Skripsi ini diharapkan bermanfaat bagi pihak-pihak yang berkepentingan.
Bogor, September 2010
DAFTAR ISI
Halaman
DAFTAR TABEL... vii
DAFTAR GAMBAR... viii
DAFTAR LAMPIRAN... ix PENDAHULUAN... 1 Latar Belakang... 1 Tujuan... 3 Hipotesis... 3 Manfaat... 3 TINJAUAN PUSTAKA... 4
Pemanfaatan Parasitoid dari Genus Trichogramma... 4
Trichogramma chilonis Ishii... 5
Corcyra cephalonica Stainton... 6
Helicoverpa (Heliothis) armigera Hubner... 7
Crocidolomia binotalis Zell... 8
Spodoptera litura Fabricius... 8
Chilo auricilius Dudgeon... 9
Pengaruh Jenis Inang yang Berbeda terhadap Kebugaran Parasitoid... 10
Kisaran Inang dan Preferensi Parasitoid Telur... 11
BAHAN DAN METODE... 13
Tempat dan Waktu Penelitian... 13
Bahan, Alat dan Metode Penelitian... 13
Perbanyakan Inang... 13
Pembuatan Pias... 15
Perbanyakan Parasitoid telur... 15
Pengujian Kebugaran T. chilonis pada Dua Jenis Inang... 16
Pengujian Preferensi T. chilonis... 18
Analisis data... 18
HASIL DAN PEMBAHASAN... 19
Kebugaran T. chilonis pada Dua Jenis Inang... 19
Tingkat Parasitisasi... 19 Lama Hidup... 21 Produksi Telur... 22 Lama Perkembangan... 22 Kematian Pradewasa... 23 Nisbah Kelamin... 24
Persentase Laju Pemunculan Imago... 26
Kisaran Inang Parasitoid Telur T. chilonis... 26
Tingkat Parasitisasi... 27
Produksi Telur... 28
Laju Survival... 29
Preferensi Parasitoid Telur T. chilonis pada Lima Jenis Inang... 30
Tingkat Parasitisasi... 30
Produksi Telur... 32
Pembahasan Umum... 34
KESIMPULAN DAN SARAN... 38
Kesimpulan... 38
Saran... 38
DAFTAR PUSTAKA... 39
DAFTAR TABEL
No. Halaman
1. Tingkat parasitisasi, lama hidup dan produksi telur T. chilonis pada
dua jenis inang... 20 2. Lama perkembangan, persentase total betina dan persentase pupa mati
T. chilonis pada dua jenis inang... 24 3. Produksi telur, persentase parasitisasi dan persentase laju survival
pada lima jenis inang... 28 4. Persentase total parasitisasi T. chilonis pada lima jenis inang... 32
DAFTAR GAMBAR
No. Halaman
1. Persentase total imago betina T. chilonis pada dua jenis inang... 25 2. Persentase laju pemunculan imago T. chilonis pada dua jenis inang... 26 3. Persentase parasitisasi T. chilonis pada lima jenis inang yang
dipaparkan selama 30 menit dan 24 jam... 31 4. Produksi telur T. chilonis pada lima jenis inang yang dipaparkan
DAFTAR LAMPIRAN
No. Halaman
1. Persentase parasitisasi T. chilonis pada lima jenis inang yang
dipaparkan selama 30 menit dan 24 jam... 44 2. Produksi telur T. chilonis pada lima jenis inang yang dipaparkan
selama 30 menit dan 24 jam... 44 3. Kebugaran T. chilonis pada inang C. auricilius... 45 4. Kebugaran T. chilonis pada inang C. cephalonica... 47
PENDAHULUAN
Latar Belakang
Hama penggerek batang Chilo auricilius (Hymenoptera: Crambidae) merupakan hama yang paling penting pada tanaman tebu. Gejala yang ditimbulkan pada tanaman terserang yaitu pucuk daun layu dan kering, pada pangkal batang terdapat ulat, pada batang terdapat bekas gerekan dan biasanya terus tumbuh anakan baru (Matnawy 1989). Serangan hama ini mampu menurunkan rendemen tebu hingga lebih dari 30% sehingga tingkat produksi gula menurun. Turunnya angka rendemen tebu menyebabkan kerugian petani.
Penggunaan insektisida sintetik mampu memberikan hasil yang cukup signifikan dalam menekan populasi hama penggerek batang, namun menimbulkan berbagai dampak negatif. Menurut beberapa ahli bioekologi, penggunaan insektisida sintetik mampu menimbulkan resistensi hama, terjadinya peledakan hama, timbulnya hama sekunder, kontaminasi lingkungan, efek residu pada hasil pertanian dan gangguan kesehatan manusia (Kusnaedi 1997). Telah dilaporkan bahwa penggunaan karbofuran menyebabkan degradasi biologi dalam tanah lebih cepat daripada penggunaan karbamat (Smith & Pimentel 1978).
Adanya dampak negatif dari penggunaan insektisida sintetik menyebabkan para ahli entomologi memikirkan cara-cara baru dalam pengendalian hama yang aman dan efektif, yaitu dengan menerapkan konsep PHT (Pengendalian hama Terpadu) yang merupakan kombinasi dari beberapa teknik untuk mengendalikan populasi hama hingga di bawah ambang ekonomi. Salah satu komponen PHT adalah dengan memanfaatkan musuh alami.
Musuh alami yang dapat digunakan untuk menekan hama penggerek batang ini salah satunya adalah parasitoid. Pada umumnya parasitoid berkembang di dalam tubuh inang, sehingga ukuran tubuh parasitoid lebih kecil dan siklus hidupnya lebih pendek. Jenis parasitoid yang telah banyak dikembangkan adalah parasitoid telur, contohnya Trichogramma chilonis. Parasitoid telur Trichogramma telah dilepaskan secara berkala di perkebunan tebu PG Tersana Baru, PTP XIV, Jawa Barat sejak tahun 1971. Beberapa tahun sebelumnya
pelepasan Trichogramma juga telah dilakukan oleh PG Sumberharjo (Samoedi et al. 1986). Pada tahun 1976, Trichogramma diintroduksi ke Lampung, kemudian dikembangkan secara massal dan dilepas secara berkala di areal perkebunan tebu Terusan Nunyai, Lampung Tengah (Sunaryo 1988 dalam Susilo et al. 2007).
Hasil evaluasi yang dilakukan pada tahun 1977 menunjukkan bahwa Trichogramma mampu hidup dan berkembang biak dengan baik di pertanaman, meskipun pelepasan sebanyak 50.000 ekor Trichogramma per hektar per musim tanam belum cukup mampu menaikkan daya parasitisasinya (Samoedi et al. 1986). Menurut Boedijono (1990 dalam Untung 1993), untuk dapat mengendalikan penggerek batang tebu diperlukan pelepasan Trichogramma sebanyak 250.000 telur per hektar.
Smith (1996) menyebutkan bahwa penelitian mendalam mengenai pengembangan dan pelepasan parasitoid dari genus Trichogramma telah banyak dilakukan di beberapa negara dan dilaporkan berhasil dalam menekan populasi hama terutama dari ordo Lepidoptera. Penelitian Nurindah dan Bindra (1989) membuktikan bahwa hasil uji laboratorium pada 5 spesies, yaitu Trichogramma australicum Girault, T. chilonis Ishii, T. japonicum Ashmead, T. minutum Riley dan Trichogrammatoidea nana (Zehntner) yang digunakan untuk mengendalikan hama penggerek, mampu memarasit dengan persentase parasitisasi berkisar antara 23%-37% (Nurafiatin 2000). Menurut Li-Ying Li (1994), T. chilonis merupakan salah satu agen antagonis yang paling sukses dalam mengendalikan populasi penggerek batang tebu di Filippina, India dan Taiwan.
Potensi yang dimiliki T. chilonis mendapat banyak perhatian sehingga saat ini T. chilonis telah banyak dikembangkan sebagai agen pengendali hayati. T. chilonis dapat dikembangkan pada inang aslinya maupun dengan memanfaatkan inang alternatif. Di beberapa negara, pengembang-biakan Trichogramma/ Trichogrammatoidea secara massal di laboratorium dilakukan dengan memanfaatkan inang alternatif atau inang pengganti, karena sangat potensial dalam menurunkan biaya mekanisasi (Untung 1993). Inang alternatif yang sering digunakan dalam pengembangan T. chilonis dalam skala laboratorium adalah Corcyra cephalonica, Sitotroga cerealella dan Ephestia kuehniella (Smith 1996).
Sebelum melaksanakan pengembangan T. chilonis, perlu diketahui terlebih dahulu bioekologi parasitoid tersebut agar dalam pemanfaatannya dapat lebih efektif dan efisien, dalam hal ini termasuk ada tidaknya pengaruh perbedaan jenis inang yang digunakan terhadap kebugaran parasitoid tersebut terutama yang berhubungan dengan tingkat kemampuan parasitisasi. Informasi mengenai kisaran inang dan preferensi juga diperlukan untuk mendukung keberhasilan penggunaan T. chilonis sebagai agens pengendalian hayati. Menurut Sosromarsono (2000 dalam Nurafiatin 2000), keberhasilan pelepasan parasitoid agar dapat menetap dan menekan populasi inang sampai tingkat yang diinginkan sangat sulit diramalkan, tetapi pemahaman terhadap bioekologi dan perilaku parasitoid dapat membantu teknik dan pola pelepasan yang lebih tepat, sehingga peluang keberhasilan akan meningkat.
Tujuan
Penelitian ini bertujuan mempelajari kebugaran T. chilonis yang terdiri dari produksi telur, lama hidup (longevity), nisbah kelamin, lama perkembangan hidup, tingkat parasitisasi dan laju pemunculan imago. Selain itu, penelitian ini juga bertujuan mengetahui kisaran inang dan pengaruh jenis inang terhadap preferensi T. chilonis.
Hipotesis
Jenis inang berpengaruh terhadap produksi telur, lama hidup (longevity), nisbah kelamin, lama perkembangan hidup, persentase parasitisasi dan laju pemunculan imago parasitoid T. chilonis. Selain mempengaruhi ciri-ciri kebugaran, jenis inang juga berpengaruh terhadap pereferensi T. chilonis.
Manfaat
Hasil penelitian ini diharapkan mampu menambah informasi mengenai kebugaran, kisaran inang dan preferensi T. chilonis sehingga dapat dijadikan dasar dalam pengembangannya sebagai parasitoid telur pada berbagai hama penggerek.
TINJAUAN PUSTAKA
Pemanfaatan Parasitoid dari Genus Trichogramma
Lebih dari 100 tahun pemanfaatan Trichogramma sebagai agens pengendali hayati untuk hama-hama Lepidoptera telah diketahui (Smith 1996). Borror et al. (1954) menyebutkan bahwa genus Trichogramma merupakan kelompok yang paling terkenal dan secara meluas telah digunakan sebagai agen pengendali hayati (musuh alami). Saat ini Trichogramma telah dimanfaatkan oleh lebih dari 23 negara. Sebanyak 18 spesies Trichogramma yang berbeda dimanfaatkan sebagai agen pengendali hayati pada tanaman jagung, tebu, padi, kacang-kacangan, kapas, sayuran dan pinus (Hassan 1993).
Keefektifan Trichogramma dalam menekan populasi hama dapat diketahui dengan melakukan monitoring dan evaluasi. Monitoring dan evaluasi dilakukan dengan mengumpulkan telur hama penggerek dari beberapa tanaman sampel, kemudian dihitung persentase parasitisasinya. Dari hasil diskusi langsung, diketahui bahwa hasil monitoring dan evaluasi yang dilakukan oleh tim monitoring PT. Sweet Indolampung menunjukkan bahwa T. chilonis mampu memarasit hingga 70% dari jumlah total telur C. auricilius.
Telah dilaporkan bahwa pelepasan 150.000 telur Trichogramma sp. per hektar dapat menurunkan populasi hama penggerek pucuk tebu, sedangkan untuk pengendalian penggerek batang tebu diperlukan 250.000 telur per hektar (Boedijono 1990 dalam Untung 1993). Di beberapa negara seperti China, Filippina, India dan Taiwan, T. chilonis dimanfaatkan sebagai agens pengendali hayati pada pertanaman tebu (Hassan 1993). Di Indonesia, T. chilonis pertama kali dikembangkan untuk mengatasi permasalahan hama penggerek batang di beberapa perkebunan tebu di Pulau Jawa, kemudian diintroduksi ke Lampung setelah muncul permasalahan yang sama. Selain mampu memarasit Chilo spp., T. chilonis juga mampu memarasit Agrotis spp., Argyroploce schistaceana, Cnaphalocrosis medinalis, Diatraea spp., Heliothis armigera, Leguminivora glycinivorella dan Spodoptera exigua (Li 1994).
Trichogramma chilonis Ishii
T. chilonis merupakan salah satu parasitoid yang termasuk dalam ordo Hymenoptera. Parasitoid ini dimanfaatkan untuk mengendalikan berbagai hama penggerek terutama dari ordo Lepidoptera. Beberapa negara yang telah melakukan pembiakan massal parasitoid ini diantaranya India, Taiwan, Jepang, Pakistan, Iran, China, Switzerland, Canada, Filippina, Hawaii dan beberapa negara lainnya (Oatman et al. 1982; Hassan 1993; Miura & Kobayashi 1998; Ahmed et al. 2003; Shirazi 2006; Shahid et al. 2007). Di Indonesia, pemanfaatan T. chilonis sebagai agens hayati pada hama penggerek batang tebu telah dilakukan sejak tahun 1970-an, yaitu di beberapa perusahaan gula di Jawa seperti PG Tersana Baru, PG Sumberharjo dan beberapa perusahaan gula lainnya (Samoedi et al. 1986). Pada tahun 1976, Trichogramma diintroduksi ke Lampung, kemudian dikembangkan secara massal dan dilepas secara berkala di areal perkebunan tebu Terusan Nunyai, Lampung Tengah (Sunaryo 1988 dalam Susilo et al. 2007)
Seperti jenis Trichogramma lainnya, T. chilonis memiliki ciri khas rambut-rambut halus pada bagian sayapnya dan tarsi yang beruas tiga (Borror et al. 1996). Perkembangan T.chilonis dari telur hingga mencapai imago dewasa berkisar antara 7-9 hari (Agus 1991). Nisbah kelamin pada populasi serangga ini biasanya 1 : 2 (jantan : betina). Imago betina mampu menghasilkan 20-50 keturunan. Betina yang tidak dibuahi, secara partenogenesis menghasilkan keturunan jantan. Sedangkan betina yang dibuahi menghasilkan keturunan betina fertil. Serangga ini meletakkan telur pada telur inang, sehingga dapat berkembang secara terus-menerus selama telur inang masih tersedia. T. chilonis menyebar secara pasif dengan bantuan angin (Kalshoven 1981). Imago dapat menemukan telur inang dengan bantuan kairomon, senyawa kimia yang dihasilkan oleh inang (Hassan 1993).
Menurut Suzuki et al. (1984 dalam Schmidt 1994), ada empat tahapan perilaku betina T. chilonis dalam meletakkan telur: (1) melakukan postur oviposisi dengan kepala terangkat, bagian abdomen ditekan, antena terangkat dan ovipositor turun; (2) berputar-putar; (3) penetrasi ovipositor selama 30 detik dan (4) oviposisi. Menurut Hassan (1993), ketika telur Trichogramma/
Trichogrammatoidea di dalam telur inang telah berubah menjadi larva, larva akan memakan isi telur inang yang dapat menyebabkan kematian telur inang. Stadia larva yang terdiri dari tiga instar dan stadia pupa berada di dalam telur inang. Telur inang yang terparasit akan berwarna hitam.
Corcyra cephalonica Stainton
C. Cephalonica merupakan salah satu hama gudang yang termasuk dalam famili Pyralidae (Lepidoptera). Ngengat yang sering disebut dengan nama ”rice moth” ini merupakan serangga kosmopolitan dan sering menimbulkan kerusakan di gudang penyimpanan beras yang kurang diperhatikan kebersihannya. Serangga ini dapat menyerang barbagai jenis komoditas pertanian diantaranya beras gilingan, tepung terigu, serbuk jagung, semolina, tepung gandum, kacang-kacangan, kopra, minyak biji palm, bungkil, biskuit, coklat dan sebagainya (Kalshoven 1981).
Telur serangga ini berwarna putih kekuningan, berbentuk oval dan berukuran 0,49 x 0,32 mm. Pada salah satu ujung telur terdapat tonjolan seperti puting yang berukuran kecil. Biasanya telur diletakkan satu per satu di antara butiran beras. Ketika akan menetas, telur tersebut berubah warna menjadi kecoklatan yang merupakan warna bakal kepala larva. Telur akan menetas menjadi larva setelah 5-7 hari dengan membuat lubang pada cangkang telur (Widayanti 1990).
Larva berwarna putih kekuningan dengan panjang tubuh mencapai lebih dari 17 mm. Larva tersebut mengeluarkan fras yang merupakan campuran antara sisa makanan dan kotoran, fras berwarna putih jika larva mengonsumsi beras. Adanya fras dapat mengurangi kualitas produk. Larva berbentuk silindris dan mempunyai seta di setiap ruasnya. Seta tersebut terdapat pada bercak-bercak coklat di abdomen dan di atas spirakel. Stadia larva terdiri dari 8 instar (Widayanti 1990).
Pupa berwarna coklat dan dilindungi oleh kokon berwarna putih. Kokon tersebut terbuat dari benang sutera yang dijalin kuat. Lamanya masa pupa ± 12 hari. Imago betina dapat menghasilkan lebih dari 400 telur dan mampu hidup hingga 10 hari. Imago serangga ini berwarna abu-abu dan memiliki panjang tubuh
8-12 mm. Antena ngengat ini bertipe filiform. Perbedaan antara imago betina dan imago jantan dilihat dari warna sayap belakang imago jantan yang lebih gelap, selain itu imago betina mempunyai palpus labia yang lebih panjang daripada imago betina. Perkembangan dari telur hingga menjadi imago dewasa membutuhkan waktu 40-60 hari (Kalshoven 1981; Widayanti 1990).
Helicoverpa (Heliothis) armigera Hubner
H. armigera (Lepidoptera: Noctuidae) sering disebut dengan istilah ”corn ear worm” dan merupakan serangga polifag yang menyerang lebih dari 160 jenis tanaman inang termasuk jagung, tomat, kentang, sorgum, jarak. kapas, tanaman hias, sayuran dan tembakau. Di Indonesia, hama ini mampu menyerang tanaman dari areal pertanaman pada dataran rendah hingga ketinggian lebih dari 2000 m (Kalshoven 1981).
Telur H. armigera berwarna kuning kehijauan, bentuknya menyerupai bola dengan diameter 0,7-0,8 mm. Warna telur akan berubah menjadi lebih gelap ketika akan menetas. Lama stadia telur 3-4 hari. Larva terdiri dari enam instar. Larva instar awal berwarna kekuningan dan ditumbuhi bulu kecoklatan, sedangkan larva instar akhir memiliki warna yang beragam sesuai dengan jenis makanannya. Pada tubuh bagian lateral larva instar akhir terdapat garis gelap terang yang memanjang. Lama stadia larva berkisar 15-23 hari (Rizal 1988). Larva Helicoverpa bersifat kanibal, sehingga jarang ditemukan lebih dari dua larva dalam satu tongkol jagung (Kalshoven 1981).
Pupa terbentuk di dalam tanah dengan warna hijau keputihan. Semakin lama pupa akan berubah warna menjadi coklat tua dan menjelang penetasan akan berwarna coklat kehitaman, licin dan mengkilat. Lama stadia pupa ± 13 hari. Imago muncul pada malam hari dan biasanya imago betina muncul lebih dulu dibandingkan imago jantan. Hal tersebut berkaitan dengan waktu yang dibutuhkan untuk pematangan ovum. Imago betina berwarna coklat kekuningan, sedangkan yang jantan berwarna coklat kelabu. Masa oviposisi imago betina dapat mencapai 17 hari. Satu individu betina rata-rata mampu menghasilkan 800-900 telur. Satu siklus hidup serangga ini membutuhkan waktu 28-40 hari (Rizal 1988).
Crocidolomia binotalis Zell.
C. binotalis atau yang dikenal dengan sebutan ulat krop kubis termasuk dalam ordo Lepidoptera, famili Pyralidae. Selain menyerang kubis, hama ini juga menyerang tanaman dari famili Brassicae lainnya serta tanaman dari famili Cruciferae (Kalshoven 1981). Imago serangga ini meletakkan telur pada permukaan atas dan bawah daun serta tangkai daun, meskipun telur lebih sering diletakkan pada permukaan bawah daun. Hal ini terjadi karena permukaan bawah daun yang berlekuk-lekuk dan tidak rata sehingga C. binotalis merasa telurnya lebih aman. Telur-telur tersebut diletakkan secara berkelompok dan saling tumpang tindih sehingga menyerupai atap rumah. Telur yang baru diletakkan berwarna hijau dan akan berubah menjadi coklat tua dan hitam ketika akan menetas. Lama stadium telur berkisar 3-6 hari (Othman 1982 dalam Dumalang 1996).
Larva C. binotalis terdiri dari empat instar dengan lama stadium larva 14-18 hari. Larva instar pertama berwarna kuning muda agak pucat dengan kepala berwarna coklat kehitaman. Larva instar kedua sampai keempat berwarna hijau dengan bintik hitam pada tiap ruas tubuhnya. Ketika memasuki tahap prapupa, larva akan membuat kokon dari bagian daun atau dari kotorannya sendiri. Pupa yang baru terbentuk berwarna hijau, kemudian berubah menjadi coklat dan mengkilat. Lama masa pupa berkisar 10-13 hari (Dumalang 1996).
Imago berwarna coklat kotor seperti jenis ngengat lainnya. Ciri khas dari imago C. binotalis adalah pada tepi sayap depan di bagian anterior dekat pangkal terdapat seberkas rambut keras. Lama hidup imago betina berkisar 15-20 hari (Dumalang 1996). Di Bogor, lama perkembangan hidup dari telur hingga menjadi imago dewasa membutuhkan waktu 22-30 hari (Kalshoven 1981).
Spodoptera litura Fabricius
S. litura merupakan serangga yang bersifat polifag, sehingga tidak hanya menyerang tanaman budidaya, serangga ini juga menyerang tanaman liar. Nama umum hama ini adalah armyworm atau di Indonesia sering disebut dengan ulat grayak karena tingkah lakunya yang hidup secara bergerombol dan aktivitas
makan yang tinggi sehingga menyebabkan tingkat serangan yang tinggi pula dalam waktu singkat.
Telur biasanya diletakkan pada permukaan atas dan bawah daun secara berkelompok dan tersusun rapi dalam dua atau tiga lapisan. telur yang baru diletakkan berwarna putih bening dan akan menjadi kelabu agak hitam ketika akan menetas. Larva yang baru menetas berwarna hijau muda. Setelah melakukan aktivitas makan warna larva menjadi hijau tua, kemudian menghitam pada larva instar akhir. Pada bagian dorsal tubuh larva terdapat tiga garis memanjang. Di setiap ruas terdapat bintik-bintik hitam berbentuk setengah lingkaran yang terletak di antara tiga garis memanjang tersebut. Larva terdiri dari enam instar dengan lama stadia 18-27 hari (Sumaryati 1993 dan Mardiningsih 1997).
Pada masa prapupa, tubuh larva akan mengecil dan memendek. Pupa terbentuk di dalam kokon yang terbuat dari tanah. Pupa yang baru terbentuk berwarna coklat kehijauan dan lunak, kemudian berubah menjadi coklat tua dan mengeras. Lama stadia pupa 9-10 hari. Tubuh imago yang baru muncul ditutupi oleh sisik-sisik halus berwarna coklat kelabu. Sayap depan berwarna coklat dengan garis-garis hitam dan putih. Sayap belakang berwarna putih kelabu. Masa praoviposisi imago betina 3-4 hari dan lama hidup imago berkisar antara 7-8 hari. Satu betina dapat menghasilkan lebih dari 1000 telur. Lama perkembangan dari telur menjadi imago dewasa membutuhkan waktu 33-36 hari (Sumaryati 1993).
Chilo auricilius Dudgeon
Serangga yang lebih dikenal dengan nama penggerek batang berkilat ini di Indonesia ditemukan pertama kali oleh Hazelhoff pada tahun 1928 di Pasuruan. Ciri khas yang membedakan spesies ini dengan spesies Chilo lainnya adalah sisik-sisik yang berkilat pada permukaan atas ujung sayap. Inang utama dari hama ini adalah tanaman tebu. Namun, hama ini juga ditemukan dapat menyerang gelagah dan jagung.
Berdasarkan pengamatan yang dilakukan oleh Ruinard (1958 dalam Wirioatmodjo 1977), imago meletakkan telur pada permukaan atas atau bawah daun. Telur tersebut berbentuk pipih dan eliptik, biasanya diletakkan dalam
kelompok yang terdiri dari dua barisan dan tersusun seperti atap genting. Jumlah telur dalam setiap kelompok berbeda-beda, berkisar antara 7 sampai 104 buah. Telur yang baru diletakkan berwarna putih susu dan berubah warna menjadi keunguan setelah dua hari. Larva di dalam telur akan tampak jelas pada hari ke-4. Penetasan telur biasanya terjadi pada pagi hari dan larva yang baru menetas memiliki panjang tubuh kurang lebih 2 mm. Bagian kepala dan perisai leher berukuran lebih besar daripada bagian badan. Protoraks dan kepala larva berwarna hitam coklat sampai hitam, sedangkan bagian badan berwarna putih kekuningan. Tidak banyak terjadi perubahan warna selama pertumbuhan, hanya bagian protoraks yang berubah menjadi kuning coklat. Larva instar akhir dapat mencapai panjang 11,5-21 mm. Lama stadia larva berkisar 21-41 hari.
Pupa awalnya berwarna coklat kuning, kemudian berubah menjadi coklat tua hingga hitam. Rata-rata panjang pupa 12,7 mm. Lama stadia pupa berkisar 5-7 hari. Imago memiliki panjang sayap 16-29 mm. Sayap depan berwarna kuning kecoklatan dengan sisik berkilat pada ujungnya dan sayap belakang berwarna kelabu kotor. Oviposisi dapat terjadi selama empat hari. Seekor imago betina dapat menghasilkan sebanyak 385-412 telur. Telur penggerek batang berkilat dapat diparasit oleh Telenomus beneficiens Zehnt dan Trichogramma sp (Ruinard 1958 dalam Wirioatmodjo 1977).
Pengaruh Jenis Inang yang Berbeda terhadap Kebugaran Parasitoid
Penelitian mengenai kebugaran Trichogramma yang dihasilkan dari inang yang berbeda telah banyak dilakukan. Salah satunya yaitu penelitian yang dilakukan oleh Corrigan & Laing pada tahun 1994. Corrigan & Laing mengemukakan bahwa kemampuan reproduksi Trichogramma dapat meningkat atau mengalami penurunan sesuai dengan jenis inang yang digunakan dalam pembiakan. Selain itu, jenis inang yang digunakan dalam pembiakan juga menentukan kualitas dan kemampuan parasitoid dalam menyerang inang target.
Hasil penelitian Quen pada tahun 1988 menunjukkan bahwa T. dendrolimi memiliki kemampuan menyerang berbagai jenis telur inang dengan ketebalan yang berbeda-beda, sedangkan T. ostriniae hanya mampu menyerang telur inang yang memiliki kulit tipis (Djuwarso & Wikardi 1997). Nurafiatin (2000)
menyimpulkan bahwa imago betina T. cojuangcoi yang dihasilkan dari inang Plutella xylostella memiliki ciri-ciri kebugaran yang lebih rendah dibandingkan imago betina yang muncul dari inang C. cephalonica dan H. armigera. Hal tersebut didasarkan pada ukuran imago yang kecil, keperidian rendah serta lama hidup dan masa reproduksi yang singkat. Namun, kebugaran parasitoid dari genus Trichogrammatoidea yang dihasilkan oleh inang P. xylostella masih lebih baik jika dibandingkan dengan inang S. litura dan C. binotalis (Yuanita 2002). Menurut Buchori et al (2000), perbedaan kebugaran parasitoid dari jenis inang yang berbeda disebabkan oleh ukuran telur inang yang berbeda-beda dan kelengkapan nutrisi yang terkandung di dalam telur tersebut.
Kisaran Inang dan Preferensi Parasitoid Telur
Trichogramma merupakan salah satu jenis parasitoid yang bersifat polifag, karena dapat hidup dan berkembang biak pada jenis inang lain selain inang utamanya. Li (1994) menyatakan bahwa T. chilonis menyerang berbagai jenis hama yang menyerang tanaman budidaya, diantaranya jenis-jenis penggerek yang menyerang jagung, tebu dan kapas. Pembiakan massal T. chilonis di laboratorium biasanya menggunakan inang C. cephalonica. Pengetahuan mengenai jenis-jenis inang yang dapat diserang oleh suatu spesies Trichogramma sangat dibutuhkan untuk mengetahui inang yang dapat digunakan sebagai alternatif dalam mempertahankan hidupnya saat inang utama tidak ditemukan di lapangan. Selain itu, juga dapat digunakan untuk memperkirakan inang yang paling efektif dan efisien dalam pembiakan massal.
Pembiakan massal di laboratorium menggunakan inang pengganti secara terus-menerus terkadang menyebabkan terjadinya pergeseran preferensi, meskipun tidak semua spesies memberikan respon yang sama (Smith 1996). Beberapa faktor yang menyebabkan perbedaan tingkat preferensi diantaranya kualitas inang dan pengaruh fisik telur seperti ketebalan kulit dan ada tidaknya bulu yang menutupi telur. Trichogrammatoidea cojuangcoi lebih memilih inang H. armigera yang memiliki kualitas inang lebih baik dibandingkan C. cephalonica dan P. xylostella (Nurafiatin 2000). Hasil penelitian Yuanita (2002) menunjukkan tingkat preferensi Trichogrammatoidea sp. pada inang P. xylostella, S. litura dan
C. binotalis cukup rendah. Hal tersebut dikarenakan ukuran telur inang yang kecil dan nutrisi inang yang kurang sesuai untuk perkembangan larva.
Tingkat preferensi juga dipengaruhi oleh umur inang. Hasil penelitian Djuwarso et al (1997) menunjukkan tingkat parasitisasi T. bactrae-bactrae pada telur Etiella sp. yang berumur satu hari lebih tinggi dibandingkan dengan telur yang berumur dua dan tiga hari. Godfray (1994) menyatakan bahwa kualitas inang didasarkan pada umur inang tersebut. Semakin muda umur inang, kualitasnya semakin baik sehingga lebih disukai oleh parasitoid.
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan mulai Februari 2010 hingga Juni 2010 di laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Bahan, Alat dan Metode Penelitian
Kegiatan yang dilakukan dalam penelitian ini meliputi perbanyakan inang dan parasitoid telur serta pengujian kebugaran, kisaran inang dan preferensi. Parasitoid telur yang diuji adalah Trichogramma chilonis yang berasal dari Lampung.
Perbanyakan Inang
Corcyra cephalonica. Serangga inang C. cephalonica dikoleksi dari gudang
penggilingan beras di daerah Ciawi. Serangga tersebut kemudian dipelihara dalam silinder peneluran yang terbuat dari bahan karton dengan diameter 8 cm dan tinggi 20 cm. Bagian atas dan bawah silinder ditutup dengan kawat kassa 25 mesh. Telur C. cephalonica yang dihasilkan dikumpulkan dengan menggunakan kuas dan kemudian disimpan dalam cawan petri. Telur yang telah terkumpul tersebut sebagian digunakan untuk perlakuan dan sisanya ditebar kembali ke dalam wadah pemeliharaan C. cephalonica di laboratorium. Wadah pemeliharaan yang digunakan terbuat dari wadah plastik berukuran 34 x 26 x 7 cm yang bagian atasnya telah dilubangi dan ditutupi dengan kawat kassa 25 mesh. Wadah pemeliharaan ini diisi dengan pakan ternak, yaitu campuran dedak dan pur ayam dengan perbandingan 1 : 2.
Chilo auricilius. Larva C. auricilius yang dikoleksi dari perkebunan tebu di
Lampung, dimasukkan dalam wadah pemeliharaan berupa tabung berukuran 3 x 15 cm yang telah berisi pakan buatan. Pakan buatan dibuat dengan memasak 80 g
kacang hijau rebus yang telah dihaluskan bersama 30 g bagas (ampas tebu) dan 7,5 g pengembang kue dalam 500 ml air steril. Selain itu juga ditambahkan bahan-bahan lain diantaranya 7 g agar; 1,3 g vitamin C; 5 tablet vitamin B Complex; 2 g sukrosa; 4 g glukosa dan 2,75 g nevagin. Seluruh bahan tersebut dicampur dengan kacang hijau dan bagas hingga mengental kemudian diberi 0,2 cc formalin 40% agar pakan buatan lebih tahan lama. Larva yang telah berubah menjadi pupa dipindahkan ke dalam toples berdiameter 25 cm. Imago yang muncul dipindahkan dalam toples lain berdiamater 15 cm yang telah ditutup dengan plastik hitam disekelilingnya, bagian dalam toples dikelilingi kertas berlipat sebagai tempat bertelur. Telur yang digunakan untuk perlakuan adalah telur yang berumur kurang ≤ 1 hari. Imago dalam toples diberi makan madu 20%.
Crocidolomia binotalis dan Spodoptera litura. Metode yang digunakan
dalam pemeliharaan kedua jenis serangga ini relatif sama. Larva C. binotalis dan S. litura yang diambil dari lapang dipelihara dalam kotak plastik berukuran 34 x 26 x 7 cm. Larva C. binotalis diberi pakan daun caisin dan/atau daun brokoli, sedangkan larva S. litura diberi pakan daun talas. Larva yang telah mencapai instar akhir diberi serbuk gergaji setebal ± 1,5 cm sebagai habitat yang cocok untuk pembentukan pupa. Imago C. binotalis dipelihara dalam toples berdiameter 15 cm yang didalamnya telah diletakkan daun brokoli sebagai tempat peletakan telur. Sedangkan imago S. litura dipelihara dalam toples berukuran sama yang sisi dalamnya dilapisi kertas buram sebagai tempat peletakan telur. Telur yang digunakan untuk perlakuan adalah telur yang berumur ± 1 hari. Imago dalam toples diberi makan madu 20%.
Helicoverpa armigera. Larva H. armigera dikoleksi dari daerah Parung,
Bogor. Larva tersebut dipelihara di laboratorium dalam wadah plastik berdiameter 3 cm. Satu wadah plastik berisi satu larva, hal ini untuk menghindari kanibalisme antar larva. Larva diberi pakan babycorn atau jagung semi. Larva yang telah mencapai instar akhir diberi serbuk gergaji untuk membantu dalam proses pembentukan pupa. Pupa yang terbentuk dicuci dengan kloroks 2%, kemudian dibilas dua kali menggunakan air biasa. Pencucian ini bertujuan untuk menghilangkan patogen yang menempel pada pupa serta memisahkan antara pupa
yang sehat dengan pupa yang kurang sehat. Imago yang muncul dipindahkan ke dalam toples berdiameter 25 cm yang bagian sisi dalamnya telah dilapisi kertas tissu bertekstur kasar sebagai tempat peletakan telur. Imago diberi makan madu 20%. Telur yang dipakai untuk perlakuan adalah telur yang berumur ± 1 hari. Pembuatan Pias
Telur C. cephalonica yang berumur ± 1 hari dimasukkan dalam freezer selama 2 jam agar telur yang tidak terparasit tidak menetas menjadi larva, karena larva serangga ini bersifat kanibal sehingga ditakutkan memangsa telur-telur lainnya termasuk telur yang terparasit. Telur tersebut kemudian ditempelkan pada karton manila berukuran 0,5 x 3,5 cm dengan menggunakan gum arabic. Jumlah telur tidak diperhatikan untuk pias perbanyakan parasitoid, sedangkan untuk pias perlakuan jumlah telur yang ditempelkan sebanyak 20 butir. Pias H.armigera dibuat dengan memotong kertas tissu yang ditempeli telur, satu potongan berisi 20 telur. Telur tersebut dimasukkan dalam freezer selama 4 jam sebelum digunakan sebagai inang dalam perlakuan. Pembuatan pias C. binotalis dilakukan dengan memotong daun brokoli yang ditempeli telur, karena telur bergerombol maka setiap potongan berisi satu gerombolan telur. Potongan daun tersebut kemudian direkatkan pada pias kertas. Sedangkan pias C. auricilius dan S. litura dibuat dengan memotong kertas yang ditempeli telur. Setiap potongan kertas berisi 20 telur yang kemudian direkatkan pada pias kertas.
Perbanyakan Parasitoid Telur
T. chilonis yang diambil dari Lampung dikembangkan dalam tabung berukuran 1,5 x 10 cm. Tabung yang digunakan diolesi madu 20% sebagai pakan. Pias untuk perbanyakan yang telah ditempeli telur C. cephalonica dimasukkan ke dalam tabung tersebut untuk diparasit. Telur yang sudah terparasit ditandai dengan perubahan warna dari putih kekuningan menjadi kelabu dan menghitam. Pias yang sudah terparasit dipindahkan ke tabung baru dan disimpan pada suhu ruang hingga imago yang baru muncul.
LP =
N PP =
M
Pengujian ini menggunakan dua jenis inang yaitu C. auricilius dan C. cephalonica. Uji kebugaran ini hanya dilakukan pada 20 butir telur dikarenakan jumlah telur inang yang terbatas. Pias dimasukkan dalam tabung yang berisi sepasang imago T. chilonis untuk diparasit. Sebelumnya, tabung tersebut diolesi madu 20% sebagai pakan parasitoid. Pengamatan yang dilakukan meliputi produksi telur, lama hidup (longevity), nisbah kelamin, persentase parasitisasi, lama perkembangan hidup, laju pemunculan imago dan kematian pradewasa. Percobaan dilakukan sebanyak 20 ulangan. Parameter pengamatan di atas dihitung dengan cara sebagai berikut:
Produksi telur. Produksi telur diketahui dengan menghitung jumlah telur yang diletakkan oleh imago betina pada telur inang dalam satu hari.
Lama hidup. Lama hidup diamati mulai dari hari pertama imago betina diberikan perlakuan sampai imago tersebut mati.
Nisbah kelamin. Jumlah imago jantan dan betina dihitung setiap hari dari total seluruh imago yang muncul. Penghitungan dilakukan di bawah mikroskop compound.
Persentase parasitisasi. Persentase parasitisasi diketahui dengan menghitung banyaknya telur inang yang terparasit setiap hari. Telur inang terparasit diketahui dengan adanya perubahan warna telur inang, dari putih kekuningan menjadi hitam.
PP = persentase parasitisasi m = jumlah telur inang terparasit M = jumlah total telur inang
Lama perkembangan hidup. Lama perkembangan dihitung mulai dari hari pertama telur parasitoid dimasukkan dalam telur inang hingga waktu pemunculan imago tiba.
∑ (Ni x ni) m
LP = Lama perkembangan
Ni = Jumlah imago yang muncul pada hari ke-i ni = Hari ke-i
N = Total imago yang muncul
Persentase laju pemunculan imago (PLPI). Besarnya nilai PLPI diketahui dengan membandingkan jumlah imago yang muncul pada hari ke-i dengan total imago yang muncul.
Kematian pradewasa. Kematian pradewasa diketahui dengan membedah telur inang pada hari terakhir pemunculan imago dan menghitung jumlah pupa mati, yaitu pupa yang tidak berhasil menjadi imago maupun imago yang tidak berkembang sempurna sehingga tidak dapat muncul dari telur inang.
Pengujian Kisaran Inang
Pengujian ini dilakukan pada lima jenis inang yaitu C. auricilius, C. cephalonica, C. binotalis, H. armigera dan S. litura. Parameter yang digunakan diantaranya produksi telur, persentase parasitisasi dan persentase laju survival. Penghitungan nilai produksi telur dan persentase parasitisasi sama seperti yang telah dijelaskan sebelumnya. Persentase laju survival merupakan kemampuan parasitoid untuk berkembang dari telur diletakkan hingga berhasil keluar menjadi imago, atau total imago yang muncul.
% Laju survival
Pengujian Preferensi T. chilonis
PLPI = Ni x 100% N
total imago yang muncul total telur yang diletakkan
Pengujian ini dilakukan menggunakan inang yang sama dengan pengujian kisaran inang. Pengujian dilakukan dua kali dengan lama pemaparan yang berbeda, 30 menit dan 24 jam. Pengujian dengan lama pemaparan 30 menit dilakukan untuk menghindari terjadinya bias data karena waktu pemaparan yang lama. Parameter yang digunakan dalam pengujian ini hanya tingkat parasitisasi dan produksi telur.
Analisis Statistik
Rancangan yang digunakan adalah Rancangan Acak Lengkap (RAL). Data diolah dengan menggunakan analisis sidik ragam dan dilanjutkan dengan uji Tukey pada taraf kepercayaan 95%.
HASIL DAN PEMBAHASAN
Kebugaran T. chilonis pada Dua Jenis Inang
Pada kedua jenis inang, telur yang terparasit dapat diketahui pada 3-4 hari setelah parasitisasi. Telur yang terparasit ditandai dengan perubahan warna menjadi kehitaman. Telur C. auricilius yang mula-mula berwarna putih susu akan berubah menjadi berwarna hitam pada seluruh permukaannya. Perubahan warna tersebut diawali dengan adanya bintik hitam yang merupakan bekas tusukan setelah terjadi oviposisi. Telur C. auricilius yang sehat dan tidak terparasit akan menghitam pada hari ke-4 setelah telur diletakkan, namun warna hitam hanya pada sebagian permukaan yang merupakan bakal kepala larva. Pada telur yang tidak sehat, tidak terjadi perubahan warna. Telur C. cephalonica yang terparasit ditandai dengan perubahan warna yang dimulai dengan adanya bintik kehitaman dan akhirnya seluruh permukaan telur menjadi berwarna hitam.
Perubahan warna pada kedua telur inang terjadi karena perkembangan parasitoid dari telur hingga menjadi pupa. Proses peletakan telur yang dilakukan oleh imago betina terjadi dalam empat tahapan: (1) melakukan postur oviposisi dengan kepala terangkat, bagian abdomen ditekan, antena terangkat dan ovipositor turun; (2) berputar-putar; (3) penetrasi ovipositor selama 30 detik dan (4) oviposisi (Suzuki et al. 1984 dalam Schmidt 1994). Hassan (1993) mengemukakan bahwa dalam menemukan inang yang dipilihnya, imago T. chilonis betina akan selalu berada pada atau dekat dengan telur inang selama beberapa lama hingga semua atau beberapa dari inang tersebut terparasit.
Tingkat Parasitisasi
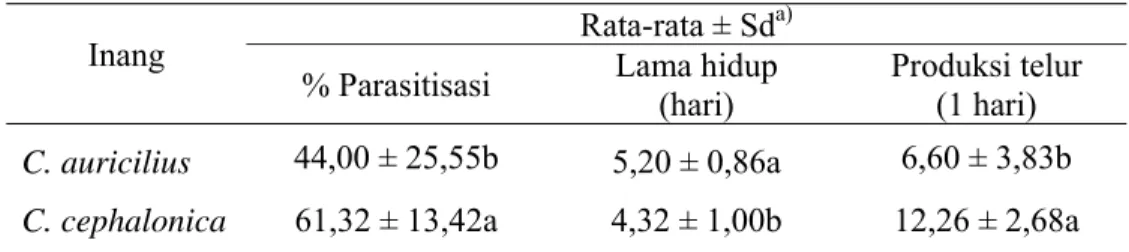
Dari hasil analisis sidik ragam, diketahui jenis inang memiliki pengaruh terhadap persentase parasitisasi. T. chilonis hanya mampu memarasit 44,00% dari jumlah total telur C. auricilius yang tersedia. Pada inang C. cephalonica, persentase telur yang terparasit lebih besar yaitu 61,32% (Tabel 1).
Tabel 1 Tingkat parasitisasi, lama hidup dan produksi telur T. chilonis pada dua jenis inang
Inang Rata-rata ± Sd
a)
% Parasitisasi Lama hidup (hari) Produksi telur (1 hari)
C. auricilius 44,00 ± 25,55b 5,20 ± 0,86a 6,60 ± 3,83b
C. cephalonica 61,32 ± 13,42a 4,32 ± 1,00b 12,26 ± 2,68a a Angka selajur yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan uji Tukey pada
taraf α 0,05.
Kemampuan parasitisasi dipengaruhi oleh kualitas inang. Sebelum memarasit atau melakukan oviposisi, imago betina akan melakukan orientasi untuk memilih telur inang yang berkualitas baik dengan cara menyentuhkan antena dan palpus pada telur inang (Yunus et al. 2004). Imago betina hanya akan meletakkan telur pada telur inang yang dianggap layak untuk perkembangan keturunannya. Kualitas telur inang yang kurang baik menyebabkan imago betina enggan meletakkan telur di dalamnya sehingga persentase parasitisasi rendah (Godfray 1994). Telur C. auricilius yang kurang baik kualitasnya diduga disebabkan oleh kondisi di laboratorium yang kurang sesuai untuk perkembangan C. auricilius sehingga produksi telur oleh imago tidak optimal. Rendahnya persentase parasitisasi pada C. auricilius diduga juga disebabkan oleh susunan telurnya yang saling tumpang tindih, sehingga menyulitkan imago untuk menemukan inang yang cocok sebagai tempat oviposisi.
C. auricilius merupakan inang asli T. chilonis di lapangan. Dari hasil evaluasi tingkat parasitisasi T. chilonis terhadap C. auricilius di lapangan yang dilakukan oleh PT. Sweet Indolampung, Lampung, persentase parasitisasi yang dihasilkan cukup tinggi, yaitu mencapai 70% (lihat bab tinjauan pustaka). Perusahaan yang bergerak dalam bidang industri gula tersebut menyimpulkan bahwa T. chilonis sangat efektif dalam menekan populasi C. auricilius, sehingga sampai saat ini pelepasan T. chilonis merupakan solusi terbaik dalam memecahkan permasalahan hama penggerek batang tebu.
Hasil penelitian yang bertolak belakang dengan hasil evaluasi yang dilakukan di lapangan tersebut dapat dipengaruhi oleh kondisi lingkungan yang berbeda. Diduga kondisi di sekitar pertanaman termasuk suhu dan iklim di
sekitarnya merupakan kondisi optimal untuk perkembangan T. chilonis, sehingga T. chilonis mampu memarasit dalam jumlah yang lebih banyak. Selain iklim, keberhasilan pengendalian dengan memanfaatkan T. chilonis juga dipengaruhi oleh faktor agronomi (Soula et al. 2003). Rana et al. (2007) menyebutkan bahwa penggunaan mulsa sampah organik yang dipadukan dengan pemanfaatan T. chilonis dapat meningkatkan kemampuan parasitisasi parasitoid tersebut di lapangan.
Tingginya tingkat parasitisasi dari hasil evaluasi menunjukkan keefektifan penggunaan T. chilonis sebagai agens pengendalian hayati di lapangan. Namun diperlukan peninjauan kembali terhadap hasil ini, contohnya dengan menggunakan teknik pengambilan sampel yang lebih baik sehingga didapatkan hasil yang lebih akurat. Hal ini perlu dilakukan mengingat tingkat parasitisasi hasil evaluasi sangat tinggi, sedangkan hasil uji laboratorium menunjukkan bahwa tingkat parasitisasi T. chilonis pada C. auricilius hanya sebesar 44,00%.
Lama Hidup
Lama hidup T. chilonis dapat dipengaruhi oleh jenis inang. Imago pada inang C. auricilius mampu bertahan hidup lebih lama jika dibandingkan pada C. cephalonica. Pada C. cephalonica imago bertahan hidup hingga ± 4 hari, sedangkan pada C. auricilius imago mampu bertahan hidup selama ± 5 hari. Lama hidup imago betina berkaitan erat dengan produksi telur. Dari tabel 1, diketahui bahwa semakin banyak imago betina meletakkan telur, maka kemampuan bertahan hidupnya semakin singkat. Hal ini diduga disebabkan oleh pemanfaatan energi selama hidupnya hanya difokuskan untuk peletakan telur, sehingga energi untuk bertahan hidup semakin menurun.
Selain dipengaruhi oleh jenis inang, lama hidup parasitoid juga dapat dipengaruhi oleh konsentrasi pakan, dalam hal ini madu. Madu dengan konsentrasi tinggi mampu meningkatkan peluang bertahan hidup imago betina (Handayani 2004). Saljoqi dan Khattak (2007) menyebutkan bahwa pemberian pakan berupa larutan madu 50% dapat meningkatkan keefektifan T. chilonis sebagai agens pengendali hayati. Pada pengendalian hayati di lapangan,
ketersediaan pakan berupa madu perlu diperhatikan agar imago parasitoid mampu bertahan hidup lebih lama, sehingga diharapkan terjadi peningkatan jumlah telur hama yang terparasit.
Produksi telur
Hasil penelitian memperlihatkan bahwa jenis inang dapat mempengaruhi produksi telur T. chilonis. Pada inang C. cephalonica, T. chilonis mampu meletakkan telur dalam jumlah yang lebih banyak dibandingkan pada inang C. auricilius (Tabel 1). Selisih produksi telur T. chilonis antara kedua inang sangat besar. Dari tabel 1 diketahui bahwa nilai produksi telur pada inang C. auricilius hanya sebesar 6,60. Diduga faktor yang menyebabkan rendahnya produksi telur T. chilonis adalah kualitas telur inang yang kurang baik dan susunan telur yang saling tumpang tindih. Kedua faktor tersebut menyebabkan jumlah telur yang terpilih sebagai tempat yang sesuai untuk oviposisi menjadi lebih sedikit sehingga jumlah telur yang dihasilkan oleh imago betina tidak maksimal. Berdasarkan hasil penelitian tersebut, rendahnya produksi telur pada C. auricilius dapat digunakan sebagai dasar dalam penentuan jumlah T. chilonis yang harus dilepaskan pada pengendalian hayati di lapangan, dalam hal ini diperlukan T. chilonis dalam jumlah yang sangat banyak agar efektif dalam mengendalikan populasi hama penggerek batang.
Produksi telur dipengaruhi oleh ukuran imago betina, ukuran telur inang, kualitas telur inang dan kemampuan oviposisi dari imago betina (Nurafiatin 2000). Menurut Yunus et al. (2004), produksi telur cenderung meningkat seiring dengan besarnya ukuran imago betina, meskipun peningkatan tersebut tidak signifikan. Nilai produksi telur yang tinggi menggambarkan potensi parasitoid dalam menghasilkan keturunan yang cukup banyak. Hal tersebut mendukung keberhasilan pembiakan massal di laboratorium.
Lama Perkembangan
Hasil analisis sidik ragam menunjukkan bahwa jenis inang tidak mempengaruhi lama perkembangan T. chilonis. Pada kedua jenis inang, waktu
yang dibutuhkan dari mulai telur diletakkan hingga pemunculan imago adalah ± 8 hari. Beberapa faktor yang dapat mempengaruhi lama perkembangan suatu spesies parasitoid diantaranya nutrisi inang, kondisi lingkungan serta suhu selama perkembangan. Suhu yang rendah menyebabkan waktu yang dibutuhkan parasitoid untuk berkembang menjadi lebih lama. Sebaliknya, semakin tinggi suhu waktu pemunculan imago semakin cepat. Lama perkembangan terpendek Trichogramma yaitu pada suhu 30°C dan terpanjang pada suhu 18°C (Sari & Buchori 2004). Suhu yang tinggi juga dapat mempersingkat waktu yang dibutuhkan untuk pre-oviposisi (Reznik et al. 2009). Nadeem et al. (2009) menyebutkan bahwa suhu yang paling optimal untuk perkembangan T. chilonis adalah 28°C.
Informasi mengenai lama perkembangan parasitoid telur pada suatu inang sangat dibutuhkan. Pada pembiakan massal di laboratorium, informasi ini digunakan sebagai referensi untuk menentukan inang yang paling efisien, dalam hal ini jenis inang yang menunjukkan lama perkembangan parasitoid lebih singkat. Informasi lama perkembangan parasitoid juga dapat dijadikan dasar dalam menentukan waktu pelepasan yang tepat ke lapangan. Pelepasan parasitoid ke lapangan biasanya dilakukan satu hari sebelum imago muncul, sehingga saat waktu pemunculan tiba imago dapat langsung menyebar dan melakukan pencarian inang.
Kematian Pradewasa
Di dalam telur inang, parasitoid mengalami perkembangan dari telur hingga menjadi imago yang siap muncul dan beraktivitas. Pupa yang tidak mampu berkembang menjadi imago maupun imago yang tidak dapat melubangi kulit telur inang sehingga tidak keluar disebut dengan pupa mati. Persentase pupa mati menggambarkan kemampuan dari telur inang dalam menunjang keberhasilan hidup T. chilonis. Komposisi nutrisi yang terkandung dalam telur inang berbeda-beda sehingga kemampuan dalam menunjang keberhasilan hidup parasitoid juga berbeda. Selain itu, ketebalan kulit telur inang juga mempengaruhi keberhasilan imago untuk melubangi telur inang. Secara tidak langsung hal ini menjadi faktor penyebab kematian pradewasa T. chilonis.
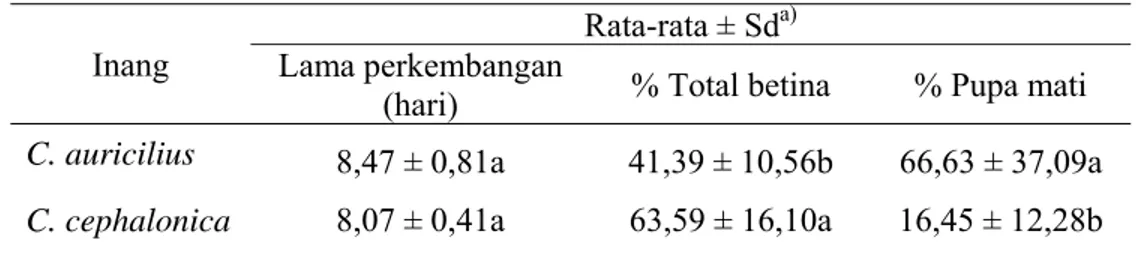
Tabel 2 Lama perkembangan, persentase total betina dan persentase pupa mati T. chilonis pada dua jenis inang
Inang
Rata-rata ± Sda) Lama perkembangan
(hari) % Total betina % Pupa mati C. auricilius 8,47 ± 0,81a 41,39 ± 10,56b 66,63 ± 37,09a C. cephalonica 8,07 ± 0,41a 63,59 ± 16,10a 16,45 ± 12,28b a Angka selajur yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan uji Tukey pada
taraf α 0,05.
Hasil penelitian menunjukkan pada inang C. auricilius persentase pupa mati sebesar 66,63%, artinya sebanyak 66,63% dari jumlah telur yang diletakkan tidak mampu berkembang dan muncul menjadi imago. Hal ini menggambarkan rendahnya kemampuan C. auricilius dalam menunjang keberhasilan hidup T. chilonis. Sebaliknya, persentase pupa mati pada inang C. cephalonica hanya sebesar 16,45%, artinya jumlah parasitoid yang berhasil menjadi imago lebih banyak dari jumlah kematian pradewasa. Rendahnya jumlah kematian pradewasa menunjukkan bahwa inang ini sangat baik untuk digunakan sebagai inang pengganti pada pembiakan massal. Selain karena mampu meningkatkan jumlah individu dalam satu populasi secara signifikan, pada saat dilepas di lapangan pun akan lebih banyak imago yang muncul sehingga diharapkan dapat meningkatan efektivitas pengendalian.
Nisbah Kelamin
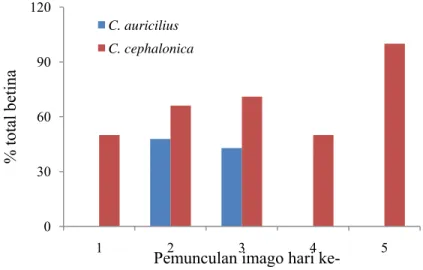
Hasil penelitian menunjukkan bahwa jenis inang mempengaruhi nisbah kelamin dari imago yang muncul. T. chilonis yang berhasil menjadi imago dari telur inang C. auricilius kebanyakan adalah jantan, ditunjukkan dengan persentase total betina sebesar 41,39%. Sedangkan pada C. cephalonica, imago yang muncul kebanyakan adalah betina. Persentase total betina yang muncul yaitu sebesar 63,59% (Tabel 2). Hal ini dapat terjadi karena ukuran telur inang yang berbeda, telur C. cephalonica berukuran lebih besar jika dibandingkan dengan telur C. auricilius. Menurut Walter (2000), imago betina akan meletakkan telur betina pada inang yang relatif besar dan telur jantan pada inang yang relatif kecil.
Persentase rata-rata betina pada hari pertama pemunculan imago rendah, artinya jumlah imago jantan lebih banyak daripada imago betina. Hal tersebut terjadi secara alamiah. Tujuannya agar pada saat imago betina muncul, imago jantan sudah siap kawin, sehingga proses reproduksi dapat berjalan secara maksimal. Hasil penelitian yang dilakukan oleh Susilo et al. (2007) menunjukkan bahwa pada inang C. cephalonica, jenis kelamin T. chilonis dapat diketahui dengan deteksi dini dari warna telur yang terparasit. Warna telur yang lebih terang menghasilkan imago betina, sedangkan warna telur yang lebih gelap menghasilkan imago jantan.
Selain dipengaruhi oleh jenis inang, nisbah kelamin juga dapat dipengaruhi oleh kepadatan inang. Hasil penelitian Khan et al. (2004) menunjukkan persentase total betina yang lebih tinggi terjadi pada kepadatan inang yang lebih rendah. Hal ini disebabkan semua telur diletakkan pada hari pertama, sehingga persentase telur betina lebih tinggi.
Inang yang memiliki nilai persentase total betina lebih tinggi lebih cocok untuk digunakan sebagai inang pengganti pada pembiakan massal di laboratorium. Hal ini dikarenakan semakin banyak imago betina yang dihasilkan dalam satu pias, maka di lapangan akan terjadi peningkatan keberhasilan pengendalian akibat jumlah telur parasitoid yang dihasilkan semakin meningkat. Berdasarkan hasil penelitian diketahui bahwa C. cephalonica lebih baik daripada C. auricilius untuk digunakan sebagai inang pada pembiakan di laboratorium.
Gambar 1 Persentase total imago betina T. chilonis pada dua jenis inang
0 30 60 90 120 1 2 3 4 5 % total betina
Pemunculan imago hari ke-C. auricilius
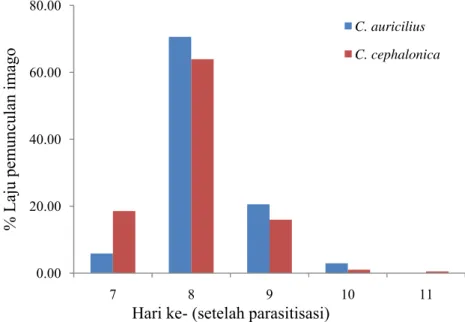
Gambar 2 Persentase laju pemunculan imago T. chilonis pada dua jenis inang
Persentase Laju Pemunculan Imago
Pada kedua inang, imago muncul pertama kali pada hari ke-7 setelah parasitisasi dengan persentase yang rendah. Laju pemunculan imago tertinggi terjadi pada hari ke-8 setelah parasitisasi. Laju pemunculan imago tertinggi berkaitan erat dengan lama perkembangan yang dibutuhkan oleh suatu populasi parasitoid. Laju pemunculan imago tertinggi menunjukkan rata-rata waktu yang dibutuhkan parasitoid pradewasa untuk berkembang.
Setelah hari ke-8, terjadi penurunan persentase laju pemunculan imago. Imago T. chilonis pada inang C. cephalonica paling lambat muncul pada hari ke-11 sehingga pemunculan imago berlangsung selama 5 hari, sedangkan pada inang C. auricilius imago terakhir muncul pada hari ke-10 setelah parasitisasi, artinya pemunculan imago berlangsung lebih singkat daripada C. cephalonica yaitu 4 hari.
Kisaran Inang Parasitoid Telur T. chilonis
Informasi mengenai kisaran inang suatu parasitoid sangat diperlukan untuk mengetahui jenis inang apa saja yang sesuai sebagai inang pengganti pada
0.00 20.00 40.00 60.00 80.00 7 8 9 10 11 % Laju pem unculan im ag o
Hari ke- (setelah parasitisasi)
C. auricilius C. cephalonica
pembiakan massal di laboratorium. Pada program pengendalian hayati, keberadaan inang pengganti di lapangan dapat menunjang T. chilonis agar tetap mampu bertahan hidup ketika populasi inang asli menurun. Hasil penelitian ini menunjukkan bahwa T. chilonis dapat memarasit kelima jenis inang yang diujikan, yaitu C. auricilius, C. cephalonica, C. binotalis, S. litura dan H. armigera meskipun dengan tingkat parasitisasi yang berbeda-beda.
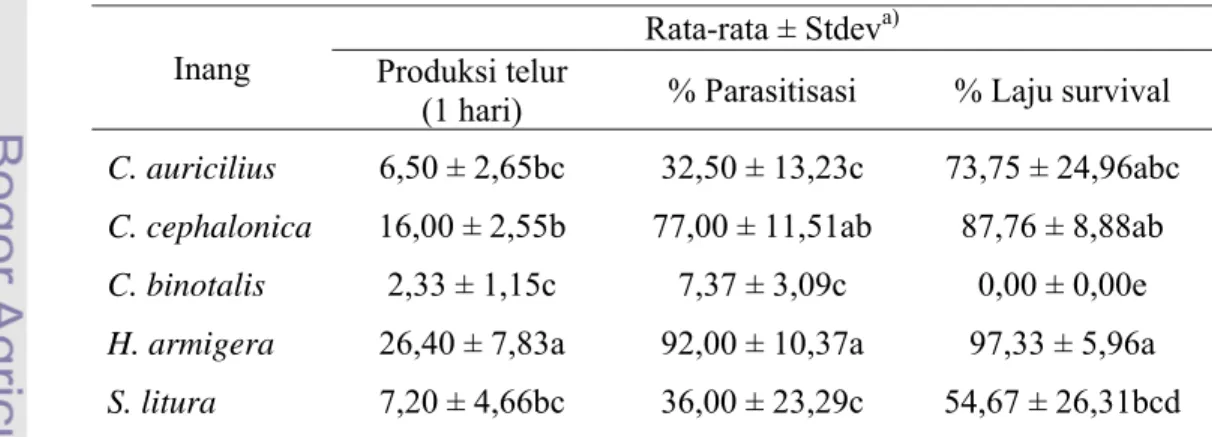
Tingkat Parasitisasi
Data persentase parasitisasi pada tabel 3 menunjukkan bahwa jenis inang mempengaruhi persentase parasitisasi T. chilonis. Pada inang C. binotalis, persentase parasitisasi T. chilonis sangat rendah, yaitu hanya 7,37%. Hal tersebut diduga disebabkan oleh kualitas telur inang yang kurang baik. C. binotalis memiliki ukuran telur yang paling kecil jika dibandingkan dengan empat inang yang lain. Selain itu, nutrisi yang terkandung dalam telur C. binotalis diduga belum memenuhi jumlah yang dibutuhkan untuk perkembangan T. chilonis.
Pada inang aslinya yaitu C. auricilius, T. chilonis hanya mampu memarasit 32,5% dari jumlah telur yang dipaparkan. Hal tersebut menggambarkan ketika T. chilonis hasil pembiakan pada inang C. cephalonica dilepas ke lapangan untuk mengendalikan C. auricilius, persentase parasitisasi yang dihasilkan tidak akan jauh berbeda dari nilai 32,5%. Hal ini berkaitan erat dengan kualitas dari masing-masing inang seperti yang telah dijelaskan sebelumnya. Namun, dalam kaitannya dengan pengendalian hayati di lapangan, persentase parasitisasi dapat meningkat atau menurun dipengaruhi oleh kondisi di sekitar pertanaman.
Berdasarkan tabel 3, persentase parasitisasi pada inang C. auricilius dan C. binotalis sangat rendah. Seperti yang telah diketahui bahwa kedua inang memiliki telur berkelompok dan saling tumpang tindih menyerupai susunan genting. Hal ini diduga menyebabkan sulitnya imago betina T. chilonis menemukan telur yang berkualitas baik di antara sekelompok telur, berbeda dengan telur S. litura, H. armigera dan C. cephalonica yang memiliki telur terpisah satu sama lain sehingga memudahkan imago betina untuk menemukan telur yang memiliki kualitas baik. Corrigan & Laing (1994) menyebutkan bahwa kelompok telur yang saling
tumpang tindih diduga merupakan faktor yang mempengaruhi kesesuaian suatu spesies sebagai inang Trichogramma.
Persentase parasitisasi pada inang S. litura, C. cephalonica dan H. armigera cukup besar, yaitu masing-masing 36%, 77% dan 92%. Nilai tersebut melebihi tingkat parasitisasi pada inang asli T. chilonis, yaitu C. auricilius. Hal ini diduga disebabkan oleh ukuran telur yang lebih besar, nutrisi yang lebih banyak serta kondisi telur yang saling terpisah seperti dijelaskan sebelumnya. Persentase parasitisasi tertinggi terjadi pada inang H. armigera. Hasil ini memiliki kesamaan dengan hasil penelitian yang telah dilakukan sebelumnya oleh Nurafiatin (2000) dan Yuanita (2002).
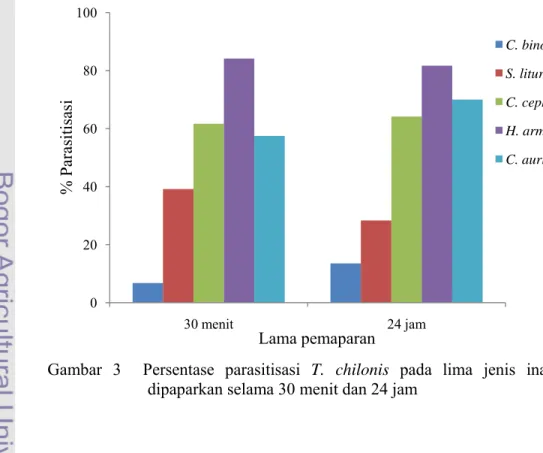
Produksi telur
Hasil analisis sidik ragam menunjukkan bahwa jenis inang mempengaruhi produksi telur. Pada inang H. armigera, T. chilonis mencapai nilai produksi telur tertinggi. Hal ini diduga disebabkan oleh ukuran telur H. armigera yang cukup besar sehingga memungkinkan imago betina meletakkan 2-3 telur pada satu telur inang. Nilai produksi telur pada inang C. cephalonica juga cukup tinggi, karena C. cephalonica merupakan inang yang digunakan untuk pembiakan massal sebelum perlakuan.
Tabel 3 Produksi telur, persentase parasitisasi dan persentase laju survival pada lima jenis inang
Inang
Rata-rata ± Stdeva) Produksi telur
(1 hari) % Parasitisasi % Laju survival C. auricilius 6,50 ± 2,65bc 32,50 ± 13,23c 73,75 ± 24,96abc C. cephalonica 16,00 ± 2,55b 77,00 ± 11,51ab 87,76 ± 8,88ab
C. binotalis 2,33 ± 1,15c 7,37 ± 3,09c 0,00 ± 0,00e
H. armigera 26,40 ± 7,83a 92,00 ± 10,37a 97,33 ± 5,96a
S. litura 7,20 ± 4,66bc 36,00 ± 23,29c 54,67 ± 26,31bcd
a Angka selajur yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan uji Tukey pada
Corrigan & Laing (1994) menyatakan bahwa kemampuan reproduksi imago betina akan meningkat hanya jika spesies inang baru memiliki kualitas yang sama atau lebih baik daripada inang sebelumnya. Berdasarkan pernyataan tersebut, nilai produksi telur pada inang C. auricilius. C. binotalis dan S. litura yang lebih rendah dibandingkan C. cephalonica menunjukkan bahwa kulitas telur ketiga inang tersebut tidak lebih baik daripada C. cephalonica. Sebaliknya, tingginya nilai produksi telur pada inang H. armigera menunjukkan bahwa telur H. armigera memiliki kualitas yang paling baik. Selain itu, ukuran telur H. amigera yang besar memungkinkan imago betina meletakkan lebih dari dua telur pada satu telur inang.
Telur S. litura memiliki ukuran yang cukup besar, tetapi nilai produksi telur T. chilonis pada S. litura tidak lebih besar jika dibandingkan dengan C. cephalonica. Hal ini dikarenakan kualitas inang tidak hanya dicirikan oleh ukuran telur, tetapi juga umur telur dan nutrisi yang terkandung di dalamnya. Semakin tua umur inang, kualitasnya semakin menurun (King 1987 dalam Godfray 1994).
Laju Survival
Persentase laju survival menggambarkan kemampuan hidup T. chilonis pada suatu inang. Hasil analisis sidik ragam menunjukkan bahwa jenis inang mempengaruhi persentase laju survival parasitoid. Pada inang H. armigera persentase laju survival mencapai nilai tertinggi sebesar 97,33%, artinya 97,33% dari jumlah telur yang diletakkan berhasil menjadi imago dewasa. Nilai ini menggambarkan nutrisi yang terkandung dalam telur H. armigera merupakan komposisi yang paling cocok untuk perkembangan T. chilonis. Nilai ini bahkan melebihi persentase laju survival pada inang C. cephalonica yang telah banyak digunakan sebagai inang pengganti pada pembiakan massal di laboratorium.
Pada inang C. binotalis, tidak ada parasitoid yang berhasil menjadi imago, sehingga persentase laju survival yang dihasilkan sebesar 0%. Dari hasil pembedahan, diketahui bahwa parasitoid mati pada fase pupa sehingga disebut dengan istilah pupa mati. Dari hasil tersebut diketahui bahwa C. binotalis hanya mampu sebagai inang sasaran, bukan sebagai inang pengganti pada pembiakan
massal di laboratorium karena tidak mampu menunjang keberhasilan hidup T. chilonis generasi berikutnya.
Pada inang C. auricilius jumlah telur yang berhasil menjadi imago cukup banyak, yaitu hingga mencapai 73,75%. Berbeda dengan S. litura, meskipun nilai produksi telur dan persentase parasitisasinya lebih besar dari C. auricilius, namun kemampuan hidupnya lebih rendah jika dibandingkan dengan C. auricilius. Menurut Djuwarso dan Wikardi (1997), nutrisi yang terdapat dalam telur inang berpengaruh terhadap pertumbuhan dan perkembangan parasitoid pradewasa, telur inang yang tidak cocok nilai nutrisinya, meskipun dapat terparasit dan parasitoid pradewasa dapat berkembang tetapi tidak akan berhasil menjadi imago. Rendahnya tingkat kemampuan hidup pada inang S. litura selain disebabkan oleh nutrisi yang kurang cocok, diduga dapat pula terjadi karena imago tidak mampu melubangi kulit telur dan keluar untuk menyempurnakan perkembangan menjadi imago seutuhnya, dalam hal ini fisiologi dan anatominya telah berkembang sempurna. Selanjutnya, imago yang tidak dapat keluar ini juga disebut dengan pupa mati.
Preferensi Parasitoid Telur T. chilonis pada Lima Jenis Inang
Tingkat Parasitisasi
Persentase parasitisasi lima jenis inang yang terjadi pada lama pemaparan 30 menit dan 24 jam memiliki perbedaan respon, meskipun pada perlakuan keduanya H. armigera menunjukkan respon tertinggi dengan persentase parasitisasi lebih dari 80%. Persentase total parasitisasi diketahui dengan membandingkan jumlah telur terparasit pada satu jenis inang dengan jumlah total seluruh telur yang dipaparkan. Dari tabel 4 diketahui bahwa persentase total parasitisasi tertinggi juga terjadi pada inang H. armigera dengan persentase parasitisasi sebesar 15,03% pada lama pemaparan 30 menit dan 14,85% pada lama pemaparan 24 jam.
Pada pemaparan 30 menit, persentase tertinggi kedua terjadi pada C. cephalonica, sedangkan pada pemaparan 24 jam terjadi pada C. auricilius. Hal ini
menunjukkan adanya bias preferensi akibat lamanya waktu pemaparan. Semakin singkat waktu pemaparan, maka data preferensi yang didapatkan akan semakin baik karena faktor yang menyebabkan data bias semakin rendah. Berdasarkan alasan tersebut, maka dapat ditentukan bahwa T. chilonis lebih banyak memilih C. cephalonica daripada C. auricilius, sesuai dengan hasil yang diperoleh pada lama pemaparan yang lebih singkat yaitu 30 menit.
Dari gambar 3 diketahui bahwa persentase parasitisasi S. litura pada pemaparan 30 menit lebih tinggi daripada pemaparan 24 jam. Seperti yang telah dijelaskan sebelumnya bahwa hal ini diduga terjadi karena variasi kualitas telur pada masing-masing pias. Selain itu, perbedaan respon ini juga dapat disebabkan oleh jumlah rambut-rambut halus yang menutupi telur inang tidak sama. Menurut Beserra et al. (2005) perilaku oviposisi dan efisiensi parasitisasi Trichogramma pada larva dari genus Spodoptera dipengaruhi oleh adanya rambut-rambut halus di sekitar telur. Gambar 3 juga menunjukkan bahwa C. binotalis merupakan pilihan terakhir imago betina untuk meletakkan telur. Hasil ini memiliki kesamaan dengan pengujian sebelumnya yaitu pengujian kisaran inang. T. chilonis mampu memarasit C. binotalis meskipun dengan tingkat parasitisasi yang sangat rendah.
Gambar 3 Persentase parasitisasi T. chilonis pada lima jenis inang yang dipaparkan selama 30 menit dan 24 jam
39,17 6,78 61,67 84,17 57,50 13,57 28,33 64,17 81,67 70,00 0 20 40 60 80 100 30 menit 24 jam % P aras it isa si Lama pemaparan C. binotalis S. litura C. cephalonica H. armigera C. auricilius
Tabel 4 Persentase total parasitisasi T. chilonis pada lima jenis inang
Inang Lama pemaparan
a)
30 menit 24 jam
C. auricilius 10,38 ± 6,37abc 12,76 ± 4,81ab
C. cephalonica 10,99 ± 2,03ab 11,75 ± 2,19abc
C. binotalis 1,83 ± 0,01d 3,65 ± 2,25d
H. armigera 15,03 ± 3,28a 14,85 ± 3,49a
S. litura 7,04 ± 2,87bcd 5,17 ± 2,31d
a Angka selajur yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan uji Tukey pada
taraf α 0,05.
Produksi telur
Hasil penelitian menunjukkan adanya kemiripan respon dari kedua perlakuan lama pemaparan. Secara umum terjadi peningkatan jumlah telur yang diletakkan seiring dengan peningkatan lama pemaparan, meskipun peningkatan tersebut tidak terlalu besar. T. chilonis meletakkan telur lebih banyak pada H. armigera jika dibandingkan dengan empat jenis inang lainnya. Telah dijelaskan pada subbab sebelumnya bahwa hal ini terjadi karena kualitas telur H. armigera lebih baik dari jenis inang lainnya. Selain itu, ukuran telur H. armigera juga memungkinkan terjadinya superparasitisme, dimana imago betina parasitoid meletakkan lebih dari satu telur dalam satu telur inang.
Pada lama pemaparan 30 menit produksi telur pada C. cephalonica dan C. auricilius masing-masing 12,33 dan 11,50. Penambahan waktu pemaparan menyebabkan peningkatan nilai produksi telur masing-masing menjadi 14,00 dan 14,00. Hal ini terjadi karena semakin lama waktu pemaparan maka semakin banyak pula kesempatan imago betina untuk menemukan inang lain.