Control of interaction strength in a network of the true
slime mold by a microfabricated structure
Atsuko Takamatsu
a,*, Teruo Fujii
a b, Isao Endo
aaBiochemical Systems Laboratory,RIKEN
(The Institute of Physical and Chemical Research),2-1,Hirosawa,Wako-shi,Saitama, 351-0198,Japan
bInstitute of Industrial Science,Uni6ersity of Tokyo,7-22-1,Roppongi,Minato-ku,Tokyo,106-8558,Japan
Abstract
The plasmodium of the true slime mold, Physarum polycephalum, which shows various nonlinear oscillatory phenomena, for example, in its thickness, protoplasmic streaming and concentration of intracellular chemicals, can be regarded as a collective of nonlinear oscillators. The plasmodial oscillators are interconnected by microscale tubes whose dimensions can be closely related to the strength of interaction between the oscillators. Investigation of the collective behavior of the oscillators under the conditions in which the interaction strength can be systematically controlled gives significant information on the characteristics of the system. In this study, we proposed a living model system of a coupled oscillator system in thePhysarumplasmodium. We patterned the geometry and dimensions of the microscale tube structure in the plasmodium by a microfabricated structure (microstructure). As the first step, we constructed a two-oscillator system for the plasmodium that has two wells (oscillator part) and a channel (coupling part). We investigated the oscillation behavior by monitoring the thickness oscillation of the plasmodium in the microstructure with various channel widths. It was found that the oscillation behavior of two oscillators dynamically changed depending on the channel width. Based on the results of measurements of the tube dimensions and the velocity of the protoplasmic streaming in the tube, we discuss how the channel width relates to the interaction strength of the coupled oscillator system. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Physarum polycephalum; Coupled oscillators; Microfabrication
www.elsevier.com/locate/biosystems
1. Introduction
Nonlinear dynamics becomes important for the studies of collective cells, for example, cardiac cells, neurons and other oscillatory or excitable cells (Winfree, 1980). In these biological systems,
the connections between the elements are not highly designed individually. Nevertheless, the collectives of such elements self-organize to show various biological functions, such as information processing in nervous systems and cooperative beating in cardiac systems. In order to understand the mechanism of biological self-organization, it is important to investigate the behavior of the sys-tems under conditions in which the connections of the elements can be systematically controlled. In * Corresponding author. Tel: +81-48-4679312; fax: +
81-48-4624658.
E-mail address:[email protected] (A. Takamatsu)
this study, we adopted the true slime mold,
Physarum polycephalum, as a living nonlinear element.
The plasmodium of P. polycephalum is a huge amoeboid multinucleated unicellular organism. The plasmodium is an aggregate of endoplasm without any highly differentiated structure like a nervous system. Multiple plasmodia can be fused into a single one and a single plasmodium can be easily divided into multiple ones. Despite its sim-ple structure, the plasmodium demonstrates so-phisticated biological functions. Namely, a large cluster of the plasmodium is able to behave as a single entity based on the information from its surroundings. The information acquired by a lo-cal partial body drives the cooperative behavior by transmitting the information throughout the whole cluster. The information is considered to be processed through the interaction among the par-tial bodies. This leads to, for example, the attract-ing/escaping behavior of the plasmodium to/from attractants/repellents (Knowles and Carlile, 1978). These functions, information transmission and processing, are said to be realized through the intrinsic oscillatory phenomena in the plasmod-ium, such as thickness oscillation (Baranowski, 1978) which synchronizes with the shuttle stream-ing of the protoplasm (Kamiya, 1950), and the oscillation in the concentrations of ATP (Yoshi-moto et al., 1981) and Ca2+ (Natsume et al.,
1992). These phenomena are supposed to be
gen-erated by mechanochemical reactions caused by complicated interactions among intracellular chemicals, proteins, organelles, etc. (Ueda et al., 1986).
From the viewpoint of nonlinear dynamics, the plasmodium can therefore be modeled as a collec-tive system that consists of nonlinear oscillators interconnected with certain interaction strength. In this model, an oscillator can be defined for each partial body in the plasmodium, and the interaction among the oscillators would be related to the protoplasmic streaming observed inside the tube structures which interconnect the partial bodies. It is therefore expected that, by con-trolling the growth pattern and the dimensions of the tube structure, we could observe various types of collective behavior which could help us to understand the characteristics of the plasmodium as a nonlinear dynamic system. In this paper, we describe the advanced patterning method of the plasmodium by a microfabricated structure (mi-crostructure) and the results of observation of the plasmodium in the microstructure for a two-oscil-lator system, as the first step.
2. Materials and methods (Takamatsu et al., 1998, 2000)
2.1. Geometry control of the plasmodium
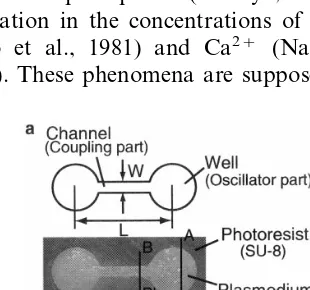
The plasmodium prefers a wet medium such as an agar plate (1.5%), on which the plasmodium is usually cultured, to a dry one. It is possible to control the geometry of the plasmodium by using the agar plate covered with a dry medium having variously designed openings. As the dry medium, we adopted an ultra-thick negative photoresist resin (NANO™ SU-8 50, Microlithography Chemical) which enabled us to fabricate mi-crostructures with about 10 mm precision that is the same order as the tube diameter of the plas-modium in the natural situation. The microstruc-tures for the two-oscillator system is a thick sheet (100 mm) having an opening consisting of two circular wells (oscillator part) connected by a channel (coupling part) as shown in Fig. 1. The diameters of the wells are determined to be 2 mm Fig. 1. Microstructure for coupled plasmodial oscillators. (a)
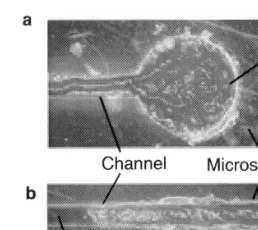
Fig. 2. Growing process of the plasmodium on the microstruc-ture.
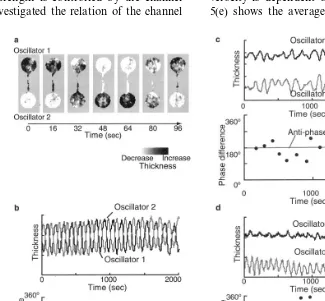
In order to enhance the change in thickness in the images obtained by the above method, we calculated differential images between the two images at time t andt+Dt (Dt=4 s), where the black and white images mean an increase and decrease in the thickness of the plasmodium, re-spectively (Fig. 3(a)).
2.3. Measurement of the tube diameter and
6elocity of the protoplasmic streaming
The plasmodia with the microstructure were placed under a inverted microscope (TMD series, Nikon, objective lens ×10 or ×20) and the protoplasmic streaming in the tube structure was observed. The images were recorded by the VCR (CCD-TRV92, Sony). The tube diameter was defined as the width of the area where the proto-plasmic streaming was observed. The velocity of the protoplasmic streaming was determined by measuring the time that organelles observed at around the center of the tube structure passed a certain distance on the video monitor.
3. Results and discussion
The thickness oscillation of the plasmodium on the microstructure for the two-oscillator system was observed over various channel width (W) ranges from 50 to 800 (mm) where the channel length was fixed at L=4 (mm). Fig. 3(a) shows the spatio-temporal patterns of the change in the thickness oscillation when W=400 (mm). Alter-nating wave propagation from one plasmodial oscillator to another was observed. The upper graph of Fig. 3(b) shows the time course of the thickness of each oscillator that was obtained by averaging the images over each well. The lower graph of Fig. 3(b) shows the time course of the phase difference between the two oscillators that was determined by a conventional method (for example, Nakata et al., 1998). These graphs demonstrate that the two oscillators are synchro-nized and the phases are locked in anti-phase. WhenWis smaller (W=200 – 300mm), the phase difference sometimes comes off the anti-phase line (180°) as shown in the lower graph of Fig. 3(c), (Fig. 1(b)), so that we can consider the
plasmod-ium in a well as a single oscillator. The size is comparable to half of the spatial wavelength of oscillation in the natural plasmodium and, within this area, the plasmodium shows almost synchro-nized oscillation.
The plasmodium was cultured on 1.5% agar medium with oatmeal as the food. The tip por-tions of the plasmodium were placed at the both wells in the microstructure on the agar plate. The plasmodia in the wells grew along the channel pattern during the culture for 5 – 15 h under the conditions of 25°C and RH 85%, as shown in Fig. 2. Finally, the plasmodia from both sides of the wells contacted at around the center of the chan-nel to be physically connected through the tube structure (Fig. 1(a)), where the endoplasmic streaming was observed.
2.2. Obser6ation of the thickness oscillation
which can be categorized as quasi-periodic oscilla-tion. WhenWis much smaller (W=50 – 100mm), the oscillators were not synchronized and oscil-lated in their own periods (Fig. 3(d)). In the extremely narrow channel below 50mm in width, the tube structure was never formed. On the contrary, in a wide channel of more than 400mm in width, the oscillations became very compli-cated. Here, the wave propagation was observed in directions other than the major axis of the channel.
This oscillation behavior depending on the channel width suggests that the interaction strength between the oscillators is controlled by the channel dimension of the microstructure in a systematic manner. In order to confirm how the interaction strength is controlled by the channel width, we investigated the relation of the channel
width to the diameter of the tube structure in the channel. The plasmodium in the channel formed a single tube structure up to W=400 mm and the diameter increased depending on W as shown in Fig. 4. Beyond this critical width, the increase in the tube diameter became saturated and the plas-modium in the channel formed a dendritic tube structure (Takamatsu et al., 2000). The compli-cated oscillation behavior in the wide channel would be caused by the dendritic tube structure. Hereafter, we considered only the case under the critical channel width (W=400 mm).
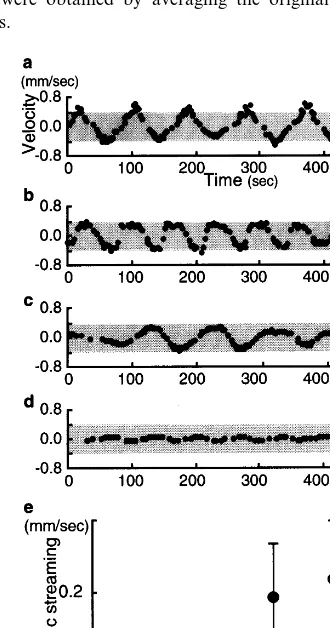
Fig. 5(a – d) show the time course of the velocity of the protoplasmic streaming with various chan-nel widths, where periodic changes in the velocity can be observed. In addition, the amplitude of the velocity is dependent on the channel width. Fig. 5(e) shows the averaged absolute velocity of the
Fig. 4. Relation of channel width W to tube diameter. The data were obtained by averaging the original data over ten points.
This result indicates that the amount of proto-plasm transported by the streaming is controlled by the channel width.
The amount transported by the protoplasmic streaming is estimated to be about 0.1 ml per half of the period of oscillation, when W=400
mm (the tube diameter is about 100 mm and the velocity is about 0.25 mm/s). On the other hand, the amount of the protoplasm changed accord-ing to the thickness oscillation in the oscillator part, whose change was about 9100 mm in our observation, is estimated to be about 0.3 ml. This is the same order as the estimated amount calcu-lated from the velocity of the protoplasmic streaming. It can therefore be considered that the protoplasmic streaming directly affects on the change in the thickness of the oscillator parts. The anti-phase oscillation which appears in the system with the relatively large channel width (W=400 mm) can be explained by the fact that the effects of the transportation of the pro-toplasm on the thickness in the oscillator parts are reciprocal. Consequently, the thickness in one oscillator part increases, and that in another decreases.
It is concluded that the interaction strength between the oscillators is closely related to the amount of the protoplasm transported by the streaming and, accordingly, is also related to the channel width.
4. Conclusion
In conclusion, we succeeded in controlling the growth pattern of the Physarum plasmodium by the microfabricated structure. Using this mi-crostructure for a two-oscillator system, we eluci-dated that the strength of interaction among the plasmodia depends on the tube dimensions which can be systematically controlled by the channel width.
With this system, it is also possible to inte-grate a stimulator and observe the response to stimuli in a systematic way. Furthermore, rich oscillation behavior will be observed using the 2-D microstructures as shown in Fig. 6, which gives us further information about the system as a collective nonlinear oscillator system.
Fig. 5. Relation of channel widthWto protoplasmic streaming inside the tube. (a)W=400mm, (b)W=300mm, (c)W=200 mm, (d) W=100 mm. (e) Averaged velocity of the absolute value over 10 periods of oscillation for each channel width.
Fig. 6. Multiple-oscillator system. (a) Triple oscillators. (b) Multiple oscillators.
Kamiya, N., 1950. The rate of the protoplasmic flow in the myxomycete plasmodium. II. Cytologia 15, 194 – 204. Knowles, D.J.C., Carlile, M.J., 1978. The chemotactic
re-sponse of plasmodia of the myxomycete Physarum poly-cephalum to sugars and related compounds. J. Gen. Microbiol. 108, 17 – 25.
Nakata, S., Miyata, T., Ojima, N., Yoshikawa, K., 1998. Self-synchronization in coupled salt-water oscillators. Physica D 115, 313 – 320.
Natsume, K., Miyake, Y., Yano, M., Shimizu, H., 1992. Development of spatio-temporal pattern of Ca2+ on the
chemotactic behavior of Physarum plasmodium. Proto-plasma 166, 55 – 60.
Takamatsu, A., Fujii, T., Hosokawa, K., Endo, I., 1998. Microfabricated structure for nonlinear dynamics obser-vation of cell network, Proceedings for the 20th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, pp. 2987 – 2990.
Takamatsu, A., Fujii, T., Yokota, H., Hosokawa, K., Higuchi, T., Endo, I., 2000. Controlling the geometry and the coupling strength of the oscillators system in Physarumplasmodium by microfabricated structure. Pro-toplasma, in press.
Ueda, T., Matsumoto, K., Akitaya, T., Kobatake, Y., 1986. Spatial and temporal organization of intracellular adenine nucleotides and cyclic nucleotides in relation to rhythmic motility in Physarum plasmodium. Exp. Cell. Res. 162, 486 – 494.
Winfree, A.T., 1980. The Geometry of Biological Time. Springer-Verlag, NY, Heiderberg, Berlin.
Yoshimoto, Y., Sakai, T., Kamiya, N., 1981. ATP oscilla-tion in Physarum plasmodium. Protoplasma 109, 159 – 168.
Acknowledgements
The authors thank Dr Hosokawa for helping in the microfabrication process. This study was partly supported by Special Coordination Funds for promoting Science and Technology of the Science and Technology Agency of the Japanese Government.
References
Baranowski, Z., 1978. The contraction-relaxation waves in Physarum polycephalum plasmodia. Acta Protozool. 17, 377 – 388.