LAPORAN SKRIPSI
Taksonomi kimiawi dan molekular isolat Aktinomisetes dari
Jambi, Timor dan Lombok
Oleh :
Cornellius Yudha Wijaya 11/316157/BI/08737
Pembimbing I : Prof. Dr. Langkah Sembiring, M.Sc. Pembimbing II : Dr. Endah Retnaningrum, S. Si, M. Eng.
FAKULTAS BIOLOGI
UNIVERSITAS GADJAH MADA
YOGYAKARTA
TAKSONOMI KIMIAWI DAN MOLEKULAR ISOLAT
AKTINOMISETES DARI JAMBI, TIMOR DAN LOMBOK
SKRIPSI
Untuk memenuhi sebagai persyaratan
guna mencapai derajad Sarjana Sains
Program Studi Biologi
Oleh :
Cornellius Yudha Wijaya 11/316157/BI/08737
Pembimbing I :
Prof. Dr. Langkah Sembiring, M.Sc. NIP. 195905011985031003
Pembimbing II :
Dr. Endah Retnaningrum, S. Si, M. Eng. NIP. 197203191999032002
FAKULTAS BIOLOGI
UNIVERSITAS GADJAH MADA
YOGYAKARTA
HALAMAN PENGESAHAN
Taksonomi Kimiawi dan Molekular Isolat Aktinomisetes dari
Jambi, Timor dan Lombok
Oleh :
Cornellius Yudha Wijaya 11/316157/BI/08737
Telah dipertahankan di depan Tim Penguji
Pada tanggal 9 Oktober 2015
dan dinyatakan telah memenuhi syarat
Yogyakarta, Oktober 2015
Fakultas Biologi
Universitas Gadjah Mada
Tim Penguji
Pembimbing Skripsi I Dekan Fakultas Biologi
Prof. Dr. Langkah Sembiring, M.Sc. Prof. Dr. Suwarno Hadisusanto, S.U.
Pembimbing Skripsi II
Dr. Endah Retnaningrum, S Si., M. Eng.
Penguji II
Rina Sri Kasiamdari, S.Si., Ph.D.
Penguji III
HALAMAN PERYATAAN
Dengan ini saya menyatakan skripsi ini tidak terdapat karya yang pernah diajukan untuk memperoleh gelar kesarjanaan di suatu Perguruan Tinggi, dan sepanjang pengetahuan saya juga tidak terdapat karya atau pendapat yang pernah dituliskan atau diterbitkan oleh orang lain, kecuali yang tertulis diacu dalam naskah ini dan disebutkan dalam daftar pustaka.
Yogyakarta, 28 Oktober 2015
PRAKATA
Laporan ini disusun dalam rangka memenuhi tugas mata kuliah Skripsi BIO 5993 dengan bobot 6 SKS berdasarkan usulan penelitian dengan judul Karakterisasi dan Identifikasi Isolat Aktinomisetes Indonesia dengan Metode Kemotaksonomis. Dengan selesainya laporan skripsi ini penulis ingin menyampaikan ucapan terima kasih kepada berbagai pihak yaitu Pertama-tama kepada Bapak Prof. Dr. Langkah Sembiring, M.Sc. dan ibu Dr. Endah Retnaningrum, S. Si., M. Eng. sebagai pebimbing skripsi yang telah banyak memberikan arahan dalam menyelesaikan laporan skripsi, Ibu Dr. Puspita Lisdiyanti, M. Agr. Chem. yang telah banyak memberikan arahan dan bantuan di dalam penelitian ini serta memberikan ijin untuk melakukan penelitian di LIPI Cibinong, Ibu Dra. Siti Susanti, S.U. dan Ibu Dwi Umi Siswanti, S.Si., M.Si. selaku penyelenggara skripsi yang telah memfasilitasi penulis untuk melaksanakan skripsi, ibu Rina Sri Kasiamdari S.Si., Ph.D. dan bapak Dr. Budi Setiadi Daryono M.Agr.Sc. selaku penguji yang telah banyak memberikan arahan dan bantuan di dalam menuliskan laporan ini serta membantu penulis untuk melaksanakan ujian sidang skripsi . Penulis juga ingin mengucapkan terima kasih bagi seluruh pihak yang telah membantu dalam penyusunan laporan ini.
Yogyakarta, Oktober 2015
DAFTAR ISI
BAB II. TINJAUAN PUSATAKA DAN HIPOTESIS... 4
A. Tinjauan Pustaka... 5
A. Tempat dan Waktu Penelitian... 27
B. Strain Bakteri... 27
C. Kultur Isolat... 27
D. Persiapan Isolat untuk Analisis Kemotaksonomi dan Molekular... 27
E. Analisis Kemotaksonomi... 28
F. Analisis Molekular... 30
G.
Generic Assignment... 35H.
Analisis Data... 35BAB IV. HASIL DAN PEMBAHASAN... 36
BAB V. SIMPULAN DAN SARAN... 57
A. Simpulan... 57
B. Saran... 57
DAFTAR PUSTAKA... 58
Taksonomi Kimiawi dan Molekular Isolat Aktinomisetes dari Jambi, Timor dan Lombok
Oleh :
Cornellius Yudha Wijaya 11/316157/BI/08737
INTISARI
Aktinomisetes merupakan kelompok bakteri yang memiliki kemiripan dengan fungi. Aktinomisetes menyumbang sebesar 45% metabolit sekunder yang telah ditemukan atau sekitar 10000 metabolit sekunder. Meskipun demikian informasi mengenai aktinomisetes seperti ciri khas maupun manfaatnya telah diketahui tetapi analisis secara kemotaksonomi dan molekular perlu untuk dipelajari. Hal ini dikarenakan klasifikasi aktinomisetes sering mengalami perubahan seiring dengan banyaknya penelitian dan studi yang dilakukan. Tujuan penelitian ini adalah mempelajari keanekaragaman hingga tingkat genus isolat aktinomisetes Indonesia dari koleksi kultur LIPI Cibinong menggunakan metode kemotaksonomi dan molekular. Penelitian ini dilakukan di LIPI Cibinong, Bogor selama 6 bulan. 10 isolat aktinomisetes digunakan dalam penelitian kali ini dipersiapkan untuk analisis kemotaksonomi dan molekular. Isolat ID04-1019,ID04-1052, ID04-1066, ID04-1067, ID04-1071, ID04-1088, ID04-1093, ID04-1103 diisolasi dari seresah daun di Jambi, Isolat ID05-0653 dari tanah di Timor dan Isolat ID06-0464 dari tanah di Lombok. Penelitian ini menggunakan metode kemotaksonomi yaitu analisis diamino pimelic acid, analisis gula sel, analisis asam lemak, analisis menaquinon, analisis fosfolipid serta metode molekular yaitu sekuensing gen 16S rRNA dan determinasi G+C Content. Hasil analisis menggunakan pohon filogenetik menunjukkan bahwa isolat ID04-1071 memilki kedekatan dengan spesies Actinoplanes rectilleneatus, isolat ID04-1066, ID04-1052 dan ID04-1033 memiliki kedekatan dengan spesies Actinoplanes ferrugineus dan isolat ID04-1067, ID04-1093 dan ID04-1088 memiliki kedekatan dengan spesies Actinoplanes globisporus. Hasil keseluruhan menunjukkan isolat 1052, 1066, 1067, 1071, 1088, 1093, ID04-1103 merupakan anggota genus Actinoplanes. Isolat ID05-0653 dan ID-0464 merupakan anggota genus Actinophytocola.
Chemotaxonomy and Molecular Taxonomy Actinomycetes Isolate from Jambi, Timor and Lombok
By :
Cornellius Yudha Wijaya 11/316157/BI/08737
ABSTRACT
Actinomycetes is bacterial group that have similarity to Fungi. Actinomycetes contribute around 45% secondary metabolite that has been found or around 10000 secondary metabolite. Even though information about actinomycetes such as unique trait or the benefit of it is known already but cehmotaxonomy analysis and molecular analysis still need to be learn. This thing happen because classification of actinomycetes always changing following the research and study that has been done. The aim for this research is to studying diversity until genus level actinomycetes Indonesia LIPI Cibinong culture collection using chemotaxonomy and molecular methods. This research was done in the LIPI Cibinong Bogor for 6 months. 10 isolate actinomycetes that was used in this research are being prepare for chemotaxonomy and molecular analysis. Isolate ID04-1019,ID04-1052, ID04-1066, ID04-1067, ID04-1071, ID04-1088, ID04-1093, ID04-1103 was isolated from leaves in Jambi, Isolate ID05-0653 was from soil in Timor and isolate ID06-0464 was from soil in Lombok. This research using chemotaxonomy method that is Diamino pimelic acid analysis, cell sugar analysis, fatty acid analysis, menaquinone analysis, phospholipid analysis also molecular method that is gen sequencing of 16S rRNA and G+C Content determination. Analysis result using phylogenic tree showed that isolate ID04-1071 related to species Actinoplanes rectilleneatus, isolate ID04-1066, ID04-1052 dan ID04-1033 related to species Actinoplanes ferrugineus and isolate ID04-1067, ID04-1093 and ID04-1088 related to species Actinoplanes globisporus. Result showed isolate 1052, 1066, 1067, 1071, 1088, ID04-1093, ID04-1103 are genus member of Actinoplanes. Isolate ID05-0653 and ID-0464 are genus member of Actinophytocola.
DAFTAR TABEL
Halaman
Tabel 1. Tahap-tahap klasifikasi numerik pada analisis bakteri 7 Tabel 2. Analisis bakteri dengan secara taksonomi numerik 8 Tabel 3. Klasifikasi berdasarkan tipe dinding sel strain anggota
aktinomisetes melalui metode kemotaksonomis
12
Tabel 4. Analisis melalui metode molekular 16
Tabel 5. Beberapa produk metabolit primer aktinomisetes 20 Tabel 6. Beberapa produk metabolit sekunder aktinomisetes 21 Tabel 7. Ukuran genom DNA anggota genus Streptomisetes 22 Tabel 8. Struktur gen biosintetetik antibiotik pada kromosom dan
ekstrakromosom aktinomisetes 23
Tabel 9. Produk biokonversi oleh aktinomisetes 25 Tabel 10. Kondisi HPLC untuk Derminasi G+C Content 32 Tabel 11. Primer yang digunakan untuk analisis sequencing gen
16S rRNA
33
Tabel 12. Kondisi PCR untuk mendapatkan produk PCR 33 Tabel 13. Kondisi PCR untuk sequence gen 16S rRNA 34 Tabel 14. Isolat yang digunakan di dalam penelitian 36
Tabel 15. Analisis DAP menggunakan TLC 40
Tabel 16. Analisis gula sel menggunakan TLC 41
Tabel 17. Analisis menaquinon menggguakan LC-MS 42 Tabel 18. Analisis asam lemak menggunakan GC-MS 43 Tabel 19. Analisis fosfolipid menggunakan TLC 44 Tabel 20. Hasil pencarian isolat menggunakan BLAST 46
Tabel 21. Analisis G+C content 54
Tabel 22. Karakteristik khas pada genus Actinoplanes 54 Tabel 23. Analisis kemotaksonomi dan profil genetik pada isolat
ID05-0653 dan isolat ID06-0464. 55
DAFTAR GAMBAR
Halaman
Gambar 1. Sistem hierakri taksonomi yang sekarang digunakan 5 Gambar 2. Dendogram dengan menggunakan komputer 9 Gambar 3. Pohon filogenetik berdasarkan 16S/18S rRNA 10 Gambar 4. Representasi skematik siklus hidup anggota genus
Streptomyces
19
Gambar 5. Proses bioprospek aktinomisetes 19
Gambar 6.Gambar Isolat pada Medium ISP2 Padat. A. Isolat ID04-1019, B. B. Isolat ID04-1052, C. Isolat ID04-1066, D. Isolat ID04-1067, E. Isolat E. ID04-1071, F. Isolat ID04-1088, G. Isolat ID04-1093, H. ID04-H. 1103, I. ID05-0653, J. ID-06-0464.
37
Gambar 7. Hasil TLC untuk analisis DAP isolat pada TLC plastic sheet
39
Gambar 8. Interface program MEGA6 48
Gambar 9. Hasil alignment sequence gen 16S rRNA menggunakan ClustalW
48
Gambar 10. Parameter analisis untuk pembuatan pohon filogenetik pada program MEGA6
49
Gambar 11. Pohon Filogenetik 7 isolat dengan mennggunakan
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Medium ISP2 62
BAB I. PENDAHULUAN
A. Latar Belakang
Mikroorganisme merupakan makhluk hidup dengan tingkat keanekaragaman yang tinggi dan mampu baik di udara, tanah ataupun air. Selain itu mikroorganisme dapat hidup dalam berbagai macam kondisi seperti kurangnya oksigen, salinitas tinggi dan sebagainya (Pelczar & Chan, 1981). Mikroorganisme dengan kemampuannya sebagai penghasil metabolit yang memiliki diversitas tinggi telah digunakan oleh manusia didalam berbagai bidang seperti kesehatan, lingkungan, pertanian dan industri (Furest, 2014). Berkaitan dengan hal tersebut salah satu mikroorganisme yang penting dari segi ilmu pengetahuan untuk dipelajari adalah aktinomisetes yang terkenal di dunia industri sebagai penghasil metabolit sekunder penting yaitu antibiotik (Maheshwari et al., 2010).
Aktinomisetes merupakan kelompok bakteri yang memiliki kemiripan dengan fungi (Crawfor et al., 1993). Bakteri ini dapat ditemukan melimpah di tanah ataupun seresah daun dan bakteri tersebut berperan untuk membantu menguraikan sisa tanaman atau hewan yang telah mati. Didalam dunia bioteknologi, aktinomisetes menjadi salah satu objek yang diminati untuk dipelajari dari berbagai bidang seperti kesehatan (Goodfellow et al., 1988).
Aktinomisetes menarik untuk dipelajari dikarenakan memiliki kemampuan sebagai sumber enzim yang baik dibandingkan dengan sumber lainnya. Hal ini dikarenakan aktinomisetes memiliki tingkat kestabilan yang tinggi, mampu berkativitas pada suhu tinggi dan spesifikasi substrat yang berbeda dengan mikroorganisme lainnya (Goodfellow et al., 1988).
dengan cara mencari isolat baru di berbagai tempat seperti permukaan tanah sampai kedalamlaut yang dalam (Sharma, 2014).
Walau telah banyak informasi yang diketahui mengenai bakteri tersebut seperti ciri khas maupun manfaatnya tetapi masih banyak yang perlu untuk dipelajari. Berkaitan dengan hal tersebut salah satu yang diperlukan untuk dipelajari lebih lanjut adalah klasifikasi dari aktinomisetes. Hal ini dikarenakan klasifikasi aktinomisetes sering mengalami perubahan seiring dengan banyaknya penelitian dan studi yang dilakukan (Goodfellow et al., 1982). Selain itu metode pengklasifikasian juga telah banyak mengalami kemajuan dari awal dikenalnya aktinomisetes oleh Waksman pada tahun 1940 dengan mengamati perbedaan morfologi hingga perkembangan sekarang dengan menggunakan metode molekular sehingga dapat diketahui perbedaan setiap isolat yang didapatkan (Waksman & Henrichi, 1943).
Di Indonesia telah banyak dilakukan penelitian untuk mencari isolat baru beserta aplikasinya di mana penelitian mengenai aktinomisetes di Indonesia telah diawali dengan penemuan beberapa strain seperti Streptomyces asiaticus, cangkringnensis, indonesiensis, javensis, rhizosphaerius dan yogyakartensis. Oleh karena itu dengan penemuan strain baru ini,maka membuka banyak kesempatan mengenai potensi penelitian aktinomisetes di Indonesia (Sembiring et al., 2000). Salah satu upaya lain untuk pengembangan aktinomisetes lokal Indonesia telah dilakukan sejak tahun 2003 dan telah ditemukan banyak isolat yang memiliki potensi penghasil berbagai macam enzim. Berkaitan dengan hal tersebut, para peneliti telah meneliti mengenai aktinomisetes dan telah berhasil melaksanakan isolasi aktinomisetes dari berbagai tempat di Indonesia sehingga jumlah isolat aktinomisetes yang telah diisolasi dari beberapa daerah di Indonesia dari tahun 2003 sampai dengan 2008 yaitu sekitar 3193 isolat (Wahyuni, 2014)
digunakan untuk pengklasifikasikasian aktinomisetes karena mampu memberikan hasil klasifikasi yang lebih baik untuk bakteri, mampu melihat kedekatang antara bakteri, menelaah aspek evolusi mereka dan mengidentifikasi organisme penting secara klinis. (Brondz, 1986). Selain itu pendekatan secara molekular juga terbukti mampu memberikan hasil klasifikasi yang mampu mendekati hasil klasifikasi yang diiberikan oleh klasifikasi secara metode lain sehingga metode klasifikasi secara molekular mampu mendukung hasil klasifikasi yang didapatkan melalui metode kemotaksonomi (Singh, 2012).
B. Permasalahan
1. Apakah isolat aktinomisetes koleksi LIPI Cibinong dapat diindentifikasi melalui metode khusus kemotaksonomis analisis DAP, analisis gula sel, asam lemak, menaquinon, fosfolipid dan molekular?
2. Bagaimanakah kenaekaragaman hingga tingkat genus isolat aktinomisetes Indonesia dari koleksi kultur LIPI menggunakan pendekatan kemotaksonomis metode khusus kemotaksonomis anallisis DAP, gula sel, asam lemak, menaquinon, fosfolipid dan molekular?
C. Tujuan
Tujuan penelitian ini adalah untuk untuk mengetahui keanekaragaman hingga tingkat genus isolat aktinomisetes Indonesia dari koleksi kultur LIPI menggunakan pendekatan kemotaksonomis metode khusus kemotaksonomis anallisis DAP, gula sel, asam lemak, menaquinon, fosfolipid dan molekular
D. Manfaat
Diharapkan dari penelitian ini dapat diketahui secara jelas sifat kimiawi yang dimiliki oleh aktinomisetes strain lokal Indonesia secara jelas dan klasifikasinya sehingga dapat membantu untuk penelitian yang akan datang khususnya mengenai aktinomisetes. Selain itu dengan dikenalnya isolat aktinomisetes Indonesia hingga tingkat Genus dapat diketahui potensi yang dimiliki dan dapat diketahui plasma nutfah yang dimiliki oleh Indonesia.
A. Tinjauan Pustaka
1. Klasifikasi Mikroorganisme
Taksonomi adalah cabang ilmu pengetahuaan yang mempelajari penamaan, pendeskripsian dan klasifikasi dari suatu organisme makhluk hidup dan taksonom menggunakan pengamatan morofologi, genetik dan biokimia untuk membantu kegiatan taksonomi (Anonim, 2014).
Taksonomi dapat didefinisikan sebagai studi dan deskripsi dari variasi suatu organisme, mengapa bisa terjadi variasi dan akibat dari variasi tersebut serta menganalisis data yang didapatkan untuk menghasilkan suatu sistem klasifikasi (Stace, 1989). Oelh karena itu taksonomi dapat disimpulkan sebagai cabang dari ilmu pengetahuan yang mempelajari penamaan, deskripsi serta variasi baru suatu organisme untuk menghasilkan sistem klasifikasi (Anonim, 2014).
Klasifikasi adalah suatu metode yang digunakan oleh peneliti untuk mengelompokkan suatu organisme dan memasukkanya ke dalam suatu sistem hierakri. Selain itu, identifikasi adalah penamaan dari suatu organisme dengan menggunakan referensi suatu klasifikasi yang ada (Stace, 1989).
Domain
Kingdom
Phyllum
Class
Ordo
Family
Genus
Species
Gambar 1. Sistem hierakri yang sekarang digunakan (Glodberg, 2010)
Selain itu, pembagian domain sekarang telah dilakukan berdasarkan analisis sequence gen 16S rRNA sehingga sekarang telah dibagi menjadi 3 domain yaitu Eukarya, Archaea dan Bacteria.
Di dalam penelitian mengenai sistematika mikroorganisme, hal yang penting untuk dilakukan yaitu memahami konsep dasar mengenai klasifikasi mikroorganisme. Klasifikasi mikroorganisme merupakan penyusunan mikroorganisme kedalam suatu kelompok berdasarkan kemiripan atau perbedaan yang dimiliki (Priest & Goodfellow, 2000). Hasil dari kegiatan ini merupakan suatu sistem klasifikasi yang menggambarkan hubungan antar organisme dan menyimpan informasi menganai organisme tersebut. Hubungan ini biasa berdasarkan dari kombinasi data genotipik dan fenotipik. Data genotipik merupakan data kumpulan informasi dari gen (informasi genetik) sedangkan seluruh informasi yang dapat diamati disebut phenome (Informasi fenotipik) (Priest & Goodfellow, 2000).
Taksonomi numerik atau fenetik adalah nama yang diberikan untuk bermacam-macam metode yang menyusun suatu unit taksonomi berdasarkan karakteristik yang dimiliki. Prinsip dasar dalam taksonomi numerik adalah taksonomi yang menggunakan sebanyak-banyaknya karakter biologis suatu organisme yang disebut Operational Taxonomic Units (OTU). Semakin banyak informasi (karakter) yang ada maka akan dihasilkan pengelompokkan yang bersifat teliti, reprodusibel serta padat informasi. Secara umum klasifikasi numerik mengunakan 50—200 karakteristik biokimia, morfologi, dan karakter koloni yang digunakan untuk menentukan derajat kesamaan di antara beberapa organisme. Dalam kajian numerik, peneliti sering mengkalkulasi koefisien persentase persamaan di antara strain ataupun spesies. Dendogram atau matriks persamaan memperlihatkan hubungan individual antar strain dan dibuat berdasarkan koefisien persentase persamaan di antara mereka (Priest & Goodfellow, 2000).
Di dalam klasifikasi numerik masih adanya kontrovesi antara taksonom jaman dulu dan sekarang, dan menurut taksonom jaman dulu di dalam klasifikasi suatu sifat dianggap memiliki nilai sifat yang lebih dibandingkan sifat lainnya sedangkan di dalam klasifikasi numerik seluruh sifat dianggap memiliki nilai yang sama tanpa adanya perbedaan. Ahli taksonomi yang menggunakan klasifikasi secara taksonomi numerik menyatakan bahwa suatu sifat dapat memiliki nilai lebih setelah dilakukan klasifikasi untuk tujuan identifikasi (Priest & Austin, 1986). Di dalam taksonomi numerik, analisis bakteri dapat melewati tahap-tahap seperti yang disajikan dalam Tabel 1.
Tabel 1. Tahap-Tahap klasifikasi numerik pada analisis bakteri ( Priest & Austin, 1986)
No Tahapan
1 PEMILIHAN STRAIN
Memasukkan Pengulangan
Memasukan kultur untuk referensi
2 TES SELEKSI
Melakukan bermacam-macam tes
Minimal berjumlah 50
Menggunakan metode rapid jika dimungkinkan
3 MENYIMPAN HASIL
Analisis data error dan penolakan data yang tidak bisa diulang
4 DATA CODING
5 ANALISIS KOMPUTER
Perhitungan kemiripan
Analisis Kluster
6 INTEPRETASI HASIL
Pengklusteran
Identifikasi
Pemilihan perwakilan strain untuk studi lebih lanjut
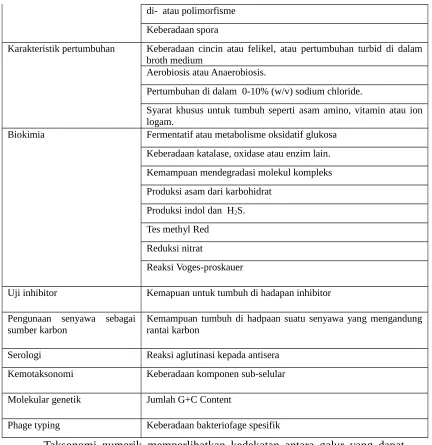
Di dalam taksonomi numerik banyak sekali karakter-karakter yang dapat digunakan untuk melakukan klasifikasi secara numerik, sebagai contoh karakter yang digunakan dalam analisis bakteri secara taksonomi numerik disajikan di dalam Tabel 2.
Tabel 2. Analisis bakteri dengan taksonomi numerik (Priest & Austin, 1986).
Kategori Analisis
Morfologi Koloni Keberadaan non-difusi atau difusi, fluorosens, non-fluorosens atau pigmen bercahaya
Ukuran dan bentuk koloni (Koloni yang menyebar) Mikromorfologi Pewarnaan Gram
keberadaan cocci, miselia, baccili Ukuran sel
Struktur tambahan
Keberadaan intraselular granular
di- atau polimorfisme Keberadaan spora
Karakteristik pertumbuhan Keberadaan cincin atau felikel, atau pertumbuhan turbid di dalam broth medium
Aerobiosis atau Anaerobiosis.
Pertumbuhan di dalam 0-10% (w/v) sodium chloride.
Syarat khusus untuk tumbuh seperti asam amino, vitamin atau ion logam.
Biokimia Fermentatif atau metabolisme oksidatif glukosa Keberadaan katalase, oxidase atau enzim lain.
Kemampuan mendegradasi molekul kompleks Produksi asam dari karbohidrat
Produksi indol dan H2S.
Tes methyl Red Reduksi nitrat
Reaksi Voges-proskauer
Uji inhibitor Kemapuan untuk tumbuh di hadapan inhibitor
Pengunaan senyawa sebagai
sumber karbon Kemampuan tumbuh di hadpaan suatu senyawa yang mengandungrantai karbon
Serologi Reaksi aglutinasi kepada antisera
Kemotaksonomi Keberadaan komponen sub-selular
Molekular genetik Jumlah G+C Content
Phage typing Keberadaan bakteriofage spesifik
Gambar 2. Dendogram dengan menggunakan komputer (Priest & Austin, 1986).
Klasifikasi ini juga menghasilkan suatu produk yang biasa disebut pohon filogenetik. Pohon Filogenetik sering digunakan oleh peneliti, khususnya peneliti yang memfokuskan penelitiannya kepada sistematika karena pohon ini mampu membantu melihat kedekatan antara suatu spesies dengan lainnya lebih mudah. Ukuran dari pohon filogenetik ini bermacam-macam tergantung dari data yang digunakan untuk mengukur kedekatan antara spesies, seperti data molekular 16S/18S rRNA yang melihat kedekatan antara spesies berdasarkan data rRNA yang dimiliki setiap spesies (Lecointre & Guyader, 2006).
Gambar 3. Pohon Filogenetik berdasarkan 16S/18S rRNA (Lecointre & Guyader, 2006)
dengan memasukannya kedalam monophyletic groups atau kelompok yang terdiri dari nenek moyang dan seluruh keturunannya (Lecointre & Guyader, 2006).
2. Kemotaksonomi
Kemotaksonomi merujuk kepada metode pengklasifikasian berdasarkan perbedaan dan persamaan data kimiawi (Penyusun dinding sel, lipid, protein seluruh sel dan lainnya) (Elke, 2010). Telah banyak penelitian jaman sekarang yang mengklasifikasikan bakteri berdasarkan metode kemotaksonomis tetapi kebanyakan dari metode ini memerlukan aparatus dan kemampuan spesial agar dapat sukses dilaksanakan (Dass et al., 1976). Berkaitan dengan hal tersebut pengunaan secara rutin metode kemotaksonomi hanya dapat dilakukan di laboratorium khusus sehingga tidak dapat dilakukan di laboratorium yang tidak memiliki peralatan khusus untuk melakukan metode kemotaksonomi (Hill & Kirsop, 1991). Akan tetapi karena pentingnya pendekatan ini untuk memahami secara jelas hubungan antar spesies, koleksi kultur dapat berada di dalam laboratorium tersebut, terutama jika suatu penelitian terkait dengan salah satu aspek taksonomi (Hill & Kirsop, 1991).
bakteri. Sebagai contoh, bakteri asam laktat hanya memiliki sitokrom b dan pada Bakteri Gram-Positif Anaerob fakultatif sitokrom c secara umum tidak ada. Selain itu genus Clostridium tidak memiliki sitokrom dan sitokrom d juga karakteristik pada banyak Bakteri Gram-Negatif. (Hill & Kirsop, 1991).
Beberapa contoh metode kemotaksonomi yang bisa diaplikasikan kepada bakteri dapat bermacam-macam dari Pyrolysis gas-liquid chromatography dari seluruh sel, hingga ekstraksi dan perbandingan dari fraksi subselular (seperti asam lemak, quinon, komponen dinding sel, protein) dari asam nukleid, bagian dari asam nukleid hingga gen. Selain itu metode kemotaksonomi memberikan data berupa data kualitatif dan kuantitatif dan juga beberapa data komponen kimia yang dapat diuji ke dalam satu metode atau separasi molekular lainnya dan disimpan sebagai profil kimia kuantitatif atau ‘Fingerprint’ (Hill & Kirsop, 1991). Labeda (1985) memberikan contoh klasifikasi melalui metode kemotaksonomi yang disajikan pada Tabel 3.
Elytrosporangium Sama seperti
Geodermatophilus Hifa terbagi ke segala arah, membentuk konidia
Microtetraspora 4 – 6 rantai konidia pada miselium aerial
Planobispora Sporangia silindris, setiap sporangia terdapat 2 spora
isomer LL-DAP dan meso-DAP
dihasilkan pada miselium aerial
Pengunaan metode kemotaksonomi di dalam penelitian, terutama penelitian terkait dengan klasifikasi mikroorganisme mampu memberikan daa kimiawi yang mendukung identifikasi keanekaragaman suatu mikroorganisme (Wilkins & Lay, 2006).
DAP (Diamino Pimelic Acid) adalah salah satu karakter yang diperhatikan dalam penelitian menggunakan metode kemotaksonomi dikarenakan DAP mampu memberikan ciri khas tersendiri pada aktinomisetes diantaranya DAP berupa meso-DAP, LL-DAP atau DD-DAP (Clarkson & Meadow, 1971).
Gula dari sel merupakan ciri khas penting juga di dalam penelitian kemotaksonomi. Gula ini memberikan ciri khas tersendiri bagi aktinomisetes saehingga dibagi menjadi 5 tipe dinding sel tergantung dari gula sel yang dimiliki yaitu tipe I, II, III, IV dan X. Dapat dilihat pada tabel 3 juga perbedaan untuk setiap tipe dinding sel tergantung dari gula sel yang dimiliki. Secara umum, hasil dari analisis gula sel adalah gula berupa galactose, arabinose, xylose, rhamnose, mannose, glucose dan madurose (Labeda, 1985).
Menaquinone atau Vitamin K2 merupakan hasil sintesis biokimia yang dapat
menjadi penciri dari aktinomisetes. Secara umum, hasil menaquinon yang dapat dihasilkan berupa MK-4 hingga MK-9 dengan hidrogenasi sebanayak 4 buah (H4)
(Shearer, 2008).
Asam lemak adalah penyusun dinding sel bakteri. Setiap bakteri memiliki ciri khas tersendiri yang mampu menjadi indikator identifikasi spesies suatu bakteri. Secara umum, panjang rantai dari aktinomisetes berkisar antara C12 hingga C18 (Bolsover, 2004).
adalah bakteri yang memiliki fosfolipid berupa phosphatidyl ethanolamine. Tipe III adalah bakteri yang memiliki fosfolipid berupa phosphatidyl choline. Tipe IV adalah bakteri yang memiliki fosfolipid berupa glucosamine. Tipe V adalah bakteri yang memiliki fosfolipid berupa phosphatidyl glycerol. Selain itu terdapat juga fosfolipid dalam bentuk lain yang menjadi penciri khas yaitu phosphatidyl inositol, phosphatidyl inositol mannosides, phosphatidyl methylethanolamine, acyl phosphatidyl glycerol, diphosphatidyl glycerol dan ninhydrin-positive glycophospholipid (Lechelavier et al., 1977).
3. Klasifikasi Molekular
Teknik molekular pada keilmuan biologi telah banyak membantu di dalam membantu mengetahui kedekatan genetik antara anggota yang memiliki kategori taksonomi yang berbeda. Berbeda dengan pendekatan melalui metode lain, metode molekular menekankan kepada karakter molekular seperti sekuens basa DNA. G+C Content dan hibridisasi asam nukleat. Metode ini semakin diminati seiring dengan berkembangnya jaman dikarenakn waktu yang diperlukan untuk menganalisis tidak lama dan hasil yang didapatkan mampu memberikan hasil yang maksimal (Singh, 2012).
Di dalam klasifikasi molekular seseorang mampu menganalisis kedua DNA dan RNA melalui berbagai metode. Sebagai contoh, berbagai metode yang dapat dilakukan di dalam klasifikasi molekular disajikan pada Tabel 4.
Tabel 4. Analisis melalui metode molekular (Singh, 2012)
Metode Penjelasan Metode
Sekuens DNA Segmen DNA dari kedua organisme dilakukan sekuensing dari ujung satu ke ujung lainnya. Sekuensing dari keduanya menunjukkan kemiripan atau ketidak-miripan.
Restriction mapping Segmen DNA dari spesies yang berbeda dilakukan restriction mapping. Spesies yang berdekatan akan memiliki restriction map yang dekat.
Pemitaan Kromosom Kromosom dari spesies yang berbeda dilihat melalui mikroskop.
Sekuens Asam Amino Asam amino dari kedua organisme yang berbeda dilakukan sekuens. Spesies yang berdekatan akan memberikan protein yang hampir sama juga.
Salah satu metode yang sering digunakan didalam penelitian taksonomi molekular mikrobia adalah sekuens 16S rRNA. Teknik ini secara luas digunakan sebagai biomarker ataupun studi ekologi mikrobia. Ribosom merupakan mesin untuk mentransalasikan gen yang ada pada seluruh makhluk hidup. Gen pada ribosom RNA hanya mengalami perubahan sedikit selama jutaan tahun sejarah evolusi suatu organisme. Perubahan sedikit ini memberikan petunjuk seberapa dekatkah suatu spesies dengan spesies lain (Anonim, 2014).
4. Aktinomisetes
berasosiasi dengan tanaman sehat (Endofit) dan hewan termasuk organisme laut sehingga bakteri ini secara umum bersifat saprofit (Maheshwari et al., 2010).
Anggota dari aktinomisetes membentuk substrat dan/atau miselium serta spora aseksual. Anggota dari aktinomisetes ini memiliki pembentukan morofologi yang beragam seperti pada Genus Streptomyces, Actinoplanes, Nocardia, Actinomadura dan Dactylosporangium membentuk miselia yang berbeda pada kulutur permukaan dan tenggelam. Selain itu konidia juga muncul dari hifa yang bercabang di mana cabang tersebut membentuk cabang yang morfologinya beragam seperti membentuk filamentus ataukah miselia agregat padat (Shapiro, 1989). Di tahap lebih lanjut, miselia ini dapat membentuk fragmen berbentuk partikel-partikel kecil seperti pellet tetapi morofologi aktinomisetes ini dapat beragam tergantung dari medium dan faktor fisik. Untuk sporogenesis juga beragam, tergantung dari strain dan lingkungan. Begitu juga dengan bentuk spora, pigmen, dan germinasinya dapat bervariasi (Shapiro, 1989).
Gambar 4. Siklus hidup strain anggota genus Streptomyces (Shapiro, 1989).
Aktinomisetes telah diketahui sebagai sumber potensial yang dapat menghasilkan senyawa bioaktif dan kaya akan sumber metabolit sekunder. Aktinomisetes telah diketahui menghasilkan sekitar dua pertiga substansi bioaktif yang ada selain itu aktinomisetes merupakan penghasil enzim yang dilirik di industri bioteknologi karena kemampuannya (Maheshwari et al., 2010). Terdapat 5 kelompok besar bioproduk yang dihasilkan oleh aktinomisetes yaitu metabolit primer, metabolit sekunde,. produk biokonversi, sel Aktinomisetes sebagai produk dan produk rekombinan (Goodfellow et al., 1988;Maheshwari et al., 2010).
Gambar 5. Proses bioprospek aktinomisetes (Maheshwari et al., 2010).
Bioprospek dilakukan untuk memahami lebih dalam mengenai keankearagaman biodiversitas suatu isolasi seperti genetikanya ataupun produk yang dihasilkan sehingga dapat menguntungkan manusia (Acharya & Chaudary, 2012).
Tabel 5. Produk metabolit primer Aktinomisetes (Maheshwari et al., 2010).
Primary metabolites Producing actinomycetes
Enzymes
Lipas Streptomyces levendulae
Amylase Streptomyces sp., Thermonospora sp.
Protease S. hygroscopicus
Vitamin B12 S. olivaceus, Micromonospora sp. &Nocardia gardnen
Selain metabolit primer, aktinomisetes juga menghasilkan metabolit sekunder yang tidak kalah pentingnya. Metabolit sekunder ini bervariasi sekali, terutama pada struktur kimiawinya. Beberapa produk metabolit sekunder yang penting seperti antibiotik, agen antikanker, antivirus, antiparasit, insektisida, promotor pertumbuhan dan lainnya. Walaupun antibiotik ini adalah metabolit sekunder yang sekarang dianggap paling penting, tetapi masih banyak produk metabolit lain yang memiliki aktivitas biologis yang besar. Karakteristik yang khas dari metabolit sekunder adalah metabolit yang tidak dihasilkan pada fase eksponensial tetapi pada fase stasione oleh karena itu. produksi metabolit sekunder ini dimulai sewaktu pertumbuhan mulai dibatasi oleh salah satu sumber nutrien seperti karbon, nitrogen/fosfat (Maheshwari et al., 2010). Diperkirakan hasil dari metabolit sekunder yang dihasilkan oleh mikroorganisme berjumlah sekitar 23.000. 45% diantaranya atau sekitar 10.000 dihasilkan oleh aktinomisetes, ada sekitar 7.600 dihasilkan oleh anggota genus Streptomyces. Berkaitan dengan hal tersebut, produk metabolit sekunder adalah produk yang banyak diminati oleh
para peneliti di dunia untuk mempelajarinya dikarenakan fungsinya (Sharma, 2014). Beberapa produk dari metabolit sekunder yang dihasilkan oleh aktinomisetes disajikan di dalam Tabel 6.
Aktinomisetes memiliki satu kelompok genom berbentuk sirkular besar dan memiliki beberapa variasi dari elemen ekstrakromosomal. Ukuran genom pada strain anggota genus Streptomyces diperkirakan 1.5 – 2 kali ukuran dari Escherichia coli. Strain anggota genus Streptomyces ini memiliki DNA dengan GC content yang tinggi yaitu sekitar 70-74% tetapi DNA dari strain anggota genus Mycobacterium memiliki G+C Content lebih rendah.Selain itu sekuens DNA yang berulang muncul secara merata ada pada strain anggot genus Streptomyces .Berkaitan dengan hal tersebut aktinomisetes sering membawa extrakromosomal DNA (Plasmid) dalam ukuran dan jumlah yang bervariasi. Fungsi plasmid yang telah diketahui yaitu untuk fertilitas, transfer gen, pengaturan ulang genom dan produksi antibiotik. Selain itu aktinomisetes umumnya memiliki plasmid yang kaya akan GC, plasmidnya berukuran sekitar sekitar 10-40 kb ukurannya dan jumlahnya kurang dari 30. Plasmid Aktinomisetes dapat berbentuk sirkular ataupun linear (Goodfellow et al., 1988). Ukuran Genom dari Streptomyces disajikan di dalam Tabel 7.
Tabel 6. Produk metabolit sekunder dari Aktinomisetes (Maheshwari et al., 2010).
Biological activity Secondary metabolite
Producer
Anibacterial Streptomycin Streptomyces griseus
Chloramphenicol Streptomyces venezulae Kanamycin Streptomyces kanamyceticus Tetracyline Streptomyces rimosus
Abyssomycin Verrucosispora sp.
Antifungal Amphotericin B Streptomyces nodosus
Candicidin Streptomyces griseus Nystatin Streptomyces noursei
Antivirasls Fattiviracin Streptomyces sp.
Insecticide and antiparasitic Avermectin Streptomyces avermetilis Milbemycin Streptomyces hygroscopicus
Tabel 7. Ukuran DNA genom strain anggota genus Streptomyces (Maheshwari et al., 2010).
Organisme Genome size (md x 109)
Streptomyces coelicolor A3(2) 7.09 + 0.73 (mycelium) 7.23+0.51 (spores) Streptomyces rimosus ATCC 10970 6.77 + 0.26 (mycelium)
6.33 + 0.30 (spores)
(1 megadelton (md) berkoresponden dengan 1500 pasang basa (base pair (bp)).
Selain itu dengan perkembangan jaman serta kebutuhan dari produk yang dihasilkan dari Aktinomisetes terutama antibiotik, telah banyak dilakukan penelitian mengenai genetika dari aktinomisetes. Banyak dari penelitian yang dilakukan mencoba untuk menghasilkan antibiotik yang dengan memanipulasi struktur gen dari aktinomisetes (Maheshwari et al., 2010). Sebagai contoh berikut adalah lokasi dari biosintetik antibiotik pada kromosm dan ekstrakromosm aktinomisetes yang disajikan pada Tabel 8.
Tabel 8. Struktur gen biosintetetik antibiotik pada kromosom dan ekstrakromosom aktinomisetes (Maheshwari et al., 2010).
Antibiotics Organisme
Di alam, aktinomisetes dapat berperan merugikan ataupun menguntungkan, di antara peran merugikan yang dimiliki adalah sifat patogenik yang dimiliki aktinomisetes sehingga dapat menyebabkan penyakit kepada hewan, manusia dan tumbuhan selain itu spora dari aktinomisetes dapat menyebabkan polusi lingkungan baik di pemukiman, di peternakan ataupun di industri (Goodfellow et al., 1988) Sebagai hasilnya, penyakit seperti “Farmers lung” dan hipersensitivitas pneumonia dapat terjadi. Aktinomisetes juga dapat menyebabkan polusi air dan menyebabkan masalah pada pembuangan air seperti menghalangi ataupun pembentukan gelembung yang menggangu aliran air (Goodfellow et al., 1988).
Diantara manfaat aktinomisetes yang menguntungkan, salah satunya adalah biodegradasi melalui biotransformasi suatu senyawa, biotransformasi ini telah mendapatkan perhatian lebih juga dibandingkan dengan metode kimia biokonversi lainnya (Maheshwari et al., 2010). Biokonversi yaitu perubahan suatu substansi (Substrat) menjadi lainnya (Produk), di sini enzim memainkan peran penting di dalam melakukan biokonversi yang dilakukan bahwa enzim menjadi katalis dari reaksi kimia sebagai contoh beberapa tipe biokonversi yang penting seperti oksidasi, reduksi, reaksi hidrolitik dan kondensasi (Maheshwari et al., 2010). Beberapa biokonversi yang dilakukan oleh Aktinomisetes beserta produknya disajikan di dalam Tabel 9.
Tabel 9. Produk biokonversi oleh Aktinomisetes (Maheshwari et al., 2010).
Aktinomisetes Substrate Product
Nocardia sp. N- dedecyl benzene Phenyl acetic acid
Streptomyces sp. Chitin Chitosan
Streptomyces aureofaciens Nitropentachlorobenzal Pentachloraniline Streptomyces aureofaciens
(ATCC10762)
Anhydrotetracyline Tetracyline
Streptomyces griseus Streptomycin Streptomycin – P Streptomyces
roseochromogenes
9 alpha fluorocartisol 9 alpha fluoro – 16 alpha hydroxycartisol
Streptomyces aureus Progesterone 15 alpha hydroxyl 4 pegene
Streptomyces sp. 2 amino 4 alkyl imidazole
3 zodione 2 nitro 4 alkyl imidazole
B. Hipotesis
Berdasarkan tinjauan pustaka yang telah dilakukan, isolat aktinomisetes Indonesia dari LIPI Cibinong dapat diindentifikasi melalui metode kemotaksonomis dan Molekular. Analisis kemotaksonomi dan molekular aktiknomisetes dari koleksi kultur LIPI Cibinong dapat memberikan data kimiawi khas dari aktinomisetes Indonesia sehingga dapat diindentifikasi hingga tingkat genus. Melalui analisis kemotaksonomi, diperkirakan aktinomisetes strain lokal Indonesia koleksi kultur LIPI Cibinong memberikan hasil analisis DAP berupa meso-DAP, LL-DAP atau DD-DAP, hasil analisis gula sel berupa galactose, arabinose, xylose, rhamnose, mannose, glucose dan madurose, hasil analisis menaquinon berupa menaquinon MK-4 hingga MK-9 (H4), hasil analisis asam
BAB III. METODE PENELITIAN
A.
Tempat dan Waktu Penelitian
Penelitian ini dilakukan di Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong, Bogor, Indonesia pada bulan Maret hingga Agustus 2015.
B. Strain Bakteri
Strain bakteri berupa aktinomisetes Indonesia yang akan digunakan didapatkan dari koleksi kultur yang dimiliki oleh LIPI Cibinong. Isolat yang digunakan berjumlah 10 isolat yang isolat tersebut sebelumnya telah diisolasi dari tanah dan diperservasi ke dalam ampul dengan metode Dry Freezing.
C. Kultur Isolat
Isolat yang berada di dalam ampul diambil dengan cara memasukkan sedikit medium cair ISP2 ke dalam ampul dengan menggunakan pipet pasteur dan diambil kembali medium cair berisi isolat tersebut menggunakan pipet pasteur. Medium cair berisi isolat selanjutnya sebagian dimasukkan ke dalam medium ISP2 broth dan sebagian disebar di atas medium padat menggunakan spreader. Setelah selesai isolat diinkubasikan di dalam inkubator dengan suhu 30oC selama
1 minggu. Isolat yang telah tumbuh tersebut selanjutnya diremajakan kembali sebanyak dua kali dengan cara menumbuhkan kembali ke medium ISP2 padat.
D. Persiapan Isolat untuk analisis Kemotaksonomi dan molekular
dalam erlenmeyer 500 ml. Setelah itu isolat diletakkan pada shaker dan diinkubasikan dalam suhu ruang selama 2 minggu. Isolat yang telah tumbuh selanjutnya disentrifugasi dan dicuci dengan aquades sebanyak 3 kali. Sel yang masih basah selanjutnya disimpan di dalam suhu -20oC untuk ekstraksi DNA dan
sel yang lain selanjutnya dilakukan lyophilization dan dihancurkan untuk mendapatkan sel kering yang selanjutnya disimpan di dalam suhu ruang dan siap untuk dilakukan analisis selanjutnya.
E. Analisis Kemotaksonomi
Beberapa metode dilakukan untuk mendapatkan data kemotaksonomi dari isolat. Metode yang dilakukan adalah sebagai berikut.
1. Analisis Asam Diamino Pimelat (Diamino Pimelic Acid Analysis)
Analisis diawali dengan cara mengambil isolat pada medium ISP2 padat menggunakan metode sumuran sebanyak 3 bulatan dan dimasukkan ke dalam cryotube 2 ml. Isolat tersebut selanjutnya ditambahkan 90µl 6N HCl. Isolat kemudian di hidrolisis dengan menggunakan autoclave selama 15 menit pada suhu 121oC. Setelah dilakukan hidrolisis. Isolat dan DAP Standar selanjutnya di
spot menggunakan mikropipet ke TLC plastic sheet (Merck 5577). Setelah selesai, TLC plastic sheet selanjutnya di masukkan kedalam TLC chamber yang berisi methanol-pyridine-10N HCl- air (80:10:2.5:17.5) sebanyak 110 ml dan dibiarkan semalam. Setelah itu TLC plastic sheet disemprot menggunakan ninhydrin di dalam acetone (0.1%, w/v) menggunakan sprayer dan dikeringanginkan dilanjutkan pemanasan dengan suhu 100-110oC didalam oven
selama 2 menit.
2. Analisis Gula dari Dinding Sel
Analisis diawali dengan cara melakukan hidrolisis dari 50 mg biomassa kering dengan menggunakan 1 ml 1N HCl di dalam tabung kaca yang disegel dengan suhu 100 oC selama 2 jam. Setelah itu didinginkan dan ditambahkan 1 ml
Ba(OH)2 dan dikeringkan. Setelah itu pH diatur hingga 5,2-5,5. Setelah selesai
eppendorf tube yang baru untuk disentrifugasi selama 5 menit dengan kecepatan 12.000 rpm. Supernatan kemudian di spot pada TLC plastic sheet yang selanjutnya dimasukkan ke dalam chamber yang berisi campuran n-butanol : Aquades : Pyridine :Toluene = 10:6:6:1 (v/v) selama 1 malam. Standar yang digunakan adalah campuran 0,1% (w/v) galactose, arabinose, xylose, rhamnose, mannose, glucose dan madurose. Setelah itu TLC plastic sheet disemprot menggunakan campuran 10 ml n-butanol, 0,325 g phtalic acid dan 0,2 ml aniline menggunakan sprayer dan dikeringanginkan dilanjutkan pemanasan dengan suhu 100-110oC didalam oven selama 4 menit.
3. Analisis Menaquinon
Analisi diawalli dengan100 mg biomassa kering dimasukkan ke dalam larutan 10 ml chloroform:methanol (2:1) dan dicampur selama 1 malam. Setelah itu larutan diuapkan dan hasil ekstrasi dilarutkan dengan 1,5 ml acetone dan diuapkan kembali. Setelah itu hasil ekstraksi dispot ke TLC plate silica gel Kielselgel F254
didalam chamber berisi toluene selama 30-40 menit. Quinon ditandai menggunai lampu UV. Silica gel dikorek dan dimasukkan kedalam tube, dilarutkan menggunakan 1,5 ml acetone dan disentrifugasi selama 5 menit dengan kecepatan 3000 rpm. Setelah diuapkan lalu dilakukan analisis LC-MS.
4. Analisis Asam Lemak
Analisis diawali dengan 50 mg biomasssa kering dimasukkan kedalam tube dengan tutup yang ditambahkan 1 ml saponification reagent (15g NaOH, 50 mg Methanol dan 50 ml Aquades). Di-vortex selama 5 detik kemudian dipanaskan di water bath selama 5 menit dengan suhu 100 oC. Didinginkan lalu dilihat apakah
dengan kecepatan 2000 rpm. Bagian atas lalu dipindahkan untuk dilakukan analisis GC-MS.
5.
Analisis FosfolipidAnalisis diawali dengan 100 mg biomassa kering dicampur dengan 2,3 ml campuran chloroform:methanol:aquades (9:10:3) kemudian dicampur selama 1 jam dan disentrifugasi selama 10 menit degan kecepatan 3000 rpm. Supernatan kemudian dipindahkan ke tube baru dan ditambahkan 0,7 ml campuran chloroform : ethanol 0,3 % NaCl (5:1:4), dicampur selama 4 jam dan disentrifugasi selama 5 menit dengan kecepatan 3000 rpm. Perlakuan ini diulang sebanyak 3 kali. Supernatan yang didapatkan kemudian ditambahkan 1,75 ml chloroform dan 1,75ml 0.3% NaCl. Larutan kemudian dicampur selama 10 menit dan disentrifugasi selama 5 menit dengan kecepatan 5000 rpm. Bagian atas dihilangkan, dan bagian bawah dikeringkan. Residu kemudian dicampur dengan 50 µl etanol dan di spot pada TLC plastic sheet beserta standar dan dikeringkan.
F. Analisis Molekular
Beberapa metode dilakukan untuk mendapatkan data molekular dari isolat. Metode yang dilakukan adalah sebagai berikut.
1. Isolasi Genom DNA
Ekstrasi DNA diawali dengan memasukkan 1,5 ml 1xTE Buffer ke dalam isolat basah dan disentrifugasi. Perlakuan ini diulangi sebanyak 4 kali untuk mencuci sel. Setelah dicuci, pellet ditambahkan 500 µl 5 mM EDTA (pH 8.0) dan 50 µl lysozyme. Isolat kemudian di inkubasi di dalam water bath selama 1 jam pada suhu 37 oC. Setelah diinkubasi, isolat ditambahkan 50 µl 20% SDS (Sodium
Dodecyl Sulfate) dan 50 µl proteinase K solution yang selanjutnya diinkubasi di water bath selama 1 jam pada suhu 55 oC. Selama inkubasi, setiap 5 menit isolat
eppendof tube yang baru. Perlakuan ini diulangi sebanyak 3 kali. Supernatan yang telah didapatkan kemudian ditambahkan 3M Na acetate 1/10 volume dari supernatan dan 2-propanol 2 kali volume dari supernatan. Dikocok, DNA akan terlihat berwarna putih seperti benang-benang lendir. Eppendof tube berisi DNA kemudian disentrifugasi selama 10 menit dengan kecepatan 13.000 rpm. Supernatan kemudian dibuang, dan pellet ditambahkan 70% ethanol untuk membersihkan sisa-sisa larutan. Setelah itu DNA dikerinkan dengan cara membiarkan diuapkan pada temperatur ruang. DNA yang telah kering kemudian ditambahkan 500 µl 1x TE Buffer dan dilarutkan. Setelah itu ditambahkan 50 µl Rnase A solution dan 0.5 µl Rnase T1 solution kemudian diinkubasi di dalam water bath selama 30 menit pada suhu 37 oC. Larutan kemudian ditambahkan 50
µl Proteinase K dan di inkubasi selama 1 jam pada suhu 37 oC. Setelah selesai
kemudian dilakukan lagi perlakuan dengan PCI sebanyak 3 kali. Supernatan kembali ditambahkan 3M Na acetate 1/10 volume dari supernatan dan 2-propanol 2 kali volume dari supernatan. Eppendof tube berisi DNA kemudian di sentrifugasi selama 10 menit dengan kecepatan 13.000 rpm. Supernatan kemudian dibuang, dan pellet ditambahkan 70% ethanol untuk membersihkan sisa-sisa larutan. DNA kering kemudian disimpan di dalam kulkas suhu 4 oC.
2.
Determinasi G+C ContentDNA kering dilarutkan di dalam 25 µl aquades steril, direbus selama 10 menit dan langsung didinginkan di dalam es. Setelah itu ditambahkan 10 µl nuclease P1 solution dan diinkubasi di dalam water bath selama 1 jam pada suhu 50 oC.
Setelah itu ditambahkan 10 µl BAP solution dan di inkubasi di dalam water bath selama 24 jam pada suhu 37 oC. Setelah itu ditambahkan 10 µl BAP solution dan
25 µl nuclease P1 solution, diinkubasi selama 24 jam pada suhu 37 oC. Sampel
Tabel 10. Kondisi HPLC untuk Derminasi G+C Content
Tahapan Kondisi
Kolom ODS C-18 (4.6 x 150 mm, Cosmosil
5C18-MS-II, 38019-81, Nakalai tesque, Tokyo, Japan.
Temperatur 35 oC
Fase Mobile Campuran 0,2 M NH4H2PO4-CH3CN
(40 :1 )
Flow Rate 1,0 ml/min
Deteksi UV Detector (270 nm)
Larutan Pencuci 20% CH3CN
3. Isolasi Gen 16S rRNA
Isolasi diawali dengan cara melisiskan sel bakteri secara fisik. Isolat diambil dengan metode sumuran sebanyak 5 koloni dan dimasukkan kedalam tabung reaksi berisi bead. Koloni kemudian di fragmentasi selama 10 menit. Hasil lisis kemudian dimasukkan kedalam mikrotube untuk selanjutnya disentrifugasi dengan kecepatan 15.000 rpm selama 5-10 menit. Setelah selesai, supernatan dibuang. Pellet yang tersisa kemudian dicuci dengan 1xTE solution sebanyak 3 kali dan ditambahkan TE solution hingga berjumlah 150 µl. Selanjutnya ditambahkan 20 µl lysozyme dan dikocok. Isolat kemudian diinkubasi pada water bath selama 1 jam dengan suhu 37 oC. Setelah selesai kemudian ditambahkan 50
µl 20% SDS dan 50 µl Proteinase K. Isolat kemudian di inkubasi pada suhu 55 oC.
diletakkan pada magnetic stand dan dipindahkan ke tube baru. Selanjutnya dilakukan amplifikasi gen 16S rRNA menggunakan PCR. Primer yang digunakan adalah sebagai berikut (Brosius et al., 1981).
Tabel 11. Primer yang digunakan untuk analisis sequencing gen 16S rRNA (Brosius et al., 1981)
Primer Basa
9F 5’-GAGTTTGATCCTGGCTCAG
1541R 5’-AAGGAGGTGATCCAGCC
PCR dilakukan menggunakan kit TaKaRa Ex Taq. Larutan PCR yang digunakan adalah campuran dari Template DNA 1 µl 10 ng/ µl, 10xreaction buffer 5 µl, dNTP mix 4 µl, primer 9F 1 µl, Primer 1541R 1 µl, Taq Polymerase 0,25 µl dan aquades steril 37,75 µl. Kondisi PCR yang digunakan disajikan pada Tabel 12.
Tabel 12. Kondisi PCR untuk mendapatkan produk PCR
Tahapan Kondisi
3 menit, 95 oC
Pre-denaturation
Denaturation 30 detik, 95 oC
Annealing 15 detik, 55 oC 30 siklus
Elongation 1 menit, 72 oC
Final Extension 5 menit, 72 oC
yang telah dimurnikan selanjutnya dilakukan sekuensing gen 16S rRNA menggunakan BigDye terminator V3. 1 Cycle Sequencing Ready Reaction Kit (Applied Biosystem) dengan 2 primer yaitu forward primer adalah 9F serta reverse primer 1541R. Larutan yang digunakan untuk sequencing PCR merupakan campuran Template DNA 1 µl 10 ng/µl, BigDYE Terminator 2 µl, 5x Premix Buffer 1 µl, Setiap primer 1 µl dan aquades steril 5 µl. Kondisi yang digunakan disajikan pada Tabel 13.
Tabel 13. Kondisi PCR untuk Sequencing gen 16S rRNA
Tahapan Kondisi
Pre-denaturation 1.5 menit, 96 oC
Denaturation 10 detik, 96 oC
Annealing 5 detik, 50 oC 25 Siklus
Elongation 4 menit, 60 oC
Final Extension 5 menit, 72 oC
4. Pembuatan Pohon Filogenetik
Pembuatan pohon filogenetik 10 isolat yang digunakan dalam penelitian kali ini diawalai dengan mendapatkan sequence 16S rRNA yang sebelumnya telah dilakukan deteksi oleh peneliti dari LIPI sebelumnya. Hasil sequencing ini selanjutnya dilakukan pencarian spesies menggunakan website BLAST (Basic Local Assignment Search Tool). Hasil yang didapatkan paling mendekati selanjutnya dicatat. Untuk mendapatkan hasil spesies yang paling mendekati, selanjutnya dilakukan perbandingan dengan menggunakan pohon filogenetik. Pembanding untuk spesies yang didapatkan dari hasil BLAST digunakan seluruh data 16S rRNA spesies yang terdaftar di dalam satu genus yang berada di website LPSN (List of Prokaryotic names with standing in Nomenclature). Seluruh data yang telah didapatkan kemudian dianalisis menggunakan program MEGA6 yang berfungsi untuk membantu pembuatan pohon filogenetik. Data sequence 16S rRNA dari isolat dan LPSN kemudian di align menggunakan ClustalW yang berada di dalam MEGA6. Setelah di align kemudian dilakukan pembuatan pohon filogenetik dengan menggunakan parameter Kimura-2 method dan bootstrap sebanyak 1000 kali.
Data kimiawi yang telah didapatkan dari hasil analisis kemotaksonomi setelah itu menjadi acuan untuk mengindentifikasi isolat. Acuan yang digunakan dapat menggunakan buku identifikasi seperti Bergey’s Manual of Derminative Bacteriology ataupun acuan lainnya.
H. Analisis Data
BAB IV. HASIL DAN PEMBAHASAN
Penelitian mengenai analisis profil genetik dan kemotaksonmi aktinomisetes yang dilakukan di LIPI Cibinong dari bulan Maret hingga Agustus 2015 ini menggunakan 10 isolat yang sebelumnya telah diisolasi dan diperservasi menggunakan metode Freeze Drying. Isolat yang digunakan disajikan pada Tabel 14.
Tabel 14. Isolat yang digunakan di dalam penelitian
Timor dan Lombok. 10 ampul berisi isolat yang telah diperservasi kemudian dibuka dan ditumbuhkanpadai medium ISP 2 cair kemudian ke padat pada cawan petri menggunakan metode pour plate. Isolat di inkubasi di dalam inkubator selama 1 minggu dengan suhu 37 oC. Setelah isolat di dalam petri tumbuh,
dilakukan purifikasi sebanayak 2 kali agar didapatkan isolat yang murni. Hasil isolat yang telah dipurifikasi disajikan pada Gambar 6.
1. 2. 3.
4. .
Gambar 6. Isolat pada Medium ISP2 Padat. 1. Isolat ID04-1019, 2. Isolat ID04-1052, 3. Isolat 1066, 4. Isolat 1067, 5. Isolat 1071, 6. Isolat ID04-1088, 7. Isolat ID04-1093, 8. ID04-1103, 9. ID05-0653, 10. ID-06-0464.
Pada analisis DAP, digunakan metode Thin-Layer Chromatoghraphy atau TLC bahwa secara plastic sheet sebagai fase stationer nya dan solvent sebagai fase gerak nya. Hasil DAP yang terlihat akan berwarna ungu ataupun coklat gelap dan jika dibandingkan dengan standar akan terlihat isomer apakah dari DAP yang dianalisis. Hasil dari analisis DAP disajikan pada Gambar 7.
7. 8. 9.
Gambar 7. Hasil TLC untuk analisis DAP isolat pada TLC plastic sheet
bergerak ke atas pada TLC plastic sheet. Hasil analsis DAP pada isolat disajikan pada Tabel 15.
Tabel 15. Analsis DAP menggunakan TLC.
No. Kode Isolat Hasil analisis DAP
1 ID04-1019 meso-DAP gram-negatif bahwa biasanya asam amino ini melalui ikatan peptida terikat pada ikatan NAM-NAG dinding sel serta di beberapa bakteri gram-positif bahwa DAP ditemukan pada mucopeptida atau peptidoglikan bakteri tersebut. Terdapat 3 macam isomer dari DAP yaitu meso-DAP, LL-DAP dan DD-DAP. Pada bakteri gram-positif seperti aktinomisestes ataupun gram-negatif umum dijumpai DAP dalam bentuk meso-DAP tetapi LL-DAP dan DD-DAP jarang dijumpai pada bakteri gram-negatif. Pada aktinomisetes memang umum dijumpai meso-DAP pada dinding sel bakteri aktinomisetes tersebut sehingga analisis DAP ini bisa menjadi salah satu penunjuk untuk identifikasi akan tetapi masih perlu didukung data-data lainnya seperti data kemotaksonomi lainnya ataupun data pendukung seperti profil genetik yang mampu memberikan hasil identifikasi juga sehingga hasil identifikasi akan lebih akurat walaupun data yang ada masih memiliki kekurangan seperti belum seluruh data mengenai kemotaksonomi isolat yang dilakukan.
untuk melakukan penelitian, keterbatasan waktu dan diharuskannya berpindah antara laboratorium. Analisis selanjutnya yang dilakukan yaitu gula dari sel. Prinsip dasar yang digunakan masih sama yaitu menggunakan Thin-Layer Chromatoghraphy bahwa TLC plastic sheet merupakan fase stationer dan solvent merupakan fase geraknya. Pada analisis gula sel hasil disajikan pada Tabel 16.
Tabel 16. Hasil analisis gula sel menggunakkan TLC
No Isolat Gula Sel
1 ID05-0653 Galactose, Ribose, Mannose, Arabinose, Rhamnose 2 ID06-0464 Galactose, Ribose, Mannose, Arabinose, Rhamnose
Analisis gula dari sel memerlukan biomassa kering dari sel yang telah disiapkan sebelumnya. Prinsip pada analisis gula sel memiliki persamaan dengan analisis DAP dengan perbedaan pada analisis gula sel hanya melihat gula dari sel saja dengan menggunakan standar TLC gula yang memberikan ciri khas tersendiri bagi suatu spesies dan dari sini dapat terlihat juga tipe dinding sel yang dimiliki oleh suatu spesies. Berdasarkan Tabel 16. dapat terlihat bahwa isolat memilki gula pada selnya berupa Galactose, Ribose, Mannose, Arabinos dan Rhamnose. Deteksi gula sel ini terutama gula yang terdapat pada dinding sel setelah dilakukan hidrolisis sel sehingga dapat terlihat gula yang ada.
Analisis gula sel sering digunakan untuk mengindentifikasi bakteri, terutama aktinomisetes. Secara umum hasil analisis gula sel pada aktinomisetes dapat dihasilkan gula sel berupa Galactose, Arabinose, Xylose, Rhamnose, Mannose, Glucose dan Madurose. Gula sel ini merupakan penyusun dinding sel pada aktinomisetes. Meskipun hasil analisis gula sel menjadi penanda kemotaksonomi yang penting di dalam proses identifikasi aktinomisetes, gula sel pada aktinomisetes dapat bervariasi tergantung dari kondisi pengkulturan sehingga menjadi karakter yang kurang stabil dibandingkan karakter lain yang bisa diperhatikan
dapat mengindikasikan kemungkinan isolat yang diteliti merupakan anggota dari class Actinobacteria (Rainey & Oren, 2011).
Selanjutnya dilakukan analisis Menaquinon. Analisis Menaquinone dilakukan dengan Liquid Chromatoghraphy – Mass Spectrometry. LC-MS menggunakan kombinasi pemisahan yang digunakan pada HPLC dan analisis yang dilakukan oleh MS. Aplikasi dari LC-MS lebih berorientasi pada pemisahan substansi dan deteksi berdasarkan substansi kimia lainnya. HPLC adalah pemisahan dengan menggunakan kolom khusus sebagai fase stasioner bahwa cairan dipaksa melewati kolom tersebut sehingga terjadilah pemisahan. Untuk MS, prinsip teknik ini adalah untuk mengukur massa partikel. Dari hasil analisis yang dilakukan disajikan pada Tabel 17.
Tabel 17. Analisis menaquinon menggunakan LC-MS
No Isolat Menaquinon
1 ID05-0653 MK-9(H4) (100%)
2 ID06-0464 MK-9(H4) (100%)
Menaquinon merupakan vitamin K2 yang dapat disintesis oleh bakteri. Menaquinon dapat menjadi suatu penanda kemotaksonomi yang bernilai tinggi karena hasil sintesis menaquinon untuk setiap bakteri memberikan hasil sintesis yang unik sehingga mampu menjadi penanda kemotaksonomi. Berdasarkan hasil analisis yang dilakukan, dapat diketahui bahwa kedua isolat memiliki menaquinon MK-9(H4). MK-9 adalah senyawa kimia Vitamin K2 yang memilki rantai samping
isoprene sejumlah 9 buah dan H4 berarti 4 diantaranya mengalami hidrogenasi.
Analisis selanjutnya yang dilakukan adalah analisis asam lemak. Analisis asam lemak dilakukan dengan menggunakan Gas Chromatoghrapy – Mass Spectrometry. GC-MS menggunakan prinsip bahwa pemisahan dilakukan menggunakan alat Gas Chromatograph yang fase geraknya merupakan gas dan fase diamnya merupakan kolom. Hasil analisis asam lemak disajikan pada Tabel 18.
Tabel 18. Analisis asam lemak menggunakan GC-MS
No Isolat Asam Lemak 2 ID06-0464 16:1 ISO H 21.74% 16:0 ISO 59.98% 16:0 ISO 2H 6.25%
Sesuai dengan penamaan pada asam lemak angka didepan menunjukkan banyaknya rantai karbon pada asam lemak, angka kedua menunjukkan seberapa banyak rantai yang memiliki ikatan ganda, ISO menunjukkan adanya gugus methyl pada rantai terakhir dan H menunjukkan posisi bahwa terdapat gugus H pada rantai karbon tersebut. Dapat terlihat pada Tabel 18. terdapat perbedaan antara asam lemak yang dimiliki oleh isolat ID05-0653 dan ID06-0464. Hal ini dapat menjadi indikator pembeda dan karakteristik unik setiap spesies.
pertumbuhan adalah medium cair ISP2 sehingga hasil yang didapatkan lebih valid.
Selanjutnya analisis yang dilakukan adalah analisis fosfolipid. Analisis fosfolipid dilakukan dengan menggunakan Thin-Layer Chromatoghraphy. Hasil analisis fosfolipid disajikan pada Tabel 19.
Tabel 19. Analisis fosfolipid menggunakan TLC
No Isolat Tipe Fosfolipid Fosfolipid
PE = Phosphatidylethanolamin
PME = Phosphatidylmethylethanolamine lyso-PE = lyso- Phosphatidylethanolamin DPG = Diphosphatidylglycerol
PIM = Phosphatidylinositol mannoside NPG = Ninhydrin-positive glycophospholipid
Fosfolipid merupakan lipid yang merupakan bagian penting dari membran sel. Mereka dapat membentuk lipid bilayer karena karakterisitiknya yang merupakan amphiphilic. Struktur dari fosfolipid secara umum terdapat 1 bagian kepala gugus fosfat yang hidrofilik dan 2 bagian ekor yang hidrofobik. Fosfolipid terikat oleh ikatan gliserol. Pada fosfolipid terdapat Tipe fosfolipid P yang merupakan fosfolipid tanpa gugus nittrogenous dan tipe fosfolipid PII bahwa phosphatidylethanolamine sebagai fosfolipid yang terdeteksi. Pada kedua isolat terdapat fosfolipid yang terdeteksi seperti yang terlihat pada tabel 19 yaitu PE ( Phosphatidylethanolamin), PME (Phosphatidylmethylethanolamine), lyso-PE, DPG (Diphosphatidylglycetol), PIM (phosphatidylinositol mannoside) dan NPG (ninhydrin-positive glycophospholipid).
bakteri yang memiliki fosfolipid berupa phosphatidyl choline. Tipe IV adalah bakteri yang memiliki fosfolipid berupa glucosamine. Tipe V adalah bakteri yang memiliki fosfolipid berupa phosphatidyl glycerol. Selain itu terdapat juga fosfolipid yang bukan merupakan penanda pola fosfolipid akan tetapi mampu menjadi indikator untuk melakukan identifikasi dikarenakan secara umum spesies yang masuk di dalam suatu anggota genus tertentu akan memiliki kemiripan pada pola dan fosfolipid yang dimiliki (Bora et al., 2015).
Untuk mendukung identifikasi dari isolat, maka dilakukan profil genetik seperti pembuatan pohon filogenetik dan determinasi G+C content dari isolat. Untuk pembuatan pohon filogemetik, didasari berdasarkan kedekatan antar spesies melalui hasil sequencing gen 16S rRNA dari isolat yang sebelumnya menggunakan BLAST untuk mencari kedekatannya dan selanjutnya dibandingkan dengan spesies dalam genus yang ditemukan mendekati dari hasil BLAST menggunakan pohon filogenetik sehingga bisa terlihat jelas kedekatan antar isolat dengan spesies yang ada.
Sequencing 16S rRNA sebelumnya telah dilakukan oleh peneliti dari LIPI sehingga dalam penelitian kali ini hasil sequencing ini akan langsung digunakan untuk membantu mendukung identifikasi isolat. Hasil sequencing gen 16S rRNA dengan menggunakan bantuan BLAST yang dibandingkan dengan data daari GENE BANK dapat ditemukan genus yang paling mendekati isolat. Hasil yang didapatkan dari pencarian menggunakan BLAST disajikan pada Tabel 20.
Tabel 20. Hasil pencarian isolat menggunakan BLAST
No. Kode
Actinolpanes sp. 98% LC049065.1 1458
2
ID04-1066
Actinolpanes sp. 99% LC049066.1 1462
3
ID04-1067
4
ID04-Actinolpanes sp. 98% KC554663.1 1461
6
ID04-1093
Actinolpanes sp. 99% KM396267.1 1423
7
ID04-1103
Actinolpanes sp. 99% LC049066.1 1461
8
ID05-Dikarenakan tidak adanya data hasil sequence gen 16S rRNA pada isolat ID04-1019 maka dalam penelitian kali ini tidak dimasukkan untuk diberlakukan pencarian menggunakan BLAST. Hasil BLAST yang tidak mencapai 100% selanjutnya dianalisis lebih lanjut menggunakan pohon filogenetik dan genus yang telah didapatkan mendekati sebelumnya dari hasil BLAST selanjutnya dicari seluruh spesies yang terdaftar di dalam Euzeby : List of Procaryotes Name with Standing in Nomenclature untuk menjadi pembanding dari isolat di dalam pohon filogenetik sehingga dapat terlihat dengan jelas sedekat apakah isolat yang dianalisis dengan suatu spesies. Tujuan pembuatan pohon filogenetik ini secara umum untuk memperkirakan hubungan antar sequemce dan untuk mengetahui asal usul ataupun spesies pendahulu yang sama dari sequence. Pembuatan pohon filogenetik sekarang sudah didasari dari sequence DNA. Menurut Hall (2013) secara dasar pembuatan pohon filogenetik dibagi menjadi 4 tahap yaitu 1. Identifikasi dan mendapatkan set DNA atau sequence protein homolog, 2. Meng-align sequence, 3. Memperkirakan pohon yang akan digunakan dan 4. Mempresentasikan pohon sebaik mungkin sehingga seluruh informasi dapat diketahui oleh pembaca.
sebagai data untuk diketahui kedekatan antar isolat dan membantu identifikasi. Dipergunakannya genn 16S rRNA ini dikarenakan gen tersebut adalah gen yang tidak terlalu berubah dari jaman dulu hingga sekarang sehingga lebih mudah untuk mengetahui kedekatan antar spesies menggunakan gen 16S rRNA Pembuatan pohon filogenetik diawali dengan memasukkan sequence gen kedalam notepad agar selanjutnya dibuat dalam file FASTA bahwa program MEGA6 hanya dapat membaca file untuk sequence dalam bentuk FASTA saja. Pembandingnya yaitu seluruh spesies yang terdaftar di dalam genus Actinoplanes juga dicari gen 16S rRNA melalui Euzeby dan dimasukkan kedalam satu file FASTA yang sama dengan isolat yang ingin dicari kedekatannya. Isolat yang masih belum diketahui secara pasti dikarenakan dari hasil BLAST belum mencapai kedekatan sebesar 100% maka perlu dipergunakan pohon filogenetik untuk melihat sedekat apa ketujuh isolat lainnya dengan spesies yang telah terdaftar pada Euzeby. Program MEGA6 digunakan untuk membantu pembuatan pohon filogenetik (Gambar 8.).
Gambar 8. Interface program MEGA6
Gambar 9. Hasil alignment sequence gen 16S rRNA menggunakan ClustalW
Gambar 10. Parameter analisis untuk pembuatan pohon filogenetik pada program MEGA6
sequence dengan memformulasikanya dalam suatu matrix sehingga dapat dihasilkan suatu pohon filogenetik dengan tingkat kepercayaan tertentu. Berbeda model akan menghasilkan pohon filogenetik yang berbeda pula. Pemilihan Kimura-2 parameter di dalam penelitian kali ini didasari karena menurut Prasetya et al. (2011) bahwa pengunaan kimura-2 parameter di dalam metode pohon filogenetik neighbour-joining akan memberikan hasil yang lebih baik dibandingkan model lain, terutama jika berhadapan dengan banyak data nukleotida.