BAB I
RESEPTOR TIROSIN KINASE
RESEPTOR TIROSIN KINASE
RESEPTOR TIROSIN KINASE
RESEPTOR TIROSIN KINASE
DAN TRANSFORMING GROWTH FACTOR β
DAN TRANSFORMING GROWTH FACTOR β
DAN TRANSFORMING GROWTH FACTOR β
DAN TRANSFORMING GROWTH FACTOR β
Transforming Growth Factor β (TGF β).
Transforming growth factor beta (TGF β) adalah protein yang disekresikan untuk meregulasi proliferasi, diferensiasi dan kematian dari berbagai jenis sel. Semua jenis sel kekebalan, termasuk sel B, sel T dan sel dendritik serta makrofag, mensekresi TGF β, yang mengatur proliferasi, diferensiasi dan aktivasi oleh sitokin lain. TGF β adalah imunosuppressor utama yang berhubungan dengan autoimun, peradangan dan kanker. TGF β merupakan protein sekresi yang terdiri dari tiga isoform yakni TGF β1, TGF β2 dan TGF β3. TGF β1, merupakan anggota utama dari golongan sinyal ini yang telah banyak diketahui perannya. TGF β merupakan superfamili protein yang dikenal sebagai faktor pengatur transformasi beta superfamili, yang meliputi inhibins, aktivin, anti mullerian hormon, tulang morphogenetic protein, dan decapentaplegik. Kelompok pertama dari superfamili TGF β diidentifikasi berdasarkan kemampuan untuk mengekspresikan fenotif dan mengubah ekspresi sel sel dalam kultur. Namun faktor faktor pertumbuhan yang disekresi ini sekarang diketahui memiliki spektrum efek yang luar biasa pada pertumbuhan dan perkembangan sel normal. Faktor pertumbuhan ini juga merangsang produksi sel molekul adhesi, faktor pertumbuhan lainnya, dan molekul matriks ekstraselular.
Asam asam amino terminal berguna untuk mengatur sekresi sel pada sel target. TGF β juga memiliki terminal C yang terdiri dari 112 114 asam amino yang berperan sebagai signal pembentukan TGF β itu sendiri. TGF β yang matang membentuk protein dimer untuk menghasilkan molekul 25 kDa aktif dengan struktur yang mempunyai banyak motif. TGF β mempunyai sembilan residu sistein. Delapan residu sistein membentuk ikatan disulfida dalam molekul untuk membentuk karakteristik struktur simpul sistein dari TGF β superfamili. Sistein kesembilan dimanfaatkan untuk membentuk sebuah ikatan dengan model dimer. Banyak residu pada TGF β yang berfungsi untuk membentuk struktur sekunder melalui interaksi hidrofobik. Daerah sistein pada posisi antara kelima dan keenam merupakan daerah yang paling berbeda dari molekul TGF β yang ditampilkan pada permukaan molekul dan terlibat dalam pengikatan reseptor.
protein ini menunjukkan asal usul nenek moyang yang sama dengan banyak sekuens selama evolusi. Namun, organisasi dari subunit dalam protein dimer bervariasi di antara ketiga faktor pertumbuhan.
Ikatan silang iodinated mplekul TGF β terdapat pada permukaan sel dan telah diketahui tiga polipeptida dengan berat molekul jelas 55, 85, dan 280 kDa disebut sebagai reseptor TGF β I, II, dan III. Reseptor TGF β tipe I dan tipe II keduanya merupakan protein transmembran serin atau treonin kinase. Pengikatan TGFβ menginduksi pembentukan reseptor multimerik yang sebagian besar berupa heterotetramers, terdiri dari reseptor tipe I dan tipe II. Jenis reseptor reseptor III TGF β berupa molekul permukaan sel, yaitu β proteoglycan disebut glikan, yang muncul untuk mengatur aksesibilitas TGF β untuk sinyal transduksi reseptor TGF β dari tipe I dan tipe II heterotetramer. Fenomena ini mirip dengan pengikatan faktor pertumbuhan fibroblast (FGF) oleh proteoglikan dan presentasi FGF terikat pada reseptor.
Pembentukan TGF β.
Proses pembentukan TGFβ
Aktivasi TGF β oleh Faktor Transkripsi Smads.
reseptor yang terkait. Ini merupakan fenomena umum pada sistem sinyal intersellular. Jalur signaling TGF β memberikan satu contoh strategi yang sangat baik untuk mencapai respon yang spesifik. Sebagai contoh, pengikatan TGF β ke reseptornya menyebabkan fosforilasi Smad2, dimerization dengan Smad4, translokasi dari Smad2 atau Smad4 ke nukleus, dan mengaktivasi transkripsi gen gen target tertentu. Di sisi lain, pengikatan BMP2, anggota lain dari TGF β superfamili, pada reseptor yang dapat memfosforilasi Smad1 menyebabkan dimerisasi dengan Smad4. Pada kasus ini aktivasi dan tanggapan transkripsional tertentu dapat berbeda dari yang disebabkan oleh Smad2 dan Smad4.
Tanggapan spesifik dari reseptor TGF β dan BMP2 ditentukan oleh tiga asam amino dalam tipe I subunit reseptor dan saling melengkapi dalam R residu Smads. Spesifitas masing masing reseptor dapat diubah hanya dengan penggantian asam amino pada posisi ketiga. Demikian juga, pertukaran sekuens yang komplementer antara Smad1 dapat membalikkan spesifitas aktivasi Smad2, sehingga sekarang Smad1 diaktifkan oleh reseptor TGF β dan diaktifkan Smad2 oleh reseptor BMP2. Meskipun komplementer ini sesuai urutan reseptor spesifik dengan R Smads tertentu, wilayah lain dalam C terminal domain R Smads sangat penting untuk menentukan spesifikasi target gen induksi, kemungkinan melalui interaksi dengan faktor faktor transkripsi spesifik.
Fosforilasi dapat terjadi pada dua terminal C residu serine pada motif SXS domain aktif MH2 pada reseptor Smad. Ligan akan mengikat, Smad 2 atau 3 yang difosforilasi oleh bentuk aktif reseptor tipe I dan juga berasosiasi dengan Smad4 untuk membentuk komplek hetero oligodimer yang akan bertranslokasi menuju nukleus. Setelah melakukan translokasi komplek akan berikatan dengan skuen SNA spesifik pada daerah promoter gen yang meregulasi transkripsinya. Respon transkripsi dari TGF beta tergantung dari aktivitas Smad. Jalur signaling dari Smad akan dihambat dengan regulasi oleh inhibitor Smad6 dan Smad7. Inhibitor Smad7 akan bekerja untuk menghambat sinyal yang dimediasi oleh Smads dengan membentuk ikatan stabil dengan receptor tipe 1 dan mencegah terjadinya fosforilasi reseptor Smads serta meregulasi feedback negative. Selain itu transkripsi juga diaktivasi oleh signal TGF beta, dan Smad7 yang menginduksi terbentuknya ubiquitine dan mendegradasi reseptor melalui protein Smurf 1 atau 2. Smad 6 bersaing dengan Smad1 untuk berikatan dengan Smad4.
(LTBP), membentuk kompleks yang lebih besar yang disebut Laten Large Complex (LLC). Dalam hal ini LLC yang didapatkan disekresikan ke ECM.
simultan menginduksi perubahan konformasi LAP dan protease. Terlepas dari terlibat maupun tidaknya MMPs, mekanisme ini masih memerlukan asosiasi intergrins dan membuat jalur non proteolitik.
Peran TGF β PadaTumor.
menyebabkan hilangnya efek TGF β pada individu yang menerima perlakuan itu.
Sebuah pendekatan yang melibatkan transfeksi gen dengan decorin kecil, yang mana leusin proteoglycan mengikat dan menonaktifkan TGF β akan mengganggu homeostasis. Dalam penelitian ini, dipastikan bahwa transfeksi decorin mengakibatkan penghapusan kegiatan TFG β in vitro dan yang lebih penting, sel sel yang mengekspresikan decorin pada level yang tinggi tidak memiliki sifat sebagai sel tumor pada sistem in vivo. Namun, ada beberapa kontroversi mengenai apakah efek terapeutik dimediasi dengan decorin menghilangkan efek TGF β. Untuk menjawab hal tersebut diperlukan eksperimen lebih lanjut. Potensi penggunaan decorin dalam terapi gen mempunyai kemungkinan berpengaruh pada interaksi TGF β yang terlibat dalam hilangnya tumorgenisitas.
Fungsi TGF β Pada Apoptosis.
Sel bisa mati dalam dua cara. Pertama, melalui kematian sel terprogram (apoptosis dan autophagy), yang merupakan proses fisiologi normal dan melalui nekrosis, yang merupakan kematian dari penyebab lain, seperti kekurangan oksigen atau racun. TGF β menginduksi apoptosis dalam berbagai jenis sel. TGF β dapat menginduksi apoptosis dalam dua cara yaitu melalui SMAD pathway dan DAXX pathway. SMAD pathway adalah jalur sinyal kanonik dimana sinyal TGF β dimer mengikat ke reseptor tipe II yang merekrut tipe I phosphorylates sebuah reseptor. Jenis reseptor ini kemudian merekrut dan reseptor phosphorylates diatur oleh SMAD (R SMAD). SMAD3, R SMAD, yang terlibat dalam menginduksi apoptosis. R SMAD kemudian mengikat ke SMAD umum (coSMAD) SMAD4 dan membentuk sebuah heterodimeric kompleks. Kompleks ini kemudian memasuki inti sel di mana ia bertindak sebagai faktor transkripsi untuk berbagai gen, termasuk untuk mengaktifkan mitogen activated protein kinase 8 jalur, yang memicu apoptosis.
siklin: CDK kompleks bertanggung jawab atas Retinoblastoma protein (Rb) fosforilasi. Jadi TGF β blok melalui fase siklus G1. TGF β menekan ekspresi gen c myc yang terlibat dalam siklus sel G1. Sistem kekebalan TGF β diyakini penting dalam regulasi, sistem kekebalan Regulatory T CD25 + sel. TGF β muncul untuk memblokir aktivasi limfosit dan monosit yang diturunkan phagocytes.
Peran ganda TGF β1 dalam apoptosis.
Pemaparan sel untuk TGF beta1 dapat memicu berbagai respon selular termasuk penghambatan pertumbuhan sel, migrasi, diferensiasi dan apoptosis. TGF beta1 diatur adalah jenis sel apoptosis dan tergantung pada konteks, memang TGF beta1 memberikan sinyal baik untuk kelangsungan hidup sel atau apoptosis. Mekanisme molekuler yang mendasari peran TGF beta1 dalam apoptosis tetap tidak jelas. Protein yang terutama menengahi sinyal intraselular dari TGF beta1 adalah anggota keluarga Smad. Namun demikian, TGF beta1 signaling juga dapat bekerja sama dengan kematian reseptor apoptotic jalur (FAS, TNF), dengan modulator apoptosis intraselular JNK dan P38 MAP kinase, Akt, NF kappaB, dan dengan jalur apoptotic mitokondria dimediasi oleh anggota anggota Bcl 2 keluarga. Selain itu, keterlibatan TGF beta1 dalam produksi stres oksidatif dan mencegah proses peradangan yang diperlukan untuk pembersihan tubuh apoptotic bukti lebih lanjut integrasinya ke jalur apoptotic. Interaksi dan keseimbangan antara rangsangan yang berbeda memberikan dasar bagi pro atau anti output apoptotic TGF beta1 pensinyalan dalam suatu sel.
Peran TGF β pada Jantung.
Peran TGF β pada
TGF β signaling juga mungkin memainkan peran utama dalam patogenesis sindrom Marfan, penyakit yang ditandai dengan tidak proporsional tinggi, arachnodactily, ectopia lentis dan komplikasi hati seperti prolaps katup mitral dan pembesaran aorta meningkatkan dan ada kemungkinan terjadinya diseksi aorta. Sementara cacat mendasar dalam sindrom Marfan ialah rusaknya sintesis glikoprotein fibrillin, biasanya merupakan komponen penting dari serat elastis, fenotipe sindrom Marfan dapat dihilangkan dengan penambahan dari TGF β antagonis dalam tikus yang terkena dampak. Hal ini menunjukkan bahwa untuk sementara gejala sindrom Marfan mungkin tampak konsisten dengan gangguan jaringan ikat, mekanisme lebih mungkin berkaitan dengan pengurangan karantina dari TGF β oleh fibrillin. Prekursor TGFβ4 ditemukan dengan peningkatan susunan gen selama fase pra menstruasi dalam endometrail stroma (Kothapalli et al. 1997) dan disebut EBAF (endometrium pendarahan faktor terkait). Kemudian secara terpisah ditemukan embrio vertebrata asimetri kiri kanan dan diberi nama lefty2 (juga disebut Lefty A).
Aktivasi Reseptor TGFβ I.
Jalur aktivasi TGFβ Smad
permukaannya. Karena jenis sel yang berbeda mengandung faktor transkripsi yang berbeda, respon seluler yang diinduksi oleh TGFβ juga bervariasi di antara jenis sel. Pada sel epitel dan fibroblas misalnya, TGFβ tidak hanya menginduksi ekspresi protein matriks ekstraseluler (misalnya kolagen), tetapi juga ekspresi protein yang menghambat serum protease.
Hilangnya TG β signaling berperan atas proliferasi sel.
. Efek mutasi pada protein protein yang berperan dalam jalur TGFβ Smad.
Gangguan yang berhubungan dengan aktivasi TGF β signaling.
Efek pada FKBP12 RI.
Hambatan perubahan konformasi, disebabkan oleh adanya FKBP12 dan kestabilan orientasi. Struktural katalitik domain memungkinkan perlindungan komponen vital yang melaksanakan proses fosforilasi Smad. Seperti disebutkan, perlindungan FKBP12 loop GS menyebabkan perubahan konformasi reseptor di seluruh struktur. Pengikatan menggeser FKBP12 C alfa heliks dengan memaksa interaksi dengan GS loop, urutan aktivasi, dan alpha C heliks sendiri. Interaksi sekitar Fosfat mengikat ATP loop dan mencegah ikatanATP t. Tiga residu bertanggung jawab atas hal ini dengan mendekati lingkaran yang mengikat dan menciptakan interaksi mereka sendiri, Lys 232 blok alfa phopsphate, Phe 216 beta blok fosfat, dan Arg 372 dengan blok Phe 216 gamma fosfat . Dilihat dari model rantai yang berpadu sisi Arg 372 terlihat memperluas ke pusat katalitik RI dan membentuk pasangan ion dengan Asp 351, suatu asam amino esensial yang dibutuhkan untuk koordinasi ion Mg. Yang menghambat konformasi alfa heliks C akan distabilkan oleh sejumlah interaksi residu panjang dengan dua lembar beta struktural penting 9 dan 10. Kontak Van Der Walls dibuat antara C heliks alfa dan beta untai 10. Glu 247 residu akan menstabilkan interaksi antara Arg 244 dan Asp 366. H ikatan terjadi antara alpha helix dan beta C untai 9. Residu Asp 366 dari 10 untai kelompok beta Arg 357 ke dalam konformasi di tempat yang terdapat H ikatan dengan Thr 251. Urutan aktivasi tampaknya berefek pada kegiatan RI dalam menggeser konformasi. Hal itu sendiri tidak terfosforilasi oleh RII kinase. Seperti disebutkan, fosforilasi dari lingkaran GS mengganggu interaksi dengan C alfa heliks dan memungkinkan segmen aktivasi untuk pindah dari mengikat ATP loop
Fosforilasi Smad.
dari ATP terikat ke substrat Smad. Meskipun urutan ini bervariasi antara RI dari spesies yang berbeda, lima residu sangat dilestarikan dan muncul untuk memfasilitasi fosforilasi Smads, lys 232, glutathione 245, asp 333, asn 338, asp 351.
TGF β dan Smad.
domain, dan domain intraseluler. Domain ekstraselular bertanggung jawab untuk mengenali dan mengikat faktor pertumbuhan, sementara intraselular domain, ketika dibawa ke ruang dengan jarak yang berdekatan, bertanggung jawab untuk mengaktifkan mesin hilir yang membawa sekitar satu respon selular yang disetel tepat. Faktor pertumbuhan dari (TGF b) superfamili telah sangat beragam selama evolusi, dengan enam faktor faktor seperti nematoda, sembilan di lalat buah, dan empat puluh dua pada manusia. Mereka termasuk bone morphogenetic protein (BMP), yang memainkan peran mendasar dalam pola embrionik, yang berhubungan erat dengan faktor pertumbuhan dan diferensiasi (GDFS), yang mengatur kerangka tulang rawan dan pembangunan, activins, yang mengatur pelepasan hormon hipofisis, dan terlambatnya evolusioner, (TGF bs), yang mengatur pertumbuhan sel dan morfogenesis. BMP, GDFS, activins, dan TGF bs adalah homodimers, terdiri dari dua monomer yang diperpanjang dan dilakukan bersama di sebagian besar, tapi tidak semua kasus, dengan rantai antar ikatan disulfida.
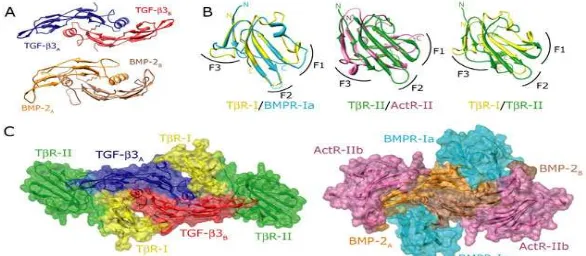
masing masing, dan ekstensif kontak satu sama lain. BMP tipe I dan tipe II reseptor berarsir cyan dan magenta masing masing, tidak kontak satu sama lain.
TGF βs dan faktor faktor yang terkait mendorong respons mereka dengan menggabungkan sebuah kompleks heterotetrameric terdiri dari dua pasang tipe I tipe II reseptor. Tipe I dan tipe II reseptor yang sama struktur domain secara keseluruhan, termasuk kaya sistein ekstraselular domain yang mengadopsi tiga lipat finger toksin, satu transmembran heliks, dan domain serin treonin intraseluler kinase. Encode genom manusia ada tujuh, lima reseptor tipe I dan tipe II. Melalui studi silang yang berbasis sel, BMP dan GDFS yang telah terbukti untuk mengikat beberapa reseptor tipe I dan tipe II di mixed order, sedangkan TGF βs mengikat satu reseptor tipe I dan tipe II di sequential order, pertama dengan mengikat tipe II, TbR II, diikuti oleh tipe I, TβR II. Temuan biokimia ini menunjukkan bahwa kompleks reseptor faktor pertumbuhan dari family ini mungkin berbeda secara struktural. Perbedaan perbedaan ini telah dibuktikan melalui analisis struktural langsung BMP dan TGF b: tipe I reseptor: tipe II reseptor kompleks terner. Struktur kompleks terner BMP disimpulkan pertama kali didasarkan pada struktur struktur independen BMP BMP terikat pada tipe I dan tipe II reseptor (masing masing Kirsch, et al. Dan Greenwald, et al.,), dan kemudian dikonfirmasi di analisis langsung oleh dua BMP kompleks terner (Allendorph, et al. dan Weber, et al.). Struktur terner TGF β kompleks, baru baru ini dilaporkan oleh Groppe, et al dan subjek ini menekankan, ditentukannya analisis struktural langsung dari TGF β3: TβR I: TβR II kompleks terner menggunakan kristal yang terdifraksi untuk sebuah resolusi 3,0 Å menggunakan SSRL beamline 11 1.
Structural fitur dari tipe I dan ligan reseptor yang mempromosikan peralihan dari BMP ke TGF b mode pengikatan. (A) Short ekstensi, dari D53 ke L55, di BMP 2 (coklat) pocket komplementer pada reseptor tipe I BMP, BMPR Ia (cyan). Seperti sebuah ekstensi absen di TGF b3 (merah), perlu sebuah cara alternatif tipe I reseptor yang mengikat. (B) TβR I, termasuk sebuah loop ekstensi (merah) dibandingkan dengan reseptor BMP tipe I, Ia dan BMPR BMPR pon. This extension berisi dua prolines (P55 dan P59), serta fenilalanin (F60), yang mengikat pocket di bawah jari TGF b yang dibentuk oleh W30, dengam W32, Y90, dan L101. (C) Daerah N terminal TβR II melibatkan TbR I oleh packing pada permukaannya dan dengan memasukkan dua hidrofobik residu, F24 dan V22, ke dalam pocket hidrofobik pada permukaan TβR I. TβR II lebih melibatkan TβR I oleh pasangan ion yang terbentuk antara karboksilat dari TβR II D118 dan kelompok guanidinium TβRI R58.
Jalur Signaling TGF β.
dimulai dengan pengikatan dari TGF beta superfamili ligan ke TGF beta reseptor tipe II. Tipe II reseptor adalah serin / treonin reseptor kinase, yang mengcatalyses fosforilasi Tipe I reseptor. Setiap klas mengikat ligan tipe tertentu II reseptor. Pada mamalia diketahui ada tujuh tipe I reseptor dan lima tipe II reseptor. Ada tiga activins: Aktivin A, B dan Aktivin Aktivin AB. Activins yang terlibat dalam embriogenesis dan osteogenesis. Mereka juga mengatur banyak hormon termasuk hipofisis, hormon gonad dan hipotalamus serta insulin. Mereka juga faktor survival sel saraf. The BMP mengikat protein reseptor morphogenetic Bone tipe 2 (BMPR2). Mereka terlibat dalam banyak fungsi sel, termasuk osteogenesis, diferensiasi sel, anterior / posterior axis spesifikasi, pertumbuhan, dan homeostasis.
Signaling TGF β
Keluarga TGF beta meliputi: TGFβ1, TGFβ2, TGFβ3. Seperti BMPS, TGF beta yang terlibat dalam embriogenesis dan diferensiasi sel, mereka juga terlibat dalam apoptosis, serta fungsi lainnya. Mereka mengikat reseptor TGF beta tipe 2 (TGFBR2). Nodal mengikat untuk aktivin A reseptor, ketik IIB ACVR2B. Ini dapat juga membentuk kompleks dengan reseptor aktivin A reseptor, ketik IB (ACVR1B) atau dengan aktivin A reseptor,tipe IC (ACVR1C) .
Perekrutan dan fosforilasi reseptor.
transmembran domain dan sebuah domain sitoplasma kaya serin / treonin. The GS domain dari tipe I reseptor terdiri dari serangkaian sekitar tiga puluh serin glisin yang berulang. Pengikatan keluarga TGF beta ligan menyebabkan rotasi reseptor sehingga sitoplasma kinase domain tersebut diatur dalam orientasi catalytically yang baik. Reseptor tipe II memfosforilasi residu serin reseptor tipe I, yang akan mengaktifkan protein.
Fosforilasi reseptor tipe I
phosphorylates serin R SMAD. Fosforilasi menginduksi perubahan konformasi dalam domain MH2 R SMAD dan selanjutnya disosiasi dari reseptor kompleks dan SARA.
Fosforilasi RSMAD memiliki afinitas tinggi untuk coSMAD (misalnya SMAD4) dan membentuk suatu kesatuan kompleks. Gugus fosfat tidak bertindak sebagai situs docking coSMAD, bukan membuka fosforilasi asam amino yang memungkinkan peregangan interaksi.
Perekrutan SMADs
Transkripsi.
Fosforilasi RSMAD menyebabkan transkripsi
Mekanisme Regulasi.
peran spesifikasi pada kiri kanan. Lefty bertindak mencegah fosforilasi R SMADs. Ia melakukannya melalui konstitutif tipe I TGFβ aktif reseptor dan melalui proses downstream dalam aktivasinya. Obat berbasis (drug base) antagonis juga telah teridentifikasi, seperti SB431542, yang secara selektif menghambat ALK4, ALK5, dan ALK7.
Transforming growth factor receptor 3 (TGFBR3) adalah kelompok TGF β reseptor yang paling banyak namun , ia tidak memiliki tanda yang dikenali domain . Namun dapat menjalankan peningkatkan pengikatan ligan TGF beta pada reseptor tipe II TGF beta oleh ikatan TGFβ dan dinyatakan sebagai TGFBR2. Salah satu target downstream sinyal TGF β, GIPC, mengikat pada PDZ domainnya, yang mencegah para proteosomal degradasi, kemudian meningkatkan aktivitas TGFβ. Mungkin juga berfungsi sebagai coreceptor inhibin untuk ActivinRII. BMP dan Aktivin membran mengikat inhibitor (BAMBI), yang memiliki domain ekstraselular yang sama seperti tipe I reseptor. Ketiadaan domain intraselular akan serin / treonin protein kinase dikenal sebagai pseudoreceptor. Ia mengikat reseptor tipe I untuk mencegah pengaktifannya. Berfungsi sebagai pengatur sinyal negatif TGF beta dan mungkin membatasi ekspresi TGF beta selama embryogeneis. Hal ini membutuhkan sinyal BMP untuk berekspresi. FKBP12 mengikat daerah GS tipe I mencegah fosforilasi reseptor reseptor oleh tipe II reseptor. Hal ini diyakini bahwa FKBP12 membantu untuk mencegah aktivasi homologs tipe I reseptor dalam ketiadaan ligan, karena mengikat ligan menyebabkan disosiasi.
Peran inhibitory Smad.
Protein Kinase.
Protein kinase adalah enzim kinase yang mengubah protein lain dengan menambahkan gugus fosfat kimia (fosforilasi). Fosforilasi biasanya menghasilkan perubahan fungsional protein target (substrat) dengan mengubah aktivitas enzim, lokasi selular, atau asosiasi dengan protein lain. Genom manusia mengandung sekitar 500 gen protein kinase dan sekitar 2% dari seluruh gen manusia. Protein kinase juga ditemukan pada bakteri dan tanaman. Hingga 30% dari semua protein manusia dapat dimodifikasi oleh aktivitas kinase, dan kinase dikenal untuk mengatur sebagian besar jalur selular, terutama yang terlibat dalam transduksi sinyal.
Fosforilasi oleh protein kinase
memerlukan adanya rangsang kimia. Sementara protein kinase A (PKA) dan protein kinase G (PKG) yang merupakan protein serin treonin kinase yang terlarut dalam sitoplasma memerlukan akstifasi dari nukleotida dan penyeranta kedua seperti cAMP dan cGMP. Sedangkan PKC yang merupakan keluarga terbanyak dari protein kinase distimuli oleh aktifitas penyeranta kedua DAG dan ion kalsium. Pada tingkatan transduksi yang dikatalisa oleh komplek kalmodulin kalsium akan teraktifasi suatu enzim kinase yang dikenal sebagai protein kinase yang bergantung pada kompleks kalsium kalmodulin.
Aktivitas Kimia.
dan Ca2 + kalmodulin. Satu kelompok yang sangat penting protein
kinase adalah MAP kinase (singkatan dari: mitogen mikrotubula activated protein kinase). Subkelompok penting adalah kinase dari subfamili ERK, biasanya diaktifkan dengan mitogenic sinyal, dan stres protein kinase JNK diaktifkan oleh P38. Sementara MAP kinase serin dan treonin spesifik, mereka diaktifkan oleh kombinasi fosforilasi pada serin treonin dan residu tirosin. Kegiatan MAP kinase dibatasi oleh jumlah protein fosfatase, yang menghapus gugus fosfat yang ditambahkan ke serin spesifik atau treonin residu dari kinase dan diperlukan untuk menjaga kinase dalam konformasi aktif. Dua faktor utama kegiatan tersebut mempengaruhi MAP kinase: a) sinyal yang mengaktifkan reseptor transmembran (baik ligan alam, atau agen silang) dan protein yang terkait dengan mereka (mutasi yang mensimulasikan keadaan aktif), b) sinyal bahwa menonaktifkan fosfatase yang membatasi suatu MAP kinase. Sinyal semacam itu termasuk stres oksidan. Spesifik tirosin protein kinase (EC EC 2.7.10.1 dan 2.7.10.2) memfosforilasi residu tirosin asam amino, seperti serin dan treonin kinase spesifik yang digunakan dalam signal transduksi. Mereka berperan terutama sebagai reseptor faktor pertumbuhan dan sinyal dari faktor pertumbuhan. Beberapa contoh reseptor tersebut antara lain:
Platelet Derived Growth Factor Receptor (PDGFR) Epidermal Growth Factor Receptor (EGFR) Insulin Growth Factor 1 Receptor (IGF1R) Colony Stimulating Factor Receprot (CSFR).
domain bertanggung jawab terhadap aktivitas kinase, serta beberapa fungsi regulasi. Apabila protein transmembran tersebut mengikat ligan menyebabkan dua reaksi:
1. Dimerization dari dua monomer reseptor kinase atau stabilisasi dimer yang longgar. Banyak ligan dari reseptor tirosin kinase multivalent. Beberapa reseptor tirosin kinase (misalnya, berasal dari platelet reseptor faktor pertumbuhan) yang dapat terbentuk heterodimers tetapi tidak identik kinase subfamili yang sama, sehingga respon sangat bervariasi untuk sinyal ekstraselular 2. Trans autophosphorylation (fosforilasi oleh kinase lain dalam
dimer) dari kinase.
histidin. Diregulasi aktivitas kinase sering menjadi penyebab penyakit, terutama kanker, dimana kinase mengatur banyak aspek yang mengontrol pertumbuhan sel, gerakan dan kematian. Obat yang menghambat kinase tertentu sedang dikembangkan untuk mengobati beberapa penyakit, dan beberapa saat ini dalam penggunaan klinis, termasuk Gleevec (imatinib) dan Iressa (gefitinib). Perkembangan obat untuk kinase inhibitor dimulai dari tes, biasanya senyawa diprofilkan untuk kekhususan sebelum pindah ke tes lebih lanjut. Banyak layanan profil yang tersedia dari neon berbasis radioisotop tes untuk deteksi. Seperti halnya protein kinase, protein kinase A (juga dikenal sebagai cyclic AMP dependent protein kinase atau A kinase) adalah enzim protein yang kovalen dengan gugus fosfat. Karakteristik yang unik dari protein kinase A adalah bahwa aktivitas diatur oleh tingkat fluktuasi siklik AMP dalam sel (maka nya alias sebagai cyclic AMP dependent protein kinase). Sehingga enzim ini berfungsi sebagai efektor akhir untuk berbagai hormon yang bekerja melalui jalur sinyal AMP siklik. Dengan kata lain, protein kinase A pada akhirnya bertanggung jawab atas semua respon selular karena AMP merupakan siklik sistem pembawa pesan kedua.
Genetika dan Struktur.
Protein kinase A holoenzyme adalah heterotetramer yang terdiri dari dua jenis subunit:
1. Katalitik subunit: subunit ini berisi situs aktif enzim. Itu juga memuat sebuah domain yang mengikat ATP (sumber fosfat) dan domain yang mengikat subunit regulasi.
Gen yang mengatur aktivasi protein kinase
Regulatory subunit ada dalam dua bentuk utama, RI dan RII,
dengan masing masing bentuk memiliki dua subtipe ditunjuk alfa dan
beta. Masing masing dari empat isotypes dari subunit peraturan
dikodekan oleh gen yang berbeda. Selain itu, tiga isotypes dari subunit
katalitik telah diidentifikasi (alfa, beta dan gamma). Isotypes yang
berbeda cenderung memiliki distribusi yang berbeda di dalam sel dan
di antara jaringan. Tipe I enzim sitoplasma, yang larut dari sel,
sedangkan tipe II enzim cenderung untuk bergabung dengan
membran selular. Konsentrasi intraselular siklik AMP memberikan
informasi paling mendasar yang mengontrol aktivitas protein kinase
A:
1. Ketika AMP siklik tingkat rendah, catalytic subunit terikat pada
subunit regulasi dimer dan tidak aktif.
2. Sebagai konsentrasi siklik AMP meningkat, ia mengikat ke
subunit regulasi, yang mengarah ke perubahan alosterik
konformasi yang menyebabkan melepaskan dari subunit katalitik.
3. Free subunit katalitik aktif dan mulai memfosforilasi target
Peranan cAMP pada pembentukan subunit bebas
1. Enzim: Fosforilasi secara luas digunakan sebagai mekanisme switching molekuler untuk mengaktifkan atau menonaktifkan aktivitas enzim. Dalam banyak kasus, enzim yang terfosforilasi itu sendiri adalah suatu kinase. Contoh klasik adalah bahwa protein kinase A fosforilase phosphorylates enzim kinase, yang, pada gilirannya, phosphorylates glikogen phorphorylase, yang mengarah pada pemecahan glikogen di hati dan otot.
2. Ion saluran: saluran kalsium tertentu dalam sel sel otot jantung diaktifkan oleh protein kinase A, pada akhirnya menyebabkan kontraksi otot. Medis lain contoh penting adalah bahwa protein kinase A phosphorylates dan dengan demikian mengaktifkan sebuah saluran klorida penting dalam sekresi air di usus kecil. 3. Kromosomal protein: Histone H1 adalah sasaran pertama
diidentifikasi untuk protein kinase A.
4. Faktor faktor Transkripsi: CREB’s (siklik AMP respon mengikat unsur protein) adalah faktor faktor transkripsi yang, ketika terfosforilasi oleh protein kinase A, menjadi kompeten untuk mengikat daerah promotor gen responsif dan merangsang transkripsi. Protein kinase A terlibat dalam proses proses penting seperti energi metabolisme, kontraksi otot, membran transportasi dan ekspresi gen.
Protein Kinase C.
Tirosin Kinase.
Tirosin kinase adalah subclas dari protein kinase. Prinsip dari protein adalah posporilasi.
Protein tirosin kinase
Tyrosin kinase adalah enzim yang dapat mentransfer suatu kelompok pospat dari ATP ke suatu residu tyrosin dalam suatu protein. Tyrosin kinase adalah subgrup dari kelas besar protein kinase. Posporilasi protein oleh kinase adalah suatu mekanisme penting dalam sinyal transduksi untuk regulasi aktivitas enzim. Tyrosin kinase dikelompokkan pada proten tyrosin pospatase. Protein kinase adalah group enzim yang mempunyai subunit katalitik yang mentransfer pospat gamma dari nukleotida tripospat (ATP) ke satu atau lebih residu asam amino dalam sisi rantai protein substrat yang menghasilkan perubahan konformasi yang mempengaruhi fungsi protein. Enzim dikelompokkan dalam dua kelas besar, dikarakter dengan respeknya terhadap spesifik substrat: spesifik serin/treonin dan spesifik tyrosin (domain ini).
Tirosin
Fosfat
!
Ada labih dari 100 3D struktur tyrosin kinase yang tersedia di Protein data Bank. Contoh adalah PDB 1IRK, struktur kristal tyrosin kinase domain human insulin receptor. Famili tyrosin kinase dibagi dalam dua famili utama : 1. Transmembran receptor linked kinase dan protein cytoplasmic. Rata rata terdapat 2000 kinase yang diketahui dan lebih dari 90 Protein Tyrosin Kinase (PTKs) yang terdapat di human genom. Mereka dibagi dalam dua kelas, receptor dan non receptor PTKs. Saat ini terdapat 58 receptor tyrosin kinase (RTKs) yang diketahui, dikelompokkan dalam 20 subfamili. Mereka berperan pivotal pada bermacam macam aktivitas seluler termasuk growth, differensiasi, metabolism, adhesi, motility, dan kematian. RTKs disusun suatu extracelluler domain yang dapat mengikat spesifik ligan, suatu transmembran domain dan intracelluler catalitik domain yang dapat mengikat dan posporilasi seleksif substrat.
kanker. Imatinib (jenis nama Gleevec dan Glivec) obat yang dapat mengikat potongan katalitik tyrosin kinase yang menghambat aktivitasnya. Dalam sel sel normal, TGF β, bertindak melalui jalur signaling, menghentikan siklus sel pada tahap G1, menghentikan proliferasi, menginduksi diferensiasi, atau apoptosis. Ketika sebuah sel berubah menjadi sel kanker, bagian dari TGF β jalur signaling bermutasi, dan TGF β tidak lagi mengendalikan sel. Sel sel kanker tersebut berkembang biak. Sel stroma di sekitarnya (fibroblas) juga berkembang biak. Kedua sel meningkatkan produksi TGF β. TGF β ini bekerja pada sel sel stroma di sekitarnya, sel sel kekebalan, endotel dan sel otot halus. Hal ini menyebabkan imunosupresi dan angiogenesis, yang membuat kanker lebih invasif. TGF β juga mengkonversi T sel efektor, yang biasanya menyerang kanker dengan peradangan (imun) reaksi, ke dalam regulasi T sel, yang mematikan reaksi peradangan.
Receptor TGF β
Interaksi aktivasi segmen bergerak ke posisi di tempat yang dapat menghambat ikatan ATP dan mencegah Smad fosforilasi. Ketika para GS phosphorylates RII loop, FKBP12 dilepas, loop GS interaksi dengan heliks spesifik adalah destryoed, dan pergeseran segmen aktivasi dari ikatan ATP yang memungkinkan ATP untuk mengikat dan menjadi Smad terfosforila. Dalam tutorial ini, lima RI protein mengkristal sebagai lima subunit pentamer, namun pada vivo RI hanya ada sebagai satu rantai protein, bukan agregasi lima ditampilkan.
KUIS
1. Apakah pentingnya TGF beta dalam homeostasis?
2. Apabila dilakukan knockout gen TGF beta fenotip apakah yang anda prediksi
BAB II
RESEPTOR TIROSIN KINASE DAN
RESEPTOR TIROSIN KINASE DAN
RESEPTOR TIROSIN KINASE DAN
RESEPTOR TIROSIN KINASE DAN
AKTIVASI RAS
AKTIVASI RAS
AKTIVASI RAS
AKTIVASI RAS
Reseptor Tirosin Kinase.
Reseptor tirosin kinase (RTK) adalah reseptor yang terlibat dalam sinyal transduksi, dan proses berbagai lingkungan serta sinyal intersellular. Sebaliknya, protein tirosin kinase (PTK) adalah enzim yang mengkatalisis fosforilasi residu tirosin. Dari 91 protein tirosin kinase diidentifikasi, 59 adalah reseptor tirosin kinase dan 32 non reseptor. Sebagai komponen sentral dari jaringan sinyal sel, RTK memainkan peran penting dalam proses fisiologis, seperti embriogenesis, perkembangan dan diferensiasi neuron, proliferasi sel, sinyal anti apoptosis dan kematian sel (apoptosis). Beberapa signaling molekul bertindak sebagai reseptor adhesi. Bagian adhesi kaya dengan tirosin protein yang mengalami fosforilasi sehingga terjadi coupling adhesi sel ke jalur sinyal transduksi dalam sel. Berbagai reseptor adhesi, seperti integrins, berhubungan erat dengan protein kinase dan fosfatase.
Reseptor Tirosin Kinase (RTKs) terdiri dari empat domain: 1. Domain ekstraselular ligan.
2. Domain tirosin kinase intraseluler, dengan sekuens asam amino dalam substrat ATP dan cAMP dependent protein kinase (cAPK, PKA).
3. Domain intraselular. 4. Domain transmembran.
immunoglobulin, domain kringle, dan daerah yang kaya leusin. Aktivasi kinase ini dilakukan dengan pengikatan ligan ke domain ekstraselular, yang menginduksi reseptor dimerization. Mengaktifkan reseptor tirosin autophosphorylate residu katalitik di luar domain melalui jalur fosforilasi. Ini secara otomatis menstabilkan konformasi fosforilasi reseptor aktif dan menciptakan area untuk phosphoTirosin docking protein yang mengirim sinyal tranduksi di dalam sel.
protein adaptor seperti Grb2 atau NCK hanya mengandung domain SH2 dan SH3. Signaling protein lainnya mengandung tambahan protein enzimatis seperti protein kinase PTPase (Shp2) fosfolipase C (PLC ), Ras GAP atau Rho GRF (Vav). (B) Docking protein yang berfungsi sebagai bentuk untuk perekrutan sinyal protein. Semua docking protein mengandung sebuah membrane terminal. FRS2 adalah target ke membran oleh myristoylation, dan LAT adalah ditargetkan ke membran sel oleh domain transmembran (TM) dan oleh palmytoylation. Kebanyakan protein docking ditargetkan ke membran sel oleh domain PH. Docking protein mengandung banyak bagian pTyr phosphorylation yang berfungsi sebagai tempat pengikatan untuk area SH2 berbagai sinyal protein.
domain dari PDK1 dan PKB. Translokasi membran disertai dengan pelepasan sebuah autoinhibition PKB kinase yang menyebabkan aktivasi dari kinase PDK1 dan PKB. Aktivasi PKB memerlukan phosphorylasi oleh PDK1 (dan juga oleh PDK2). Berbagai sasaran mencegah kematian dan apoptotic serta mengatur berbagai proses metabolisme. (B) aktivasi oleh perubahan konformasi. Pengikatan SH2 area p85, PI 3 kinase untuk bagian pTyr diaktifkan reseptor sebuah autoinhibition yang merangsang katalitik domain (p110). PI 3 kinase mengkatalisis fosforilasi dari 3 posisi Inositol PtdIns (4) P dan PtdIns (4,5) P2 untuk menghasilkan masing masing PtdIns (3,4) P 2 dan PtdIns (3,4,5) P3. (C) aktivasi oleh fosforilasi tirosin. Pengikatan wilayah SH2 PLC untuk pTyr dalam memfasilitasi baru diaktifkan oleh reseptor fosforilasi sinus PLC serta translokasi membran. Proses ini diperantarai sebagian oleh pengikatan domain PH ke PI 3. Tirosin. Fosforilasi tirosin adalah aktivasi penting untuk PLC menuju hydrolysis dari PtdIns (4,5) P2 dan pembentukan dua second messenger Ins (1,4,5) P 3 dan diacyglycosol.
polisakarida bermuatan negatif pada matriks ekstraseluler. Asosiasi ini meningkatkan pengikatan ligan terhadap monomer reseptor dan pembentukan kompleks ligan dimer reseptor. Ligan untuk beberapa RTK berbentuk dimer yang mengikat dua monomer bersama sama secara langsung. RTK yang lain, misalnya reseptor insulin, membentuk dimer dengan ikatan disulfida ketika tidak terdapat hormon. Pengikatan ligan terhadap RTK jenis ini akan mengubah konformasinya sehingga reseptor menjadi teraktivasi.
Dalam kondisi resting atau tidak terstimuli, aktivitas kinase intrinsik pada RTK sangat rendah. Dalam kondisi reseptor dimer, knase pada satu subunit mampu mmfosforilasi satu atau lebih residu Tirosin di dekat sisi katalisis pada subunit yang lain. Hal ini akan mengakibatkan perubahan konformasi yang memfasilitasi pengikatan ATP pada beberapa reseptor (misalnya reseptor insulin) dan pengikatan substrat protein pada reseptor lainnya (misalnya reseptor FGF) peningkatan aktivitas kinase ini kemudian memfosforilasi sisi lain dalam domain sitosolik reseptor. Aktivasi aktivitas RTK kinase yang diinduksi ligan ini mirip dengan aktivasi JAK kinase yang berasosiasi dengan reseptor kinase. Perbedaannya terletak pada lokasi sisi katalitik kinase, dimana sisi katalitik pada RTK terdapat pada domain sitosolik, sedangkan JAK kinase terpisah dari reseptor sitokin. Kebanyakan residu fosfoTirosin dalam RTK yang teraktivasi berinteraksi dengan protein adapter, protein kecil yang mengandung domain SH2, PTB, atau SH3, tetapi tidak memiliki aktivitas enzimatik intrinsik atau signaling. Protein protein tersebut merangkai RTK ke komponen yang lain dari jalur sinyal transduksi, misalnya aktivasi Ras.
Ras, GTPase Switch Protein.
mengikat GDP. Protein G trimerik secara langsung terikat dengan reseptor permukaan sel, sedangkan Ras tidak secara langsung terikat dengan reseptor permukaan sel. Aktivasi Ras dipercepat oleh guanine nucleotide exchange factor (GEF), yang berikatan dengan kompleks Ras GDP. Karena konsentrasi GTP dalam sel lebih tinggi daripada GDP, GTP berikatan secara spontan dengan molekul Ras, dengan melepaskan GEF dan membentuk Ras GTP aktif. Selanjutnya terjadi hidrolisis pada ikatan GTP menjadi GDP sehingga terjadi deaktivasi Ras. Tidak seperti deaktivasi Gα GTP, deaktivasi Ras GTP membutuhkan protein lain yang disebut GTPase activating protein (GAP) yang berikatan dengan Ras GTP dan mempercepat aktivitas GTPase intrinsik lebih dari seratus kali. GAP mengikat fosfoTirosin spesifik pada RTK yang teraktivasi, sehingga akan mendekatkan ke Ras GTP yang terikat membran untuk menggunakan kecepatan hidrolisis GTP. Hidrolisis GTP dikatalisis baik oleh Ras maupun GAP.
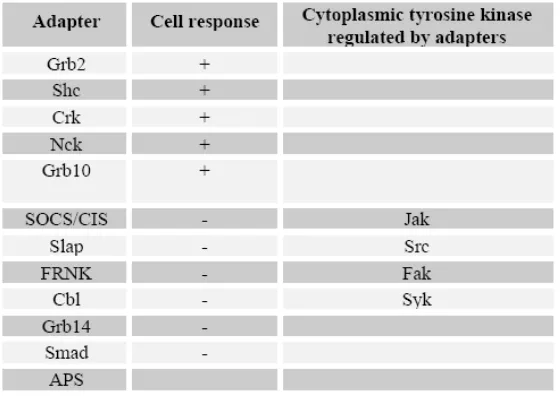
Protein adapter dan Guanine Nucleotide–Exchange Factor menghubungkan Ras dengan RTK yang teraktivasi.
Tabel 1. Protein adapter yang mengaktifkan (+) dan menghambat ( ) respon sel.
Beberapa kelompok reseptor yang termasuk dalam TRK menurut Zwick, 2001, di antaranya adalah:
• Epidermal growthfactor receptor (EGFR) family
Epidermal growthfactor receptor (EGFR) family terdiri atas empat anggota, yaitu EGFR/ErbB1, HER2/ErbB2, HER3/ErbB3 dan HER4/ErbB4. Keempat reseptor tersebut memiliki dua domain ekstraseluler yang kaya sistein dan bagian intraseluler dengan rantai C terminus yang panjang yang berfungsi sebagai tempat autofosforilasi. Anggota EGFR family diaktifkan oeh sekelompok besar EGF related growth, yang semuanya mengandung EGF like domain dan disintesis sebagai protein prekursur transmembran, termasuk transforming growth factor α (TGFα), epiregulin, betacellulin, heparin binding EGF like growth factor, dan amphiregulin.
IGFR family terdiri atas insulin receptor (IR) and the insulin like growth factor (IGF) receptor (IGF IR). Kedua resepto tersebut tersusun atas dua subunit α ektraseluler, yang berperan dalam mengikat ligan, dan dua membrane spanning β subunits yang menunjang domain Tirosin kinase dan autophosphorylation site. Ligan untuk kedua reseptor ini adalah insulin, IGF I dan IGF II. Insulin merupakan hormon metabolik, sedangkan IGF I dan IGF II berperan penting dalam perkembangan secara normal dan karsinogenesis.
• Vascular endothelial growth factor receptor (VEGFR) family Vascular endothelial growth factor (VEGF) merupakan salah satu inducer utama pada proliferasi sel endotel dan permeabilitas pembuluh darah. Terdapat dua RTK yang mampu mengikat VEGF, yaitu VEGFR 1 dan VEGFR 2, yang diekspresikan pada sel endotel selama perkembangan embrio dan merupakan regulator utama untuk angiogenesis. Beberapa penelitian menunjukkan bahwa sistem ligand– receptor VEGF–VEGFR berperan dalam vaskularisasi dan metastatis tumor. Sel tumor mensekresikan VEGF yang mengaktifkan VEGFR 2 dan menginduksi proliferasi sel endotel stroma.
• Fibroblast growth factors receptor (FGFR) family
• Hepatocyte growth factor receptor (HGFR)
Hepatocyte growthfactor receptor (HGFR), yang dikode oleh proto oncogene met, diidentifikasi sebagai regulator berbagai proses, seperti migrasi sel, persebaran sel dan invasi matriks ekstraseluler. HGFR merupakan heterodimer yang diikat oleh ikatan disulfida dengan α chain and a β chain ekstraseluler yang mengalami glikosilasi. HGFR tersusun atas domain transmembran dan domain Tirosin kinase sitoplasmik. Hepatocyte growth factor (HGF) atau scatter factor (SF), ligan reseptor ini, diekspresikan pada mesenchymal derived cells.
• RET receptor Tirosin kinase
RET receptor Tirosin kinase (RET RTK) merupakan gen yang bertanggung jawab atas Multiple endocrine neoplasia type 2 (MEN2). MEN2 merupakan syndrome cancer yang diturunkan dan dikarakterisiasi melalui pertumbuhan medullary thyroid carcinoma. RET proto oncogene mengkode protein yang dikarakterisasi oleh cadherin like domain dan cadherin like. RET diekspresikan selama embrigenesis pada sistem saraf tepi dan sistem urogenital. Kanker MEN2 terjadi karena dominan mengaktifkan germline ke dalam RET proto oncogene.
• Platelet derived growthfactor receptor (PDGFR) family
Ada dua anggota PDGFR family, yaitu platelet derived growth factor receptor (PDGFR) dan Kit. Protein tersebut dikarakterisasi melalui domain ekstraseluler dengan 5 Ig like domain dan sebuah domain Tirosin kinase intraseluler. Dua gen yang mengkode PDGFR α dan PDGFR β telah diidentifikasi. Kedua reseptor diaktivasi oleh ligan dimer yang tersusun atas PDGF A dan/atau PDGF B. Hal ini akan menginduksi dimerisasi reseptor dengan tiga kemungkinan konfigurasi, yaitu αα, ββ, αβ.
"#$
1. Apabila terjadi mutasi gen yang menyandi reseptor tirosin kinase, apakah prediksi yang saudara bisa kemukakan.
BAB III
G
G
G
G PROTEIN DAN SECOND
PROTEIN DAN SECOND
PROTEIN DAN SECOND
PROTEIN DAN SECOND
MESSENGER
MESSENGER
MESSENGER
MESSENGER
Organism khususnya organism multiselular untuk dapat melakukan regulasi dan koordinasi untuk tetap berada dalam keadaan yang homeostatis adalah dengan melakukan koordinasi antar sel di dalam tubuhnya. Sel untuk mampu melakukan koordinasi dengan sel lain adalah dengan melakukan komunikasi. Komunikasi antar sel ini dapat dilakukan karena adanya signal transduksi. Sel mampu menerima sinyal dari luar berupa sinyal kimia, sinyal elektromagnetik maupun sinyal mekanik. Dalam 1 sel mampu menerima lebih dari 1 sinyal kimia untuk tetap berada dalam kondisi homeostatis.Sinyal dapat diterima oleh sel lain melalui 3 tahap yaitu reception (sinyal di terima oleh reseptor), transduction (sinyal yang diterima kemudian ditindak lanjuti untuk proses berikutnya),dan respon (sinyal mentriger aktivitas celuler yang spesifik). Setiap sel target memiliki reseptor yang akan mengenali spesifik sinyal. Proses binding antara ligan (sinyal) dan reseptornya akan memberikan perubahan bentuk pada reseptor sehingga sinyal dapat masuk ke dalam sel dan berinteraksi dengan molekul di dalamnya. Sebagian besar reseptor merupakan protein membrane plasma dan sebagian besar sinyal merupakan molekul hidrofilik (water soluble) sehingga tidak mampu untuk menembus membrane plasma yang hidrofobik. Ada 3 macam tipe reseptor yaitu a) ion chanel reseptor, b) G protein reseptor dan c) Tirosin kinase reseptor.
G Protein Reseptor.
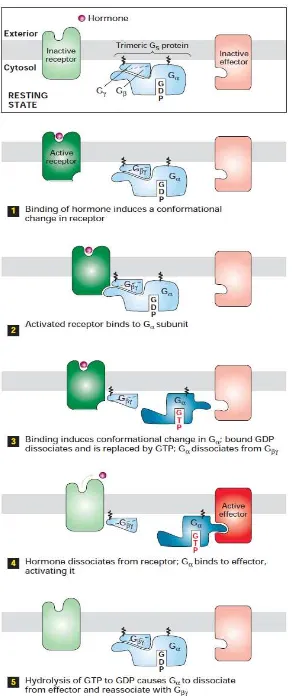
berhenti saat molekul sinyal ekstraselular tidak lagi berikatan pada reseptor G protein reseptor merupakan reseptor dari berbagai macam signal molekul antara lain hormone, neurotransmitter, dan local mediator yang mana G protein reseptor memiliki variasi struktur berbeda sesuai dengan fungsinya. Ligan yang sama akan mengaktifkan berbagai macam family berbeda. Kurang lebih 9 G protein linked reseptor di aktifkan oleh adrenalin, 5 reseptor yang lain diaktifkan oleh acetylcoline dan kurang lebih 15 oleh serotonin. G protein reseptor bekerja secara tidak langsung untuk melakukan regulasi aktivitas separasi protein target ikatan membrane plasma yang dapat berupa enzyme atau ion chanel. Atau dengan kata lain, G protein reseptor secara tidak langsung mengaktivkan atau menon aktifkanikan membrane plasma enzyme atau ion chanel. Interaksi antara reseptor dan protein target melalui 3 protein yang dikenal dengan nama trimeric GTPbinding regulatory protein (G protein) (Gambar 10).
G protein linked reseptor. Mekanisme reseptor G protein (Gambar 10) memperlihatkan ligan menempel pada reseptor. Dengan menempelnya ligan dengan reseptor akan mengaktifkan G protein dengan cara menempel pada reseptor. G protein yang menempel pada reseptor akan melakukan phosporilasi dan G protein lepas dari reseptor kemudian menempel dan mengaktifkan enzim atau ion chanel. Dan enzim dan ion chanel yang telah aktif akan mengaktifkan proses downstream nya.
Second Messenger.
molekul kecil, non protein, molekul water soluble atau ion yang di sebut second messenger. Molekul molekul ini secara cepat berdifusi masuk ke dalam sel. Second messenger ini berpartisipasi dalam inisiasi pathway melalui G protein linked reseptor, tyrosin kinase reseptor dan beberapa ion chanel. Secara umum terdapat 2 second messenger yang berperan penting yaitu cyclic AMP (cAMP), IP3,
maupun Ca2+.
Ada 3 tipe dasar molekul second messenger:
• Molekul hydrofobik (tidak larut dalam air) seperti diacylglycerol dan phosphatidylinositol, yang mana membrane berasosiasi dan berdifusi dari membrane plasma menuju ke dalam intermembran yang dapat mencapai dan meregulasi asosiasi membrane protein effektor.
• Molekul hydrofilik (larut dalam air) seperti cAMP, cGMP, IP3 dan Ca+ yang berada di dalam sitosol
• Gas sepeti NO dab CO dimana mampu berdifusi melalui sitosol dan keluar membrane selular.
Messenger intraselular secara umum memiliki kemampuan:
• Bisa disintesis atau di lepaskan dan di pecahkan kembali pada reaksi yang spesifik melalui mekanisme enzyme atau ion chanel
• Beberapa (seperti Ca+) dapat disimpan pada organela spesifik
dan secara cepat dilepaskan ketika dibutuhkan
alpha subunit dari G protein lepas dari ikatan subunit beta dan gamma. Kemudian G protein akan memproduksi suatu molekul primary effektor yang aktifitasnya kemudian membuat sinyal baru yang mampu berdifusi masuk ke dalam sel.sinyal ini yang kemudian disebut sebagai secondary messenger (Neurotransmitter sebagai first messenger). Second messenger dapat mengaktifkan secondary effector yang pengaruhnya tergantung pada particular dari sistem second messenger.
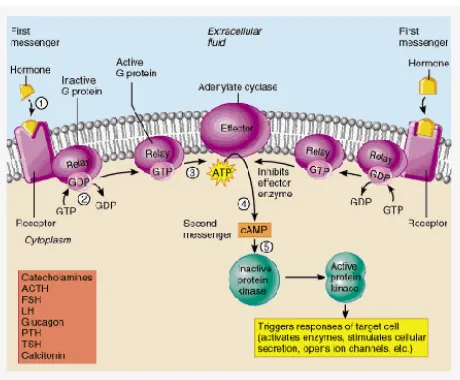
G Protein Reseptor Activated Adenylate Cyclase.
G protein berfungsi sebagai intermediary antar reseptor reseptor hormone dan enzim enzim effektor. G protein berfungsi sebagai mekanisme regulasi metabolism molekul molekul dalam merespon sinyal hormonal dan merubahnya. Komplek G protein terdapat alpha (α), beta (β) dan gamma (λ) subunit dan diikuti dengan disosiasi komplek, porsi dari masing masing bagian bertugas dalam mekanisme aktivitas adenylyl cyclase. Ketika hormone ekstraselular berikatan dengan sel reseptor, reseptor kemudian menginisiasi pelepasan GDP dari subunit G alpha dan kemudian menginisiasi binding antara GTP dan G alpha subunit. Sehingga terjadi disassosiasi antara G alpha subunit dari komplek G beta/gamma. Pada titik ini, aktifnya g alpha subunit dan lepasnya kompleks G beta/gamma dapat menstimuli aktifitas adenylyl cyclase (Gambar 10). Pada umumnya, adenylyl cyclase bertindak sebagai enzyme yang mengkatalisi ikatan cyclisasi. Secara umum, adenylyl cyclase juga bertindak sebgai enzim effektor dimana mengkatalisis 5’Adenosis Triphosphat (ATP) menjadi cyclic Adenosine Monophosphat (cAMP). Supaya adenylyl cyclase mampu berubah menjadi cyclize ATP, G alpha harus menempel pada sisi penempelan G alpha. Ketika G alpha subunit berikatan, adenylyl cyclase akan menkonformasi perubahan, katalitik C1 dan C2 mengorientasi untuk memulai mengambil energy dari ATP dan memulai proses cyclisasi. Adenylate cyclase mengkatalisi ATP dari plasma membrane menjadi cAMP + PPi. Umumnya ligan yang mengaktifkan adenylate cyclase adalah berupa hormone (ex: epinephrine). Hormon menempel pada reseptor kemudian mengakitfkan adenylate cyclase menjadi cAMP. cAMP ini merupakan second messenger. Kemudian setelah proses selesai, enzyme phosphodiester mengkatalisis cAMP + H2O → AMP.
Kinase A sehingga cAMP menstimuli proses degradasi sehingga sinyal cAMP berhenti. Ketika cAMP dikatalisis, GTP pada G alpha berubah menjadi GDP dan G alpha subunit lepas adari cyclase. Saat terlepas, g alpha subunit berasosiasi kembali dengan komplek G beta/gamma. Produksi dari cAMP di mulai ketika hormone ekstraselular yang lain menempel pada protein reseptor dan memulai proses kembali.
Gambar 21.
G Protein Coupled Reseptor – Activated Adenylate cyclase
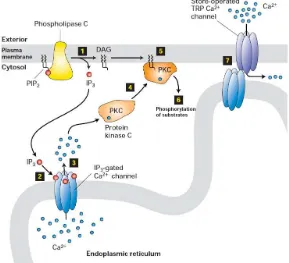
G Protein Reseptor Activated Phospholipase C.
Phospholipase C merupakan kelas dari enzyme yang membelah (cleave) phospholipid saat sebelum gugus phosphate. sama seperti enzyme enzym lainnya dimana phospholipase C juga berperan penting dalam mekanisme fisiologi sel eukaryotic, particular dari sinyal transduksi pathway. Terdapat 13 macam phospholipase C dari mamalia yang diklasifikasikan berdasarkan bentuk strukturnya kedalam 6 model yaitu β, γ, δ, ε, ζ, η.
Reseptor yang mengakitifkan jalur ini utamanya adalah Reseptor G protein yang berpasangan dengan Gαq subunit, yang
termasuk didalamnya antara lain: Reseptor 5 HT2 serotonergic
Reseptor α1 (Alpha 1) adrenergic
Reseptor calcitonin Reseptor H1 histamin
Reseptor metabotropic glutamate Reseptor M1, M3, M5 musarinic
Reseptor minor activator selain Gαq antara lain
MAP kinase. Mengaktivasi jalur PDGF dan FGF
Βγ kompleks dari heterotrimeric G protein sebagai jalur minor dilepasnya growth hormone oleh growth hormone releasing hormone
PLC membelah phospholipid. Dalam prosesnya, phosphatidylinositol 4,5 bisphosphate (PIP2) membelah menjadi
diacyl glycerol (DAG) dan inositol 1,4,5 triphosphate (IP3). DAG
tetap berikatan dengan membrane dan IP3 dilepaskan sebagai struktur
yang soluble ke dalam sitosol. IP3 kemudian berdifusi melalui sitosol
untuk berikatan dengan repetor IP3, particular chanel calcium dan
reticulum endoplasma. Hal ini menyebabkan konsentrasi calcium dalam sitosol meningkat yang menyebabkan perubahan cascade dari intraselular dan aktivitasnya. Selain itu, calcium dan DAG bekerja bersama untuk mengaktivkan protein kinase C dimana akan berfosforilasi menjadi molekul lain, yang menimbulkan adanya aktifitas selular. IP3 akan mentriger untuk melepaskan Ca
2+ dari RE.
Sebagian besar ion Ca2+ disimpan pada mitokondria dan di lumen RE
dan vesikel yang lain. Sel memiliki mekanisme untuk meregulasi konsentrasi ion Ca2+ di sitosol dimana biasanya dibawah 0,2M. Ca2+
ATPase memompa ion Ca2+ melalui plasma membrane ke sel
eksterior atau lumen intraselular Ca2+. Berikatnnya banyak hormone
pada reseptor membrane di hati liver, fat, dan sel lain menginduksi adanya elevasi di dalam Ca2+ sitosolik meskipun ketika ion Ca2+ tidak
tersedia dari sekitar ekstraselular. Pada situasi ini Ca2+ akan dilepas ke
dalam sitosol dari RE melalui aktivasi dari IP3 yang membuka ion
memiliki IP3 binding site di sisi N terminal. IP3 binding menginduksi
membukanya ion chanel sehingga ion Ca2+ dapat keluar dari RE ke sitosol (Gambar 12). Satu dari bermacam respon selular yang diinduksi keluar dari ion chanel yaitu sitosolik Ca2+ merupakan
rekrutmen protein kinase C (PKC) menuju membrane plasma dimana telah diaktifkan sebelumnya oleh DAG. Aktifnya PKC menstimuli phosphorilasi bermacam enzim selular dan reseptor. Sepeti halnya pada Ca2+ RE, chanel IP
3gated Ca
2+ berikatan, sehingga membuka
cahnel TRP Ca2+ di dalam plasma membrane sehingga ion Ca2+
masuk dari ekstraselular ke dalam sitosol.
Gambar 23. G Protein Coupled – Activated Phospholipase C
Konsentasi hormon dalam cairan ekstrasel sangat rendah
molekul lain. Derajad pembeda dilakukan oleh molekul pengenal yang terikat pada sel target disebut Reseptor →Reseptor Hormon merupakan molekul pengenal spesifik dari sel tempat hormon berikatan sebelum memulai efek biologiknya. Umumnya pengikatan Hormon :Reseptor ini bersifat reversibel dan nonkovalen reseptor hormon bisa terdapat pada permukaan sel (membran plasma% atau pun intraselluler Interaksi hormon dengan reseptor permukaan sel akan memberikan sinyal pembentukan senyawa yang disebut sebagai second messenger (hormon sendiri dianggap sebagai first messenger) Jika hormon sudah berinteraksi dengan reseptor spesifiknya pada sel sel target, maka peristiwa peristiwa komunikasi intraseluler dimulai.Hal ini dapat melibatkan reaksi modifikasi seperti fosforilasi dan dapat mempunyai pengaruh pada ekspresi gen dan kadar ion. Peristiwa peristiwa ini hanya memerlukan dilepaskannya zat zat pengatur. Hormon dapat diklasifikasikan melalui berbagai cara yaitu menurut komposisi kimia, sifat kelarutan, lokasi reseptor dan sifat sinyal yang memperantarai kerja hormon di dalam sel.
Klasifikasi hormon berdasarkan senyawa kimia pembentuknya:
1. Golongan Steroid→turunan dari kolestrerol 2. Golongan Eikosanoid yaitu dari asam arachidonat
3. Golongan derivat Asam Amino dengan molekul yang kecil (Thyroid,Katekolamin)
4. Golongan Polipeptida/Protein (Insulin,Glukagon,GH,TSH )
Berdasarkan sifat kelarutan molekul hormon:
Berdasarkan lokasi reseptor hormon:
1. Hormon yang berikatan dengan hormon dengan reseptor intraseluler
2. Hormon yang berikatan dengan reseptor permukaan sel (plasma membran)
Berdasarkan sifat sinyal yang mengantarai kerja hormon di dalam sel:kelompok
Hormon yang menggunakan kelompok second messenger senyawa cAMP,cGMP,Ca2+, Fosfoinositol, Lintasan Kinase sebagai mediator intraseluler
&! ' (
) ! *( + ( , + %
+( !
. Target molekul Protein G
1. Kanal ion Asetilkolin menyebabkan disosiasi α dan ßγ 2. Enzim yang terikat pada membran plasma
a. Adenil siklase merubah ATP → cAMP
b. Fosfolipase C akan merubah inositolfosfolipid menjadi IP3 dan
Adenylate Cyclase.
Transmembran reseptor dari berbagai hormon yang digabungkan untuk adenilat cyclase (AC) melalui heterotrimeric G protein. Ligan mengikat reseptor, mengubah konformasi reseptor, sehingga memungkinkannya untuk berasosiasi dengan G protein. Hal ini menyebabkan aktivasi G protein spesifik melalui pertukaran GTP menjadi GDP dan terikat pada subunit α G protein. Yang diaktifkan G protein pada gilirannya mengaktifkan AC menghasilkan konversi ATP untuk cAMP. cAMP lalu bertindak untuk mengatur berbagai proses selular. AC dapat berpasangan dengan baik dan menstimulasi inhibitor G protein (Gs dan Gi, masing masing). Gs merangsang interaksi aktivitas dan interaksi dengan Gi menghambat aktivitas enzimatik. Adenilate cyclase yang teraktifkan oleh sub unit dari protein G akan menghasilkan cAMP, dimana cAMP akan berperan sebagai penyeranta kedua yang mengaktifasi protein kinase A (PKA). PKA akan mengaktifasi beberapa protein factor transkripsi yang akan menstimulasi proses penyandian gen structural. Tiga subkelompok reseptor adrenergik berhubungan dengan sistem adenilat siklase. Hormon yang terikat pada reseptor β1 dan β2 akan mengaktifkan enzim adenilat siklase, sedangkan hormon yang terikat pada reseptor α2 akan menghambat enzim ini. Kerja hormon epineprin dapat meningkatkan kadar cAMP dalam sel otot melalui pengaktifan sistem β adrenergik ini yang melalui perangkaian reseptor pada Potein G. Protein G mengikat GTP dan GTP merangsang adenilat siklase untuk mensintesis cAMP. cAMP yang terbentuk akan mengaktifkan enzim fosforilase kinase dan menginaktifkan enzim glikogen sintase melalui aktifitas protein kinase. Enzim Adenilat Siklase berada pada permukaan internal membran plasma mengkatalisasi pembentukan cAMP dari ATP. Apabila aktifitas enzim Adenilat Siklase meniongkat maka jumlah cAMP juga meningkat. Pengaturan aktivasi dan inaktivasi enzim Adenilat siklase oleh hormon berlangsung dengan perantara:
1. Reseptor spesifik hormon pada permukaan luar membran plasma (Rs atau Ri)
Protein pengatur ini diberi simbol Gs(stimulasi) dan Gi(inhibisi) yang masing masing tersusun tiga subunit α,β,γ. Subunit β dan γ dalam Gs identik dengan dalam Gi, sedangkan subunit α dalam Gs berbeda dengan dalam Gi diberi tanda αs dan αi. Pengikatan sebuah hormon dengan reseptor meningkatkan interaksi reseptor dengan kompleks perangsang Gs .Dengan pengantaraan reseptor berlangsung pengikatan GTP yang tergantung pada Mg2+ oleh α dan disosiasi sekaligus β dan γ dari α. Subunit α dapat juga merupakan ADP ter ribosilasi sebagai respon terhadap toksin Kolera yang mengaktivasinya.Dalam menimbulkan proses tersebut akan membuat inaktif enzim GTPase,dengan demikian αs dibekukan dalam bentuk aktif.Toksin Pertusis dapat memblokir inaktivasi dari adenilat siklase melalui aktivitas ribosiltransferase ADP pada subunit αi
Phospolipase C.
Protein Kinase dan Protein Fosfatase.
Protein kinase terdapat baik di membrane plasma maupun di dalam sitoplasma. Pengklasifikasian protein kinase biasanya dilakukan berdasar jenis asam amino yang difosforilasinya seperti tirosin kinase atau serin treonin kinase. Protein kinase secara fungsional bertugas membantu fosforilasi beberapa protein agar dapat menjalankan tuganya selaku sinyal transduktor. Sinyal transduksi yang memerlukan kehadiran protein kinase antara lain cAMP yang bergantung pada enzim kinase. Tirosin kinase membran dan serin treonin kinase sebagai protein kinase di tingkat membran untuk dapat teraktifasi memerlukan adanya rangsang kimia. Sementara itu protein kinase A (PKA) dan protein kinase G (PKG) yang merupakan protein serin treonin kinase yang terlarut dalam sitoplasma memerlukan aktifasi dari nukleotida dan penyentara kedua seperti cAMP dan cGMP. Sedangkan Protein Kinaase C yang merupakan keluarga terbanyak dari protein kinase distimulasi oleh aktifitas penyentara kedua DAG dan ion kalsium.
( ! ( " - . /& (
Dalam sel eukariot, cAMP berikatan dengan Protein Kinase yaitu sebuah molekul heterotetramer terdiri atas 2 subunit regulasi dan 2 subunit katalitik. Pengikatan cAMP menghasilkan reaksi: 4 cAM + R2C2 ↔ 2 (R 2cAMP) + 2C
. Pengaktifan G protein
cGMP sebagai Second Messenger.
cGMP merupakan senyawa second messenger yang dibentuk dari GTP oleh kerja enzim Guanilil Siklase, yang terdapat dalam bentuk larut dan terikat membran.Hormon Atriopeptin, suatu famili peptida dihasilkan dalam atrium jantung, menyebabkan natriuresis, diuresis,vasodilatasi otot dan inhibisi sekresi aldosteron. Hormon peptida ini akan mengaktifkan enzim guanilil siklase sehingga cGMP meningkat dan dapat menyebabkan hormone bekerja efisien. Senyawa nitroprusida,nitrogliserin, natrium nitrit, natrium azida, nitogen oksida (NO) meningkatkan cGMP dengan mengaktifkan guanilil siklase . Peningkatan cGMP akan berikatan dan mengaktifkan Protein Kinase Spesifik (Kinase G ) yang analog dengan Kinase A. Enzim ini akan melakukan fosforilasi terhadap sejumlah protein otot polos. Peristiwa ini agaknya terlibat dalam proses relaksasi otot polos dan vasodilatasi.
Ca2+ Sebagai Second Messenger.
1. Dihambat dalam media kalsium bebas atau bila kadar kalsium intrasel berkurang
2. Mempengaruhi aliran kalsium sel
Diketahui konsentrasi Ca2+ sitosol lebih rendah dibandingkan konsentrasi Ca2+ dalam cairan ekstraseluler dan organela intraseluler.Keadaan ini dipertahankan oleh adanya pompa Ca2+ / Mg2+ ATPase dependent.Hormon dan zat efektor lain dapat merangsang pelepasan ion kalsium ke dalam sitosol. Jalan utama hormon meningkatkan penambahan Ca2+ adalahmelalui stimulasi dari produksi InsP3 yang dihasilkan oleh pemecahan dari PIP2 yang diperantarai fosfolipase C. PIP2 (Phosphatidil Inositida 4,5 Bisphosphat) merupakan senyawa phospholipid dari membran, memainkan peranan dalam aksi hormon yang tergantung Ca2+. Produk metabolisme PIP2 diduga menyediakan komunikasi antara reseptor hormon membran plasma dengan reservoir Ca2+ intrasel dan mempengaruhi Ca2+ channel. Dalam aksi hormon PIP
2 akan
dihidrolisis menjadi dua senyawa yaitu :
1. Inositol 1,4,5 Triphosphat (InsP
3), merupakan senyawa yang
efektif mempengaruhi mitokhondria dan RES mengeluarkan Ca2 + k sitoplasma.
2. Diasil Gliserol, mampu mengaktifkan protein kinase sehingga terjadi fosforilasi sejumlah protein, sebahagian merupalan komponen pompa ion dan mendorong peningkatan ion kalsium sitoplasma
"#$
1. Apakah kaitan G protein dengan pembentukan Energi level sel?
BAB IV
PERTAHANAN TUBUH
PERTAHANAN TUBUH
PERTAHANAN TUBUH
PERTAHANAN TUBUH
Mikroorganisme yang menyebabkan penyakit pada manusia dan hewan masuk ke dalam tubuh dengan berbagai cara. Secara umum mikroorganisme yang dapat menimbulkan penyakit disebut patogen. Patogen yang telah masuk akan menimbulkan penyakit dengan pelbagai mekanisme. Segala macam mikroorganisme yang menginvasi vertebrata akan berhadapan dengan imunitas innate sebagai pertahanan pertama yang terjadi beberapa menit setelah infeksi. Imunitas adaptif akan timbul apabila pertahanan pertama ini tidak mampu mengeliminasi patogen yang masuk.
. ! . Mekanisme
efektor yang membersihkan agen penginfeksi, contohnya makrofag dan komplemen. Dua fase pertama tidak memerlukan spesifikasi antigen, artinya semua antigen akan dikenali oleh sistem imun yang bekerja pada dua fase pertama ini. Fase ketiga adalah fase imunitas adaptif. Fase ini memerlukan
Imunitas innate (cepat: 0-4 jam)
Respon yang terinduksi (segera: 4-96 jam)
Respon imunitas adaptive (lambat: > 96 jam)
Infeksi
Infeksi
Infeksi
Pengenalan oleh efektor nonspesifik
Perekrutan sel efektor
Transpor antigen menuju organ limfoid
Pemusnahan agen penginfeksi
Pengenalan, aktivasi sel efektor
Pengenalan oleh sel T dan B naive
Ekspansi klon dan diferensiasi sel
efektor
Pemusnahan agen penginfeksi Pemusnahan agen
reseptor yang spesifik yang terbentuk dari gene rearrangement. Imunitas adaptif bekerja pada fase akhir.
Pertahanan pertama tidak dapat menuntaskan tugasnya antara lain karena besarnya jumlah invader yang masuk, cacat genetik, maupun lemahnya sistem pertahanan itu sendiri akibat kurang gizi. Sel sel epitel pada permukaan tubuh mempunyai peran penting sebagai penghalang masuknya mikroorganisme dalam tubuh. Sekresi kelenjar minyak maupun keringat juga mempunyai peran dalam sistem pertahanan pertama. Makrofag dan neutrofil merupakan komponen selluler pertahanan pertama yang bersifat fagosit, sedangkan NK berperan sebagai sitotoksik pada pertahanan pertama. NK merupakan sel yang memiliki jalur sama dengan sel limfosit hanya saja tidak mempunyai antigen khusus yang dikenali pada targetnya. NK mengenali sel yang mengalami kanker dengan cara mendeteksi penurunan ekspresi molekul MHC.
memori setelah terjadinya infeksi. Kerja imunitas innate ini pada umumnya berhasil menghalangi terjadinya infeksi. Apabila imunitas innate tidak berhasil mengeliminasi agen penginfeksi, makrofag dan sel lain yang telah teraktivasi pada respon innate akan segera membantu inisiasi respon imunitas adaptif.
Pengenalan Antigen Oleh Sel B dan T.
Telah dijelaskan sebelumnya bahwa tubuh dipertahankan dengan imunitas innate, tetapi sistem tersebut hanya mengontrol patogen yang mempunyai susunan molekul tertentu atau patogen tersebut menginduksi tersintesisnya interferon atau molekul efektor lain. Imunitas innate tidak membentuk memori dan imunitas innate ini bekerja dengan reseptor yang dikode di dalam genom. Imunitas innate sangat penting untuk menjaga agar patogen tidak berkembang bebas di dalam tubuh, namun imunitas innate tidak memiliki sifat yang dimiliki inunitas adaptif. Imunitas adaptif memiliki memori yang bertahan dalam waktu sangat lama terhadap antigen spesifik. Untuk mengenali dan melawan patogen yang memiliki diversitas tinggi, limfosit sebagai komponen imunitas adaptif telah berkembang dan dapat mengenali diversitas yang tinggi dari antigen bakteri, virus, dan organisme penyebab penyakit lainnya. Molekul pengenalan sel B adalah imunoglobulin, Ig. Imunoglobulin diproduksi oleh sel B dalam keadaan yang sangat beragam sesuai dengan keragaman antigen. Setiap sel B memproduksi imunoglobulin tunggal. Imunoglobulin yang berada pada permukaan sel berfungsi sebagai re