Mikoriza Arbuskuler Meningkatkan Toleransi Tanaman
TerhadapTanah Serpentin
Badruzsaufari
1,a), Akhmad Rizali Saidy
2), dan Noor Faiqoh Mardatin
3)Program Studi Biologi, FMIPA, UNLAM, Banjarbaru Program Ilmu Tanah, FAPERTA, UNLAM, Banjarbaru
Puslit Sumberdaya Hayati dan Bioteknologi IPB badruzsaufari@gmail.com
Abstrak. Tanah serpentin terdapat di Kalimantan Selatan, merupakan hasil pelapukan batuan ultramafik sehingga banyak mengandung logam berat, seperti Cr, Mn, Ni, Co, Fe, dan Mg. Rendahnya rasio Ca dan Mg menyebabkan tanaman sulit tumbuh di tanah tersebut. Cendawan mikoriza arbuskuler (CMA) banyak mengkolonisai perakaran Ischaemum barbatum yang tumbuh pada tanah serpentin. Hal ini menimbulkan dugaan bahwa mikoriza arbuskuler memiliki peranan dalam adaptasi tanaman terhadap tanah serpentin. Dua isolat cendawan tersebut diisolasi berdasarkan metode ekstraksi dengan penyaringan bertingkat. Kedua isolat tersebut beserta inolukum komersial dinokulasikan pada I. barbatum yang berasal dari tanah serpentin dan non-serpentin dan tanaman penutup tanah non-serpentin Pueraria japanica, dan kedua tanaman tersebut ditumbuhkan pada tanah serpentin. Inokulasi dengan CMA mampu meningkatkan berat kering tanaman dan serapan logam berat Cr dan Ni. Inokulum komersil berisi beragam spesies CMA mampu meningkatkan berat kering dan serapan logam berat sama seperti atau lebih baik daripada isolat asal tanah serpentin. Hal tersebut menunjukan bahwa mikorizsa arbuskuler mempunyai peranan penting dalam toleransi tanaman terhadap kondisi ekstrim tanah serpentin.
Kata Kunci. serpentin, mikoriza arbuskuler, logam berat, toleransi
PENDAHULUAN
Tanah serpentin merupakan hasil pelapukan batuan ultramafik yang dicirikan oleh tingginya kandungan unsur krom (Cr), mangan (Mn), kobal (Co), nikel (Ni), magnesium (Mg), dan besi (Fe) tetapi kandungan unsur esensial lainnya seperti kalsium (Ca), nitrogen (N), dan fosfor (P) rendah. Miskinnya unsur hara dan tingginya kandungan logam berat pada tanah tersebut menyebabkan hanya tumbuhan tertentu yang dapat beradaptasi. Selain itu, logam berat seperti pada lahan serpentin berpotensi menjadi polutan tanah dan air yang dapat menimbulkan bahaya bagi kesehatan dan lingkungan. Eksplorasi pertambangan pada lahan serpentin berpotensi menimbulkan pencemaran terhadap lingkungan di sekitarnya. Usaha untuk mengatasi atau paling tidak mengurangi masalah di atas, diantaranya
dapat dilakukan dengan melakukan pengurangan kadar logam berat dalam tanah yang tercemar melalui fitoremediasi. Selain itu, pilihan lain adalah melakukan revegetasi dengan tanaman yang mampu beradaptasi (toleran) terhadap tanah serpentin.
Tumbuhan Ischaemum barbatum merupakan rumput yang mendominasi lahan serpentin, seperti yang terdapat di Desa Mandiangin, Kabupaten Banjar, Kalimantan Selatan dan mempunyai kemampuan dalam mengakumulasi logam Cr dan Ni dalam jaringannya. Tumbuhan tersebut mempunyai laju pertumbuhan, produksi biomasa yang tinggi, dan perakaran yang kuat . Karakter di atas menunjukan bahwa I. barbatum tersebut merupakan tumbuhan yang toleran terhadap tanah serpentin dan mempunyai potensi untuk digunakan dalam upaya revegetasi atau reklamasi lahan. Sampai saat ini belum diketahui dengan jelas faktor yang mempengaruhi toleransi tumbuhan tersebut terhadap tanah serpentin.
Perakaran tumbuhan I. barbatum yang tumbuh pada berbagai vegetasi di lahan serpentin Mandiangin dikolonisasi sebanyak 40 – 65% oleh cendawan mikoriza arbuskuler (CMA). Kolonisasi tersebut berkorelasi positif dengan banyaknya logam Cr dan Ni yang diakumulasi di akar. CMA termasuk dalam kelas Glomeromisetes dan merupakan fungi yang membentuk berbagai struktur seperti arbuskuler dan beberapa spesies mempunyai vesikel serta hifa internal dan eksternal akar. Hifa eksternal membentukan jaringan ekstramatrikal yang luas dan memiliki berbagai fungsi seperti absorpsi air dan fosfor. Karena itu, CMA memiliki beragam efek positif terhadap pertumbuhan tanaman, produktivitas, dan melawan cekaman lingkungan. Adanya hubungan simbiotik dengan cendawan mikoriza tersebut diduga menjadi hal yang membantu tanaman dalam mentoleransi tanah serpentin. Untuk itu, isolat CMA dari tanah serpentin diuji kemampuannya mengkolonisasi akar dan mengakumulasi logam Cr dan Ni serta meningkatkan pertumbuhan I. barbatum asal tanah non-serpentin dan tanaman penutup tanah, Pueraria javanica, yang ditanam pada tanah serpentin.
METODE PENELITIAN
BAHAN DAN ALAT
Bahan tanaman yang digunakan adalah I. barbatum asal tanah inceptisol (sepertin) Mandiangin (IBMD), I. barbatum asal tanah non-serpentin Lianganggang (IBLA), dan P. javanica. Inokulum lainya yang digunakan adalah inokulum mikoriza
komersil (‖Techno‖) dan inokulum CMA
yang diisolasi dari tanah non-serpentin Lianganggang (LA). Bahan kimia yang digunakan untuk mewarnai spora adalah
larutan Melzer‘s, PVLG, pewarna trypan
blue 0,05%, gliserol 50%, KOH 10%, HCl 1%. Larutan sukrosa 30% dan 60% digunakan untuk memisahkan spora dalam larutan gradien sukrosa dengan sentrifus (Celement 2000). Kultur tabung menggunakan zeolit aktif 2 mm asal Bogor. Kultur tabung menggunakan lampu 75 watt (200 – 300 µmol m-2 s-1 ) yang diukur dengan Qantum meter. Larutan hara yang digunakan untuk memelihara kultur adalah Hyponex merah (25-5-20).
ISOLASI SPORA
Spora CMA diisolasi dari perakaran I. barbatum yang tumbuh di tanah tersebut dengan metode yang dikembangkan oleh International (Vescicular) Arbuscular Mychorrhizae (INVAM). Campuran akar tanaman dan tanah di aduk merata, lalu disaring bertingkat dengan saringan 2mm, 1mm, 500m, 250m, 125m dan 53m. Hasil saringan berupa lapisan hancuran akar dan spora dipisahkan dengan sentrifus gradien sukrosa 20-60% pada kecepatan 960 x g selama 2 – 3 menit. Supernatant disaring dengan kertas saring dan dicuci dengan akuades, lalu spora dikoleksi dengan menggunakan pipet pasteur ujung sangat lancip.
MD1 dan MD2, yang berbeda diameter, bentuk skutelum, dan dinding sporanya.
KULTUR SPORA TUNGGAL DAN UJI EFEKTIVITAS KOLONISASI
Spora yang didapat diuji efektivitasnya mengkolonisasi akar P. javanica melalui kultur spora tunggal media zeolit steril dalam tabung reaksi. Biji P. javanica
disterilsasi permukaan dengan 5% NaOCl dan dikecambahkan pada zeolit steril. Akar benih P. javanica yang berdaun dua, berumur 7 – 10 hari, diinokulasi dengan satu spora yang berasal dari isolat MD1 atau MD2. Selain itu, satu benih lainnya diinokulasi secara bersama (ko-inokulasi) oleh isolat MD1 dan MD2. Masing-masing inokulasi dilakukan terhadap 50 benih tanaman. Benih tersebut dikultur dalam tabung reaksi berukuran 2 cm x 15 cm yang berisi 50 g zeolit steril. Kultur tersebut tanaman dipilih secara acak dari masing-masing perlakuan untuk diamati jaringan hifa pada permukaan akar (persentase infektivitas) dan kolonisasi hifa. Kolonisasi hifa diamati dengan mewarnainya dengan trifan biru. Persentase kolonisasi dan panjang akar terkolonisasi mikoriza pada akar P. javanica dan I. barbatum dihitung dipelihara dengan menyiraminya dengan air sesuai keperluan dan dengan larutan hara
Hyponex merah (1g.L-1) setiap minggu sebanyak 20 mL, sampai berumur dua bulan. Setelah itu jumlah air yang disiramkan dikurangi secara bertahap untuk memicu pembentukan spora sampai tanaman menguning. Kerapatan spora dalam inokulum dihitung secara langsung dengan mengekstrak spora dari 100 g tanah (INVAM).
UJI EFEKTIVITAS ASOSIASI CMA ISOLAT MD1
Untuk mengetahui efektivitas asosiasi isolat MD1 dengan tanaman adalah dengan melihat peranannnya dalam pertumbuhan dan akumulasi logam Cr dan Ni tanaman I. barbatum asal tanah serpentin (IBMD) dan I. barbatum asal tanah non-serpentin (IBLA), serta tanaman penutup tanah, P. javanica. Spora isolat MD1 diisolasi dari inokulum MD1 dengan metode sama seperti percobaan di atas. Sebanyak 10 spora isolat MD1 diinokulasikan pada akar benih tanaman I. bartatum yang tingginya 15 cm dan P. javanica yang berdaun dua. Tanaman itu dikulturkan dan dipelihara sebagaimana kultur spora tunggal. Percobaan ini dilakukan tiga ulangan dengan menggunakan rancangan split-plot dengan tanaman sebagai petak utama dan perlakuan inokulasi serta kontrol sebagai anak petak.
dan Ni dalam jaringan diukur. Pengukuran kandungan logam tersebut dilakukan dengan menggunakan spektrofotometer serapan atom (AAS) di Laboratorium Badan Lingkungan Hidup (BLH), Kabupaten Banjar.
UJI EFEKTIVITAS ASOSIASI
KOMBINASI ISOLAT MD1 DAN MD2
Uji efektivitas kolonisasi ko-inokulasi isolat MD1 dan MD2 menunjukan inokulasi tersebut menghasilkan efekvitas kolonisasi yang lebih baik daripada inokulasi tunggal masing-masing isolat. Pengujian ini menggunakan metode yang dimodifikasi dari metode trapping untuk isolasi spora. Sebanyak 80 g zeolit steril diletakan di dasar pot berukuran 220 ml, lalu ditambahkan 80 g inokulum MD1+2 (berisi isolat MD1 dan MD2) yang mengandung sekitar 100 spora, kemudian diletakan benih tanaman yang sama seperti uji efektivitas isolat MD1, dan terakkhir ditutup dengan zeolit steril 60 g. Perlakuan lainnya adalah
inokulasi dengan inokulum ―Techno‖, yang
berisi spora berbagai spesies CMA, serta inokulasi dengan spora CMA isolat LA yang diisolasi dari perakaran I. barbartum asal tanah spodosol (non-serpentin) Lianganggang. Tanaman diletakan di rumah kaca dan dipelihara sama dengan percobaan sebelumnya. Setelah berumur 1 bulan, tanaman dipindah ke pot berukuran 660 ml yang telah disterilisasi permukaan dan berisi 500 g tanah serpentin steril. Percobaan menggunakan rancangan split-plot dengen tiga ulangan. Pengamatan yang dilakukan sama dengan yang dilakukan pada percobaan uji efektivitas asosiasi isolat MD.
HASIL DAN PEMBAHASAN
Isolasi spora pada daerah perakaran I. barbatum yang tumbuh di tanah serpentin Mandiangin mendapatkan dua macam spora yang dikelompokan menjadi dua isolat
yakni MD1 dan MD2. Kedua isolat tersebut mempunyai karakter fisik yang berbeda (Gambar 1). Spora isolat MD1 berukuran 80-135 µm, berwarna coklat kehitaman, berbentuk bulat dengan skutelum kurang tampak dan dinding spora lebih tebal. Spora isolat MD2 berukuran 70-95 µm, berwarna kekuningan, berbentuk lonjong dengan terlihat jelas skutelum dan dinding spora lebih tipis.
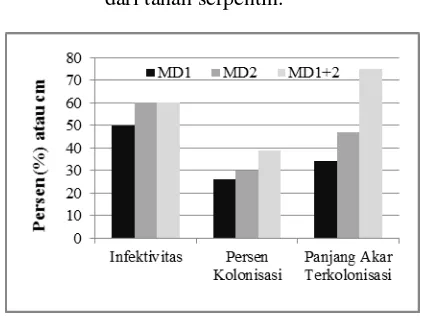
Efektivitas inokulasi suatu isolat dapat dinilai dari infektivitas dan derajat kolonisasi (persen kolonisasi dan panjang akar terkolonisasi) akar tanaman inang. Infektivitas isolat MD1 dan MD2 berkisar 50 % merupakan nilai yang setara dengan infektivitas inokulum mikoriza komersial.
Isolat MD1 Isolat MD2
Gambar 4. Spora cendawan mikoriza arbuskuler isolat MD1 dan MD2 yang diisolasi dari tanah serpentin.
Uji efektivitas inokulasi menunjukan inokulasi secara bersama (ko-inokulasi) oleh isolat MD1 dan MD2 menghasilkan persen kolonisasi dan panjang akar terkolonisasi yang lebih baik daripada inokulasi oleh isolat tunggalnya. Kedua isolat tersebut berinteraksi secara sineristik dalam mengkolonisasi akar I. barbatum, yang mana kondisi ini juga kemungkinannya terjadi di alam. Interaksi sinerggisitik ini menjadi hal yang sangat penting bagi CMA untuk meningkatkan biomasa dan daya tahan hidup bagi tanaman di lahan tambang emas. Tanaman yang diinfeksi secara bersama oleh Glomus versiforme dan Paraglomus occultum mempunyai luas daun spesifik, rasio area daun, dan volume akar yang lebih baik daripada tanaman yang diinokulasi oleh salah satu CMA tersebut. Inokulasi sinergistik juga bisa terjadi antara CMA dengan berbagai fungi dan bakteri termasuk fungi safrofitik. Antagonistik antar isolat dalam mengeksploitasi nutrisi tidak ditemui pada tipe strategi kolonisasi Glomus namun ditemui beberapa pada tipe Gigaspora. Gambar 1 mengindikasikan bahwa kedua isolat berasal dari genera Glomus atau Paraglomus.
Efektivitas inokulasi isolat MD1 terhadap tanaman I. barbatum asal tanah serpentin (IBMD), I. barbatum asal tanah non-serpentin (IBLA), dan P. javannica tidak menunjukan perbedaan yang signifikan. Infektivitas isolat MD1 pada ketiga tanaman berkisar 70 sampai 90 %, yang tampak lebih tinggi dibandingkan dengan percobaan uji efektivitas kolonisasi (Gambar 2). Perbedaan ini karena jumlah spora yang digunakan untuk inokulasi berbeda.
Inokulasi isolat MD1 secara signifikan mampu meningkatkan berat kering tajuk tanaman pada I. barbatum asal tanah non-serpentin (IBLA) namun tidak mampu pada dua tanaman lainnya. (Gambar 3). Hal ini menunjukan isolat MD1 efektif berasosiasi dengan IBLA namun tidak terjadi pada
tanaman lainnya, sedangkan inokulum
―Techno‖ bisa efektif berasosiasi dengan
tanaman PJ dan IBLA. Hasil tersebut mengindikasikan dua hal, pertama asosiasi antara tanaman IBMD dengan isolat MD1 tidak memberikan kontribusi pada pertumbuhan. Tanaman IBMD merupakan tanaman yang sudah beradaptasi dengan lingkungan serpentin yang rendah rasio Ca-Mg dan toksisitas logam beratnya tinggi, sehingga dalam kondisi kultur yang nutrisi yang mencukupi peranan mikoriza menjadi berkurang. Efektivitas asosiasi (mikoriza) tergantung kepada ketersediaan hara, terutama unsur fosfor (P). Tanaman IBLA adalah tanaman non-serpentin yang belum beradaptasi dengan lingkungan serpentin sehingga tetap memerlukan mikoriza untuk mengatasi efek negatif rendahnya rasio Ca-Mg dan mendetoksifikasi logam berat. CMA mengsekresi protein Glomalin yang dapat mengkelat logam tersebut sehingga menjadi tidak toksik. Kedua, tanaman P. javanica (PJ) tidak menunjukan respon positif terhadap inokulasi satu jenis CMA (isolat MD1) namun sangat signifikan terhadap inokulum multi spesies seperti
―Techno‖. Hal ini mengindikasikan bahwa
rendahnya spesifisitas antara PJ dengan isolat MD1.
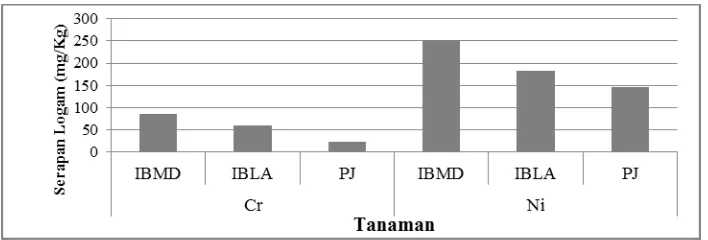
Tanaman yang diinokulasi dengan CMA mengandung logam berat Cr dan Ni konsentrasi yang lebih tinggi daripada tanaman kontrol (Gambar 4). Mikoriza yang terbentuk oleh inokulasi tersebut membentuk jaringan hifa yang sangat luas sehingga memungkinkan penyerapan logam berat dari volume tanah yang lebih besar. Tanaman bermikoriza menyerap logam berat tersebut melalui protein transporter logam spesifik atau non-spesifik yang terdapat pada akar maupun hifa lalu menyimpan dalam vakuola atau mentransportasikannya melalui hifa masuk ke korteks akar, kemudian dapat diedarkan melalui sistem pembuluh. Inokulum
―Techno‖ mengandung spora yang bukan
kemungkinannya spora tersebut tidak toleran terhadap logam berat. Namun pada percobaan ini, tanaman diinokulasi dulu dengan CMA pada media zeolit setelah kolonisasi terbentuk lalu dipindah ke tanah serpentin, sehingga CMA sudah tidak sensitif lagi terhadap logam. Spora CMA
dapat mentoleransi dengan kehadiran logam berat namun sifat toleransi ini merupakan plastisitas fenotipe bukan karena perubahan genetik, karena sifat itu akan hilang pada generasi berikutnya ketika logam berat tidak ada.
Gambar 6. Berat kering tajuk tanaman I. barbatum asal tanah serpentin (IBMD), I. barbatum asal tanah non-serpentin (IBLA), dan P. javanica yang diinokulasi dengan isolat MD1, inokulum Techno, dan tanpa inokulasi (K). Keterangan: Kolom yang diikuti oleh huruf yang sama, tidak berbeda nyata dengan uji BNT (LSD) pada taraf 0.05%; Bar pada kolom menunjukkan standar deviasi.
Gambar 7. Kandungan logam berat kromium (Cr) dan nikel (Ni) pada jaringan berbagai tanaman yang diinokulasi dengan isolat MD1, inokulum Techno, dan Kontrol (tanpa inokulasi).
Logam berat seperti Cr dan Ni dapat bersifat toksik bagi tanaman namun beberapa tanaman tertentu diketahui bisa mengakumulasi logam berat. Kemampuan tersebut bisa dibantu oleh inokulasi CMA yang dapat meningkatkan metabolisme asam amino prolin sehingga bisa melindungi dari logam berat kromium. Tanaman I. barbatum asal tanah serpentin (IBMD) mengakumulasi logam Cr dan Ni lebih tinggi daripada tanaman lainnya (Gambar 5). Kemampuan tanaman IBMD mengakumulasi logam tersebut tampaknya tidaklah semata-mata karena adanya mikoriza tapi adanya mekanisme seluler yang diatur secara genetik atau plastisitas fenotipe. Pertama adalah induksi asam organik, seperti asam oksalat, dan protein pengkelat logam, seperti fitokelatin dan metallotionien, yang mengkelat logam tersebut di vakuola. Kedua adalah induksi protein kejutan panas (heat-shock) yang dapat melindungi membran dan protein dari kerusakan seperti yang ditimbulkan kejutan panas. Protein pengkelat dan kejutan panas diatur oleh gen tertentu. Gen CAD pada
Arabidopsis menyandikan protein PC sintase yang terlibat dalan sintesis fitokelatin. Selain itu Arabidopsis memiliki gen MT1 dan MT2 yang menyandikan gen metallotionien. Tanaman IBLA mampu mengakumulasi lebih baik daripada PJ, hal ini mengindikasikan bahwa
I. barbatum mempunyai daya adaptasi yang lebih baik daripada tanaman PJ. Untuk beradaptasi itu tanaman mengembangkan jaringan kompleks mekanisme homeostatis yang sangat efektif untuk mengendalikan masukan (uptake), akumulasi, peredaran, dan detoksifikasi logam.
Untuk mengkonfirmasi bahwa asosiasi CMA isolat MD1 dengan tanaman I. barbatum asal tanah serpentin (IBMD) adalah tidak efektif seperti pada uji efektivitas asosiasi isolat MD1, maka tanaman tersebut dan dan P.javanica
diinokulasi dengan inokulum MD1+2 yang mengandung spora isolat MD1 dan MD2,
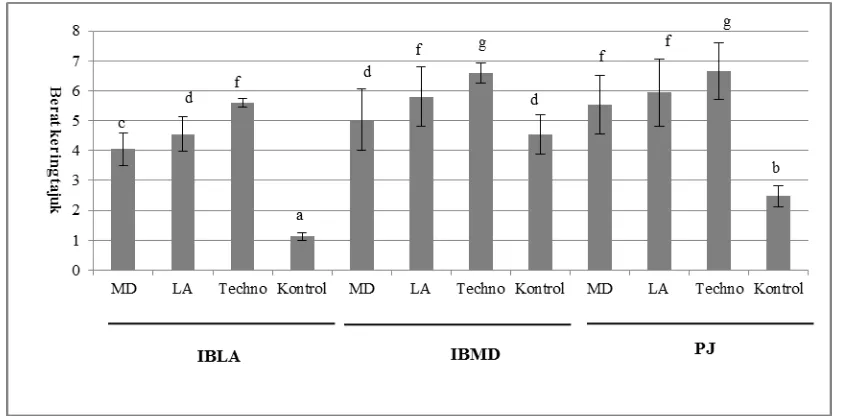
inokulum LA yang mengandung spora CMA dari perakaran I. barbatum tanah non-serpentin, dan inokulum ―Techno‖. Hasil percobaan menunjukan bahwa semua tanaman yang diinokulasi mempunyai berat kering yang nyata lebih tinggi daripada tanaman kontrol kecuali tanaman IBMD yang diisolasi dengan isolat MD (Gambar 6). Hal ini karena tingginya berat kering tanaman kontrol yang mengindikasikan bahwa tanaman tersebut tidak tergantung pada mikoriza dalam memacu pertumbuhan sedangkan tanaman lainnya adalah tergantung pada mikoriza. Tanaman PJ yang diinokulasi CMA mempunyai berat kering yang nyata lebih tinggi daripada tanaman kontrol. Ini berarti bahwa tanaman merespon baik kolonisasi inokulum MD yang berisi isolat MD1 dan MD2, hal ini berbeda dengan percobaan sebelumnya (Gambar 3). Hasil ini memperkuat indikasi bahwa tanaman PJ mempunyai efektitivitas asosiasi yang spesifik terhadap isolat atau spesies CMA. Adanya respon pada tanaman yang spesifik untuk spesies CMA juga diamati pada rumput-rumputan. Respon spesifik-spesies tersebut kemungkinannya dimediasi oleh kombinasi kompleks sinyal tanaman dan CMA oleh bahan yang dikendalikan secara genetik.
Gambar 9. Berat kering tajuk tanaman I. barbatum asal tanah serpentin (IBMD), I. barbatum asal tanah non-serpentin (IBLA), dan P. javanica yang diinokulasi oleh CMA isolat MD1+2 (MD), CMA isolat asal tanah non-serpentin (LA) dan inokulum ―Techno‖, serta kontrol. Keterangan: Kolom yang diikuti oleh huruf yang sama, tidak berbeda nyata dengan uji BNT (LSD) pada taraf 0.05%; Bar pada kolom menunjukkan standar deviasi.
KESIMPULAN
Asosiasi yang efektif antara cendawan mikoriza arbuskuler dengan tanaman I. barbatum dan P. javanica mampu meningkatkan berat kering tanaman dan juga serapan logam berat kromium (Cr) dan nikel (Ni) yang ditanam di tanah serpentin. CMA non-serpentin dapat membentuk asosiasi yang efektif dengan tanaman di tanah serpentin setelah kolonisasi terbentuk. Interaksi antara CMA dengan jenis tanaman mengindikasikan adanya respon spesfik-spesies, untuk itu perlu ditelaah efektivitas inokulasi dan assosiasi isolat MD2 dengan tanaman pada tanah serpentin.
UCAPAN TERIMA KASIH
Tulisan ini berdasarkan sebagian data dari Penelitian Fundamental dari ketiga penulis di atas yang didanai oleh Dikti pada tahun 20009 dan 2010. Penulis berterima kasih dan memberikan penghargaan
setinggi-tingginya kepada Dwi Purno Widekdo yang membantu dalam isolasi isolat MD1 dan MD1 serta pembuatan inokulum, Hidayatun Mutia untuk isolasi spora LA, Adityawarman untuk menguji isolat MD1, dan Deviana Ayu Aresma untuk menguji inokulum MD1+MD2. Penulis juga mengucapkan terima kasih kepada Kepala Sub-laboratorium Biologi beserta seluruh staf yang telah mendukung dan memfasilitasi kegiatan penelitian ini.
DAFTAR PUSTAKA
Ross, S., (1994). Sources and Forms of Potentially Toxic Metals in Soil-Plant System. Brisbane: John Wiley & Sons.
Brady, K.U., A.R. Kruckeberg, and H.D. Bradshaw.J, (2005). Evolutionary Ecology of Plant Adaptation to Serpentine Soils. Annual Review of Ecology, Evolution, and Systematics, Vol. 36, issue p. 243-266. phytoremediation of arsenic contaminated land. Environ Pollut, Vol. 118, issue 3, p. 453-61
Odjegba, V.J. and I.O. Fasidi, (2004). Accumulation of trace elements by Pistia stratiotes: implications for phytoremediation. Ecotoxicology, Vol. 13, issue 7, p. 637-46.
Hall, J.L., (2002). Cellular mechanisms for heavy metal detoxification and tolerance. J Exp Bot, Vol. 53, issue 366, p. 1-11.
Norliana. (2007). Kolonisasi mikoriza pada rumput Ischaemum barbatum Retz yang tumbuh pada berbagai vegetasi di tanah serpentin.Skripsi Program Studi Biologi Lambung Mangkurat, Banjarbaru.
Orlowska, E., D. Orlowski, J. Mesjasz-Przybylowicz, and K. Turnau, (2011). Role of mycorrhizal colonization in plant establishment on an alkaline gold mine tailing.Int J Phytoremediation, Vol. 13, issue 2, p. 185-205.
Bradshaw, H.D., Jr., (2005). Mutations in CAX1 produce phenotypes characteristic of plants tolerant to serpentine soils.New Phytol, Vol. 167, issue 1, p. 81-8.
Mansur, I. (2000). Diversity of rhizobia nodulating the tree legumes Acacia mangium and Paraserianthes falcataria and their interaction with arbuscular
mycorrizal fungi in young seedling.PhD Dissertation University of Kent at Canterbury, Kent.
Phillips, J.M. and D.S. Hayman, (1970). Improved procedures for clearing roots and staining parasitic and vesiculararbuscular mycorrhizal fungi for rapid assessment of infection. .Trans Br. Mycol. Soc., Vol. 55, issue p. 158-161.
Giovannetti, M. and B. Mosse, (1980). An Evolution of Techniques to Measure VAM Infection in Roots New Phytologist Vol. 84, issue 489-500.
Brundrett, M.C., L. Melville, and L. Peterson, (1994). Practical Methods in Mycorrhiza Research. . Ontario, Canada. : Mycologue Publications.
Biermann, B., (1981). Quantifying vesicular-arbuscular mychorrizhae : proposed method toward standardization. New Phytologist, Vol. 87, issue p. 63-67.
Corkidi, L., B. Allen, D. Merhaut, M. Allen, J. Downer, J. Bohn, and M. Evans, (2004). Assessing the Infectivity of Commercial Mycorrhizal Inoculants in Plant Nursery Conditions. J. Environ. Hort. , Vol. 22 issue 3, p. 149-154.
Tian, Y., Y. Lei, Y. Zheng, and Z. Cai, (2013). Synergistic effect of colonization with arbuscular mycorrhizal fungi improves growth and drought tolerance of Plukenetia volubilis seedlings. Acta Physiologiae Plantarum, Vol. 35, issue March 2013, 3, p. 687-696
Cano, C. and A. Bago, (2005). Competition and substrate colonization strategies of three polyxenically grown arbuscular mycorrhizal fungi. Mycologia, Vol. 97, issue 6, p. 1201-14.
Gohre, V. and U. Paszkowski, (2006). Contribution of the arbuscular mycorrhizal symbiosis to heavy metal phytoremediation. Planta, Vol. 223, issue 6, p. 1115-22.
Taylor, D.L., T.D. Bruns, J.R. Leake, and D.J. Read, (2002).Mycorrhizal Specificity and Function in Myco-heterotrophic Plants, in Mycorrhizal Ecology, M.G.A. van der Heijden and I. Sanders, Editors. Springer-Verlag Berlin Heidelberg. p. 375-411.
Shalaby, A.M., (2003). Responses of Arbuscular Mycorrhizal Fungal Spores Isolated from Heavy Metal-polluted and Unpolluted Soil to Zn, Cd, Pb and Their Interactions in vitro.Pakistan Journal of Biological Sciences, Vol. 6 issue 16 p. 1416-1422.
Memon, A.R., D. Aktoprakligul, A. Zdemur, and A. Vertii, (2001). Heavy Metal Accumulation and Detoxification Mechanisms in Plants. Turk J Bot, Vol. 25, issue p. 111-121.
Ruscitti, M., M. Arango, M. Ronco, and J. Beltrano, (2011). Inoculation with mycorrhizal fungi modifies proline metabolism and increases chromium tolerance in pepper plants (Capsicum annuum L.). Brazilian Society of Plant Physiology, Vol. 23, issue 1, p. 15-25.
Yang, Z. and C. Chu, (2011).Towards Understanding Plant Response to Heavy Metal Stress, in Towards Understanding Plant Response to Heavy Metal Stress, Abiotic Stress in Plants - Mechanisms and Adaptations,, A. Shanker, Editor. InTech Europe: Rijeka, Croatia.
Wilson, G.W.T. and D.C. Hartnett, (1998). Interspecific variation in plant responses to mycorrhizal colonization in tallgrass prairie. American Journal of Botany, Vol. 85, issue 12, p. 1732-1738.
Gianinazzi-Pears, V., S. Gianinazi, J.P. Guillemian, A. Trouvelot, and G. Duc, (1991).Genetic and cellular analysis of resistance of vesicular arbuscular (VA) mycorrhizal fungi in pea mutants., in