i
IDENTIFIKASI ALEL GEN SITOKROM P450 2A6*9 PADA SUBJEK UJI NONPEROKOK RAS KULIT HITAM PAPUA INDONESIA DENGAN
METODE POLYMERASE CHAIN REACTION
SKRIPSI
Diajukan untuk Memenuhi Salah Satu Syarat Memperoleh Gelar Sarjana Farmasi (S. Farm)
Program Studi Farmasi
Oleh :
Mensiana Lodvi Putarato
NIM : 168114014
FAKULTAS FARMASI
UNIVERSITAS SANATA DHARMA YOGYAKARTA
ii
IDENTIFIKASI ALEL GEN SITOKROM P450 2A6*9 PADA SUBJEK UJI NONPEROKOK RAS KULIT HITAM PAPUA INDONESIA DENGAN
METODE POLYMERASE CHAIN REACTION
SKRIPSI
Diajukan untuk Memenuhi Salah Satu Syarat Memperoleh Gelar Sarjana Farmasi (S. Farm)
Program Studi Farmasi
Oleh :
Mensiana Lodvi Putarato
NIM : 168114014
FAKULTAS FARMASI
UNIVERSITAS SANATA DHARMA YOGYAKARTA
v
HALAMAN PERSEMBAHAN
Saya persembahkan skripsi ini untuk : Tuhan Yesus
Bapak Victor Ngongo Putarato, Mama Dorkas Lende, Endo Putarato, Alm. Rian Putarato, Ney Putarato dan Ona.
Keluarga Besar Loli dan Wewewa.
Sahabat dan teman-teman saya tercinta, serta Almamater saya.
viii PRAKATA
Puji dan syukur ke hadirat Tuhan yang Maha Esa karena atas berkat, kasih dan penyertaan-Nya penulis dapat menyelesaikan skripsi dengan judul “Identifikasi Alel Gen Sitokrom P450 2A6*9 pada Subjek Uji Nonperokok Ras Kulit Hitam Papua Indonesia dengan Metode Polymerase Chain Reaction” untuk memenuhi persyaratan memperoleh gelar Sarjana Farmasi (S.Farm) Program Studi Farmasi Universitas Sanata Dharma. Skripsi ini merupakan bagian dari penelitian Dr. Christine Patramurti, Apt., dengan judul “Identifikasi Gen CYP2A6 Alel *4, *7, dan *9 pada Subjek Uji Suku Tionghoa dan Ras Kulit Hitam Papua di Indonesia dengan Metode Polymerase Chain Reaction” .
Selama proses penyusunan skripsi ini tidak lepas dari dukungan dan bantuan dari berbagai pihak. Pada kesempatan ini penulis ingin menyampaikan terima kasih kepada:
1. Ibu Dr. Yustina Sri Hartini, Apt., selaku Dekan Fakultas Farmasi Universitas Sanata Dharma, Yogyakarta.
2. Ibu Dr. Christine Patramurti, Apt., selaku Ketua Program Studi Fakultas Farmasi Universitas Sanata Dharma, Yogyakarta dan sekaligus dosen pembimbing atas segala kesabaran, bimbingan, ilmu, semangat, nasihat serta dukungan kepada penulis.
3. Ibu Damiana Sapta Candrasari, M.Sc., selaku Kepala Laboratorium Fakultas Farmasi Universitas Sanata Dharma dan juga sebagai dosen penguji yang telah memberi semangat, kritik serta saran yang sangat membangun dalam penelitian ini.
4. Bapak Maywan Hariono Ph.D., Apt., selaku dosen penguji yang telah memberi semangat, kritik dan saran yang sangat membangun dalam penelitian ini.
5. Ibu Dr. Dewi Setyaningsih, Apt., selaku Dosen Pembimbing Akademik (DPA) atas bimbingannya selama ini.
ix
7. Segenap dosen Fakultas Farmasi yang dengan sabar dan tulus membagikan ilmu pengetahuan kepada penulis serta karyawan Fakultas Farmasi Universitas Sanata Dharma, Yogyakarta atas pelayanan dan bantuannya selama ini.
8. Bapak Victor Ngongo Putarato, Mama Dorkas Lende serta adik-adik penulis Yuninto Medvi Putarato, Alm. Aprianto Nidvi Putarato, Nerliyati Radvi Putarato, Ona Rostanti Pati Iwu, Elisabet Sarinto Ina Kii, dan Devandi Giovani Porta Lero atas cinta kasih, motivasi, dukungan dan doa yang terus membuat penulis kuat dan bersemangat dalam menggapai segala harapan dan cita-cita. Terkhusus untuk satu-satunya adik laki-laki penulis Alm. Aprianto Nidvi Putarato terima kasih pernah hadir dan membuat hidup penulis berwarna serta selalu bersyukur atas kebaikan Tuhan dalam keluarga kami “Terang Indah” dan seluruh keluarga besar kami.
9. Alm. Opa Dato Toda dan Oma Ester Waingu Bella (Ina Ngongo), Alm. Opa Matius Lende Domba, dan Almh. Oma Apliana Peda Daido, serta Opa Benyamin Bora Busha dan Oma Lidia K. Wolla atas segala perhatian, cinta kasih dan dukungan doa yang selalu menguatkan penulis sehingga penulis dapat melangkah sejauh ini. Terkhusus untuk Alm. Opa Dato Toda yang telah berpulang sebelum penulis lahir, melalui cerita Bapak dan keluarga penulis tahu Opa sangat menyayangi penulis dan cucu-cucu Opa yang lain, serta Alm. Opa Matius Lende Domba dan Almh. Oma Apliana Peda Daido terima kasih untuk segala kasih sayang dan kenangan dimasa kecil yang pernah Opa dan Oma berikan kepada penulis.
10. Keluarga besar Loli terkhusus Keretoma dan Kurutepe serta keluarga besar Wewewa terkhusus Pantendabola yang selalu mendoakan dan memberikan semangat serta dukungan kepada penulis.
11. Sahabat-sahabat penulis “LORY” yakni Resita Wedu, Ocha Valentine dan Yani Etna yang selalu setia dan memotivasi penulis untuk terus berkembang serta selalu menjadi sahabat berbagi cerita baik dalam suka dan duka. 12. Wiwy, Indah Gery Cho, Rani Langkeru, Agatha Maia, dan Ayu P.D. yang
xi
DAFTAR ISI
HALAMAN SAMPUL ... i
HALAMAN JUDUL ... ii
HALAMAN PERSETUJUAN PEMBIMBING ... iii
HALAMAN PENGESAHAN ... iv
HALAMAN PERSEMBAHAN ... v
PERNYATAAN KEASLIAN KARYA ... vi
PERNYATAAN PERSETUJUAN PUBLIKASI ... vii
PRAKATA ... viii
DAFTAR ISI ... xi
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xiii
DAFTAR LAMPIRAN ... xiv
ABSTRAK ... xv
ABSTRACT ... xvi
PENDAHULUAN ... 1
METODE PENELITIAN ... 3
HASIL DAN PEMBAHASAN ... 7
KESIMPULAN ... 13
SARAN ... 14
DAFTAR PUSTAKA ... 15
LAMPIRAN ... 18
xii
DAFTAR TABEL
xiii
DAFTAR GAMBAR
Gambar 1. Hasil analisis kemurnian isolat DNA... 9
Gambar 2. Situs penempelan primer pada sekuen alel gen CYP2A6*1 ... 10
Gambar 3. Urutan nukleotida alel gen CYP2A6*1 dan alel gen CYP2A6*9 ... 11
xiv
DAFTAR LAMPIRAN
Lampiran 1. Ethical Clearance... 19
Lampiran 2. Perhitungan Jumlah Sampel ... 20
Lampiran 3. Hasil Kuesioner ... 21
Lampiran 4. Data Subjek Uji Hasil Kuesioner ... 24
Lampiran 5. Usage Information of FavorPrepTM ... 25
Lampiran 6. Go Taq Green Master Mix Certificate of Analysis ... 27
Lampiran 7. Usage Information of Go Taq Green Master Mix... 28
Lampiran 8. Product Information of DNA Ladder ... 29
Lampiran 9. Product Datasheet of primer forward and reverse ... 30
Lampiran 10. Hasil Analisis Kemurnian Isolat DNA ... 31
Lampiran 11. Hasil Identifikasi Alel Gen CYP2A6*1 dan *9 ... 31
xv ABSTRAK
Tobacco-specific N-nitrosamines (TSNA) seperti 4-(metilnitrosamino)-1-(3-piridil)-1-butanon (NNK) dan N-nitrosonornikotin (NNN) merupakan senyawa prokarsinogen yang terdapat dalam asap rokok. TSNA perlu diaktivasi untuk dapat menjadi senyawa karsinogen yang menyebabkan kanker. Enzim sitokrom P450 2A6 (CYP2A6) merupakan salah satu enzim yang mengaktivasi TSNA yang diekspresikan oleh gen CYP2A6 dan diketahui memiliki polimorfisme yang tinggi. Salah satunya adalah alel gen CYP2A6*9 yang mengalami single nucleotide polymorphism (SNP) dalam kotak TATA di daerah promotor (T-48G). Polimorfisme CYP2A6*9 telah dilaporkan terjadi pada populasi Afrika berkulit hitam di Ghana yang memiliki kesamaan ras dengan ras kulit hitam Papua Indonesia, yaitu ras Negroid. Penelitian ini adalah studi deskriptif observasional dengan tujuan untuk mengidentifikasi keberadaan CYP2A6*9 dan frekuensi CYP2A6*9 pada subjek uji nonperokok ras kulit hitam Papua Indonesia. Subjek uji berjumlah 30 orang dengan kriteria inklusi dan eksklusi yang diambil sampel darahnya lalu dilakukan isolasi DNA. Polimorfisme pada CYP2A6*9 diidentifikasi dengan menggunakan metode Polymerase Chain Reaction (PCR). Hasil penelitian menunjukkan bahwa tidak ditemukan alel gen CYP2A6*9 dari 30 subjek uji, sehingga enzim CYP2A6 akan bekerja secara normal dalam proses aktivasi metabolisme TSNA.
Kata Kunci: CYP2A6*9, polimorfisme, TSNA, nonperokok, ras kulit hitam Papua, PCR.
xvi ABSTRACT
Tobacco-specific N-nitrosamines (TSNA) such as 4-(metilnitrosamino)-1-(3-piridil)-1-butanon (NNK) and N-nitrosonornikotin (NNN) are procarcinogenic compounds found in cigarette smoke. TSNA needs to be activated to change it into carcinogen that causes cancer. Cytochrome P450 2A6 (CYP2A6) is an enzyme that activates TSNAs expressed by the CYP2A6 gene and known to have a high polymorphism. One of them is the CYP2A6*9 gene allele which experiences Single Nucleotide Polymorphism (SNP) in the TATA box of the promoter region (T-48G). The CYP2A6*9 polymorphism has been reported in black African populations in Ghana who have racial similarities to the black Papuan, the Negroid race. This is an observational descriptive study with the aim to identify the presence of CYP2A6*9 and the frequency of CYP2A6*9 in black non-smoker Papua Indonesia. Thirty people are involved as subjects in this study with inclusion and exclusion criteria. Blood samples were collected and then DNA furtherly isolated. Polymorphisms in CYP2A6*9 were identified using the Polymerase Chain Reaction (PCR) method. The results showed that there were no CYP2A6*9 gene alleles from 30 test subjects, so the CYP2A6 enzyme would work normally in the process of TSNA metabolism activation.
1 PENDAHULUAN
Polusi asap rokok merupakan salah satu masalah yang sedang diselesaikan pemerintah Indonesia. Hal ini dilakukan dengan mengeluarkan Undang-undang No. 36 tahun 2009 tentang Kesehatan pada pasal 115 yang mengatur tentang larangan merokok di tempat umum atau Kawasan Tanpa Rokok (KTR) dan menyediakan area khusus untuk merokok (Suryantisa, 2018). Kota Yogyakarta merupakan salah satu daerah yang telah menerapkan KTR dengan mengeluarkan Peraturan Daerah Kota Yogyakarta No. 2 tahun 2017 tentang KTR untuk mengatasi polusi asap rokok. Masalah polusi asap rokok penting untuk diselesaikan karena asap rokok menyebabkan pencemaran udara yang dapat dihirup oleh perokok itu sendiri dan juga perokok pasif. Perokok pasif adalah orang yang bukan perokok tetapi menghirup asap rokok orang lain atau orang yang berada dalam satu ruangan tertutup dengan orang yang sedang merokok (Kementerian Kesehatan RI, 2019). Asap rokok yang dihirup oleh perokok pasif jauh lebih berbahaya bagi kesehatan karena berisiko terkena berbagai penyakit seperti serangan jantung atau stroke dan kanker paru-paru (Suryantisa, 2018).
Asap rokok merupakan aerosol kompleks yang mengandung sekitar 4000 senyawa toksik dan karsinogenik (Ashley et al., 2010; Geiss and Kotzias, 2007; Suryantisa, 2018). Asap rokok mengandung Tobacco-specific nitrosamines (TSNA) (IARC, 2004) seperti 4-(metilnitrosamino)-1-(3-piridil)-1-butanon (NNK) dan N-nitrosonornikotin (NNN) (Ashley et al., 2010) yang merupakan senyawa karsinogen bagi manusia apabila dimetabolisme aktivasi oleh suatu enzim, namun apabila tidak teraktivasi maka senyawa ini bersifat prokarsinogen (Geiss and Kotzias, 2007; IARC 2007). Sitokrom P450 2A6 (CYP2A6) merupakan salah satu enzim utama yang mengaktifkan TSNA seperti NNN dan NNK terutama dalam asap rokok (Liu et al., 2013; Mwenifumbo et al., 2008; Raunio and Rahnasto-Rilla, 2012) dan NNAL yang merupakan hasil metabolik dari NNK. Aktivasi NNK, NNN dan NNAL terjadi melalui reaksi α-hidroksilasi yang menghasilkan senyawa dengan ion diazonium yang dapat terikat pada DNA sehingga dapat membentuk DNA adduct yang menyebabkan kanker pada jaringan yang terpapar asap rokok seperti paru-paru dan laring (Chiang et al., 2011; Hecht
2
and Hoffmann, 1989; Raunio and Rahnasto-Rilla, 2012). Selain senyawa TSNA, enzim CYP2A6 juga berperan dalam memetabolisme obat-obatan dan bahan kimia (Alsanosi et al., 2014; Fujieda et al., 2004). Enzim CYP2A6 merupakan enzim yang dikode oleh gen CYP2A6 (Rossini et al., 2008). Gen CYP2A6 telah dilaporkan banyak mengalami polimorfisme (Raunio and Rahnasto-Rilla, 2012).
Polimorfisme pada CYP2A6 menyebabkan kerja enzim menjadi menurun, meningkat atau bahkan menghilang (Mwenifumbo et al., 2010; Raunio and Rahnasto-Rilla, 2012). Hingga saat ini telah diidentifikasi 89 jenis alel CYP2A6 (PharmVAr, 2012). Alel gen CYP2A6*1 merupakan bentuk normal atau wild type dari gen CYP2A6, sedangkan sebagian lainnya merupakan bentuk yang mengalami polimorfisme. Menurut Minematsu et al. (2006), polimorfisme gen CYP2A6 yang umum pada populasi Asia adalah variasi alel gen CYP2A6 *4,*7 dan *9. Pada alel gen CYP2A6*9 terjadi single nucleotide polymorphism (SNP) dalam kotak TATA di daerah promotor (T-48G), yang dapat menurunkan aktivitas transkripsi dan metabolik in vivo dari CYP2A6 (Minematsu et al., 2006; Yoshida et al., 2003). Hal ini menyebabkan penurunan aktivasi senyawa TSNA yang berpotensi mengurangi risiko kanker paru-paru (Liu et al., 2013; Raunio and Rahnasto-Rilla, 2012).
Alel gen CYP2A6*9 dapat ditemukan pada populasi maupun ras tertentu dalam jumlah frekuensi yang berbeda. Pada populasi Afrika berkulit hitam di Ghana ditemukan alel gen CYP2A6*9 dengan frekuensi 5,7% (Gyamfi et al., 2005). Populasi Afrika di Ghana ini memiliki kesamaan ras dengan ras kulit hitam Papua Indonesia yaitu ras Negroid (Gayanti, 2017). Hal ini menunjukkan terdapat kemungkinan populasi Papua di Indonesia juga memiliki polimorfisme CYP2A6*9. Penelitian yang bertujuan untuk mengidentifikasi CYP2A6*9 telah dilakukan oleh Prabowo (2017) pada subjek uji perokok ras kulit hitam Papua Indonesia dan didapatkan frekuensi alel gen CYP2A6*9 adalah 0%. Oleh karena itu, perlu dilakukan identifikasi alel gen CYP2A6*9 pada subjek uji yang berbeda yaitu nonperokok untuk mengetahui lebih lanjut mengenai frekuensi polimorfisme alel gen CYP2A6*9 pada ras kulit hitam Papua Indonesia.
3
Polimorfisme pada alel gen CYP2A6*9 ini dapat dideteksi dengan metode Polymerase Chain Reaction (PCR). PCR adalah metode yang dapat melipatgandakan (amplifikasi) potongan DNA dalam waktu singkat (Bintang, 2018). Menurut Bintang (2018) dan Querci et al. (2006) metode PCR terdiri atas 3 tahapan yaitu: pemisahan untai DNA (denaturasi), annealing dan sintesis DNA (extension).
METODE PENELITIAN Bahan Penelitian
Sampel darah subjek uji nonperokok ras kulit hitam Papua, Primer forward: GATTCCTCTCCCCTGGAAC-3’) dan primer reverse: (5’-GGCTGGGGTGGTTTGCCTTTA-3’) (Yoshida et al., 2003), Promega Go Taq Green Master Mix (yang mengandung taq DNA polymerase, dNTPs, MgCl2, dan buffer), FavorPrepTM Genomic DNA Mini Kit (Blood/Cultured Cell), 10X Tris-Borate-EDTA (TBE) Buffer pH 8,3 Ultra Pure Grade (Vivantis), agarose (Vivantis), FluoroVueTM Nucleic Acid Gel Stain (SMOBIO), 1KB (0.25-10 kb) DNA Ladder (SMOBIO), loading dye, etanol absolut (Sigma Chemical Co., St. Louis), dan Water For Injection (WFI) (Ikapharmindo).
Alat Penelitian
Alat-alat gelas, Thermal cycler Perkin Elmer 2400, satu set Elektroforesis horizontal (Sigma Aldrich), UV Transilluminator (fisher scientific), dispossable gloves, microtube 1,5 mL, hot plate (Thermo scientific), mikropipet, waterbath, blue tip, yellow tip, white tip, Microcentrifuge (Thermo fisher scientific), kamera Mirrorless Fuji Film XA3, spuit injeksi 3 mL (Terumo), vacutainer K2 yang mengandung EDTA 5,4 mg, PCR® Tubes (Axygen), FABG column, collection tube 2 mL, timbangan analitik (METTLER AE 260), dan vortex.
Pemilihan Subjek Uji
Subjek uji dalam penelitian ini berjumlah 30 orang dengan kriteria yaitu nonperokok, berusia ≥18 tahun, keturunan ras kulit hitam Papua asli minimal sampai third degree relativies (kakek dan nenek orang Papua asli), dan tinggal di Yogyakarta. Pada penelitian ini dilakukan eliminasi pada subjek uji yang sedang
4
terkena penyakit yang disebabkan oleh virus dan bakteri. Subjek uji dengan sadar dan bersedia menandatangani inform consent. Penelitian yang dilakukan di Laboratorium Biokimia Fakultas Farmasi Universitas Sanata Dharma ini telah memenuhi Kode Etik yang disetujui oleh Komisi Etik Fakultas Kedokteran Universitas Kristen Duta Wacana, Yogyakarta.
Pengambilan Sampel Darah pada Subjek Uji
Sampel darah dari subjek uji akan diambil oleh petugas profesional perawat. Sampel darah subjek uji diambil dari pembuluh vena, yang kemudian ditampung dalam vacutainer K2 yang berisi EDTA 5,4 mg. Sampel darah yang diperoleh disimpan pada suhu ± 4°C, sebelum dianalisis. Sampel darah ini kemudian akan digunakan untuk identifikasi alel gen CYP2A6*9.
Isolasi DNA
Sampel darah yang telah disiapkan kemudian diisolasi dengan menggunakan FavorPrepTM Blood Genomic DNA Extraction Mini Kit. Sampel darah diambil sebanyak 200 µL dan dipindahkan ke microtube 1,5 mL. Proteinase K sebanyak 20 µL dan FABG buffer 200 µL ditambahkan ke dalam sampel lalu divortex sebentar. Campuran tersebut kemudian diinkubasi pada suhu 60°C selama 15 menit untuk melisiskan sampel serta pada setiap menit ke-3 divortex selama 30 detik. Setelah selesai diinkubasi campuran tersebut disentrifugasi pada kecepatan 10.000 rpm selama 30 detik. Kemudian ditambahkan 200 µL etanol absolut ke dalam sampel dan divortex selama 30 detik. Campuran tersebut kemudian disentrifugasi pada kecepatan 10.000 rpm selama 30 detik. Kolom FABG disiapkan dan dipasangkan pada collection tube, lalu campuran sampel ditransfer dari microtube 1,5 mL ke dalam kolom FABG yang telah terpasang collection tube. Campuran sampel yang telah dipindahkan disentrifugasi pada kecepatan 13.000 rpm selama 1 menit, lalu cairan yang ada pada collection tube dibuang. Selanjutnya kolom FABG dipasangkan pada collection tube baru. Kolom FABG segera dicuci dengan ditambahkan 500 µL W1 buffer lalu disentrifugasi pada kecepatan 13.000 rpm selama 1,5 menit lalu sisa cairan pada collection tube dibuang. Kolom FABG kemudian dicuci dengan 750 µL wash buffer lalu disentrifugasi pada kecepatan 13.000 rpm selama 1,5 menit lalu sisa cairan pada
5
collection tube dibuang. Selama 4 menit dilakukan sentrifugasi untuk mengeringkan kolom FABG. Kolom FABG yang telah kering dipasangkan pada elution tube dan ditambahkan 200 µL elution buffer pada FABG kolom. Kemudian dibiarkan berdiri diam selama 3 menit dan disentrifugasi selama 3 menit untuk mengelusi DNA pada kecepatan 13.000 rpm. Isolat DNA disimpan pada suhu -70°C.
Analisis Kemurnian Isolat DNA
Analisis kemurnian isolat DNA dapat dilakukan dengan metode elektroforesis yang menggunakan gel agarose 1,0%. Cara pembuatan gel agarose 1,0% yaitu dengan menggunakan 250 mg agarose yang dilarutkan dalam larutan 1x TBE buffer sebanyak 25 mL. Campuran tersebut kemudian dipanaskan dengan hot plate sambil diaduk dengan magnetic stirrer hingga larut dan menjadi jernih. Larutan agarose 1% yang telah jernih diangkat, lalu ditambahkan 2,5 μL larutan Nucleic Acid Gel Stain dan diaduk hingga homogen. Larutan gel agarose yang telah siap, dituang ke dalam cetakan gel yang sebelumnya telah diberi sisiran pada tepi gel dan gel dibiarkan mengeras. Sisiran dicabut setelah gel mengeras sehingga terbentuk sumur-sumur, kemudian ditempatkan ke dalam gel tray elektroforesis yang sudah berisi larutan 1x TBE buffer. Isolat DNA diambil sebanyak 4,0 μL, ditambahkan WFI sebanyak 1,0 μL dan dicampur dengan loading dye sebanyak 1,0 μL. Campuran tersebut diambil sejumlah 6,0 μL dan dimasukkan ke dalam sumur-sumur gel menggunakan mikropipet. Satu sumur gel diisi dengan 1KB (0.25-10 kb) DNA Ladder sebanyak 3,0 μL sebagai marker dan dilakukan elektroforesis dengan kecepatan 100 volt/cm selama 30 menit. Pada pH netral molekul DNA yang bermuatan negatif akan bergerak menuju kutub positif karena adanya pengaruh medan listrik. Hasil elektroforesis dideteksi di bawah UV Transilluminator dan didokumentasikan menggunakan kamera Mirrorless Fuji Film XA3. Panjang pita DNA dapat diketahui dengan cara menarik garis lurus masing-masing pita sampel isolat DNA dengan posisi pita DNA ladder.
Amplifikasi Isolat DNA dengan Metode PCR
Fragmen alel gen CYP2A6*9 diidentifikasi melalui proses amplifikasi menggunakan primer yang diadopsi dari Yoshida et al. (2003), yaitu primer
6
forward: 5'-GAT TCC TCT CCC CTG GAA C-3' dan primer reverse: 5'- GGC TGG GGT GGT TTG CCT TTA-3'. Amplifikasi DNA dilakukan dengan PCR menggunakan bahan yang terdiri atas Promega Go Taq Green Master Mix 12,5 μL, primer forward 1,25 μL, primer reverse 1,25 μL, isolat DNA 5,0 μL, dan WFI 5,0 μL. Kondisi PCR untuk proses amplifikasi yang digunakan didasarkan pada hasil optimasi yang dilakukan oleh Cindy (2017). Amplifikasi dilakukan dengan mesin PCR (Thermal cycler Perkin Elmer 2400). Kondisi PCR diatur sebagai berikut: initial denaturasi pada suhu 94ºC selama 3 menit; dilanjutkan dengan denaturasi pada suhu 94ºC selama 30 detik; annealing pada suhu 60ºC selama 30 detik dan ekstensi pada suhu 70ºC selama 25 detik. Siklus amplifikasi dilakukan sebanyak 35 kali dan diakhiri dengan final ekstensi pada suhu 72ºC selama 5 menit. Hasil produk PCR disimpan pada suhu -70°C.
Identifikasi Alel Gen CYP2A6*9
Hasil produk PCR dapat dideteksi dengan elektroforesis pada kecepatan 100 volt/cm selama 30 menit menggunakan fase diam agarose 1,5% yang ditambahkan dengan Nucleic Acid Gel Stain dan fase gerak larutan 1x TBE buffer. Hasil elektroforesis kemudian dilihat dengan menggunakan lampu UV Transilluminator. Hasil produk PCR ditunjukkan dengan adanya pita yang kemudian didokumentasi dengan kamera Mirrorless Fuji Film XA3. Produk PCR yang terbentuk pada penelitian ini adalah alel gen CYP2A6*1 yang berada pada pita 368 bp (Yoshida et al., 2003).
Analisis Hasil
Untuk mengetahui adanya alel gen CYP2A6*1 dan alel gen CYP2A6*9 dapat dideteksi dengan metode elektroforesis. Adanya produk yang terbentuk menunjukkan alel gen CYP2A6*1, sedangkan jika tidak terbentuk produk maka menunjukkan adanya alel gen CYP2A6*9.
Frekuensi alel gen CYP2A6*1 dan alel gen CYP2A6*9 dapat dihitung dengan rumus:
Frekuensi CYP2A6*1 =
Frekuensi CYP2A6*9 =
7 HASIL DAN PEMBAHASAN
Penelitian ini bertujuan untuk mengidentifikasi keberadaan alel gen CYP2A6*9 pada subjek uji nonperokok ras kulit hitam Papua Indonesia. Penelitian ini dilakukan dalam 4 tahap yaitu pemilihan subjek uji, isolasi DNA, analisis kemurnian isolat DNA, dan identifikasi alel gen CYP2A6*9 pada produk PCR yang diamplifikasi dengan metode PCR. Metode yang digunakan dalam mengidentifikasi alel gen CYP2A6*9 adalah metode PCR yang dapat melipatgandakan (amplifikasi) potongan DNA dalam waktu singkat (Bintang, 2018). CYP2A6*9 merupakan alel yang akan diidentifikasi dalam penelitian ini karena telah dilaporkan terdapat pada beberapa ras dan populasi di dunia.
Pemilihan Subjek Uji
Subjek uji dalam penelitian ini berjumlah 30 orang yang terdiri atas 19 orang laki-laki dan 11 orang perempuan yang berusia 18-23 tahun (Lampiran 4). Subjek uji merupakan keturunan ras kulit hitam Papua asli minimal sampai third degree relatives dan tinggal di Yogyakarta. Jumlah sampel minimal pada penelitian genetik menurut B-Rao (2001) adalah 29 orang (Lampiran 2), sehingga jumlah sampel subjek uji yang digunakan sudah memenuhi kriteria. Pada penelitian ini terdapat kendala dalam mencari subjek uji yang bersedia berpartisipasi dalam penelitian ini serta sebagian besar calon subjek uji adalah perokok. Pada subjek uji yang merupakan seorang perokok akan langsung dieksklusikan. Hal ini karena penelitian ini hanya akan mengidentifikasi alel gen CYP2A6*9 pada subjek nonperokok. Pada subjek uji yang sedang sakit akibat virus dan bakteri juga akan dieksklusikan karena dapat mempengaruhi kemurnian hasil isolasi DNA.
Isolasi DNA
Isolasi DNA menggunakan reagen FavorPrepTM Blood Genomic DNA Extraction Mini Kit yang terdiri atas Proteinase K, FABG Buffer, W1 buffer, wash buffer, dan elution buffer serta etanol absolut. Pada isolasi DNA dengan Extraction Mini Kit bahan yang digunakan telah dirancang untuk memudahkan dalam melakukan isolasi DNA serta terdapat fasilitas FABG column yang merupakan tabung dengan matriks yang berfungsi untuk mengikat DNA. Matriks
8
tersebut terlindung oleh tabung sehingga tidak mudah rusak (Kamaliah, 2017). Selain reagen terdapat beberapa alat yang digunakan untuk membantu proses isolasi seperti vortex dan microcentrifuge. Vortex adalah suatu alat yang berfungsi untuk mencampur sampel darah dengan reagen sehingga dapat tercampur dengan homogen (Maftuchah et al., 2014), sedangkan microcentrifuge berfungsi untuk memisahkan suatu bahan berdasarkan berat molekul dengan kecepatan tertentu (Bintang, 2018).
Pada isolasi DNA dengan Extraction Mini Kit diawali dengan pelisisan sel dengan menggunakan proteinase K dan FABG buffer (Anonim, 2019; Khosravinia et al., 2007), selain itu proteinase K juga berfungsi untuk pemurnian DNA dari protein (penghancuran protein) (Anonim, 2019; Fatchiyah et al., 2011; Kamaliah, 2017). DNA yang diperoleh setelah pelisisan sel masih tercampur dengan isi sel yang lain sehingga dilakukan proses presipitasi DNA dengan menggunakan etanol absolut (Kamaliah, 2017). DNA yang telah terpresipitasi kemudian akan dicuci dengan W1 buffer untuk menghilangkan residu protein dan residu RNA serta dicuci juga dengan wash buffer untuk menghilangkan residu garam serta etanol (Geneaid, 2019). Tahap terakhir ditambahkan elution buffer untuk melarutkan atau mengelusi DNA. Isolat DNA kemudian disimpan di dalam freezer pada suhu -70°C. Hasil isolat DNA dapat dianalisis kemurniannya secara kualitatif dengan teknik elektroforesis.
Analisis Kemurnian Isolat DNA
Analisis kemurnian isolat DNA dilakukan dengan teknik elektroforesis sebelum isolat DNA diamplifikasi dengan metode PCR. Elektroforesis merupakan teknik untuk memisahkan makromolekul seperti DNA dan protein yang bermuatan karena adanya pengaruh medan listrik berdasarkan berat molekul dan berguna dalam analisis kualitatif produk PCR. Selain untuk mendeteksi kemurnian DNA elektroforesis dapat digunakan untuk menentukan berat molekul (BM) dan menetapkan titik isolistrik (Bintang, 2018). Hasil analisis kemurnian isolat DNA yang baik ditandai dengan terbentuknya pita tunggal dan tebal serta tidak terbentuk pita ganda pada ukuran >10.000 bp yang menunjukkan isolat DNA murni, tidak tercemar oleh protein atau bagian sel lain serta tidak
9
terdegradasi (Faatih, 2009; Patramurti et al., 2014). Posisi pita >10.000 bp dapat diketahui dengan menggunakan 1KB (0.25-10 kb) DNA Ladder sebagai marker yang dapat menampilkan pita dari 250 bp sampai 10.000 bp. Berdasarkan hasil penelitian bentuk pita yang dihasilkan adalah pita tunggal dan tebal yang memiliki ukuran DNA >10.000 bp (Gambar 1). Hal ini menunjukkan isolat DNA murni dan tidak terkontaminasi.
Gambar 1. Hasil analisis kemurnian isolat DNA. Keterangan:
M : 1KB (0.25-10 kb) DNA Ladder sebagai marker 5, 10, 15, 20, dan 25 : Isolat DNA dengan pita tunggal dan tebal
Kondisi elektroforesis : Fase diam gel agarose 1%, fase gerak larutan 1x TBE buffer, kecepatan 100 volt/cm selama 30 menit.
Identifikasi Alel Gen CYP2A6*9
Identifikasi alel gen CYP2A6*9 menggunakan metode PCR. Kondisi PCR pada penelitian ini diatur menurut hasil optimasi oleh Cindy (2017), yaitu initial denaturasi pada suhu 94ºC selama 3 menit, denaturasi pada suhu 94ºC selama 30 detik, annealing pada suhu 60ºC selama 30 detik dan ekstensi pada suhu 70ºC selama 25 detik. Siklus amplifikasi dilakukan sebanyak 35 kali dan diakhiri dengan final ekstensi pada suhu 72ºC selama 5 menit.
Keberhasilan dalam proses amplifikasi juga ditentukan oleh ketepatan primer dan penggunaan beberapa bahan. Primer merupakan potongan DNA yang terdiri atas beberapa basa nukleotida yang berfungsi mengawali proses amplifikasi dengan menempel pada template DNA CYP2A6 sebagai target amplifikasi (Gambar 2). Penempelan (annealing) primer terjadi setelah pemisahan (denaturasi) untai DNA ganda menjadi untai DNA tunggal (Yuwono, 2005).
10.000 bp
M 5 10 15 20 25
10
Primer yang digunakan diadopsi dari Yoshida et al. (2003), yaitu primer forward 5'-GAT TCC TCT CCC CTG GAA C-3' dan primer reverse 5'- GGC TGG GGT GGT TTG CCT TTA-3'.
GATTCCTCTCCCCTGGAAC TAAAGGCAAACCACCCCAGCC
CTAAGGAGAGGGGACCTTG ATTTCCGTTTGGTGGGGTCGG
Gambar 2. Situs penempelan primer pada sekuen alel gen CYP2A6*1. Keterangan:
: Arah penempelan primer forward : Arah penempelan primer reverse
Selain primer penambahan bahan-bahan seperti WFI dan Promega Go Taq Green Master Mix yang mengandung taq DNA polymerase, dNTPs, MgCl2, dan buffer juga memiliki peran penting dalam proses amplifikasi. Taq DNA polymerase berperan dalam membuat untai baru atau memperpanjang (ekstensi) rantai DNA setelah penempelan primer pada template DNA dengan menambahkan basa nukleotida pada template DNA. Sumber basa nukleotida dalam amplifikasi DNA secara in vitro dengan PCR ini berasal dari deoksiribosanukleotida trifosfat (dNTPs), karena dNTPs mengandung deoksiadenosina trifosfat (dATP), deoksisistidina trifosfat (dCTP), deoksiguanosina trifosfat (dGTP), dan deoksitimina trifosfat (dTTP). MgCl2 merupakan salah satu faktor penting karena akan mempengaruhi aktivitas enzim Taq DNA polymerase, sedangkan buffer berfungsi untuk mempertahankan pH sehingga Taq DNA polymerase tetap berada dalam kondisi optimum selama proses amplifikasi berlangsung (Bintang, 2018).
Hasil amplifikasi ditunjukkan dengan terbentuknya pita tunggal dan tebal pada 368 bp yang menunjukkan alel gen CYP2A6*1, sedangkan bila tidak terbentuk pita pada 368 bp maka produk merupakan alel gen CYP2A6*9. Terbentuknya pita pada alel gen CYP2A6*1 karena primer forward yang digunakan dapat menempel pada ekson urutan 395 sampai 376 dan primer reverse
5’ 3’
5’ 3’
Primer forward
11
dapat menempel pada ekson urutan 48 sampai 28. Namun, pada alel gen CYP2A6*9 primer reverse yang digunakan tidak dapat melakukan amplifikasi karena terjadi SNP dalam kotak TATA di daerah promotor (T-48G) yang ditunjukkan pada gambar 3. Hal ini mengakibatkan tidak terjadi proses penempelan primer reverse (Yoshida et al., 2003).
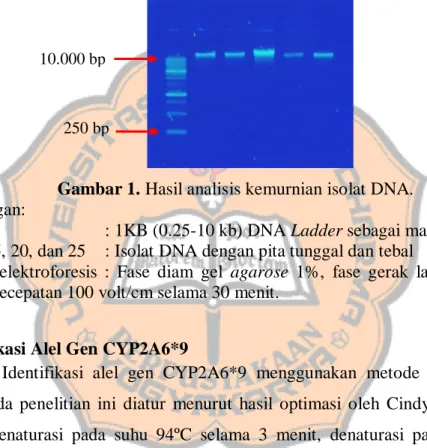
A. CYP2A6*1 GGATTCCTCTCCCCTGGAACCCCCAGATCCACAACTTTGGGGTGC B. CYP2A6*9 GGATTCCTCTCCCCTGGAACCCCCAGATCCACAACTTTGGGGTGC A. CYP2A6*1 ATTCTCACTCTCAGACCCCAAATCCAAAGCCCAAGTGCTCCCCTA B. CYP2A6*9 ATTCTCACTCTCAGACCCCAAATCCAAAGCCCAAGTGCTCCCCTA A. CYP2A6*1 TGCAAATATTCCAAACTCCTCAGTTCTACAGCTTATCTGTTGCCC B. CYP2A6*9 TGCAAATATTCCAAACTCCTCAGTTCTACAGCTTATCTGTTGCCC A. CYP2A6*1 CCTCCTAAATCCACAGCCCTGCGGCACCCCTCCTGAAGTACCACA B. CYP2A6*9 CCTCCTAAATCCACAGCCCTGCGGCACCCCTCCTGAAGTACCACA A. CYP2A6*1 GATTTAGTCTGGAGGCCCCCTCTCTGTTCAGCTGCCCTGGGGTCC B. CYP2A6*9 GATTTAGTCTGGAGGCCCCCTCTCTGTTCAGCTGCCCTGGGGTCC A. CYP2A6*1 CCTTATCCTCCCTTGCTGGCTGTGTCCCAAGCTAGGCAGGATTCA B. CYP2A6*9 CCTTATCCTCCCTTGCTGGCTGTGTCCCAAGCTAGGCAGGATTCA A. CYP2A6*1 TGGTGGGGCATGTAGTTGGGAGGTGAAATGAGGTAATTATGTAAT B. CYP2A6*9 TGGTGGGGCATGTAGTTGGGAGGTGAAATGAGGTAATTATGTAAT A. CYP2A6*1 CAGCCAAAGTCCATCCCTCTTTTTCAGGCAGTATAAAGGCAAACC B. CYP2A6*9 CAGCCAAAGTCCATCCCTCTTTTTCAGGCAGTAGAAAGGCAAACC A. CYP2A6*1 ACCCCAGCCG B. CYP2A6*9 ACCCCAGCC
Gambar 3. Urutan nukleotida alel gen CYP2A6*1 dan alel gen CYP2A6*9 (perbedaan pada basa nukleotida berwarna hijau dan biru yang mengalami SNP). Keterangan :
A. Potongan urutan basa alel gen CYP2A6*1 menurut Gen Bank B. Urutan basa alel gen CYP2A6*9 menurut Yoshida et al. (2003).
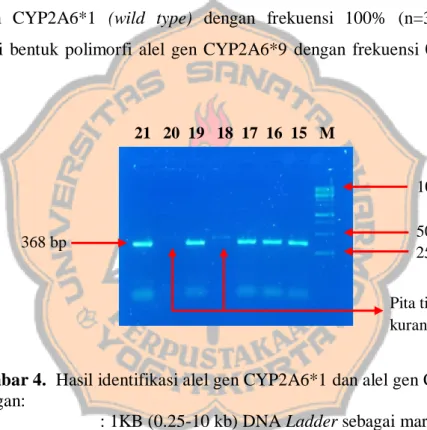
Hasil amplifikasi dianalisis menggunakan elektroforesis dan dilihat di bawah UV Transilluminator. Berdasarkan hasil analisis, semua sampel produk PCR menunjukkan terbentuknya pita tunggal, tebal dan terang pada 368 bp. Namun demikian, terdapat beberapa pita tunggal tipis yang kurang terang pada 368 bp (Gambar 4) yang disebabkan oleh konsentrasi DNA yang kecil. Hal ini dipengaruhi oleh kecilnya jumlah fragmen DNA yang teramplifikasi serta
12
kecilnya konsentrasi template DNA sehingga mempengaruhi intensitas pita yang dihasilkan (Azizah, 2009; Fahlevi et al., 2017; Rahayu et al., 2016). Hal tersebut dapat dibuktikan dengan analisis kuantitatif menggunakan Spektrofotometer, namun karena jumlah sampel terbatas maka analisis tersebut tidak dapat dilakukan. Terbentuknya pita pada 368 bp dari hasil analisis 30 sampel produk PCR menunjukkan bahwa semua subjek uji memiliki enzim CYP2A6 yang bekerja secara normal dalam aktivasi metabolisme senyawa TSNA dalam asap rokok yang bersifat prokarsinogen. Hal ini dikarenakan semua subjek uji memiliki alel gen CYP2A6*1 (wild type) dengan frekuensi 100% (n=30) dan tidak memiliki bentuk polimorfi alel gen CYP2A6*9 dengan frekuensi 0% (Lampiran 12).
Gambar 4. Hasil identifikasi alel gen CYP2A6*1 dan alel gen CYP2A6*9 Keterangan:
M : 1KB (0.25-10 kb) DNA Ladder sebagai marker 15-17, 19 dan 21 : Produk PCR dengan pita tunggal, tebal dan terang 18 dan 21 : Produk PCR dengan pita tunggal, tipis dan kurang terang Kondisi elektroforesis : Fase diam agarose 1,5%, fase gerak larutan 1x TBE buffer, kecepatan 100 volt/cm selama 30 menit.
Selain pada TSNA, CYP2A6 juga memiliki peran dalam memetabolisme beberapa obat dan bahan kimia seperti nikotin, coumarin, tegafur, dan letrozole (Tabel 1). Pada individu dengan alel gen CYP2A6*1 proses metabolisme akan terjadi secara normal sedangkan pada individu dengan bentuk polimorfi CYP2A6*9 proses metabolisme akan 50% lebih lambat karena terjadi SNP (Minematsu et al., 2006; Patramurti dan Fenty, 2017). Namun pada bentuk wild
500 bp 250 bp 368 bp
Pita tipis dan kurang terang
10.000 bp
13
type maupun polimorfi CYP2A6*9 terdapat keuntungan dan kekurangannya masing-masing. Berdasarkan hasil identifikasi pada 30 sampel produk PCR didapatkan hasil bahwa semua sampel memiliki alel gen CYP2A6*1 sehingga proses metabolisme pada beberapa substrat seperti nikotin, coumarin, tegafur, dan letrozole oleh enzim CYP2A6 akan terjadi secara normal.
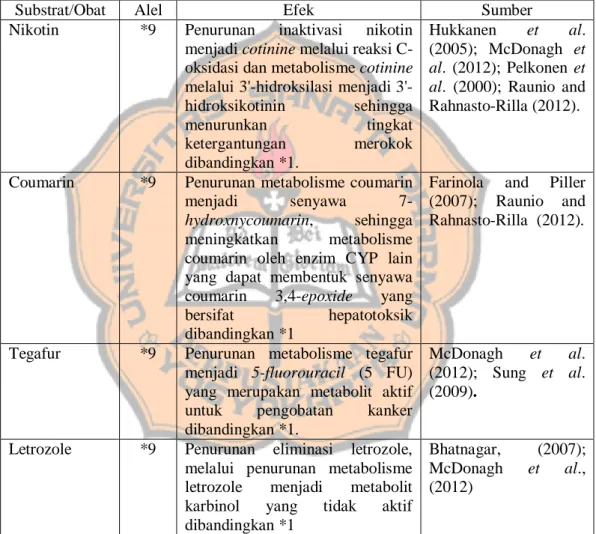
Tabel 1. Hubungan polimorfisme CYP2A6 dengan respon substrat/obat.
Substrat/Obat Alel Efek Sumber
Nikotin *9 Penurunan inaktivasi nikotin menjadi cotinine melalui reaksi C-oksidasi dan metabolisme cotinine melalui hidroksilasi menjadi 3'-hidroksikotinin sehingga menurunkan tingkat ketergantungan merokok dibandingkan *1. Hukkanen et al. (2005); McDonagh et al. (2012); Pelkonen et al. (2000); Raunio and Rahnasto-Rilla (2012).
Coumarin *9 Penurunan metabolisme coumarin
menjadi senyawa
7-hydroxnycoumarin, sehingga
meningkatkan metabolisme coumarin oleh enzim CYP lain yang dapat membentuk senyawa coumarin 3,4-epoxide yang bersifat hepatotoksik dibandingkan *1
Farinola and Piller (2007); Raunio and Rahnasto-Rilla (2012).
Tegafur *9 Penurunan metabolisme tegafur menjadi 5-fluorouracil (5 FU) yang merupakan metabolit aktif untuk pengobatan kanker dibandingkan *1.
McDonagh et al. (2012); Sung et al. (2009).
Letrozole *9 Penurunan eliminasi letrozole, melalui penurunan metabolisme letrozole menjadi metabolit karbinol yang tidak aktif dibandingkan *1
Bhatnagar, (2007); McDonagh et al., (2012)
KESIMPULAN
Hasil identifikasi alel gen CYP2A6*9 pada subjek uji nonperokok ras kulit hitam Papua Indonesia adalah tidak adanya alel gen CYP2A6*9 dengan frekuensi 0% dari 30 subjek uji.
14 SARAN
Penulis menyarankan untuk melakukan penelitian dengan jumlah subjek uji yang lebih besar. Dapat pula dilakukan penelitian pada gen CYP2A13 yang juga merupakan salah satu gen yang berperan dalam aktivasi metabolisme senyawa TSNA dalam asap rokok.
15
DAFTAR PUSTAKA
Alsanosi, S.M.M., Skiffington, C., Padmanabhan, S., 2014. Pharmacokinetic Pharmacogenomics, in: Handbook of Pharmacogenomics and Stratified Medicine. Elsevier Inc., pp. 341–364.
Anonim, 2019. Genomic DNA Extraction. https://www.prima-sci.com/14481965/genomic-dna-extraction. Accessed 19 November 2019.
Ashley, D.L., O’Connor, R.J., Bernert, J.T., Watson, C.H., Polzin, G.M., Jain, R.B., Hammond, D., Hatsukami, D.K., Giovino, G.A., Cummings, K.M., McNeill, A., Shahab, L., King, B., Fong, G.T., Zhang, L., Xia, Y., Yan, X. (Jane), McCraw, J.M., 2010. Impact of Differing Levels of Tobacco-Specific Nitrosamines in Cigarette Smoke on the Levels of Biomarkers in Smokers 1389–1398.
Azizah, A., 2009. Perbandingan Pola Pita Amplifikasi DNA Daun, Bunga, dan Buah Kelapa Sawit Normal dan Abnormal. Skripsi. Fakultas Matematika Dan Ilmu Pengetahuan Alam Institut Pertanian Bogor. Bogor.
B-Rao, C., 2001. Sample size considerations in genetic polymorphism studies. Human Heredity, 52(4), 191–200.
Bhatnagar, A.S., 2007. The discovery and mechanism of action of letrozole. SPringer, 105, 7–17.
Bintang, M., 2018. Biokimia Teknik Penelitian. Edisi 2. Erlangga, Jakarta.
Chiang, H. chih, Wang, C.Y., Lee, H.L., Tsou, T.C., 2011. Metabolic effects of CYP2A6 and CYP2A13 on 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK)-induced gene mutation-A mammalian cell-based mutagenesis approach. Toxicology and Applied Pharmacology, 253(2), 145–152.
Cindy., 2017. Optimasi dan Validasi Kondisi Polymerase Chain Reaction pada Identifikasi CYP2A6*9. Skripsi. Fakultas Farmasi Universitas Sanata Dharma. Yogyakarta.
Faatih, M., 2009. Isolasi dan Digesti DNA Kromosom. Jurnal Penelitian Sains dan Teknologi, 10(1), 61–67.
Fahlevi, M.R., Bakti, D., Sitepu, S.F., 2017. Karakterisasi Molekuler Elaeidobius kamerunicus Faust. (Coleoptera: Curculionidae) Asal Sumatera Utara Menggunakan Metode Amplified Fragment Length Polymorphism (AFLP). Jurnal Agroekoteknologi FP USU, 5(4), 941–953.
Farinola, N., Piller, N.B., 2007. CYP2A6 polymorphisms : is there a role for pharmacogenomics in preventing coumarin-induced hepatotoxicity in lymphedema patients ? 8(2), 151–158.
Fatchiyah, Arumingtyas, E.L., Widyarti, S., Rahayu, S., 2011. Biologi Molekular: Prinsip Dasar Analisis. Erlangga, Jakarta.
Fujieda, M., Yamazaki, H., Saito, T., Kiyotani, K., Gyamfi, M.A., Sakurai, M., Dosaka-Akita, H., Sawamura, Y., Yokota, J., Kunitoh, H., Kamataki, T., 2004. Evaluation of CYP2A6 genetic polymorphisms as determinants of smoking behavior and tobacco-related lung cancer risk in male Japanese smokers. Carcinogenesis, 25(12), 2451–2458.
16
Gayanti, 2017. Pemodelan Hubungan Alometri Tinggi Badan Dan Berat Badan Menggunakan Ordinary Least Product Regression. Institut Pertanian Bogor.
Geiss, O., Kotzias, D., 2007. Tobacco , Cigarettes and Cigarette Smoke, Reproduction.
Geneaid, 2019. Why does the plasmid DNA extraction kit require two washings with two types of wash buffers (W1 and Wash Buffer) while competitors kits require one washing?. https://www.geneaid.com/faq/why-does- plasmid-dna-extraction-kit-require-two-washings-two-types-wash-buffers-w1-and-wash. Accessed 19 November 2019.
Gyamfi, M.A., Fujieda, M., Kiyotani, K., Yamazaki, H., Kamataki, T., 2005. High prevalence of cytochrome P450 2A6*1A alleles in a black African population of Ghana. European Journal of Clinical Pharmacology, 60(12), 855–857.
Hecht, S.S., Hoffmann, D., 1989. The relevance of tobacco-specific nitrosamines to human cancer. Cancer Surv, 8(2), 273–294.
Hukkanen, J., Iii, P.J., Benowitz, N.L., 2005. Metabolism and Disposition Kinetics of Nicotine 57(1), 79–115.
IARC, 2007. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. The International Agency for Research on Cancer, Lyon, France.
IARC, 2004. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. International Agency for Research on Cancer, Lyon, France. Kamaliah, 2017. Perbandingan Metode Ekstraksi Dna Phenol-Chloroform dan Kit
Extraction pada Sapi Aceh dan Sapi Madura 5(1), 60–65.
Kementerian Kesehatan RI, 2019. Yuk, Mengenal Apa itu Perokok pasif?.
http://www.p2ptm.kemkes.go.id/infographic/yuk-mengenal-apa-itu-perokok-pasif. Diakses pada tanggal 22 Maret 2019.
Khosravinia, H., Murthy, H.N.N., Parasad, D.T., Pirany, N., 2007. Optimizing factors influencing DNA extraction from fresh whole avian blood. African Journal of Biotechnology, 6(4), 481–486.
Liu, T., Xie, C.-B., Ma, W.-J., Chen, W.-Q., 2013. Association Between CYP2A6 Genetic Polymorphisms and Lung Cancer: A Meta-Analysis of Case-Control Studies. Environmental and Molecular Mutagenesis, 54, 133– 140.
Maftuchah, Winaya, A., Zainudin, A., 2014. Teknik Dasar: Analisis Biologi Molekuler. Deepublish, Yogyakarta.
McDonagh, E.M., Wassenaar, C., David, S.P., Tyndale, R.F., Altman, R.B., Whirl-Carrillo, M., Klein, T.E., 2012. PharmGKB summary: very important pharmacogene information for cytochrome P-450, family 2, subfamily A, polypeptide 6. Pharmacogenetics and Genomics, 22(9), 695–708.
Minematsu, N., Nakamura, H., Furuuchi, M., Nakajima, T., Takahashi, S., Tateno, H., Ishizaka, A., 2006. Limitation of cigarette consumption by CYP2A6*4, *7 and *9 polymorphisms. European Respiratory Journal, 27(2), 289–292.
17
Mwenifumbo, J.C., Lessov-schlaggar, C.N., Zhou, Q., Krasnow, R.E., Swan, G.E., Benowitz, N.L., Tyndale, R., 2008. Identification of Novel CYP2A6 * 1B Variants : The CYP2A6 * 1B Allele is Associated With Faster In Vivo Nicotine Metabolism 83(1), 115–121.
Mwenifumbo, J.C., Zhou, Q., Benowitz, N.L., Sellers, E.M., Tyndale, R.F., 2010. New CYP2A6 gene deletion and conversion variants in a population of Black African descent. Pharmacogenomics, 11(2), 189–198.
Patramurti, C., Fenty, 2017. Studi Genotipe Sitokrom P450 2A6 Alel CYP2A6 * 4 dan CYP2A6 * 9 pada Subyek Uji Perokok Suku Jawa Indonesia 15(1), 50–56.
Patramurti, C., Sugiyanto, Nurrochmad, A., Martono, S., 2014. Polymorphism of Cytochrome P450 2A6 (CYP2A6*1 and CYP2A6*4) Among Javanese Indonesian Smoker and Non Smoker 26(1), 11–19.
Pelkonen, O., Rautio, A., Raunio, H., Pasanen, M., 2000. CYP2A6 : a human coumarin 7-hydroxylase. Elsevier, 144(2000), 139–147.
PharmVAr, 2012. CYP2A6 allele nomenclature.
https://www.pharmvar.org/gene/CYP2A6. Accessed 17 June 2019. Prabowo, D.A., 2017. Identifikasi Gen Sitokrom P450 2A6 (CYP2A6) Alel *9
pada Subyek Uji Perokok Ras Kulit Hitam Papua Indonesia. Skripsi. Fakultas Farmasi Universitas Sanata Dharma. Yogyakarta.
Querci, M., Jermini, M., Eede, G. Van de, 2006. The analysis of food samples for the presence of genetically modified organisms. JRC European Commission, Europe.
Rahayu, A.R., Pancasakti, H., Budiharjo, A., 2016. Pelacakan Gen Sitokrom Oksidase Subunit 1 (Co1) DNA Mitokondria Pada Itik Tegal (Anas sp.) 18(2), 114–122.
Raunio, H., Rahnasto-Rilla, M., 2012. CYP2A6: Genetics, structure, regulation, and function. Drug Metabolism and Drug Interactions, 27(2), 73–88. Rossini, A., Simao, Tatiana de Almeida Albano, R.M., Pinto, L.F.R., 2008.
Review CYP2A6 polymorphisms and risk for tobacco ‑ related cancers Review. Pharmacogenomics, 9(11), 1737–1752.
Sung, J.H., Dhiman, A., Shuler, M.L., 2009. A Combined Pharmacokinetic – Pharmacodynamic ( PK – PD ) Model for Tumor Growth in the Rat with UFT Administration. Journal of Pharmaceutical Sciences, 98(5), 1885– 1904.
Suryantisa, I., 2018. Situasi Umum Konsumsi Tembakau di Indonesia. Pusat Data dan Informasi Kementerian Kesehatan RI. Jakarta Selatan
Yoshida, R., Nakajima, M., Nishimura, K., Tokudome, S., Kwon, J.T., Yokoi, T., 2003. Effects of polymorphism in promoter region of human CYP2A6 gene (CYP2A6*9) on expression level of messenger ribonucleic acid and enzymatic activity in vivo and in vitro. Clinical Pharmacology and Therapeutics, 74(1), 69–76.
18
19 Lampiran 1. Ethical Clearance
20 Lampiran 2. Perhitungan Jumlah Sampel
Persamaan yang digunakan untuk menentukan jumlah sampel minimal pada penelitian genetik polimorfisme menurut B-Rao (2001)adalah :
≥
Keterangan:
: jumlah sampel minimal
: probabilitas alel utama : frekuensi alel minor
Pada penelitian ini terdapat dua alel yang akan diteliti, yaitu: CYP2A6*1 (alel utama) dan CYP2A6*9 (alel minor). Menurut B-Rao (2001), nilai dan
untuk kedua alel minor tersebut berturut-turut adalah = 0,95 dan
Jadi jumlah sampel minimal yang diperlukan untuk penelitian ini adalah:
=
= = 28,89
= 29 orang
21 Lampiran 3. Hasil Kuesioner
24 Lampiran 4. Data Subjek Uji Hasil Kuesioner
Subjek Umur (Tahun) Jenis Kelamin 1P 19 Perempuan 2P 23 Laki-laki 3P 22 Laki-laki 4P 19 Perempuan 5P 22 Perempuan 6P 23 Laki-laki 7P 19 Laki-laki 8P 22 Laki-laki 9P 20 Perempuan 10P 19 Perempuan 11P 18 Perempuan 12P 19 Perempuan 13P 22 Laki-laki 14P 20 Perempuan 15P 21 Perempuan 16P 21 Laki-laki 17P 20 Laki-laki 18P 20 Laki-laki 19P 19 Laki-laki 20P 19 Laki-laki 21P 23 Laki-laki 22P 22 Laki-laki 23P 20 Laki-laki 24P 18 Laki-laki 25P 19 Laki-laki 26P 20 Laki-laki 27P 20 Laki-laki 28P 19 Perempuan 29P 20 Laki-laki 30P 22 Perempuan
25 Lampiran 5. Usage Information of FavorPrepTM
27
28
29
30
31
Lampiran 10. Hasil Analisis Kemurnian Isolat DNA
Lampiran 11. Hasil Identifikasi Alel Gen CYP2A6*1 dan *9
21 20 19 18 17 16 15 M 28 27 26 25 24 23 22 M 1 2 3 4 5 6 7 M 14 13 12 11 10 9 8 M 29 30 M M 5 10 15 20 25
32
Lampiran 12. Frekuensi Alel Gen CYP2A6 *1 dan *9
Alel Jumlah Frekuensi
CYP2A6*1 30 100% CYP2A6*9 0 0% Frekuensi CYP2A6*1 = = = 100% Frekuensi CYP2A6*9 = = = 0%
33
BIOGRAFI PENULIS
Penulis bernama lengkap Mensiana Lodvi Putarato, lahir di Waikabubak pada tanggal 31 Mei 1997. Penulis merupakan anak pertama dari empat bersaudara dari pasangan Bapak Victor Ngongo Putarato dan Ibu Dorkas Lende. Penulis menempuh pendidikan TK sampai SMA di Sumba Barat, Nusa Tenggara Timur pada TK Pertiwi (2001- 2003), SD Masehi Waikabubak II (2003-2009), SMP Negeri 3 Waikabubak (2009-2012), dan SMA Negeri 1 Waikabubak (2012-2015). Pada tahun 2016, penulis melanjutkan pendidikan ke jenjang Perguruan Tinggi di Fakultas Farmasi Universitas Sanata Dharma Yogyakarta. Selama masa kuliah, penulis aktif dalam beberapa kepanitiaan seperti Pharmalympic tahun 2017 sebagai anggota divisi pendaftaran, Tiga Hari Temu Akrab Farmasi (Titrasi) 2018 sebagai anggota divisi medis, dan Pharmacy Performance tahun 2018 sebagai koordinator divisi konsumsi. Penulis juga berpartisipasi sebagai koordinator asisten dosen Biokimia tahun 2019.