0
METABOLISME ZAT GIZI MAKRO
Oleh :
DIAH MAUNAH
9999 – 32 – 048
JURUSAN GIZI
FAKULTAS ILMU – ILMU KESEHATAN
UNIVERSITAS ESA UNGGUL
1
KARBOHIDRAT
PENDAHULUAN
Karbohidrat (KH) atau Hidrat Arang (HA) merupakan salah satu dari zat gizi makro yang memegang peranan penting sebagai sumber energi utama bagi manusia, dimana setiap gramnya menghasilkan 4 kcal. Walaupun lemak menghasilkan energi lebih besar daripada karbohidrat yaitu 9 kcal/gram, namun karbohidrat lebih banyak dikonsumsi sehari-hari sebagai bahan makanan pokok. Di negara sedang bekembang karbohidrat dikonsumsi sekitar 70-80% dari total kalori, bahkan pada daerah miskin dapat mencapai 90% dari total kalori. Sedangkan pada negara maju karbohidrat hanya dikonsumsi sekitar 40-60% dari total kalori.
Secara kima karbohidrat didefinisikan sebagai turunan aldehid atau keton dari alkohol polihidrik (karena mengandung gugus hidroksi lebih dari satu), atau sebagai senyawa yang menghasilkan turunan tersebut (Muchtadi, Deddy. Pengantar Ilmu Gizi. 2009). Secara umum definisi karbohidrat adalah senyawa organik yang mengandung atom Karbon, Hidrogen dan Oksigen, dan pada umumnya unsur Hidrogen clan oksigen dalam
komposisi menghasilkan H2O. Di dalam tubuh karbohidrat dapat dibentuk dari beberapa
asam amino dan sebagian dari gliserol lemak. Sumber karbohidrat nabati dalam glikogen bentuk glikogen, hanya dijumpai pada otot dan hati dan karbohidrat dalam bentuk laktosa hanya dijumpai di dalam susu. Pada tumbuh-tumbuhan, karbohidrat di bentuk dari basil
reaksi CO2 dan H2O melalui proses foto sintese di dalam sel-sel tumbuh-tumbuhan yang
mengandung hijau daun (klorofil). Matahari merupakan sumber dari seluruh kehidupan, tanpa matahari tanda-tanda dari kehidupan tidak akan dijumpai. Akan tetapi sebagian besar karbohidrat diperoleh dari bahan makanan yang dikonsumsi sehari-hari, terutama sumber bahan makan yang berasal dari tumbuh-tumbuhan, dimana Pada proses fotosintesis, klorofil pada tumbuh-tumbuhan akan menyerap dan menggunakan energi matahari untuk
membentuk karbohidrat dengan bahan utama CO2 dari udara dan air (H2O) yang berasal dari
tanah. Energi kimia yang terbentuk akan disimpan di dalam daun, batang, umbi, buah dan biji-bijian.
2
Sinar matahari
6 CO2 + 6 H2O ---> C6 H12 O6 + 6 O2 Klorofil
SUSUNAN KIMIA
Semua jenis karbohidrat terdiri atas unsur-unsur karbon (C), hidrogen (H), dan oksigen (O). Paerbandingan antara hidrogen dan oksigen pada umumnya adalah 2:1 seperti halnya dalam air, dengan demikian diberi nama karbohidrat. Dalam bentuk sederhana,
formula umum karbohidrat adalah CnH2nOn. Hanya heksosa (6-atom karbon), serta pentosa
(5-atom karbon), dan polimernya memegang peranan penting dalam nutrisi.
KLASIFIKASI KARBOHIDRAT
Berdasarkan sifat ketersediaannya (availabilitas) bagi tubuh, maka karbohidrat yang terkandung dalam bahan pangan dikelompokkan menjadi dua golongan, yaitu :
1. Karbohidrat tersedia (Available Carbohydrate) adalah karbohidrat yang dapat dicerna
dan atau diserap serta dimetabolisme dalam tubuh. Kelompok ini meliputi monosakarida (misalnya glukosa, fruktosa, dan galaktosa); disakarida dan oligosakarida (misalnya sukrosa, laktosa, maltosa, trehalosa dan oligosakarida lain yang sejenis dengan maltosa dan isomaltosa); dan polisakarida glukan (misalnya pati, dekstrin, dan glikogen).
2. Karbohidrat tidak tersedia tubuh (Unvailable Carbohydrate). Karbohidrat yang termasuk
dalam kelmpok ini adalah karbohidrat yang tidak dapat hidrolisis oleh enzim-enzim yang terdapat dalam saluran pencernaan manusia, sehingga akirnya tidak dapat diserap oleh tubuh. Karbohidrat yang termasuk kelompok ini adalah oligosakarida yang tergolong sebagai seri rafinosa (rafinosa, stakhiosa, dan verbaskosa); polisakarida glukan (selulosa); polisakarida turunan (hemiselulosa, lignin, gum, pektin); serta beberapa macam disakarida misalnya laktulosa. Karbohidrat jenis ini dapat difermentasi oleh mikroflora yang terdapat dalam saluran pencernaan, menjadi asam lemak rantai pendek dan asam laktat. Sebagian dari hasil fermentasi akan diserap oleh usus besar dan akhirnya dimetabolisme oleh tubuh.
Secara umum dalam ilmu gizi, penggolongan karbohidrat didasarkan pada jumlah molekulnya, yaitu karbohidrat sederhana dan karbohidrat kompleks. Sesungguhnya semua
3
jenis karbohidrat terdiri dari karbohidrat sederhana atau gula sederhana, sedangkan karbohidrat komplek memiliki lebih dari dua unit gula sederhana didalam satu molekul.
Karbohidrat Sederhana, terdiri atas :
1. Monosakarida, yang terdiri atas jumlah C yang sama dengan molekul air, yaitu
Heksosa (mengandung 6 buah karbon [C6(H2O)6] contoh Glukosa, Fruktosa,
Galaktosa) dan Pentosa (mengandung 5 buah karbon [C5(H2O)5] contoh Ribosa,
Arabinosa, dan Xylosa).
2. Disakarida, yang terdiri atas ikatann 2 monosakarida. Dimana untuk tiap 12 atom C
ada 11 molekul air [C12(H2O)11]. Contoh Sukrosa, Maltosa, Laktosa.
3. Gula Alkohol, merupakan bentuk alkohol dari monosakarida.
4. Oligosakarida, adalah gula rantai pendek yang dibentuk oleh galaktosa, glukosa, dan
fruktosa.
1. MONOSAKARIDA (C6 H12 O6)
Merupakan jenis karbohidrat yang paling sederhana (simple sugar), oleh karena tidak dapat lagi dihidrolisa dan terdiri dari molekul tunggal. Monosakarida larut dalam air, memiliki rasa yang manis (sehingga secara umum disebut juga gula). Tata nama monosakarida tergantung dari gugus fungsional yang dimiliki, terdapa nama monosakarida yang lain yaitu: Triosa (3 karbon), Tentrosa (4 karbon), Pentosa (5 karbon), dan Heksosa (6 karbon), Sebagian besar monosakarida dikenal dan yang terpenting adalah heksosa, karena terdiri dari 6-rantai atau cincin karbon. Atom-atom hidrogen dan oksigen terikat pada rantai atau cincin secara terpisah atau sebagai gugus hidroksil (OH). Terdapat tiga jenis heksosa yang terpenting dalam ilmu gizi, yaitu glukosa, fruktosa, dan galaktosa. Ketiga macam monosakarida ini mengandung jenis dan jumlah atom yang sama, yaitu 6 atomkarbon, 12 atom hidrogen, dan 6 atom oksigen. Perbedaan terlatak pada cara penyusunan atom-atom hidrogen dan oksigen disekitar atom-atom karbon. Perbedaan penyunan atom tersebut memberikan perbedaan dalam tingkat kemanisan, daya larut, dan sifat lain dari ketiga monosakarida tersebut. Monosakarida yang terdapat pada alam umumnya terdapat dalam bentuk isomer dekstro (D).
4
-D-Galaktosa
Gambar 1. Struktur kimia monosakarida.
GLUKOSA
Disebut juga dekstrosa atau gula anggur, terdapat luas dialam dalam jumlah sedikit, yaitu di dalam sayuran, buah, sirup jagung, sari pohon, dan bersamaan dengan fruktosa terdapat dalam madu, sari tebu (molase). Dalam proses metabolisme, glukosa merupakan bentuk karbohidrat yang beredar dalam tubuh yang berasal dari hasil pemecahan glikogen (cadangan karbohidrat dalam jaringan), dari pangan yang dikonsumsi atau sebagai pemecahan karbohidrat lain yang lebih kompleks seperti hasil akhir pencernaan amilum, sukrosa, maltosa, dan laktosa. Karena hanya glukosa yang ditemukan dalam plasma darah dan sel darah merah, maka glukosa terkadang disebut sebagai gula darah yang berfungsi sebagai penyedia energi bagi seluruh sel dan jaringan tubuh. Tubuh hanya dapat menggunakan glukosa dalam bentuk D. Dalam keadaan normal sistem saraf pusat hanya menggunakan glukosa sebagai sumber energi.
Glukosa murni yang terdapat dipasaran diperoleh dari hasil olahan pati. Tingkat
kemanisan glukosa hanya separuh dari sukrosa, sehingga digunakan lebih banyak untuk tingkat kemanisan yang sama.
FRUKTOSA
Disebut juga levulosa atau gula buah, merupakan gula paling manis (dengan tingkat
kemanisan 1,2 dibandingkan dengan sukrosa dan isoglukosa 1,0; glukosa 0,7; sorbitol 0,5). Fruktosa memiliki rumus kima yang sama dengan glukosa namun strukturnya berbeda. Fruktosa terutama terdapat dalam sepertiga madu (bersama glukosa), dala buah, nektar bunga, sayur, hasil hidrolisa gula tebu. Sedangkan dalam tubuh fruktosa didapat dari hasil pemecahan sukrosa atau sakarosa. Secara komersil fruktosa yang diperoleh dari pati digunakan sebagai pemanis, seperti minuman ringan (softdrink) menggunakan sirup jagung-tinggi-fruktosa sebagai bahan pemanis.
5
GALAKTOSA
Tidak ditemukan secara bebas di alam seperti glukosa dan fruktosa, tetapi dihasilkan
di dalam tubuh selama berlangsungnya proses pencernaan laktosa. Galaktosa merupakan komponen serebrosida, yaitu lemak turunan yang terdapat di dalam otak dan jaringan saraf.
MANOSA
Jarang terdapat di dalam makanan. Di daerah gurun pasir manosa terdapat di dalam manna yang diolah untuk membuat roti.
PENTOSA
Merupakan bagian sel-sel semua bahan makanan alami. Terdapat dalam jumlah
kecil, sehingga tidak terlalu penting sebagai sumber energi. Ribosa dan deoksiribosa merupakan bagian asam nukleat dalam inti sel. Karena dapat disintesis oleh semua hewan, ribosa dan deoksiribosa tidak merupakan zat gizi esensial.
2.DISAKARIDA
Merupakan gabungan antara dua monosakarida yang melekat melalui proses kondensasi. Kedua monosakarida saling mengikat berupa ikatan glikosidik melalui satu atom oksigen. Disakarida dapat dipecah kembali menjadi dua molekul monosakarida melaui reaksi hidrolisis. Terdapat empat jenis disakarida yaitu sukrosa atau sakarosa, maltosa, laktosa, dan trehalosa.
SUKROSA
Sukrosa atau sakarosa disebut juga gula tebu atau gula bit. Merupakan jenis gula yang sering dipergunakan sehari-hari sehingga disebut gula meja (table sugar) atau gula pasir / gula invert. Mempunyai dua melekol monosakarida yang terdiri dari monosakirida yang terdiri dari satu molekul glukosa dan satu molekul fruktosa. Bila dicernakan atau dihidrolisis, sukrosa pecah menjadi satu unit fruktosa.
Surosa terdapat di tebu (100% mengandung sukrosa), bit, gula nira (50%), selai, jelly, buah, sayur dan madu.
MALTOSA (Gula Malt)
Tidak terdapat bebas di alam. Mempuyai dua molekul monosakarida yang terdiri dari dua molekul glukosa. Di dalam tubuh maltosa didapat dari hasil pemecahan amilum,
6
pati, sepertiyang terjadi pada tumbuh – tumbuhan bila benih atau bijian berkecambah dan di dalam usus manusia pada pencernaan pati. Dengan Iodium amilum akan berubah menjadi warna biru. Amilum terdiri dari 2 fraksi (dapat dipisah kan dengan air panas) yaitu :
1. Amilosa
- Larut dengan air panas
- Mempunyai struktur rantai lurus 2. Amilopektin
- Tidak larut dengan air panas
- Mempunyai sruktur rantai bercabang
Peranan perbandingan amilosa dan amilo pektin terlihat pada serelia; Contohnya beras, semakin kecil kandungan amilosa atau semakin tinggi kandungan amilopektinnya, semakin lekat nasi tersebut. Pulut sedikit sekali amilosanya (1-2%), beras mengandung amilosa > 2%. Berdasarkan kandungan amilosanya, beras (nasi) dapat dibagi menjadi 4 golongan yaitu amilosa tinggi 25-33%, amilosa menengah 20-25% amilosa rendah 09-20%, amilosa sangat rendah < 9%.
LAKTOSA
Hanya terdapat dalam susu dan terdiri atas satu unit glukosa dan satu unit galaktosa. Laktosa kurang larut dalam air. Sumber laktosa hanya terdapat pada susu sehingga disebut juga gula susu (susu sapi 4-5% dan asi 4-7%).
Laktosa diperlukan untuk pertumbu ban sel-sel otak. Laktosa dapat menimbulkan intolerance (laktosa intolerance) disebabkan kekurangan enzim laktase sehingga kemampuan untuk mencema laktosa berkurang. Kelainan ini dapat dijumpai pada bayi, anak dan orang dewasa, baik untuk sementara maupun secara menetap. Gejala yang sering dijumpai adalah diare, gembung, flatus dan kejang perut. Defisiensi laktase pada bayi dapat menyebabkan gangguan pertumbuhan, karena bayi sering diare.
TREHALOSA
Terdiri atas dua mol glukosa dan dikenal sebagai gula jamur. Sebanyak 15% bagian kering jamur terdiri atas trehalosa. Trehalosa juga terdapat dalam seranga.
7
3. GULA ALKOHOL
Terdapat 4 jenis gula alkohol, yaitu sorbitol, manitol, dulcitol, dan inositol.
SORBITOL
Terdapat didalam beberapa jenis buah dan secara komersial dibuat dari glukosa. Enzim aldosa reduktase dapt mengubah gugus aldehida (CHO) dalam glukosa menjadi alkohol (CH2OH). Tingkat kemanisan 60% bila dibandingkan dengan sukrosa, diabsorpsi lebih lambat dan diubah di dalam hati menjadi glukosa. Sorbitol tidak mudak dimetabolisme oleh bakteri dalam mulut sehingga tidak mudah menimbulkan karies gigi.
MANITOL dan DULCITOL
Merupakan alkohol yang dibuat dari monosakarida manosa dan galaktosa. Manitol terdapat di dalam nanas, asparagus, ubi jalar, dan wortel. Secara komersial manitol diekstrasi dari sejenis rumput laut.
INOSITOL
Merupakan alkohol siklis yang menyerupai glukosa. Inositol terutama terdapat dalam sekam serealia. Bentuk esternya dengan fitat akan menghambat absorpsi kalsium dan zat besi dalam usus halus.
4. OLIGOSAKARIDA
Terdiri atas polimer dua hingga sepuluh monosakarida (oligo = sedikit). Rafinosa, stakiosa, dan verbakosa adalah oligosakarida yang terdiri atas unit-unit glukosa, fruktosa, dan galaktosa. Ketiga jenis oligosakarida ini terdapat di dalam biji tumbuhan
dan kacang-kacangan serta tidak dapat dipecah oleh enzim-enzim pencernaan. Fruktan
adalah sekelompok oligo dan polisakarida yang terdiri atas beberapa unit fruktosa yang terikat dengan satu molekul glukosa. Fruktan terdapat pada serealia, bawang merah, bawang putih, dan asparagus. Fruktan tidak dicernakan secra berarti. Sebagian besar di dalam usus besar difermentasi.
KARBOHIDRAT KOMPLEKS , terdiri dari :
1. Polisakarida, yang terdiri atas lebih dari dua ikatan monosakarida.
8
Merupakan senyawa karbohidrat kompleks yang mengandung lebih dari 60.000 molekul monosakarida yang tersusun membentuk rantai lurus ataupun bercabang. Polisakarida rasanya tawar (tidak manis), tidak seperti monosakarida dan disakarida. Di dalam Ilmu Gizi jenis polisakarida yang terpenting adalahadalah amilum (pati), dekstrin, glikogen, dan selulosa maupun polisakarida nonpati..
Amilum (zat pati)
Merupakan sumber energi utama bagi orang dewasa di seluruh penduduk dunia, terutama di negara seclang berkembang oleh karena di konsumsi sebagai bahan makanan pokok. Disamping bahan pangan kaya akan amilum juga mengandung protein, vitamin, serat dan beberapa zat gizi penting lainnya. Amilum merupakan karbohidrat dalam bentuk simpanan bagi tumbuh-tumbuhan dalam bentuk granul yang dijumpai pada umbi dan akarnya.
Sumber: umbi-umbian,serealia dan biji-bijian merupakan sumber amilum yang berlimpah ruah oleh karena mudah didapat untuk di konsumsi. Jagung, beras dan gandum kandungan amilurnnya lebih dari 70%, sedangkan pada kacang-kacangan sekitar 40%.
Jumlah unit glukosa dan susunannya dalam satu jenis pati berbeda satu sama lain, bergantung pada jenis tanaman asalnya. Bentuk butiran pati berbeda satu sama lain dengan karakteristik tersendiri dalam hal daya larut, daya mengentalkan, dan rasa. Amilum tidak larut di dalam air dingin, tetapi larut di dalam air panas membentuk cairan yang sangat pekat seperti pasta; peristiwa ini disebut "gelatinisasi". Cabang-cabang dalam struktur amilopektinlah yang terutama menyebabkannya dapat membentuk gel yang cukup stabil. Selain itu proses pemasakan pati juga dapat menyebabkan melukkan dan memecahkan sel pati sehingga memudahkan proses pencernaan yang akan dihidrolisis menjadi glukosa, pada tahap pertengahan akan dihasilkan dekstrin dan maltosa.
Amilosa merupakan rantai panjang unit glukosa yang tidak bercabang, sedangkan amilopektin adalah polimer yang susunannya bercabang-cabang degan 15-30 unit glukosa pada tiap cabang. Rantai glukosa terikat satu sama lain melalui ikatan alfa yang dapat dipecah dalam proses pencernaan.
9
Komposisi amilosa dan amilopektin berbeda dalam pai berbagai bahan makanan. Amilopektin pada umumnya terdapat dalam jumlah yang lebih besar. Sebagian besar pati mengandung 15% dan 35% amilosa. Pada beras semakin kecil kandungan amilosa atau semakin tinggi kandungan amilopektinnya, semakin pulen (lekat) nasi yang diperoleh. Dekstrin
Merupakan zat antara dalam pemecahan amilum. Dektrin merupakan sumber utama karbohidrat dalam makanan lewat pipa (tube feeding). Molekulnya lebih sederhana, lebih mudah larut di dalam air, dengan yodium akan berubah menjadi warna merah. Molekul dekstrin lebih besar dari sukrosa dan glukosa sehingga memiliki pengaruh osmolar lebih kecil yang tidak mudah menimbulkan diare. Pati yang dipanggang secara kering akan menghasilkan dekstrin (contoh roti panggang). Dektrin lebih manis daripada pati dengan daya larut lebih tinggi dan lebih mudah dicernakan. Dekrin maltosa merupakan suatu produk hasil hidrolisis parsial apti, digunakan sebagi mkanan bayi karena tidak mudah mengalami fermentasi dan mudah dicerna.
Glikogen
Glikogen merupakan "pati hewani" karena merupakan bentuk simpanan karbohidrat dalam tubuh manusia dan hewan yang terutama terdapat pada hati dan otot, terbentuk dari ikatan 1000 molekul, larut di dalam air (pati nabati tidak larut dalam air) dan bila bereaksi dengan iodium akan menghasilkan warna merah. Glikogen terdiri atas unit-unit glukosa dalam bentuk rantai lebih bercabang daripada amilopektin sehingga membuat glikogen lebih mudah dipecah. Glikogen disimpan di dalam hati dan otot sebagai cadangan energi, yang sewaktu-waktu dapat diubah kembali menjadi glukosa bila dibutuhkan. Tubuh memiliki kapasitas terbatas untuk menyimpan glikogen, yaitu hanya sebanyak 350gram. Dua pertiga bagian dari glikogen disimpan dalam otot dan selebihnya disimpan dalam hati. Glikogen dalam otot dipergunakan hanya untuk keperluan energi didalam otot, sedangkan glikogen di hati dipergunakan sebagai sumber energi semua sel tubuh. Kelebihan glukosa melampaui kemempuannya menyimpan dalam bentuk glikogen akan disimpan menjadi lemak dan disimpan dalam jaringan lemak.
Pada waktu hewan disembelih, terjadi kekejangan (rigor mortis) dan kemudian glikogen dipecah menjadi asam laktat selama post mortum.
10
Definisi serat makanan adalah polisakarida nonpati yang menyatakan polisakarida dinding sel. Terdapat dua golongan serat, yaitu yang dapat larut dengan air dan yang tidak dapat larut dengan air. Serat yangg tidak dapat larut dengan aor adalah selulosa, hemiselulosa, dan lignin. Serat yang larut dalam air adalah pektin, gum, mukilase, glukan, dan algal.
Hampir 50% karbohidrat yang berasal dari tumbuh-tumbuhan adalah selulosa, karena selulosa merupakan bagian yang terpenting dari dinding sel tumbuh-tumbuhan. Selulosa tidak dapat dicerna oleh tubuh manusia, oleh karena tidak ada enzim untuk memecah selulosa. Meskipun tidak dapat dicerna, selulosa berfungsi sebagai sumber serat yang dapat memperbesar volume dari faeses, sehingga akan memperlancar defekasi. Selulosa melunakkan dan memberi bentuk pada feces karena mampu menyerap air, sehingga membantu pergerakan peristaltik usus, dengan demiian membantu defekasi dan mencegah konstipasi.
Lignin terdiri dari polimer bercabang heterogen heksosa, pentosa, dan asam uronat. Lignin memberi kekuatan pada struktur tumbuhan, oleh karena itu merupakan bagian keras dari tumbuhan sehingga jarang dimakan. Lignin terdapat pada tangkai sayuran, bagian inti di dalam worten, biji jambu biji. Menurut Garrow dan James, 1993 Lignin sesungguhnya bukan karbohidrat dan seharusnya tidak dimasukkan dalam serat makanan.
Pektin, gum, dan mukilase terdapat disekelilig dan di dalam sel tumbuhan. Ikatan ini larut atau mengembang di dalam air sehingga membentuk gel. Sehingga dipergunakan dalam industri sebagai emulsifier, stabilizer, dan pengental. Pektin merupakan polimer ramnosa dan asam galakturonat dengan cabang yang terdiri dari rantai galaktosa dan arabinosa. Pektin terdapat pada sayur dan buah, terutama jenis sitrus, apel, jambu, anggur, dan wortel. Senyawa pektin berfungsi sebagai bahan perekat antara dinding sel. Buah yang memiliki kandungan pektin tinggi baik digunakan sebagai bahan dasar pembuatan selai/jam.
Gum adalah polisakarida larut air yang terdiri atas 10.000 – 30.000 unit yang terutama terdiri atas glukosa, galaktosa, manosa, arabinosa, ramnosa, dan asam uronat.
11
Gum arabic adalah sari pohon akasia. Gum dipergunakan sebagai emulsifier, stabilizier, dan pengental.
Mukilase merupakan struktur kompleks yang mempunyai ciri khas yaitu memiliki komponen asam D-galakturonat. Mukilase terdapat di dalam biji-bijiandan akar yang berfungsi mencegah pengeringan.
Beta-glukan terutama terdiri atas polimer glukosa bercabang yang terikat dalam bentuk Beta (1-3) dan Beta (1-9). Beta-glukan terdapat dalam serealia, terutama dalam oatdan barley, dan berperan dalam menurunkan kadar kolesterol dalam darah. Polisakarida algal yang diambil dari algae dan rumput laut merupakan polimer asam-asam manuronat dan guluronat. Banyak dipergunakan sebagai bahan agar-agar dan sebagai pengental serta stabilizer.
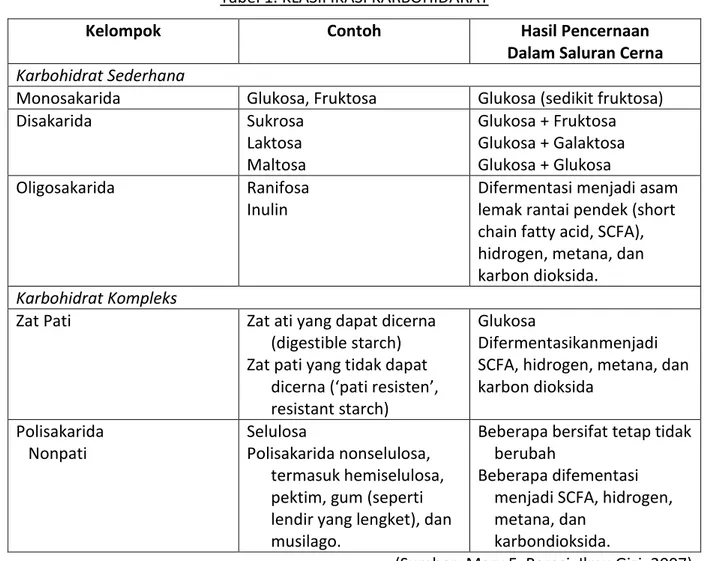
Tabel 1. KLASIFIKASI KARBOHIDARAT
Kelompok Contoh Hasil Pencernaan
Dalam Saluran Cerna
Karbohidrat Sederhana
Monosakarida Glukosa, Fruktosa Glukosa (sedikit fruktosa)
Disakarida Sukrosa Laktosa Maltosa Glukosa + Fruktosa Glukosa + Galaktosa Glukosa + Glukosa Oligosakarida Ranifosa Inulin
Difermentasi menjadi asam lemak rantai pendek (short chain fatty acid, SCFA), hidrogen, metana, dan karbon dioksida.
Karbohidrat Kompleks
Zat Pati Zat ati yang dapat dicerna
(digestible starch) Zat pati yang tidak dapat
dicerna (‘pati resisten’, resistant starch)
Glukosa
Difermentasikanmenjadi SCFA, hidrogen, metana, dan karbon dioksida Polisakarida Nonpati Selulosa Polisakarida nonselulosa, termasuk hemiselulosa, pektim, gum (seperti lendir yang lengket), dan musilago.
Beberapa bersifat tetap tidak berubah
Beberapa difementasi menjadi SCFA, hidrogen, metana, dan
karbondioksida.
12
FUNGSI KARBOHIDRAT
1. Sumber energi
Didalam tubuh karbohidrat merupakan salah satu sumber energi utama. Dalam satu gram karbohidrat menghasilkan 4kcal energi. Sebagian karbohidrat di dalam tubuh berada dalam sirkulasi darah sebagai glikosa untuk keperluan energi segera; sebagian disimpan sebagai glikogen dalam hati dan jariangan otot, dan sebagian diubah menjadi lemak untuk kemudian disimpan sebagi cadangan energi di dalam jaringan lemak. Glukosa merupakan sumber energi utama bagi sistem saraf sentral, otak, dan eritrosit.
2. Pemberi rasa manis pada makanan
Karbohidrat memberi rasa manis pada makanan, khususnya mono dan disakarida. Gula tidak mempunyai rasa manis yang sama. Fruktosa adalah gula paling manis. Bila tingkat kemanisa sakarosa diberi nilai 1 makan tinggkat kemanisan fruktosa adalah 1,7; glukosa 0,7; maltosa 0,4; laktosa 0,2.
3. Melindungi protein agar tidak dibakar sebagai penghasil energi / penghemat protein.
Bila karbohidrat makanan tidak mencukupi, maka protein akan dipergunakan untuk memenuhi kebutuhan energi tubuh, dengan mengalahkan fungsi utamanya sebagai zat pembangun. Sebaliknya, bila karbohidrat makanan mencukupi maka protein akan digunakan sebagi zat pembangun (sebagai fungsi utama protein).
4. Pengatur metabolisme lemak
Karbohidrat mencegah terjadinya oksidasi lemak yang tidak sempurna, sehingga menghasilkan bahan keton berupa asam asetoasetat, aseton, dan asam beta-hidroksi-butirat. Bahan ini dibentuk didalam hati dan dikeluarkan melelui urine dengan mengikat basa berupa ion natrium. Hal ini dapat menyebabkan ketidakseimbangan natrium dan dehidrasi. pH cairan tubuh akan menurun. Keadaan ini menimbulkan ketosis atau asidosis yang dapat merugikan tubuh, untuk mencegah keadaan tersebut dibutuhkan setidaknya 50-100gram karbohidrat dalam sehari.
5. Membantu pengeluaran feses. Karbohidarat membantu pengeluaran feses dengan cara
mengatur peristaltik usus dan memberi bentuk pada feses. Selulosa dalam serat makanan mengatur peristaltik usus, sedangkan hemiselulosa dan pektin mampu menyerap banyak air dalam usus besar sehingga memberi bentuk pada sisa makanan
13
yang akan dikeluarkan. Serat berfungsi memperbaiki kinerja peristaltik usus dan pemberi muatan pada sisa makanan, memiliki efek hipolipidemik, efek hipoglikemik, mencegah kegemukan, kostipasi, penyakit divertikulosis, kanker usus besar, membantu mengendalikan gula darah, dan jantung koroner yang berkaitan dengan kadar kolesterol yang tinggi.
6. Di dalam hepar berfungsi untuk detoksifikasi zat – zat toksik tertentu.
7. Beberapa jenis karbohidrat memiliki fungsi khusus di dalam tubuh. Misalnya laktosa
berfungsi membantu penyerapan kalsium, ribosa merupakan merupakan komponen yang penting dalam asam nukleat.
SUMBER KARBOHIDRAT
Sumber karbohidarat adalah padi-padian atau serealia, umbi-umbian, kacang-kacangan kering, dan gula. Hasil olahan bahan-bahan ibi adalah bihun, mie, roti, tepung-tepungan, selai/jam, siryp, permen, dodol, dll. Sebagian besar sayur dan buah tidak banyak mengandung karbohidrat dibandingankan dengan padi-padian. 1 penukar buah/sayur rata-rata mengandung 10gram karbohidart dibandingkan dengan kelompok padi-padian 1 penukarnya sebanyak 40gram karbohidrat. Sayur umbi-umbian, seperti wortel dan bit serta sayur kacang-kacangan relatif lebih banyak mengandung karbohidrat bila dibandingkan dengan jenis sayuran daun-daunan. Bahan makanan hewani seperti daging, ayam, ikan dan susu sedikit sekali mengandung karbohidrat. Sumber karbohidrat yang paling banyak dikonsumsi di Indonesia adalah beras, jagung, ubi, talas, singkong, dan sagu. Kandungan karbohidrat beberapa bahan makanan dapat dilihat pada tabel dibawah ini.
14
Makanan yang berasal dari hewan yang mengandung karbohidrat dalam jumlah cukup banyak adalah susu, tiram dan hati.
15
PENCERNAAN KARBOHIDRAT
Tujuan akhir pencernaan dan absorpsi karbohidrat adalah mengubah karbohidrat menjadi ikatan-ikatan yang lebih kecil, terutama berupa glukosa dan fruktosa, sehingga dapat diserap oleh pembuluh darah melalui dinding usus halus. Pencernaan karbohidrat yang tidak dicernakan akan memasuki usus besar untuk sebagian besar dikeluarkan dari tubuh.
Pencernaan Dalam Mulut
Pencemaan karbohidrat sudah dimulai sejak makanan masuk ke dalam mulut; makanan dikunyah agar dipecah menjadi bagian-bagian kecil, sehingga jumlah permukaan makanan lebih luas kontak dengan enzim-enzim pencernaan disebut dengan bolus. Bolus makanan bercampur dengan ludah/saliva yang mengandung enzim amilase (sebelumnya dikenal sebagai ptialin). Enzim amilase bekerja memecah atau menghidrolisis karbohidrat rantai panjang seperti amilum dan dekstrin menjadi bentuk karbohidrat yang lebih sedernaha, akan diurai menjadi disakarida maltosa. Sedangkan air ludah berguna untuk melicinkan bolus agar lebih mudah ditelan. Hanya sebagian kecil amilum yang dapat dicema di dalam mulut, oleh karena makanan sebentar saja berada di dalam rongga mulut. Oleh karena itu sebaiknya makanan dikunyah lebih lama, agar memberi kesempatan lebih banyak pemecahan amilum di rongga mulut. Enzim amilase ludah bekerja paling baik pada pH ludah yang bersifat netral. Dengan proses mekanik, makanan ditelan melalui kerongkongan dan selanjutnya akan memasuki lambung.
Pencernaan Dalam Lambung
Proses pemecahan amilum diteruskan di dalam lambung, selama makanan belum bereaksi dengan asam lambung. Bolus yang ditelan masuk ke dalam lambung, disini amilase ludah yangg ikut masuk ke lambung dicernakan oleh enzim pencerna protein yang terdapat di lambung, sehingga pencernaan karbohidrat di dalam lambung terhenti
Pencernaan Dalam Usus
Sebagian besar pencernaan karbohiddrat terjadi di dalam usus halus. Enzim amilase yang dikeluarkan oleh pankreas, mencerna pati menjadi dekstrin dan maltosa. Di usus halus, maltosa, sukrosa/sakarosa, dan laktosa yang berasal dari makanan maupun penguraian karbohidrat kompleks akan diubah menjadi mono sakarida. Penyelesaian pencernaan karbohidrat dilakukan oleh enzim-enzim disakaridase yang dikeluarkan oleh sel-sel mukosa
16
terjadi di dalam mikrovili dan monosakarida yang dihasilakan adalah sebagai berikut : maltase
Maltosa --->2 mol molekul glukosa sukrase
Sakarosa ---> 1 mol glukosa + 1 mol fruktosa laktase
Laktosa ---> 1 mol glukosa + 1 mol galaktosa
ABSORPSI KARBOHIDRAT
Semua jenis karbohidrat diserap dalam bentuk monosakarida, proses penyerapan ini terjadi di usus halus. Monosakarida glikosa, fruktosa, dan galaktosa kemudian diabsorpsi melalui sel epitel usus halus dan diangkut oleh sistem sirkulasi darah melaui vena porta. Bila konsentrasi monosakarida di dalam usus halus atau pada mukosa sel cukup tinggi, absorpsi dilakukan secara pasif atau fasilitatif. Tetapi, bila konsentrasi mrnutun, absorpsi dilakukan secara aktif melawan gradien konsentrasi dengan menggunakan energi dari ATP dan ion natrium. Glukosa dan galaktosa lebih cepat diabsorpsi dibandingkan fruktosa. Glukosa dan galaktosa memasuki aliran darah dengan jalan transfer aktif, sedangkan fruktosa dengan jalan difusi. Monosakarida melalui vena porta dibawa ke hati dimana fruktosa dan galaktosa diubah menjadi glukosa. Para ahli sepakat bahwa karbohidrat hanya dapat diserap dalam bentuk disakarida. Hal ini dibuktikan dengan dijumpainya maltosa, sukrosa dan laktosa dalam urine apabila mengkonsumsi gula dalam jumlah banyak. Akhimya Semua disakarida pada akhirnya diubah menjadi glukosa begitupula dengan berbagai jenis karbohidrat diubah menjadi glukosa sebelum diikut sertakan dalam proses metabolisme.
Berdasarkan urutan, yang paling cepat di absorpsi adalah galaktosa, glukosa dan
17
METABOLISME KARBOHIDRAT
Peranan utama karbohidrat di dalam tubuh adalah menyediakan glukosa bagi sel-sel tubuh, yang kemudian akan diubah menjadi energi. Glukosa memegang peranan utama dalam metabolisme karbohidrat.
Setelah proses penyerapan melalui dinding usus halus, sebagian besar monosakarida dibawa oleh aliran darah ke hati. Di dalam hati, monosakarida mengalami
proses sintesis menghasilkan glikogen, oksidasi menjadi CO2 dan H2O atau dilepaskan untuk
dibawa dengan aliran darah kebagian tubuh yang memerlukan (gambar 3). Sebagian lain monosakarida dibawa langsung ke sel jaringan organ tertentu dan mengalami proses metabolisme lebih lanjut. Karena pengaruh berbagai faktor dan hormon insulin yang dihasilkan oleh kelenjar pankreas, hati dapat mengatur kadar glukosa dalam darah. Bila kadar glukosa dalam darah meningkat sebagai akibat naiknya proses pencernaan dan
penyerapan karbohidrat, sintesis glikogen dari glukosa oleh hati akan naik. sehingga kadar
gula darah dapat dipertahankan dalam batas-batas normal (80-120 mg%).
Sebaliknya bila kadar glukosa menurun (misalkan akibat latihan olahraga) glikogen akan diuraikan menjadi glukosa untuk selanjutnya mengalami proses katabolisme menghasilkan
18
glukosa dalam darah merupakan faktor yang penting untuk kelancaran kerja tubuh. Gambar 3
Gambaran Umum Metabolisme Karbohidrat: Hubungan antara hati, darah, dan otot
HATI DARAH OTOT
Glikogen fruktosa galaktosa glukosa ATP Piruvat Lipida CO2 + H2O Sterol kolesterol fruktosa galaktosa glukosa ATP Piruvat laktat glikogen glukosa ATP Piruvat ATP Laktat CO2 + H2O
Karbohidrat yang terdapat dalam darah, praktis dalam bentuk glukosa, oleh karena fruktosa dan galaktosa akan diubah terlebih dahulu sebelum memasuki pembuluh darah.
Apabila jumlah karbohidrat yang dimakan melebihi kebutuhan tubuh, sebagian besar (2/3) akan disimpan di dalam otot dan selebihnya di dalam hati sebagai glikogen. Kapasitas pembentukan glikogen ini sangat terbatas (maksimum 350 gram), dan jika penimbunan dalam bentuk glikogen ini telah mencapai batasnya, kelebihan karbohidrat akan diubah menjadi lemak dan disimpan di jaringan lemak. Bila tubuh memerlukan kembali energi tersebut, simpanan glikogen akan dipergunakan terlebih dahulu, disusul oleh mobilisasi lemak. Jika dihitung dalam jumlah kalori, simpanan energi dalam bentuk lemak jauh melebihi jumlah simpanan dalam bentuk glikogen.
Sel-sel tubuh yang sangat aktif dan memerlukan banyak energi, mendapatkan energi dari basil pembakaran glukosa yang di ambil dari aliran darah. Kadar gula darah akan diisi kembali dari cadangan glikogen yang ada di dalam hati. Kalau energi yang diperlukan lebih banyak lagi, timbunan lemak dari jaringan lemak mulai dipergunakan. Dalam jaringan lemak diubah ke dalam zat antara yang dialirkan ke hati.
19 Skema Perubahan Karbohidrat Di dalam Tubuh
Disini zat antara itu diubah menjadi glikogen, mengisi kembali cadangan glikogen yang telah dipergunakan untuk meningkatkan kadar gula darah. Peristiwa oksidasi glukosa di dalam jaringan-jaringan terjadi secara bertahap dan pada tahap-tahap itulah energi dilepaskan sedikit demi sedikit, untuk dapat digunakan selanjutnya.
Melalui suatu deretan proses-proses kimiawi, glukosa dan glikogen diubah menjadi asam pyruvat. Asam pyruvat ini merupakan zat antara yang sangat penting dalam metabolisme karbohidrat. Asam pyruvat dapat segera diolah lebih lanjut dalam suatu proses pada "lingkaran Krebs". Dalam proses siklis ini dihasilkan CO2 dan H2O dan terlepas energi dalam bentuk persenyawaan yang mengandung tenaga kimia yang besar yaitu ATP (Adenosin Triphosphate). ATP ini mudah sekali melepaskan energinya sambi}berubah menjadi ADP (Adenosin Diphos phate). Sebagian dari asam piruvat dapat diubah menjadi "asam laktat". Asam laktat ini dapat keluar dari sel-sel jaringan dan memasuki aliran darah menuju ke hepar.
20
menjadi glikogen, dengan demikian akan menghasilkan energi.
Hal ini hanya terdapat di dalam hepar, tidak dapat berlangsung di dalam otot, meskipun di dalam otot terdapat juga glikogen. Sumber glikogen hanya berasal dari glukosa dalam darah. Metabolisme karbohidrat selain di pengaruhi oleh enzim-enzim, juga diatur oleh hormon-hormon tertentu. Hormon Insulin yang dihasilkan oleh "pulau-pulau Langerhans" dalam pankreas sangat memegang perananan penting. Insulin akan mempercepat oksidasi glukosa di dalam jaringan, merangsang perubahan glukosa menjadi glikogen di dalam sel-sel hepar maupun otot. Hal ini terjadi apabila kadar glukosa di dalam darah meninggi. Sebaliknya apabila kadar glukosa darah menurun, glikogen hati dimobilisasikan sehingga kadar glukosa darah akan menaik kembali. Insulin juga merangsang glukoneogenesis, yaitu mengubah lemak atau protein menjadi glukosa.
Juga beberapa horrnon yang dihasilkan oleh hypophysis dan kelenjar suprarenal merupakan pengatur-pengatur penting dari metabolisme karbohidrat. Enzim sangat diperlukan pada proses-proses kimiawi metabolisme zat-zat makanan. vitamin-vitamin sebagian dari enzim, secara tidak langsung berpengaruh pada metabolisme karbohidrat ini. Tiamin (vitamin B1) diperlukan dalam proses dekarboksilase karbohidrat. Kekurangan vitamin B1 akan menyebabkan terhambatnya enzim-enzim dekarboksilase, sehingga asam piruvat dan asam laktat tertimbun di dalam tubuh.
PEMBAGIAN METABOLISME KARBOHIDRAT
Secara lanjut proses metabolisme karbohidrat dibagi menjadi beberapa jalur metablisme, dimana merupakan jalur kesatuan, yang mana jalur yang paling banyak dilalui tergantung pada keadaan (situasi nutrisi) pada saat proses metabolisme karbohidrat berlangsung. Pembagiannya adalah :
1) Glikolisis ("glycolysis")
2) Glikogenesis ( "glycogenesis" ). 3) Glikogenolisis ( "glycogenolysis" ).
4) Oksidasi asam piruvat.
5) Jalur fosfoglukonat oksidatif ( "Hexose Mono-phosphate Shunt" atau "Pentose
Phosphate Pathway" ).
21
7) Metabolisme fruktosa, galaktosa dan heksosamin
Glikolisis
Glikolisis adalah pemecahan glukosa menjadi asam piruvat atau asam laktat. Jalur ini terutama terjadi dalam otot bergaris, yang dimaksudkan untuk menghasilkan energi (ATP). Apabila glikolisis terjadi dalam suasana anaerobik maka akan berakhir dengan asam laktat, dan mengha-silkan dua ATP, apabila dalam keadaan aerobik berakhir menjadi asam piruvat dengan 8 ATP.
Tahapan reaksi glikolisis:
Jalur ini disebut juga jalur Embden-Meyerhof. Semua enzim yang terlibat terdapat dalam fraksi ekstra mitokhondria (dalam sitosol). Mula-mula glukosa mengalami esterifikasi dengan fosfat, reaksi ini disebut juga fosforilasi glukosa oleh ATP menjadi glukosa 6-P.
Heksokinase (glukokinase) Mg++
22
sebagai glukosa bebas, sebagian besar terdapat dalam bentuk ester glukosa 6-P. Reaksi ini dikatalisis dua enzim : hexokinase dan glukokinase.
Hexokinase terdapat dalam ber-macam2 sel, kecuali di sel hepar dan pankreas. Enzim ini sesuai dengan namanya dapat pula mengkatalisis esterifikasi heksosa lainnya dengan ATP; contoh: fruktosa menjadi fruktosa 6-P. Dalam sel binatang dan manusia enzim ini merupakan enzim regulator, karena dapat dihambat oleh hasil reaksinya.
Glukokinase terdapat dalam hepar dan pankreas. Mempunyai Km untuk D-glukosa jauh lebih tinggi dari enzim hexokinase. Glukokinase memerlukan glukosa lebih tinggi untuk menjadi aktif bila dibandingkan dengan heksokinase.
Berbeda dengan heksokinase glukokinase tidak dihambat oleh hasil reaksinya yaitu glukosa 6-P. Glukokinase berperan biasanya pada waktu kadar glukosa darah tinggi (sesudah makan). Pada penderita Diabetes Mellitus enzim ini jumlahnya berkurang. Reaksi fosforilasi ini boleh dikatakan reaksi satu arah.
Selanjutnya glukosa 6-P diubah menjadi fruktosa 6-P. Reaksi ini dikatalisis enzim fosfoheksosa isomerase, dimana terjadi aldosa-ketosa isomerasi. Hanya D-anomer dari glukosa 6-P yang bisa dipakai sebagai substrat. Reaksi ini merupakan reaksi bolak-balik.
Reaksi selanjutnya adalah pembentukan fruktosa 1,6-difosfat oleh enzim fosfofruktokinase-1 (PFK-1). Reaksi ini boleh dikatakan reaksi satu arah. Enzim fosfofruktokinase-1 merupakan enzim yang bisa diinduksi. Enzim ini memegang peran yang penting dalam mengatur kece-patan glikolisis.
fosfofruktokinase-1
Fruktosa 6-P + ATP --->Fruktosa 1,6-BP + ADP. Mg++
Aktifitas enzim ini meningkat apabila konsentrasi ADP, AMP, fosfat inorganik (Pi) meningkat. Enzim fosfofruktokinase-1 dihambat oleh ATP, asam sitrat dan 2,3-DP gliserat (dalam sel darah merah). Apabila pemakaian ATP meningkat (kadar ATP menurun) maka aktifitasnya meningkat, sebaliknya apabila kadar ATP tinggi aktifitas enzim tersebut menurun. Enzim ini juga dihambat oleh meningkatnya kadar asam lemak bebas, sehingga apabila senyawa ini meningkat dalam darah, yang akhirnya masuk ke dalam sel , maka pemakaian glukosa akan
23
berkurang. Keadaan ini bisa terjadi pada waktu kelaparan, yang mana juga dapat terbentuk senyawa keton.
Peran fruktosa 2,6 bisfosfat (dalam hepar).
Dalam hepar fruktosa 2,6-bisfosfat merupakan allosterik efektor positif yang paling kuat bagi enzim fosfofruktokinase-1, dan merupakan inhibitor bagi enzim fruktosa 1,6-bisfosfatase ("enzim glukoneogenesis"). Fruktosa 2,6-bisfosfat menghilangkan pengaruh hambatan (inhibisi) ATP terhadap fosfofruktokinase-1, dan meningkatkan affinitas enzim ini terhadap fruktosa 6-P. Fruktosa 2,6-bisfosfat menghambat fruktosa-1,6-bisfosfatase dengan jalan meningkatkan harga Km untuk fruktosa-1,6-bisfosfat. Kadar fruktosa 2,6-bisfosfat dibawah pengaruh kontrol substrat dan hormonal.
Fruktosa 2,6-bisfosfat dibentuk dengan fosforilasi fruktosa 6-fosfat (fruktosa 6-P)
yang dikatalisis enzim fosfofruktokinase-2 (PFK-2). Enzim yang sama bertanggung jawab juga
terhadap pemecahan fruktosa 2,6 bisfosfat (F 2,6-BP), karena enzim ini mempunyai aktifitas fruktosa 2,6 bisfosfatase, namun enzim ini telah mengalami fosforilase menjadi PFK-2P (fosfo frukto kinase fosfat). Aktifitas bifungsi enzim fosfofruktokinase-2 ini dibawah pengaruh (kontrol) allosterik fruktosa 6-P. Apabila kadar senyawa ini meningkat, sebagai akibat meningkatnya kadar glukosa, maka akan meningkatkan aktifitas fosfofruktokinase-2 pada waktu yang sama aktifitas fosfatasenya menurun. Apabila kita memerlukan glukosa
(pada waktu puasa), dengan perkataan lain glukosa darah menurun, glukagon akan
menyebabkan terbentuknya cAMP, selanjutnya "cAMP- dependent protein kinase" teraktifasi, yang menyebabkan fosfofruktokinase-2 dihambat, sedangkan aktifitas fruktosa 2,6BPase meningkat, karena diubah menjadi PFK-2P.
Dalam keadaan dimana kadar glukosa meningkat, menyebabkan fruktosa 6P meningkat, ini memacu PFK-2 membentuk fruktosa 2,6 bisfosfat. Selanjutnya F 2,6 BP akan merangsang akti-fitas fosfofruktokinase-1 dan menghambat fruktosa 1,6 bisfosfatase. Fruktosa 1,6-BP akan dipecah menjadi dua triosa oleh enzim aldolase.
Aldolase
Fruktosa 1,6-BP → Dihidroksi asetonfosfat + gliseraldehida 3-P
(Pada sel binatang sedikitnya ada dua macam aldolase, aldolase A yang terdapat dalam se-bagian besar jaringan , aldolase B terdapat dalam sel hepar dan ginjal. Semuanya terdiri dari empat subunit polipeptida yang berbeda komposisi asam aminonya).
24
Kedua triosa tersebut diatas "interconverted", dapat saling berubah dengan adanya enzim fosfotriosa isomerase. Sampai dengan reaksi ini satu glukosa terpakai dan memerlukan dua ATP.
Selanjutnya glikolisis berjalan dengan oksidasi gliseraldehid 3-fosfat (gliseraldehida 3-P) menjadi 1,3-bisfosfogliserat. Karena adanya enzim fosfotriosa isomerase, dihidroksi asetonfos-fat juga dioksidasi.Enzim yang bertanggung jawab pada reaksi ini adalah
gliseraldehida 3-P de-hidrogenase yang mana aktifitasnya tergantung adanya NAD+. Enzim
ini terdiri dari empat polipeptida yang identik membentuk tetramer. Empat gugusan -SH terdapat pada tiap polipep-tida, mungkin berasal dari residu sistein (cysteine). Satu gugusan
-SH terdapat pada "active site". Reaksinya berjalan mula-mula substrat berikatan
dengan "cysteinyl moiety" pada dehidrogenase membentuk suatu tiohemiasetal, yang
kemudian dioksidasi menjadi tiol-ester. Atom hidrogen yang terlepas dipindah pada NAD+
yang terikat pada enzim. NADH yang terbentuk,akan terikat pada enzim juga tapi tidak
sekuat NAD+, sehingga NADH ini mudah diganti oleh NAD+ yang lain.
Energi yang terjadi pada oksidasi ini terwujud dalam ikatan sulfat energi tinggi, yang ke-mudian dengan fosforolisis menjadi ikatan fosfat energi tinggi pada posisi satu dari 1,3-bisfosfo-gliserat.Pada fosforolisis diatas, Pi ditambahkan dan enzim bebas serta gugus -SH be-bas terbentuk. Fosfat berenergi tinggi ini ditangkap menjadi ATP pada reaksi dengan ADP yang dikatalisis enzim fosfogliserat kinase. Reaksi ini menghasilkan 3-fosfogliserat.
Jadi oksidasi fosfogliseraldehid menjadi fosfogliserat, dimana terlepas suatu energi, energi ini dipakai oleh reaksi pengambilan fosfat inorganik dan sintesis ATP; rangkaian reaksi-reaksi
ini merupakan suatu "coupled reaction". Karena ada dua molekul triosafosfat yang
dioksidasi, maka akan terbentuk dua molekul ATP. Pada reaksi ini NAD+ tereduksi menjadi
NADH. Reaksi tersebut diatas adalah suatu con-toh dari fosforilasi pada tingkat substrat.
Apabila ada asam arsenat, maka zat ini akan berkompetisi dengan Pi yang akan
menghasilkan arseno-fosfogliserat, yang akan terhidrolisis spontan menghasilkan 3-fosfogliserat tanpa menghasilkan ATP. Ini suatu contoh arsenat dapat "uncoupled" oksidasi dan fosforilasi. Selanjutnya 3-fosfogliserat diubah menjadi 2-fosfogliserat oleh enzim
25
perubahan struktur molekul hingga terbentuk ikatan fosfat bertenaga tinggi pada posisi 2,
yaitu fosfoenolpiruvat. Enolase dihambat oleh fluorida ( F ). Dalam praktek fluorida
ditambahkan ke dalam larutan pada penentuan glukosa,juga kedalam pasta gigi. Kerja enzim
ini tergantung adanya Mn++ atau Mg++. Reaksinya sebagai berikut:
2-fosfogliserat ↔ Fosfoenolpiruvat + H
2O.
Fosfat bertenaga tinggi dari fosfoenolpiruvat dipindah ke ADP menjadi ATP, yang
di-katalisis enzim piruvat kinase.
Reaksinya:
ADP ATP
Fosfoenolpiruvat Enolpiruvat
Piruvat kinase
Enzim piruvat kinase hepar berbeda sifatnya dengan enzim piruvat kinase otot. Pada otot konsentrasi ATP yang tinggi akan menghambat enzim ini. Pada hepar enzim ini dapat dihambat oleh ATP dan alanin, tapi adanya fruktosa 1,6-difosfat dengan konsentrasi tinggi, akan dapat menghilangkan hambatan ini. Dalam hepar enzim ini dihambat juga oleh asam lemak rantai panjang dan asetil-KoA.
Dalam hepar glukagon menghambat glikolisis dan merangsang glukoneogenesis
dengan meningkatkan konsentrasi cAMP. Senyawa ini kemudian mengaktivasi "cAMP dependent protein kinase". Protein kinase yang aktif ini akan mengkatalisis fosforilasi enzim piruvat kinase menjadi piruvat kinase-P. Enzim piruvat kinase-P merupakan bentuk tidak aktif. Dengan demikian glukagon menghambat glikolisis.
Sampai dengan reaksi ini hasil netto dari perubahan glukosa menjadi dua asam piruvat adalah dua NADH dan dua ATP, yaitu pada awal jalur ini dibutuhkan dua ATP dan kemudian menghasilkan empat ATP. Dalam keadaan aerobik NADH dengan menggunakan rantai respirasi dapat diubah menjadi 3 ATP.
Pada keadaan anaerobik reoksidasi NADH melalui rantai respirasi tidak berjalan.
Asam piruvat akan dirubah menjadi asam laktat, yang dikatalisis enzim laktat
26
laktat dehidrogenase
Asam piruvat + NADH L-laktat + NAD+
Dengan demikian reoksidasi NADH melalui asam laktat memungkinkan glikolisis
berlang-sung tanpa oksigen, karena NAD+ yang terbentuk cukup untuk kebutuhan enzim
gliseraldehid-3-fosfat dehidrogenase. Jadi jaringan pada keadaan hipoksia ada tendensi untuk membentuk asam laktat, terutama dalam otot bergaris. Asam laktat yang terbentuk akan masuk ke peredaran darah dan bisa didapatkan dalam urine.
Sel Darah Merah
Glikolisis dalam eritrosit sekalipun dalam keadaan aerobik akan menghasilkan asam laktat, karena enzim-enzim yang dapat mengoksidasi asam piruvat secara aerobik tidak ada dalam sel darah merah. Dalam eritrosit golongan mammalia tahapan yang dikatalisis
fosfogliserat kinase di " by passed " dengan adanya enzim bisfosfogliserat mutase dan enzim
2,3-bisfosfogliserat fosfatase. Akibat adanya dua enzim ini ATP tidak terbentuk dan ini memungkinkan glikolisis berlangsung apabila kebutuhan ATP minimum. 2,3-bisfosfogliserat bergabung dengan hemo-globin sehingga menyebabkan affinitas hemoglobin terhadap oksigen menurun. Kurve dissosi-asi oksigen hemoglobin bergerak ke kanan. Dengan demikian adanya 2,3-bisfosfogliserat dalam sel darah merah membantu pelepasan oksigen untuk keperluan jaringan.
Reaksinya :
Enzim 1
1,3-bisfosfogliserat → 2,3-bisfosfogliserat Enzim 2 ↓
3-fosfogliserat. Enzim 1 : bisfosfogliserat mutase
27
Dalam glikolisis ada tiga reaksi yang dapat dikatakan secara fisiologis satu arah, yaitu reaksi yang dikatalisis enzim-enzim :
1. heksokinase ( dan glukokinase ) 2. fosfofruktokinase
3. piruvat kinase
Oksidasi Asam Piruvat Menjadi Asetil-CoA
Asam piruvat dapat masuk ke dalam mitokhondria dengan pertolongan suatu transporter. Asam piruvat mengalami oksodasi-dekarboksilasi oleh suatu enzim yang tersusun rapi dalam matriks mitokhondria. Enzim-enzim ini disebut piruvat dehidrogenase komplek
Mula-mula asam piruvat mengalami dekarboksilasi. Reaksi ini dikatalisis enzim piruvat dehidrogenase. Tiamin pirofosfat bertindak sebagai ko-enzim. Dalam reaksi ini
terbentuk CO2 dan α-hidroksietil-tiaminpirofosfat atau disebut juga "aktif asetaldehid".
Senyawa yang disebut belakangan ini dipindah ke prostetik lipoamide, yang merupakan bagian dari enzim transasetilase. Dalam perpindahan ini disulfida dari lipoamide tereduksi, asetildehida teroksidasi menjadi asetil aktif yang terikat sebagai tioester. Gugusan asetil ini kemudian bereaksi dengan koenzim-A, membentuk asetil-S-KoA, dan menghasilkan lipoamide dalam bentuk disulfhidril (tereduksi). Koenzim yang tereduksi ini dioksidasi kembali oleh suatu flavoprotein, dihidrolipoil dehidrogenase. Flavoprotein yang tereduksi
kemudian dioksidasi oleh NAD+. Ringkasnya, reaksinya adalah sebagai berikut:
CH3COCOOH + HSCoA + NAD+ → CH3CO-SCoA + NADH + H+
Piruvat dehidrogenase dihambat oleh hasil reaksinya yaitu NADH dan asetilKoA. Enzim ini juga dihambat oleh aktivitas oksidasi asam lemak, yang mana akan meningkatkan
rasio Asetil-KoA/KoA, NADH/NAD+ dan ATP/ADP. Peningkatan rasio diatas akan mengaktivasi
piru-vat dehidrogenase (PDH) kinase yang akan mengkatalisis fosforilasi enzim PDH a menjadi PDH b yang tidak aktif. PDH fosfatase akan menghidrolisis PDH b menjadi PDH a yang aktif. PDH fosfatase diaktivasi oleh insulin. Arsenit atau ion merkuri membentuk komplek dengan gugusan -SH dari asam lipoat dan menghambat piruvat dehidrogenase. Kekurangan tiamin akan menyebabkan asam piruvat tertimbun.
28
Glikogenesis
Pembentukan glikogen (glikogenesis) terjadi hampir dalam semua jaringan, tapi yang pal-ing banyak adalah dalam hepar dan dalam otot. Setelah seseorang diberi diet tinggi karbo-hidrat (hidrat arang), kemudian heparnya dianalisis , maka akan didapatkan kurang lebih 6% berat basah terdiri dari glikogen. Namun 12 sampai 18 jam kemudian, hampir semua gliko-gen habis terpakai. Dalam otot kandungan glikogen jarang melebihi satu persen, tapi untuk menghabiskan glikogen tersebut agak sulit, yaitu misalnya dengan olah raga berat dan lama.
Sintesis glikogen dimulai dengan perobahan glukosa 6-fosfat menjadi glukosa 1-fosfat yang dikatalisis enzim fosfoglukomutase (glukosa 1,6-bis1-fosfat bertindak sebagai koenzim). Selanjutnya enzim uridin difosfat glukosa pirofosforilase (UDPG pirofosforilase) meng-katalisis pembentukan uridin difosfat glukosa (UDP-glukosa).
UTP + Glukosa 1-fosfat → UDP-glukosa + Ppi
Reaksi ini boleh dikatakan reaksi searah,karena hidrolisis senyawa inorganik
pirofosfat menjadi inorganik fosfat, yang dikatalisis enzim inorganik pirofosfatase menarik
reaksi kekanan. Enzim glikogen sintetase (glikogen sintase) memindahkan glukosil aktif dari
UDP-glukosa (UDPG) pada bagian dari ujung glikogen yang tidak dapat direduksi, mem-bentuk ikatan α-1,4 glukosidik. Pemmem-bentukan ikatan tersebut terjadi ber-ulang2, sehingga cabangnya makin panjang. Apabila panjang cabang tersebut mencapai antara 6 sampai 11,
maka enzim amilo (α 1,4) α (1,6) transglukosidase ("branching enzim") memindahkan
se-bagian dari residu ikatan α-1,4 (minimum 6 residu), pada rantai didekatnya membentuk ika-tan α-1,6. Jadi terjadi titik percabangan baru. Kemudian kedua cabang tersebut bertambah panjang. Dan seterusnya kejadian berulang kembali.
Uridin difosfat yang dibebaskan ketika unit glukosil dari UDPG dipindah kebagian tertentu dari glikogen, disintesis kembali menjadi UTP dengan memakai ATP. Total kebutuhan ATP untuk menyimpan satu molekul glukosa menjadi satu molekul glikogen adalah dua molekul, dua ADP dan dua inorganik fosfat terbentuk. Berat molekul glikogen mencapai satu sampai empat juta lebih
29
Glikogenolisis
Pemecahan glikogen dalam hepar dan otot berbeda dengan enzim yang terdapat dalam pen-cernaan. Enzim glikogen fosforilase akan melepaskan unit glukosa dari rantai cabang gliko-gen yang tidak bisa direduksi. Reaksinya bisa digambarkan sebagai berikut:
(Glukosa)n + H
3PO4 → Glukosa 1-fosfat + (Glukosa)n-1
Enzim ini hanya memecah ikatan α-1-4 glikosidik, dan berhenti pada empat residu
dari titik cabang. Enzim amilo (α 1,4)(α 1,4) glukan transferase, memindah tiga unit
glukosa yang terikat pada rantai cabang (yang tinggal empat) pada rantai yang lain membentuk “rantai” lurus. Selanjutnya enzim glikogen fosforilase.akan memecah ikatan α-1,4 sampai 4 unit glukosa dari titik cabang, demikian seterusnya.
Debranching enzim (amilo 1,6-glukosidase) memecah ikatan glukosidik 1,6 dan menghasil-kan glukosa. Dalam otot glukosa yang dihasilkan tidak cukup banyak untuk dieksport keluar sel, kemungkinan dipakai oleh sel otot itu sendiri. Glukosa 1-fosfat yang
terlepas diubah menjadi glukosa 6-fosfat oleh enzim fosfoglukomu-tase. Senyawa ini bisa
masuk jalur glikolisis atau jalur lainnya. Di hepar, ginjal dan epitel usus halus glukosa
6-fosfatase yang spesifik memecah ikatan ester dan melepaskan glukosa ke peredaran darah. Enzim ini tidak didapatkan dalam otot.
Glikogenesis dan Glikogenolisis, Mekanisme dan Kontrol
Pada prinsipnya enzim yang mengatur metabolisme glikogen adalah glikogen
fosforilase dan glikogen sintase, enzim-enzim ini sendiri dibawah pengaruh suatu kontrol yang kom-plek yaitu suatu mekanisme yang melibatkan peristiwa allosterik dan modifikasi ikatan kovalen pada senyawa fosfat dari enzim.
Aktifitas dan Inktiffitas Forforilase
Dalam hepar, enzim fosforilase ada dalam keadaan aktif maupun tidak aktif. Pada fosfori-lase yang aktif (fosforilase a), gugusan hidroksil dari serin mengalami fosforilasi (dalam ikatan ester). Fosforilase a (yang aktif) bisa menjadi tidak aktif dengan
hilangnya fosfat yang terikat pada senyawa serin tersebut. Reaksi ini dikatalisis enzim
fosfatase spesifik dengan nama protein fosfatase-1. Untuk mengaktifkan enzim
fosforilase kembali diperlu-kan refosforilasi, yang dapat dikatalisis enzim fosforilase
kinase dengan adanya ATP. Fosforilase otot, berbeda secara immunologik dan genetik bila dibandingkan dengan fos-forilase hepar. Dalam otot fosforilase a merupakan
30
tergantung ada atau tidak adanya AMP. Tiap monomer mengandung satu piridoksal fosfat. Fosforilase b, yang mengalami defosforilasi hanya aktif apabila ada AMP. Dalam keadaan fisiologis fosforilase a merupakan bentuk aktif enzim enzim ini.
Aktifasi melalui cAMP
Fosforilase dalam otot dapat diaktifasi oleh epinefrin secara tidak langsung. Aktifasi ini melalui cAMP. cAMP merupakan suatu senyawa intra selluler, senyawa ini merupakan suatu senyawa antara ("intermediate compound"). Senyawa ini disebut juga "second mes-senger". Banyak hormon yang bekerja dengan perantaraan
senyawa ini. cAMP dibentuk dari ATP oleh enzim adenilil siklase sebelumnya dikenal
dengan nama adenilat siklase (adenylate cyclase), yang terdapat pada permukaan dalam membran sel. Adenilil siklase dapat diaktifasi oleh hormon-hormon seperti: epinefrin dan norepinefrin yang bekerja melalui reseptor adrenergik beta. Reseptor ini terletak pada sel membran.
Pada hepar glukagon bekerja melalui reseptor yang lain yaitu reseptor glukagon.
cAMP dirusak enzim fosfodiesterase; dengan adanya enzim ini kadar cAMP diatur
dalam kadar yang rendah. Insulin dapat meningkatkan aktifitas enzim fosfodiesterase
dalam hepar, dengan demikian menyebabkan kadar cAMP rendah.
Meningkatnya cAMP menyebabkan meningkatnya aktifitas enzim protein kinase "cAMP-dependent", yang mempunyai spesifisitas luas.
Protein kinase ini mengkatalisis fosforilasi oleh ATP, enzim fosforilase kinase b (tidak aktif) menjadi fosforilase kinase a (aktif), yang selanjutnya juga dengan proses fosforilasi fosforilase kinase a yang aktif mengkatalisis perubahan fosforilase b menjadi fosforilase a.
Protein kinase "cAMP-dependent" yang tidak aktif terdiri dari dua pasang subunit. Tiap pasang terdiri dari suatu subunit regulator (R), yang dapat mengikat dua cAMP, dan sub-unit katalitik (C), yang mengandung "active site" membentuk R2C2. Apabila bereaksi dengan cAMP maka kompleks protein kinase yang tidak aktif itu akan berdisosiasi dan melepaskan monomer C yang aktif.
31
R2C2 + 4cAMP ↔ 2C + 2(R-cAMP2)
enzim enzim
inaktif aktif
Aktifasi melalui Ca++ dan sinkronisasi dengan kontraksi otot.
Beberapa saat setelah kontraksi otot dimulai glikogenolisis meningkat beberapa ratus kal lipat. Hal ini disebabkan karena aktifitas fosforilase yang diaktifkan oleh
fosforilasekinase. Enzim fosforilase kinase sendiri diaktifkan oleh Ca++, yang mana
juga merupakan signal untuk mengaktifkan kontraksi. Fosforilase kinase otot
terdiri dari 4 subunit, α, β, γ dan δ yang membentuk struktur (αβγδ)4. Subunit α dan β mengandung residu serin yang dapat difosforilasi oleh protein kinase
"cAMP-dependent". Subunit β dapat mengikat 4 Ca++. Subunit β identik dengan kalmodulin
protein (suatu "Ca++ binding protein").Terikatnya Ca++ pada subunit β dapat
mengaktifkan "catalytic site γ" biarpun enzim fosforilase kinase ini dalam bentuk defosforilasi (fosforilase kinase b). Akan tetapi fosforilase kinase a hanya akan
mempunyai aktivitas maksimal apabila telah mengikat Ca++. TpC adalah Ca++ binding
protein dalam otot, strukturnya mirip struktur Calmodulin. Calmodulin atau TpC dapat
berinteraksi dengan fosforilase-kinase (yang telah mengikat Ca++) dan menyebabkan
aktivasi lebih lanjut. Jadi aktifasi kontraksi otot dan glikogenolisis dijalankan oleh
kalmodulin yang sama. Efek Ca++ dalam sel dapat terlihat berkat adanya "the Ca++
binding protein" kalmodulin
Glikogenolisis Dalam Hepar
Penelitian menunjukkan bahwa selain pengaruh aktivitas glukagon melalui reseptornya, glikogenolisis dalam hepar juga dirangsang oleh katekolamin
(adrenalin) melalui proses yang melibatkan mobilisasi Ca++ dan tidak tergantung pada
cAMP (cAMP-independent mobilization of Ca++) dari mitokhondria ke sitosol.
Selanjutnya terjadi rangsangan fos-forilase kinase yang sensitif terhadap
32
bergaris adalah, protein phosphatase-1 hepar dihambat oleh fosforilase yang aktif. Inaktivasi fosforilase.
Fosforilase a dan fosforilase kinase a dapat dibuat tidak aktif oleh protein phosphatse-1 dengan jalan melepaskan gugusan fosfatnya (dephosphorylated). Protein phosphatase-1 sendiri dapat dihambat oleh suatu protein yang disebut inhibitor-1. Inhibitor-1 hanya aktif apabila sudah mengalami fosforilasi oleh cAMP-dependent protein kinase menjadi inhibi-tor-1-P. Dengan demikian cAMP dapat mengontrol aktivasi maupun inaktivasi dari phos-phorilase.
Aktivasi dan Inaktivasi “Glycogen Synthhase” (glikogen sintase)
Seperti halnya fosforilase, glikogen sintase berada dalam bentuk fosforilasi maupun defos-forilasi. Akan tetapi bentuk aktif glikogen sintase (glikogen sintase a) adalah defosforilasi (yang telah kehilangan senyawa fosfat ). Bentuk ini bisa diubah menjadi glikogen sintase b (tidak aktif) dengan jalan fosforilasi residu serin (7 buah). Reaksi ini memerlukan ATP dan dikatalisis oleh sedikitnya 6 macam protein kinase. Glikogen sintase terdiri dari 4 sub unit yang identik. Tiap-tiap sub unit mempunyai 7 buah residu serin yang dapat difosforilasi. Enam macam protein kinase tersebut diatas,
terdiri dari dua protein kinase "Ca++/Calmodulin dependent"(satu diantaranya adalah
fosforilase kinase). Kemudian ada satu protein kinase yang cAMP-dependent. Protein kinase ini memungkinkan suatu hormon menghambat sintesis glikogen dan mengaktifkan glikogenolisis secara sinkron. Sedangkan protein kinase yang lain dikenal dengan nama glikogen sintase kinase-3, kinase-4 dan kinase-5. Glukosa-6P merupakan suatu allosterik aktivator dari glikogen sintase b dengan jalan menurunkan Km untuk UDP-glukosa. Dengan demikian bentuk yang tidak aktif (glikogen sintase b yaitu bentuk yang mengalami penambahan fosfat atau bentuk yang terfosforilasi) dapat menjalankan glikogen sintesis.Glikogen dapat menghambat sintesis glikogen itu sendiri. Insulin dapat merangsang sintesis glikogen dalam otot,dengan jalan meningkatkan defosforilasi (pelepasan fosfat) dan aktivasi glikogen sintesis b. Dalam keadaan normal defosforilasi glikogen sintesa b dilakukan oleh protein phosphatase-1, yang mana berada dibawah kontrol cAMP-dependent protein kinase.
33
Regulasi Metabolisme Glikogen
Regulasi metabolisme glikogen dipengaruhi oleh keseimbangan antara enzim glikogen sintase dan fosforilase. Kedua enzim tersebut berada dibawah kontrol substrat (melalui al-losterik) dan hormon. Melalui cAMP-dependent protein kinase, fosforilase diaktifkan (dengan jalan meningkatkan cAMP) dan glikogen sintase aktif diubah menjadi tidak aktif pada waktu yang bersamaan. Jadi inhibisi glikogenolisis akan meningkatkan glikogenesisdan penghambatan terhadap glikogenesis akan menyebabkan peningkatan glikogenolisis. Hal lain yang perlu diperhatikan dalam regulasi metabolisme glikogen adalah peristiwa defosforilasi dari fosforilase a, fosforilase kinase dan glikogen sintase b dilakukan oleh satu protein dengan spesifisitas yang luas yaitu protein phosphatse-1. Selanjutnya protein phosphatase-1 dihambat oleh inhibitor-1-P. Inhibitor-1-P berasal dari inibitor-1 yang mengalami fosforilasi dan dikatalisa oleh "cAMP-dependent protein kinase". Dengan demikian glikogenolisis bisa dihentikan dan pada waktu yang bersamaan glikogenesis dirangsang, demikian juga sebaliknya, hal ini disebabkan karena adanya cAMP-dependent protein kinase. Baik fosforilase kinase maupun glikogen sintase secara reversibel dapat difosforilasi pada lebih dari satu tempat dengan bantuan kinase dan fosfatase yang berbeda. Fosforilasi yang kedua ini mengubah sensitivitas fosforilasi dan defosforilasi yang pertama ( Enzim piruvat dehidrogenase juga menunjukkan adanya "multisite phosphorylation")
Hepar
Faktor utama yang mengatur metabolisme glikogen dalam hepar adalah konsentrasi
fos-forilase a. Enzim ini tidak hanya merupakan "control the rate-limiting step" dalam gliko-genolisis, akan tetapi dapat juga menghambat aktivitas protein phosphatase-1 dengan demikian dapat mengontrol sintesis glikogen. Penghambatan aktivitas
fosforilase terjadi sebagai akibat inhibisi allosterik oleh glukosa yang meningkat
setelah makan. Sedangkan aktivasi fosforilase oleh 5`-AMP terjadi sebagai akibat menurunnya kadar ATP. Glukagon merupakan hormon utama yang menyebabkan kenaikan cAMP dalam hepar, yang berakibat meningkatnya glikogenolisis. Sebagian besar rangsangan katekolamin (adrenalin) melalui mekanisme yang tidak melibatkan cAMP akan tetapi melalui α1-reseptor adrenergik. Proses ini melalui rangsangan
34
melalui kalsium atau jalur fosfatidilinositol bis-fosfat. Insulin menyebabkan inaktivasi fosforilase secara cepat yang diikuti oleh aktivasi glikogen sintase. Pengaruh ini bisa berlangsung apabila ada glukosa.
Glikoneogenesis
Glukoneogenesis
Glukoneogenesis adalah suatu pembentukan glukosa dari senyawa yang bukan karbohidrat. Glukoneogenesis penting sekali untuk menyediakan glukosa, apabila didalam diet tidak mengandung cukup karbohidrat. Syaraf, medulla dari ginjal, testes, jaringan embriyo dan eritrosit memerlukan glukosa sebagai sumber utama penghasil energi. Glukosa diperlukan oleh jaringanadiposa untuk menjaga senyawa antara siklus asam sitrat. Didalam mammae, glukosa diperlukan untuk membuat laktosa. Didalam otot, glukosa merupakan satu-satunya bahan untuk membentuk energi dalam keadaan anaerobik. Untuk membersihkan darah dari asam laktat yang selalu dibuat oleh sel darah merah dan otot, dan juga gliserol yang dilepas jaringan lemak, diperlukan suatu proses atau jalur yang bisa memanfaatkannya. Pada hewan memamah biak, asam propionat merupakan bahan utama untuk glukoneogenesis.
Jalur yang dipakai dalam glukoneogenesis adalah modifikasi dan adaptasi dari jalur Emb-den Meyerhof dan siklus asam sitrat.
Enzim tambahan yang diperlukan dalam proses ini selain dari enzim-enzim dalam kedua jalur diatas adalah :
Piruvat karboksilase
Fosfoenolpiruvat karboksikinase
Fruktosa 1,6-bisfosfatase (tidak ada dalam otot jantung dan otot polos)
Glukosa 6-fosfatase
Dalam keadaan puasa, enzim piruvat karboksilase dan enzim fosfoenolpiruvat karbok-sikinase sintesisnya meningkat. Sintesis enzim ini juga dipengaruhi oleh hormon glukokor-tikoid. Dalam keadaan puasa, oksidasi asam lemak dalam hepar meningkat. Ini membawa akibat yang menguntungkan untuk glukoneogenesis karena akan menghasilkan ATP, NADH dan oksaloasetat. Asam lemak dan asetil-KoA akan menghambat enzim-enzim
35
fosfofruktokinase, piruvat kinase dan piruvat dehidrogenase, mengaktifkan enzim-enzim piruvat karboksilase dan fruktosa 1,6-bisfosfatase.
Substrat untuk glukoneogenesis adalah :
Asam laktat yang berasal dari otot, sel darah merah, medulla dari glandula
supra-renalis, retina dan sumsum tulang
Gliserol, yang berasal dari jaringan lemak
Asam propionat, yang dihasilkan dalam proses pencernaan pada hewan memamah
biak.
Asam amino glikogenik
Perubahan Asam Laktat Menjadi Glukosis
Asam laktat di dalam sitoplasma diubah menjadi asam piruvat, kemudian asam piruvat ma-suk ke dalam mitokhondria dan diubah menjadi oksaloasetat. Karena oksaloasetat tidak dapat melewati membran mitokhondria, maka diubah dulu menjadi malat. Di sitoplasma malat diubah kembali menjadi oksaloasetat. Oksaloasetat kemudian diubah menjadi fosfoenolpiruvat yang selanjutnya berjalan ke arah kebalikan jalur Embden-Meyerhof dan akhirnya akan menjadi glukosa.
Asam amino glukogenik masuk ke dalam jalur glukoneogenesis ditandai dengan bundaran dan panah pada siklus asam tri karboksilat ( TCA cycle ).Beberapa reaksi dan enzim-enzim tambahan untuk mengubah asam laktat menjadi glukosa (selain jalur kebalikan glikolisis dan TCA cycle) adalah :
Enzim piruvat karboksilase mengkatalisis reaksi
Piruvat → Oksaloasetat (gambar 15-16)
Dalam reaksi ini diperlukan ATP, CO
2 (berasal dari H2CO3), biotin ( yang diperlukan
untuk mengikat bikarbonat pada enzim sebelum ditambahkan pada asam piruvat ) dan ion Mg.
Enzim fosfoenolpiruvat karboksikinase mengkatalisis reaksi :
Oksaloasetat → Fosfoenolpiruvat
Dalam reaksi ini diperlukan "high energy phosphate" GTP atau ATP, dan akan terbentuk CO
2.
36
Fruktosa 1,6-bisfosfat → Fruktosa 6-fosfat
Enzim ini bisa didapatkan dalam hati, ginjal otot bergaris, sedangkan jaringan lemak, otot jantung dan otot polos tidak mengandung enzim fruktosa 1,6-bisfosfatase.
Enzim glukosa 6-fosfatase mengkatalisis reaksi :
Glukosa 6-fosfat → Glukosa
Enzim ini terdapat dalam usus halus, hati, ginjal dan platelet, akan tetapi tidak bisa dijumpai dalam otot dan jaringan lemak
Enzim gliserokinase mengkatalisis reaksi :
Gliserol → Gliserol 3-fosfat
Dalam reaksi ini diperlukan ATP dan menghasilkan ADP. Enzim ini terutama terdapat dalam hati dan ginjal.
Enzim gliserol 3-fosfat dehidrogenase mengkatalisis reaksi : Gliserol 3-fosfat → Dihidroksi aseton fosfat ( DHAP )
Asam propionat perlu diaktivasi dahulu menjadi propionil-KoA. Ensim tiokinase
mengkatalisis reaksi ini dan memerlukan ATP , KoA dan ion Mg. Selanjutnya propionil-KoA diubah menjadi D-metilmalonil-KoA, selanjutnya setelah mengalami rasemisasi akan diubah menjadi L-metilmalonil-KoA. Senyawa ini kemudian akan diubah menjadi suksinil-KoA yang akan masuk ke dalam siklus asam sitrat yang akhirnya akan diubah menjadi glukosa melalui kebalikan jalur Embden-Meyerhof
Hexose Monophosphate Shunt (HMP Shunt) = Pentose Phosphate Pathway (PPP) Oksidasi Glukosa Langsung = Jalur Fosfoglukonat
Jalur ini aktif dalam hepar, jaringan adiposa (lemak), adrenal korteks, glandula tiroid, sel darah merah,testes dan payudara yang sedang menyusui. Dalam otot aktivitas jalur ini ren-dah sekali. Fungsi utama jalur ini adalah untuk menghasilkan NADPH, yaitu dengan
mereduksi NADP+. NADPH diperlukan untuk proses anabolik di luar mitokhondria, seperti
sintesis asam lemak dan steroid. Fungsi yang lain adalah menghasilkan ribosa-5-fosfat untuk sintesis nukleotida dan asam nukleat. Jalannya reaksi sebagai berikut: