ABSTRAK
Busuk batang Sclerotium merupakan salah satu penyakit penting pada tanaman kacang tanah, yang seringkali menyebabkan kehilangan hasil yang tinggi. Penggunaan varietas yang resisten terhadap penyakit ini merupakan cara yang paling efisisen untuk mengontrol penyakit ini. Upaya untuk mendapatkan galur kacang tanah yang toleran atau resisten terhadap penyakit busuk batang. Sclerotium melalui induksi keragaman somaklonal dan seleksi in vitro menggunakan filtrat kultur S. rolft;; telah dilaporkan sebelurnnya. Kalus embriogenik yang telah dipelihara selama kurang Iebih satu tahuo dalam media regenerasi dikulturkan dalam medium selektif dengan penambahan 30% filtrat kultur S. rolfsii, dan ES yang terbentuk setelah tiga periode seleksi in vitro masing-masing selama satu bulan, dianggap sebagai insensitif terhadap filtrat kultur cendawan. Sejumlah galur kacang tanah RC, telah diregenerasikan dari embrio somatik yang insensitif terhadap filtrat kultur cendawan, dan zuriat Rl dan R2-nya telab ditanam di rurnab plastik dan dievaluasi untuk berbagai karakter kualitatif dan kuantitatifnya. Evaluasi Wltuk resistensi terhadap infeksi S. rolft;; juga telab dilakukan secara dini pada galur-galur somaklon generasi RO, yang mengindikasikan peningkatan ketahanannya dibandingkan dengan populasi tanaman kacang tanah awal. Namun demikian masih dipedukan evaluasi respons galur-galur somaklon tersebut pada generasi berikutnya, untuk. mengetahui kestabilan karakter resistensi yang telah diperoleh sebelumnya Penelitian ini bertujuan untuk mengevaluasi respons ketahanan galur-galur somaklon kacang tanah hasil seleksi in vitro generasi Rl dan R2 terhadap infeksi S. rolfsii di rumah kaca. Hasil penelitian ini menunjukkan bahwa telab didapatkan sejurn1ab galur somaklon kacang tanah zuriat dari somaklon RO yang resisten terhadap infeksi S. rolfsii. Sifat resistensi ini terdapat diantara populasi somaklon Rl dan R2, yang
bersegregasi antara resisten dan rentan terbadap infeksi S. rolfs;;. Somaklon kacang tanah yang resisten terhadap infeksi S. rolfsii pada awalnya menunjukkan gejala penyakit busuk batang setelah diinokulasi dengan S. rolfsii, dengan skor antara 1-3, tetapi daJam perkembangannya tanaman menunjukkan proses penyembuhan luka pada lesio, dan tanaman mampu menghasilkan polong bemas yang sarna atau lebih banyak dibandingkan tanaman kacang tanah awal yang tidak diinokulasi.
Kata kunci: Variasi somaklonai, seleksi in vitro, Sclerotium rolfsii. filtrat kultur endawan, resistensi terhadap penyakit. kacang tanah.
IN VITRO SELECTED PEANUT SOMA CLONAL VARIANTS USING FUNGAL CULTURE FILTRATES RESISTANT TO Sclerotium rolfsii
ABSTRACT
Sclerotium stem rot is one of the most important peanut disease which often caused significant yield loss. The use of peanut cultivars resistant to Sclerotium rolfsii infection is the most efficient way to control the disease. Attempts to obtain peanut lines with tolerance or resistance to Sclerotium stem rot through induction of somaclonal variation and in vitro selection using fungal culture filtrates (CF) have been conducted previously. Somatic embryo (SE) clumps that had been maitained in culture for approximately one year in regeneration medium were exposed onto selective medium containing 30% S. rolfsii CF for three consecutive 1 month -passages. and the SE fanned after selection periods were considered to be insensitive to the fungal CF. A number RO peanut lines have been regenerated from CF-insensitive SE, and their Rl and R2 progenies were grown in a plastic house for evaluation on qualitative and quantitative variant characters. Early identification for tolerance to S. rolftii was also conducted among RO peanut lines, and the results have shown enhanced resistance when compared to the original non-selected cultivar. However, further evaluation is needed to study responses of the RI and R2 progenies of the sornaclones against S. rolftii infection. The objective of this particular study was to evaluate responses of RI and R2 peanut somac1ones derived from fungal CF-insensitif SE against S. rolft;; infection in the plastic house. Results of this experiment showed, a number of resistant somaclonal variants were obtained among RI and R2 population, which segregated as resistance and susceptible to S. rolfsii infection. The resistance somaclonal variants initially showed stem rot symptoms after inoculation with S. rolft;; with disease score (DS) of 1 to 3. However, as the plants grew and developed, they showed wound-healing process at the lesio and they were able to produce the same or higher number of filled pod as the original non-inoculated peanut plants.
Key words: Somac1onal variation, in vitro selection, Sclerotium rolftii. fungal culture filtrates, disease resistance, peanut.
PENDAHULUAN
Kemungkinan untuk menginduksi variasi somaklonal diantara tanaman yang diregenerasikan melalui kultur sel dan jaringan telab menimbulkan minat yang tinggi untuk menggunakannya sebagai salah satu cam untuk membantu pemuliaan tanaman (Larkin dan Scowcroft 1981). Beberapa penelitian terdahulu tentang induksi variasi somaklonal dan seleksi in vitro yang ditujukan untuk karakter resistensi terhadap penyakit telah berhasil mendapatkan plasma nutfah yang taleran, lebih resisten atau resisten terhadap penyakit teItentu (Scowcroft et al. 1985, Wenzel 1985, Daub 1986, Hammerschlag 1992).
Pemuliaan tanaman untuk sifat resistensi terhadap penyakit tertentu merupakan salah satu cara untuk melindungi tanaman dari kerusakan oleh faktor abiotik. khususnya oleh mikroorganisme patogen. Karakter resisten terhadap penyakit yang dapat ditunmkan sangat penting dalam budidaya tanaman karena dapat menghemat biaya produksi secara signifikan. Beberapa penyakit tanaman sangat merusak dan tersebar di areal pertanaman yang sangat luas, sehingga menjadi prioritas utama bagi pemulia tanaman untuk mendapatkan varietas baru yang resisten.
Busuk batang Sclerotium merupakan salah satu penyakit penting pada tanaman kacang tanab, yang seringkali menyebabkan kehilangan basil yang tinggi. Penyakit ini disebabkan oleb cendawan tular-tanab Sclerotium rolfsii Sacco Penyakit yang menyebabkan kebusukan pada pangkal batang kacang tanab dan seringkali penyebab kematian atau kerusakan yang parab ini relatif sulit diberantas, karena cendawan patogennya mempunyai inang beragam dan dapat
membentuk sklerosia yang mampu bertahan hidup di dalam tanah dalam waktu lama (Backman 1984; Punja 1985).
Varietas atau genotipe kacang tanah yang ada di Indonesia tidak ada yang dilaporkan tahan terhadap infeksi cendawan ini (Rani 2001, Widyanti 2001, Yusnita dan Sudarsono 2004). Upaya untuk mendapatkan plasma nutfah kacang tallah yang toleran atau resisten terhadap penyakit busuk batang Sclerotium
melalui induksi keragaman somaklonal
dan
seleksi in vitro menggunakan filtrat kultur S. rolfsii telah dilaporkan sebelumnya (Yusnita ef al. 2004). Dalam penelitian tersebut, kalus embriogenik yang telab dipelihru:a selama kurang lebih satu tabun dalam media regenerasi dikulturkan dalam medium MS (Murashige dan Skoog 1962) dengan penambahan 30% fi1trat kultur S. rolfsii, dan ES yang terbentuk setelah tiga periode seleksi in vitro masing-masing selama satu bulan, dianggap sebagai insensitif terhadap filtrat kultur cendawan.S<jumlah galur kacang tanah RO telah diregenerasikan dari embrio somatik yang insensitif terhadap filtrat kultur cendawan, dan zuriat RI dan R2-nya telah ditanam di rumah plastik dan dievaluasi untuk berbagai karakter kualitatif dan kuantitatifnya (Yusnita ef al. 2004). HasH penelilian tersebut memmjukkan bahwa terdapat bemgam karakter varian morfologis. fisiologis dan agronomis diantara tanaman somaklon hasil seleksi in vitro. Evaluasi untuk. resistensi terhadap infeksi S. rolfsii juga telah dilakukan secara dini pada galur-galur somaklon generasi RO, yang mengindikasikan peningkatan ketabanannya dibandingkan dengan populasi tanaman kacang tanah awal (Yusnita ef al. 2004). Penelitian ini bertujuan untuk. mengevaluasi respons ketahanan galur-galur
somaklon kacang tanah hasil seleksi in vitro generasi Rl dan R2 terhadap infeksi S rolfsii di rumah plastik.
BAHAN DAN METODE
Regenerasi ES yang Insensitif Filtrat Kultur Cendawan Menjadi Galur-galur Tanaman RO.
Dari hasil seleksi in vitro1500 clump kalus embriogenik menggunakan 30% filtrat kultur cendawan, telah didapatkan sebanyak kurang lebihl-2 ES insensitif filtrat kultur cendawan
pada
sekitar 300 clump. Perbandingan jumlah ES hasil seleksi in vitro tersebut hanya 1-5% dari ES tanpa seleksi in vitro (Yusnita ef al. 2004). Untuk proses pemulihan dan proliferasinya, ES basil seleksi in vitro ditransfer ke media MS (Murashige dan Skoog 1962) dengan penambahan pikloram 16 ~M (P16). Selanjutnya, maturasi dan pengeeambahan ES dilakukan dengan mentransfemya ke media MS dengan penambahan arang aktif 2 gil (MSAC) selama periode pengulturan, masing-masing satu bulan. Untuk merangsang pertumbuhan epikotil dan pemanjangan tunas, ES yang berkecambab dipotong akamya dan ditanam selama kurang lebih 2 minggu dalam medium MS dengan penambahan kombinasi BAP 2 mgll dan kinetin 2 mgll.Untuk mendapatkan planlet, tunas yang didapat ditanam kembali dalam media MSAC hingga berakar. Aklimatisasi planlet dilakukan secara bertahap dengan menanamnya dalam pot plastik berisi campuran pasir:arang sekam steril (l: I, v/v) yang disungkup dengan kantong plastik, hingga mampu bertaban hidup di rumah plastik. Tanaman RO yang mampu bertahan hidup dari tabapan aklimatisasi selanjutnya ditanam dalam polybag dengan ukuran 45cm x 45 em
yang berisi 10 kg campuran tanab:pasir:kompos (1:1:1, v/v) dan dipelihara di rumah plastik. Penyiraman, pemupnkan (pupuk majemnk NPK [15: 15: 15]), dan pengendalian hama, penyakit dan gulma dilakukan sesuai kebutuhan hingga tanaman dapat dipanen. Benih RO:l dipanen secara terpisah dari masing-masing Domor tanaman RO dan dikeringkan.
Penanaman T anaman 8omakloD Rl dan R2 di Rumah Plastik.
Tanaman Rl ditumbuhkan dari benih RO:l zuriat dari 25 tanaman RO yang masing-masing diregenerasikan dari ES hasil seleksi in vitro. Dari setiap nomor tanaman RO ditanam I hingga 20 tanaman Rl zuriatnya, bergantung dari jurnlab polong bemas yang dihasilkan sebelumnya. Penanaman benih RO:I di rumab plastik dan pemeliharaannya dilaknkan dalam po/ybag sebagaimana dijelaskan sebelumnya untnk penanannan tanaman RO. Tanaman Rl dipelihara hingga panen, dan benih RI:2 yang dihasilkan dipanen secara terpisab dari masing-masing Domor tanaman RI. kemudian dikeringkan dan pada musim berikutnya ditanam di rumah plastik sebagaimana tanaman RI.
Tanaman R2 diturnbuhkan dari benih RI :2, zuriat dari masing-masing Domor tanaman Rl terpilih. Dari setiap tanaman Rl dipilih satu Domor yang menghasilkan polong bemas paling hanyak, dan dari masing-masing nomor R I tersebut ditanam minimal 10 tanannan R2. T anaman R2 diturnbuhkan di rurnab plastik dengan cara dan pemeliharaan sebagaimana dijelaskan sebehunnya untnk tanaman RO dan Rl. Tanaman R2 dipelihara hingga panen, benih R2:3 yang dihasilkan dipanen seeara terpisah dari masing-masing nomOl" tanarnan R2. lalu dikeringkan.
Tananum kacang tanah cv. Kelinci (tanaman awal) yang tidak melalui tahapan seleksi in vitro ditumbuhkan dari benih di rumah plastik menggunakan polybag dengan media tumbuh dan eara pemeliharaan yang sarna dengan sebelumnya. Tanaman awal tersebut digunakan sebagai pembanding uDtuk tanaman· RD. Sebagai pernbanding untuk tanarnan RI dan R2, ditanam zuriat pertama dan kedua tanaman awal dengan cara, pemeliharaan dan dalam lingkungan yang sarna.
Pembuatan Isolat S. rOlfsii dan Inokulasi pada Tanaman Somaklon Rl dan R2.
Cendawan S. rolfsii yang virulensinya telah diuji, diisolasi dari tanaman kacang tanah terinfeksi Sclerotium di Kebun Percobaan IPB - Babakan Sawah Baru, Dannaga, Bogor. Isolat cendawan diperbanyak dalam media potato dextrose agar (PDA) dan diinkubasi dalam ruang bersubu 26
'c
selama 6--7 hari, sehingga mempunyai koloni hifa yang tebal tetapi belum membentuk sklerosia. Hifa dari kultur eendawan dalam media PDA pada! seluas 1 em' dignnakan sebagai inokulurn untuk menginokulasi tanaman kacang tanah generasi RI dan R2. Tanaman somaklon Rl dan R2 dan tanaman kacang tanah cv. Kelinci awal sebagai kontrol diinokulasi pada umur 4 minggu setelah tanam.Pada generasi RI, inokulasi hanya dilakukan pada masing-masing 3 tanaman un!uk 5 nomor galur RO (SC+INO), dan 2 tanaman ev. Kelinci awaJ sebagai kontrol (SD+INO). SejumJab 144 tanaman Rl hasil seleksi in vitro yang lain tidak diinokulasi (SC-INO), sedangkan 8 tanaman ev. Kelinei awal digunakan sebagai kontrol tanpa inokulasi (SD-INO).
Pada generasi R2, inokulasi dilakukan pada setiap nomor galllr Rl yang ditanam. masing-masing 5 dari 10 tanaman (SC+INO). Lima tanaman sisanya dari masing-masing galur Rl digunakan sebagai kontrol somaklon tanpa inokulasi eSC-INO). Sebanyak 12 tanaman cv. Kelinci awal digunakan sebagai kontro1 yang diinoku1asi (SD+INO), dan 10 tanaman tanpa inoku1asi (SD-INO).
Inokulasi pada semua tanaman yang diuji dilakukan dengan carn menusuk pangkal batang, masing-masing sebanyak: 5 tusukan dengan jarum steril, kemudian menempelkan inokulum berupa miselia dan menimbunnya dengan tanah.
Skoring Gejala Peoyakit akibat Iofeksi S. ro/fsii
Pengamatan gejala penyakit dilakukan 14 hari selelah tanaman diinokulasi untuk jumJah Skor gejala penyakit busuk batang pada tanaman kacang tanah yang diipokuiasi dicatat dengan mengamati perkembangan infeksi cendawan yang terjadi pada pangkal batang tanaman yang diujj. Gejala infeksi S. rolfsii yang berkembang diberi nilai skor antara 0--5, sebagaimana dijelaskan oleh Yusnita dan Sudarsono (2004). Krileria nilai skor gejala penyakil busuk batang
Sclerotium adalah: skor 0 = tanaman sehat, tidak menunj ukkan gejala, skor 1 =
gejala nekrosis dengan luasan hingga Y: lingkar batang, skoT 2 = gejala nekrosis
antara %-% lingkar batang. skor 3 = gejala nekrosis telah melingkari batang,
bercak cokelat telah meluas, dan kulit batang kadang-kadang sobek, skor 4 ~ seperti skor 3, batang yang terserang mulai terkulai dan sebagian daun layu, dan skor 5 = tanaman mati.
Pengamatan untuk Karakter Kuantitatif setelah Inokulasi.
Pada semua tanaman yang diuji. baik yang tidak diinokulasi maupun yang diinokulasi (kecuali untuk tanaman yang mati setelah diinokulasi) dilakukan pengamatan untuk peubah kuantitatif. yang meliputi tinggi tanaman, jumlah c.bang p.d. batang utam., jumlab polong cipo dan jumlab polong bemas per tanamao. Pengamatan untuk peubah kuantitatif tersebut dilakukan setelah tanaman dipanen.
HASIL
Perkembangan Gejala Penyakit Busuk Hatang setelab Diinokulasi S. rOlfsii Semua tanaman somaklon yang diinokulasi menunjukkan gejala penyakit dengan tingkat keparahan yang berbeda antar. skor 1 sampai 5, sedangkan seroua tanaman awal yang diinokulasi mati (skor 5). Baik tanaman awal maupun somaldon yang tidak diinokulasi tidak menunjukkan gejal. penyakit (skor 0).
Secara umum, gejala mulai muneul 5-8 hari setelah inokulasi. Gejal. perry
pangkal batang yang diinokulasi busuk b.tang dimulai dengan munculnya lesio pada batang yang diselim ti miseHa S. rolfsii berwarna putih. Lesio tersebut semakin melebar dengan erjalannya waktu setelah inokulasi disertai dengan pembusukan pennukaan b tang. Bertambab luasny. lesio pada batang diikuti oleh terjadinya perubahan w a batang pada tepi lesio dari coklat muda menjadi cold.t tua (Gambar 25,). P hari 10-14, bintik-bintik slderoti. berwarn, putih kecold.tan mulai te ntuk di pennukaan b.tang yang membusuk. Setelah pembusukan batang m luas dan pembentukan sklerosia dari
hi fa celldawan leIjadi, sebagian daun pada cabang utama muJai layu (Gambar 2Sb). Pada sejumlah somaklon dan semua tanaman awal yang diinokuJasi. gejaJa tersehUl berlanjut dengan kematian tanaman (Gambar 2Sc). Pada sejumlah
tanaman somaklon yang diinokulasi. gejaJa penyakit busuk batang telap mW1cul, tetapi lama·kelamaan miselia cendawan tidak berkembang dan pembusukan batang tidak berlanjul. Tanaman somaklon tersebut terinfeksi, tetapi menunjukkan resistensi terhadap penyakit yang ditandai dengan tcrjadinya proses penyembuhan luka pada lesio di bagian batang yang terinfeksi, seiring dengan pertumbuhan taoaman. Proses penyembuhan luka pada lesio terse but ditandai
dengan pembentukan kaJus, mirip seperti pada balang yang dicangkok. Di
samping iw. pembentukan bunga, ginofor dan polong bernas telap terjadi pada y tanaman-tanaman somaklon terinfeksi, terutama pada tanaman bergejaJa penyakil
Gambar 25. Perkembangan gejala penyakit busuk batang pada pangkal batang kacang lanah setelah diinokulasi dengan Sclerotium rof/sii. (a) Lesio pada pangkaJ batang telah meluas dan berubah dari coklat muda menjadi coklal lua, (b) sebagian daun pada cabang utama layu, (c) tanaman mati.
dengan skor 1-3. Tanaman dengan skor 4 umumnya tidak. mampu menghasilkan poiong bernas, walaupun masih bidup hingga saat paneD.
Dari setiaplO tanaman somaklon R2 zuriat dari masing-galur Rl terpilih, lima tanaman yang tidak diinokulasi ,emuanya tidak bergejala penyakit (,kor 0), sedangkan 5 tanaman yang diinokulasi menunjukkan gejala beragam mulai dari skor 1 hingga skor 5. Representatif keragaan tanaman R2 zuriat dari beberapa galur RI terpilih setelah diinokulasi dapat dilihat pada Gambar 26a-c. Semua tan3man awal yang diinokulasi menunjukkan gejala penyakit dan pada hari ke-14 setelah inokulasi, semua tanaman
mfti
(Gambar 26d).Respons Somaklon Rl terhadap Inokulasi S. rolfsii
Re,pons tanaman ,omaklon RI zuriat dari 6 galur tanaman RO terhadap inokulasi S. rolftii disajikan pada Tabel IS. Pada Tabel 15 terlibat bahwa semua tanaman aWal yang tidak diinokulasi (SD-INO) maupun ,emua tanaman ,omaklon yang tidak diinokulasi (SC-INO) menunjukkan ,kor 0 atau tidak menunjukkan gejala penyakit, sedangkan semua tanaman awal yang diinokulasi (SD+INO) menunjukkan gejala penyakit dengan ,kor 5, atau tanaman mati. Sebaliknya, tanaman somaklon yang diinoknlasi (SC+INO) menunjukkan gejala penyakit dengan ,kor beragam mulai dari ,kor 2 (4 tanaman), skor 3 (6 tanaman), 'kor 4 (4 tanaman), dan ,kor 5 (4 tanaman).
Respons SomakIon R2 terhadap Inokulasi S. rolfsii
Respons tanaman somaklon R2 zuriat dari setiap galur tanaman Rl terpilih terhadap inokulasi S. rolftii di,ajikan pada Tabel 16. Pada Tabel 16
Gambar 26. Representatif keragaan somakloll R2 zuriat dari tiga galur RI
terpilih sereiah diinokulasi dcngan S. rolfsii. (a) Lima tanaman R2
zuriat galur Rl no. K4.3-SC setelah diinokulasi, tiga diantaranya masih hidup dengan skar 3, sedangkan dua tanaman mati dengan skor 5, (b) Lima tanaman R2 zuriat galur Rl no. K24.1-SC setelah
diinokulasi, dua diantaranya masih hidup dengan skor 2 dan 3, sedangkan tiga tanaman mati dengan skar 5, (e) Lima tanaman R2
zuriat galur R 1 no. K26.8-SC, setelah diinokulasi scmuanya mati
dengan skar S. (d) Dua belas
lanaman
kacang tanah cv. Kelinciterlihat bahwa semua tanaman awal yang tidak diinokulasi (SD-INO) maupun semua tanaman somaklon yang tidak. diinokulasi (SC-INO) menunjukkan skor 0 atau tidak bergejala penyakit, sedangkan semua tanaman awal yang diinokulasi (SD+INO) menunjukkan gejaia penyakit dengan skor 5, atau tanaman mati. Sebaiiknya, pada 158 tanaman .omaklon yang diinoknlasi (SC+INO) diamati gejala penyakit dengan skor beragam mulai dari skor 1 (13 tanaman), skor 2 (15 tanaman), ,kor 3 (38 tanaman), .kor 4 (8 tanaman), dan .kor 5 (84 tanaman). Seeara lehih rinei, respons somaklon R2 zuriat dari masing-masing nomOr RI terpilih terhadap inokulasi S. rolfsii berdasarkan skor gejala penyakit yang muncul pada hari ke 14 .etelab inokulasi dapat dilihat pada Tabel1?
Tabel15. Respens tanaman Rl-zuriat dari galur somaklon RO hasil seleksi in vitro eSC) terhadap inokulasi dengan S rolfsii dibandingkan dengan tanaman kacang tanah awal (SD) yang tidak melalui seleksi in vitro.
Frekuensi tanaman awal (SO) Frekuensi somaklon (SC) Skor gejala berdasarkan skor gejala pen~akit berdasarkan skor gejala Eentakit
penyakit Tanpa Setelah inokulasi Tanpa Setelah inokulasi busuk batang inokulasi dengan S. rolfsii inokulasi dengan S. rolJsii
(SD-INO) (SD+INO) (SC-INO) (SC+INO)
0 8 0 144 0 1 0 0 0 0 2 0 0 0 4 3 0 0 0 6 4 0 0 0 4 5 0 2 0 4 Jumlab tanaman 8 2 144 18 yang diamati
Tabel 16. Respons tanaman R2-zuriat dari galur somaklon RI terpilih hasil seleksi in vitro (SC) terhadap inokulasi dengan S. rolfsii dibandingkan dengan tanaman kacang tanah awal (SD) yang tidak melalui seleksi in vitro. Skor gejaJa penyakit busuk batang
o
2 3 4 5 Jumlah tanaman yang diamatiFrekuensi tanaman awal (SD) berdasarkan skor gejala penyakit
Tanpa Setelah inokulasi inokulasi dengan S. rolfsii
(SD-INO) (SD+INO) 10
o
o
o
o
o
o
o
o
o
o
12 10 12 Frekuensi somaklon (SC) berdasarkan skor gejala penyakitTanpa Setelah inokulasi inokulasi dengan S. roIfsii (SC-INO) (SC+INO) 152
o
o
13o
15o
38o
8o
84 152 158Keragaan Agronomis Tanaman R2 dengan dan taopa Inokulasi S. rolfsii Rataan tinggi tanaman (TD, jumlah eabang utama (JCU), jumlah poloug eipo (JPC), dan jumlah polong bemas (JPB) per tanaman diantara tanaman awal (kacang tanab ev. Kelinei, SD) dan tanaman R2 zuriat tanaman RO hasil seleksi in vitro tanpa inokulasi (SC), atau setelah diinokulasi dengan 8.rolfsii (SC+INO) disajikan pada Gambar 27. Pada Gambar 27 terlihat hahwa mtaan tinggi tanaman dan jumlah polong eipo diantam somaklon R2 zuriat galur Rl terpilih tanpa inokulasi (SC) atau setelah diinokulasi (SC+INO) sedikit lebih tinggi daripada rataan tinggi tanaman awal (SD). Sebaliknya, mtaan jumlah cabang utarna hampir sarna antara tanaman awal dengan tanaman somaklon R2 tanpa inokulasi, dan sedikit lebih rendah pada tanaman somaklon R2 setelah diinokulasi S. rolfsii.
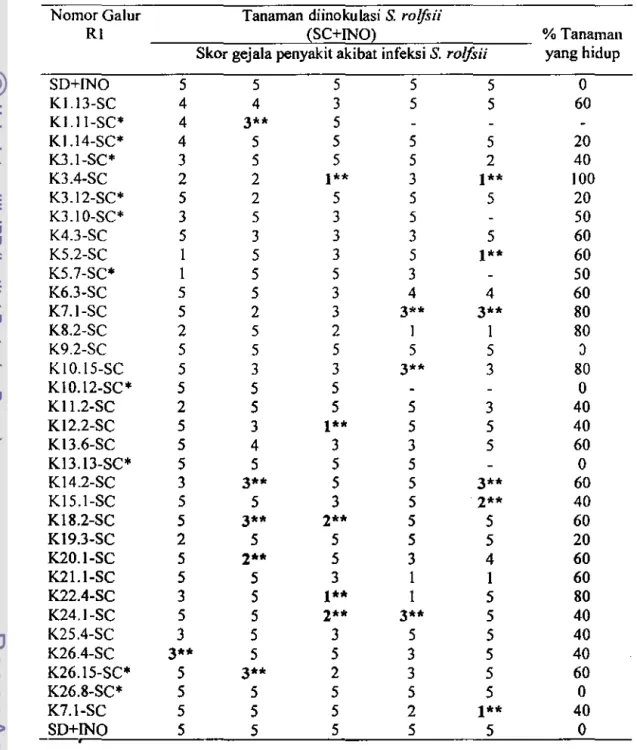
Tabel 17. Respons masing-masing galur Rl terhadap inokulasi S. rolfsii berdasarkan skor gejala penyakit yang ditimbulkan pada zuriat R2-nya dibandingkan dengan kacang tanah CV. Kelinci sebagai tanaman
awal.
Tanaman diinokulasi S. rolfsii
NomorGalur
Rl _ _ -,;;:==:;:-:=c'("'SC,,+::IN""'o':)==,-;;-==,---_ % Tanaman Skor gejala penyakit akibat infeksi S. ro/fsii yang hidup
SD+INO 5 5 5 KI.13-SC 4 4 J Kl.l1-SC* 4 3** 5 5 5 5 5
o
60 K1.l4-SC* 4 5 5 5 5 20 KJ.l-SC* 3 5 5 5 2 40 K3.4-SC 2 2 1"'* 3 1** 100 KJ.12-SC* 5 2 5 5 5 20 KJ.IO-SC* J 5 J 5 50 K4.J-SC 5 J J 3 5 60 KS.2-SC I 5 J 5 1" 60 KS.7-SC* I 5 5 J 50 K6.J-SC 5 5 J 4 4 60 K7.I-SC 5 2 3 3** 3** 80 K8.2-SC 2 5 2 I I 80 K9.2-SC 5 5 5 5 5 J KIO.15-SC 5 J 3 3" J 80 KIO.12-SC· 5 5 5 0 KlI.2-SC 2 5 5 5 J 40 KI2.2-SC 5 J 1" 5 5 40 K13.6-SC 5 4 J J 5 60 KIJ.13-SC· 5 5 5 5 0 K14.2-SC 3 3** 5 5 3** 60 KI5.1-SC 5 5 J 5 2" 40 KlS.2-SC 5 3** 2** 5 5 60 KI9.J-SC 2 5 5 5 5 20 K20.1-SC 5 2" 5 J 4 60 K21.1-SC 5 5 J I 1 60 K22.4-SC 3 5 1" I 5 80 K24.I-SC 5 5 2** 3** 5 40 K25.4-SC 3 5 3 5 5 40 K26.4-SC 3" 5 5 3 5 40 K26.15-SC* 5 3" 2 3 5 60 K26.8-SC· 5 5 5 5 5 0 K7.1-SC 5 5 5 2 I " 40SD+~~~O~--~5~_~5~--~5~--~5---~5---~0--*) Nomar tanaman yang diinokulasi pada generasi Rl dan R2. **) Skor gejala pada "tanaman R2 zwiat dari galur Rl terpi1ih yang menunjukkan resistensi terhad.p S. rolftii dan menghasilkan polong bemas hampir sama atau lebih banyak daripada tanaman awal yang tidak diinokulasi. Semua tanaman yang tidak diinokulasi (SD-lNO dan SC-lNO) tidak menunjukkan gejala penyakit (skor gejaI.~O).
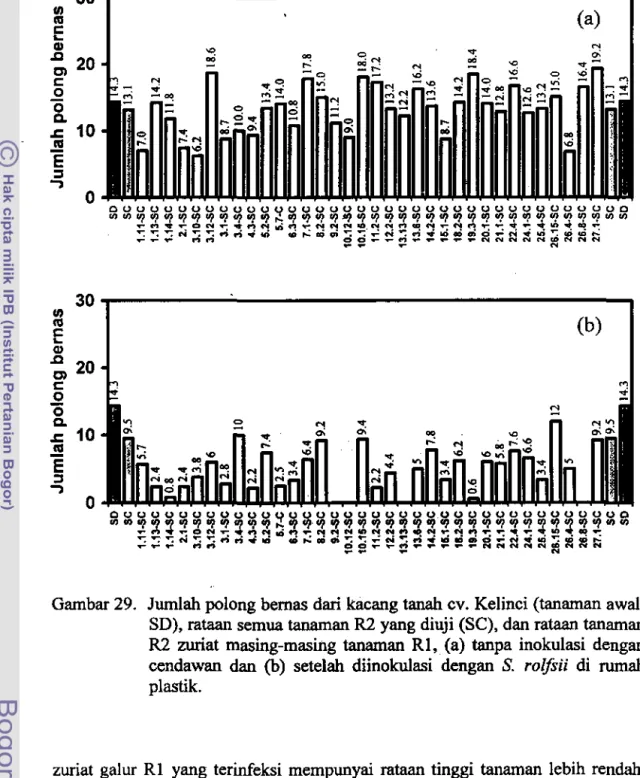
Rataan jumlah polong bemas somaklon R2 tanpa inokulasi sedikit lebih rendah dibandingkan dengan tanaman awal, sedangkan rataan jumlah polong bemas pada tanaman sornaklon R2 setelah diinokulasi lebih rendah dari tanaman awal maupun somaklon tanpa inokulasi.
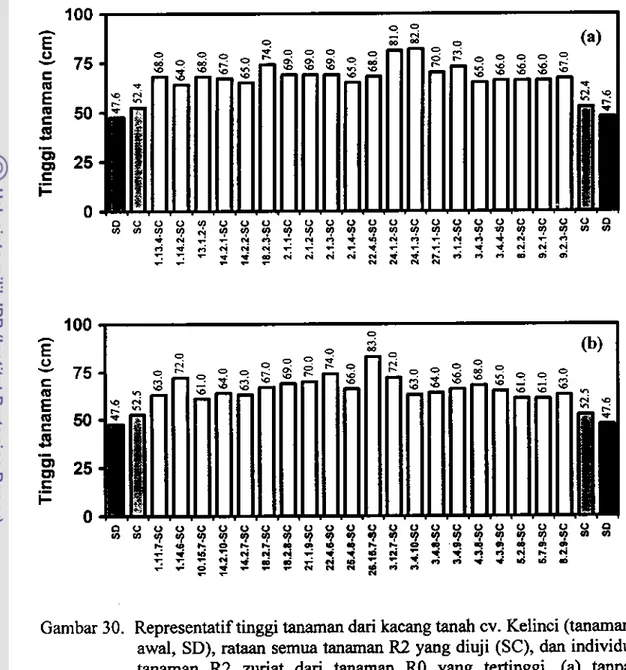
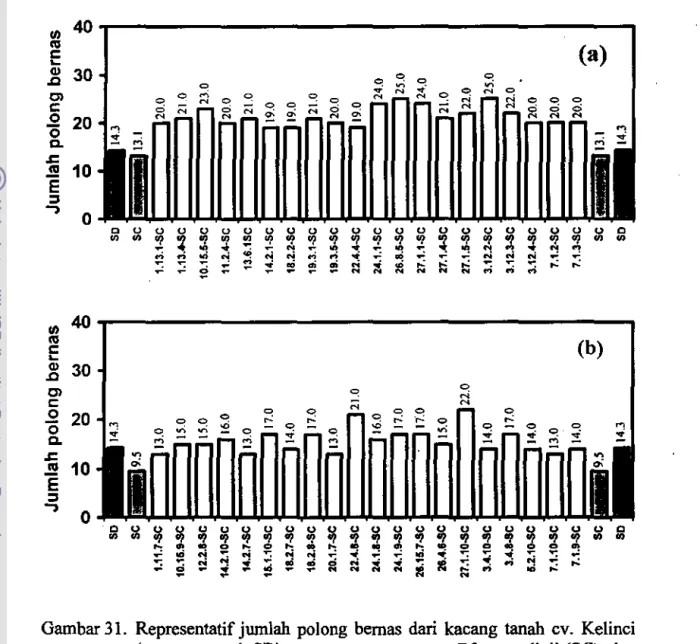
Data ini dipeIjelas dengan rataan tinggi tanaman (Gambar 28) dan jumlah polong bemas (Gambar 29) pada somaklon R2 zuriat masing-masing tanaman R 1, tanpa inokulasi dan setelah diinokulasi dengan S. rolfsii dibandingkan dengan rataan tinggi tanaman dan jumlah poiong bemas dari tanaman awal dan dari ,emua tanaman R2 yang diuji. Di sarnping itu, pada Gambar 30 dan Gambar 31 disajikan representatif tinggi tanaman dan jumlah polong bemas dari tanaman awat, rataan semua tanaman R2 yang diuji, dan individu tanaman R2 zuriat dari tanarnan RO yang tertinggi, baik tanpa perlakuan inokulasi cendawan maupun setelah diinokulasi dengan S. rolfsii.
75
.SO !!ISC CSC+INO 52.4 52.5 50 25 4.' 5.2 3 .• 4.8 5.5 0 TT JCU JPC JPB
Gambar 27. Rataan tinggi tanaman (IT - em), jumIah eabang utama (JCU), jumlah polong eipo (JPC), dan jumIah polong bernas (JPB) per tanaman diantara tanaman awat (kacang tanah cv. Kelinci, SD) dan tanaman R2, zuriat tanaman RO yang diregenerasikan dari ES insensitif ftltrat kultur cendawan hasil seleksi in vitro tanpa perlakuan inokulasi (SC), atau setelah diinokulasi dengan S. rolfsii (SC+JNO).
J
?:;
§.
a
I
~
!!!-t;!
oe..
~. ~:
aO
e.
e.
?
~ 0;<l
e
1(
-~.
s;
~
e.
i
_.
f<>ti
$
-.
~.'"
.!!.~
~~
~
§
J
t
5·
&9.
I
I
l
~
!
I
~I
~
~~
It. ~lffi.
~
1
ct.~.
I
;<l
-~.
5'
l
ct.~
e.
5·I.
j
~
e.
t
5·
~.
~
I
'"
II."
~
a
f
I
?:;
§.
a
~
'"
00H
ct.go .,.
§i'
Pi
ooSl
?"";:: ....
~a.s.
§ B ~§
e-
ill.
~
e
sJ;il~dC'~§§
';;;" _. S
e-n "~
5.
[oo ~I
~ ~
w§
e.
,<OO-:
§ "
g
(Jq Ei[;<le.~
r» _..E 0 til" ... : <:_.
.
~~R'e~~
~ ~ oo§. _.
§
'0g-
it
~~::I-~
5.~Ol
§
, , 0 "$ [
a
~: ~
" §~ill
~
§
?:;~
so sc 1.1t-8C 1.13-8C 1.14-SC usc 3.10-S0 3.12-5e 3.1-5C 304-8C U-$C u.sc 11.7-<: u·sc~
IS
T.,-5C] i4O.S u-sc i 53.3 U-SC 10.12-8C 10.115-5e 1U-8C 12.2-8C 13.13-5C 1U-5C 14.2-8C 111.1-5C 18.2-5C 19.3-5C 20.1-5C 21.1-5C " .... c 24.1-8C""'"'"
211.15-8C 72.0 68.0 ~ " .... c 28ue 27.1-5C sc SOl
~ .~
;;;
-
.
!
..
'
7.3 ~ 52.5 2.1-5C 3.10-SC 3.12-5C 8.3·SC 7.1-SC IU-SC U-SC 10.12-5C 12.2-$C 13.13-5e 13.6-5C 1U-SC 15.1-5C 18.2-8C 19.3-$C~
IS
~ .!. -w o30
.,
(a)
..
c ~..
M .c 20 ~:
~ ~ :;j ~'"
~ c 0 0 Co .c 10 .!! E ::J..,
0 30 en(b)
..
c ~"
.c 20'"
c 0'0
Co 0 .c 10 .!! E ::J..,
0Gambar 29. Jumlab polong bemas dari kacang tanab CY. Kelinci (tanaman awal,
SD), rataan semua tanaman R2 yang diuji (SC), dan rataan tanaman R2 zurlat masing-masing tanaman RI, (a) tanpa inokulasi dengan cendawan dan (b) setelab diinokulasi dengan S. rolfsii di rumab plastik.
zuriat galur Rl yang terinfeksi mempunyai rataan tinggi tanaman lebih rendah, hampir sarna atau lebih tinggi dibandingkan dengan tanarnan awal yang tidak diinokulasi, tetapi zuriat galur Rl tertenlu (K9.2-SC, K-IO.12-SC, K13.13-SC, dan K28.6-SC) terinfeksi S. roifsii dan semuanya mati, sehingga tidak dapat diamati.
100
'"
-
E ~'"
Ca) u'"
'"
'"'"
'"
75
"' "'
ci-
~ ~'"
~ ~ ~ ~ ~ ~ c ~'"
E'"
50
cS
c;,
25
'"
c ;::0
c u u u~
u u u u u u u u u u u u u u u c 0 0:
~ ~ ~ ~ ~•
~ ~ ~ ~3
w ~ ~ ~ 0 0 N ~ N ~"
,
N~
N-"
.;..
" ~ N-"
i
;: ~" "
~,
-•
~"
"
" m .;-
N100
-
E'"
"
Cb)'"
~ U ~"'.
'"
N:;;
-
75
~ ~'"
'" '"
C ~,.
~ ;0 ;0'"
E'"
50 cS
'"
25
'"
c;::
0
C U U U U U U ~ ~ U U U U U U U U U ~ C 0•
• •
• •
~•
~
• •
~ ~•
~ ~•
N ~ N '" ~ ~-
-
0-
~ ~-
•
~ ~;
-
:;
~ ~"
N, ,
~ ~ N"
..
.; .; .;-
-
~ ~"
N"
Gambar 30. Representatiftinggi tanaman dari kacang
tanah
cv. Kelinci (tanaman awal. SD), rataan semua tanaman R2 yang diuji (SC), dan individu tanaman R2 zuriat dari tanaman RO yang tertinggi. Ca) tanpa perlakuan inokulasi cendawan dan (b) setelah diinokulasi dengan S. rolftii di rumab plastik.Gambar 29a menunjukkan babwa tanaman somaklon R2 zuriat dari galur-galur Rl terpilih yang tidak diiookulasi mempunyai rataan jumlab polong bemas yang lebih rendab, hampir sama atau lebih tinggi daripada tioggi tanaman awal. Sebaliknya, setelab diinokulasi (Gambar 29b), rataanjumlab polong bernas zuriat galur-galur Rl terpilih tersebut turun drastis, sehinggalebih kocil atau hampir
<II 40
..
(a)
c ~..
30'"
'"
"
..
'"
N'"
C" "
"
N 0'"
'"
~'0
20..,
a. ~ .c .l!! 10 E ::J...,
0 c 0 0 0 u u u 0 u u u u u u u u 0 0 0 0 0 0 c w ~ ~•
•
~•
~
~• •
~•
;
~• •
~
~•
•
w•
~~
~
;
~3
•
;
~..,
.., ..,•
..,i
~ ;; N ~-
,
g
-
..,•
•
N ~-
~"
,
- - - - -
N N N ,.; ,.; <II 40..
c (b) ~..
30'"
'"
"
N C N .S! 20'"
" "
"
"
.~'"
'"
~ 0"
'"
:0 ~ ~"
~ ~ ~"
'"
~"'.
~ a.'"
'"
.c .l!! 10 E ::J...,
0 c ~ ~ ~ 0 0 ~ 0 0 0~
0 ~~
0 ~ 0 0 0~
0 0 0 c•
,
• • • •
~•
•
~• • • •
• •
:
;
0 ~ 0,
•
•
,
~ 0-
0 ~-
~,
N'"
;; ~ ~ :;•
~-
~~
~ ~;
•
;
•
-
~ ~"
..
"
•
-
-
N N N"
"
-
NGambar 31. Representatif jumlah polong bemas dati kacang tanah cv. Kelinci (tanarnan awal, SD), rataan semua tanarnan R2 yang diuji (SC), dan individu tanaman R2 zuriat dari tanaman RO yang tertinggi. (a) tanpa perlakuan
inokulasi
cendawan dan (b) setelahdiinokulasi
dengan S. rolfsii di rumah plastik.
sarna dengan tanaman awal. Hal ini karena dalam nilai mtaan tersebut tennasuk tanarnan mati akibat infeksi S. rolfs;; yang tidak mengbasilkan polong. Di samping ita, zuria! galur Rl tertenlu (K9.2-SC, K-I0.12-SC, KI3.13-SC, dan K28.6-SC) yang diinokulasi dengan S. rolf.;; semuanya mengalarni kematian, sehingga tidak dapat diarnati dan sarna sekali tidak mengbasilkan polong bemas.
Walaupun secara kole1ctif zuriat galur RI terpilih pada uroumnya mempunyai jumlah polong bemas yang lebih rendah dibandingkan dengan tanaman awal. tetapi secara individu sejumlah nomor somaklon R2 tertentu setelah diinokulasi dengan S. rolfsii mempunyai keragaan agronomis yang sarna atau lebih baik daripada taoaman awal. Pada Gambar 30a-b terlihat bahwa individu-individu tanaman R2 tertentu, mempunyai tinggi tanaman yang lebih besar dibandingkan dengan rataan tinggi tanaman awal maupun semua somaklon yang diuji. baik tanpa pedakuan inokulasi maupun dengan inokulasi cendawan.
Pada Gamhar 31a terlihat bahwa tanpa perlakuan inokulasi S. rolfsii,
sejumlah individu somaklon R2 tertentu menghasilkan jumlah polong bemas lebih tinggi dari rataan jurolah polong bemas tanaman awal maupun rataan dari semua somaklon yang diuji, sedangkan pada perlakuan inokulasi dengan S. rolfsii
(Gambar 3Ib), walaupun secara uroum nilainya lebih rendah daripada taopa perlakuan inokulasi, beberapa individu tanaman masih mampu menghasilkan polong bemas dalam jurolah yang sama atau lebih tinggi daripada taoaman awal yang tidak diinokulasi.
Contoh tanaman somaklon yang tidak diinokulasi dan somaklon yang diinokulasi tetapi masih mampu menghasilkan polong bemas dengan gejala proses penyembuhan luka disajikan pada Gambar 32a-<l, Gambar 32a menunjukkan pangkal batang yang tidak bergejala karena tidak diinokulasi, dan polong bemas yang dihasilkan jurolalmya cukup banyak. Garnbar 32b-<l ada1ah contoh-contoh tanaman somaklon yang terinfeksi S. rolfsii. Tanaman terinfeksi ini pada awal perkembangan infeksinya
terjadi
lesio di pangkal batang. tetapi seiring dengan pertumbuhan dan perkembangan taoaman hingga panen terjadiGambar 32. Contoh tanarnan somakJon yang tidak diinokulasi dan diinokulasi dengan S. rolftii. (a) PangkaJ bataog tidak menunjukkan gejaJa
penyakit. sehingga tidak terdapat lesio. (b, e, dan d) Confoh tanarnan terinfeksi yang pada tahap perkembangannya
menunjukkan proses penyembuhan luka (anak panah), yang
ditandai dengan terbentuknya kalus. Tanaman-tanaman ini mampu
menghasilkan polong bemas dalam jumlah eukup banyak.
proses penyembuhan luka dengan terbentuknya kalus dan lanamall terse but
mampu menghasilkan polong bernas dalarn jurnlah hampir sarna dengan tanaman
yang tidak diinokulasi.
PEMBAHASAN
Segl'cgasj antara Resisten dan Rentan pada Populasi Somaldon
Hasil penelitian ini menunjukkan bahwa baik pada generasi Rl maupun R2. respons lanaman somaklon kacang tanah hasil seleksi in vitro dengan fihrat
kultur cend.wan terhadap infeksi S. rolfsii hersegregasi untuk tingk.t keparahan penyakit antara skor 1-4 (tanaman masih hidup) dengan skor 5 (tanaman mati). Karena tanaman dengan gejala penyakit pada skoT 4 umumnya tidak mampu menghasilkan polong bernas, roaka tanaman terinfeksi yang dianggap resisten penyakit busuk b.tang Sclerotium ad.lab yang bergejal. dengan skor 1-3.
Pada somaklon generasi R 1 yang diinokulasi, frekuensi tanaman dengan gejaia penyakit skor 2 sebanyak 4 tanaman, sedangkan SkOT 3 sebanyak 6 tanaman. Dengan kata lain, 56% somaldon Rl menunjukkan respons resisten terhadap infeksi S. rolfsii. Dari 10 tanaman Rl yang resisten dan zuriat R2-nya ditanam untuk diinokulasi, 7 galur diantaranya menghasilkan zuriat R2 yang resisten dengan skor 2-3, sedangkan 3 galur (KIO.12-SC, KI3.13-SC, dan K26.8-SC) zuriat R2-nya rentan, semuanya mati dengan skor 5 (Tabel 15). Hal ini mengindikasikan adanya konsistensi dalam respons resistensi tanaman somaklon hasil seleksi in vitro terhadap infeksi S. rolfsji, yaitu bahwa somaklon kaeang tanah hasil seleksi in vitro dengan filtrat kultur cendawan menunjukkan gejala resistensi terhadap penyakit busuk batang Sclerotium, baik pada generasi Rl maupunR2.
Pada generasi R2, dari 158 ,omak1on zuri.t dari 34 galur RI terpilih (tennasuk 10 galur Rl yang resisten) yang diinokulasi dengan S. rolftii, 84 tanarnan mati dengan 'kor 5 (53%), 8 tanaman menunjukkan gejal. penyakit dengan skor 4 dan tidak menghasilkan polong, sedangkan 66 tanaman sisany. (42%) menunjukkan gejala resisten. Dengan demikian, dari serangkaian percobaan yang telab dilakukan sebelurnny. (Yusnita ef al. 2004., Yusnita ef al. 2004b) yang dilanjutkan dengan percobaan ini, telab didap.tkan tanaman
somaklon kacang tanah yang resisten terhadap infeksi S. rolfsii. HasH ini mengkonfirmasi hasil penelitian sebelwnnya (Song et al. 1994. Ahmed et af. 1996, Jayasankar dan Litz 1998, Jayasankar et al. 2000) bahwa induksi variasi somaklonal dan seleksi in vitro menggunakan filtrat kultur cendawan patogen dapat digunakan untuk. mendapatkan varian dengan fenotip toteran atau resisten terhadap penyakit target.
Penggoiongan toleran atau resisten suatu tanaman terhadap penyakit pada umumnya didasarkan atas produktivitas, dalam hal ini jumlah polong bemas per tanaman yang dihasilkan, relatif terhadap rataan jumlah polong bemas tanaman awal yang tidak diinoku1asi. Hasil pengarnatan 1ebih lanjut dengan jumlah polong bemas per tanaman menunjukkan bahwa setidaknya sebanyak 20 tanaman menghasilkan jumlah polong bemas per tanaman hampir sarna atau lebih banyak daripada rataan tanaman awal yang tidak diinokulasi, sehingga dapat digolongkan sebagai resisten. Rataan polong bernas per tanaman awal yang tidak diinokulasi betjumlah 14.3, sedangkan pada ke-20 somak1on tersebut jumlah polong bernas berkisar antara 13 hingga 22. Narnun demikian, tampaknya penggolongan karakter resisten dan toleran pada somaklon yang menghasilkan gejala penyakit dengan skor 1-3 setelah diinokulasi
S.
rolfsii dalarn penelitianini
kurang relevan, karena ada kemungkinan bahwa karakter kuantitatif tersebut Gumlah polong bernas) juga mengalami mutasi dan masih bersegregasi pada generasi Rl dan R2. Dengan demikian, nomor galur resisten yang pada generasi tertentu menghasilkan sedikit poiong, mungkin dapat membentuk polong bernas dalam jumlah banyak pada generasi berikutnya.Adanya segregasi pada populasi Rl dan R2 untuk karakter rentan (skor 5) dan toleran atau resisten (skor 1-3) setelah diinokulasi dengan S. rolfsii mengindikasikan bahwa karakter resistensi pada somaldon generasi RO (tetua tanaman Rl) dan generasi Rl (tetua tanaman R2) dikendalikan oleh gen mutan heterozigot. Karakter resistensi terhadap penyakit busuk batang Sclerotium diduga terjadi karena teIjadinya mutasi pada somaklon yang diregenerasikan dari ES yang sebelumnya dipelihara dalam kultur selama lebih dari satu tahun dan dilakukan seleksi in vitro menggunakan filtrat kultur S. rolft;;.
Induksi Variasi Somaklonal dan Seleksi In Vitro Efektif untuk Mendapatkan Tanaman Resisten Penyakit Busuk Batang
Dalam penelitian ini, tampak bahwa hampir semua zuriat galur RO menghasilkan· mutan resisten penyakit busuk batang Sclerotium. Hanya galur RO nomor K9-SC saja yang populasi zuriat R2-nya semuanya mati. Hal ini mengindikasikan bahwa penggunaan kultur ES yang telah berumur satu tahun dan seleksi in vitro dengan filtrat kultur S. rolfsii merupakan paduan yang efektif untuk menghasilkan ES yang insensitif terhadap agens penyeleksi, dan ekspresi fenotipe pada level sel paralel dengan ekspresi pada level tanaman utuh generasi RO, Rl dan R2. Dengan kala lain, agens penyeleksi yang digunakan untuk seleksi in vitro terbukti efektif menghasilkan ES dan somaklon yang resisten terhadap penyakit target. Dalam penelitian terdahulu, fillrat kultur cendawan Fusarium solani juga efektif digunakan sebagai agens penyeleksi dalam seleksi in vitro ES kedelai dan menghasilkan somaklon resisten (Jin et al. 1996). Jayasankar et al. (2000) juga menggunakan fillrat kultur cendawan Eisinoe ampelina untuk agens penyeleksi dalam seleksi in vitro Vilis vinifera
'Chardonnay', dan tanaman regeneran menWljukkan gejala resistensi terhadap penyakit anthraknosa.
Mekanisme ketahanan.somaklon terhadap infeksi S. rolfsii yang terjadi dalam percobaan ini tampaknya bukan merupakan reaksi hiper-sensitif yang merupakan reaksi cepat, tetapi cenderung pada terjadinya proses lignifikasi dan pembentukan kalus pada bagian batang yang terinfeksi, seiring dengan mengeringnya miselia cendawan di sekitar lesio. Hal ini mengingat bahwa pada awalnya, nekrosis dan lesio pada pangkal batang sudah teIjadi dan sempat meluas hingga melingkari seluruh batang (skor 3), namun pada perkembangan selanjutnya miselia cendawan lama kelamaan mengering dan terjadi proses penyembuhan luka (Gambar 32b-d).
Faktor Patogenisitas S. rolfsii dan Kemungkinan Mekanisme Resistensi Dalam proses patogenesisnya, S. ro/fsii menyintesis dan mensekresikan fitotoksin asam oksalat (OA) dalam jumlab besar (konsentrasi milimolar) dan enzim pendegradasi dinding sel seperti endo-poligakturonase (endo-PG) dan selulase ke dalam jaringan yang terinfeksi. OA menyebabkan pH asam pada jaringan tanaman, sehingga optimum bagi aktivitas enzim eoda-PG. Di samping itu, OA mampu mengkelat Ca ++ dari dinding sel, sehingga sel dan jaringan
tanaman inang menjadi sangat rentan terhadap enzim pendegradasi dinding sel yang dike1uarkan oleh cendawan (Punja 1985, Punja et al., 1985, Rowe 1993). OA juga dapat menghambat alrtivitas O-diphenol oxidase, sehingga menghambat oxidative burst (Cessna et al. 2000). Penghambatan oxidative burst dapat menyebabkan reaksi hiper-sensitif tidak tetjadi. Pentingnya OA dalam proses
patogenesis juga telah ditunjukkan oleh adanya mutan cendawan S. sclerotiorum yang tidak dapat menyintesis OA, dan temyata mutan cendawan tersebut tidak virulen, sementara cendawan wild-type dan mutan revertant-oya dapat menyebabkan penyakit busuk batang pada Phaseolus vulgaris L (Godoy et al. 1990, Jamaux et al. 1995). Dalam jwn1ah rendah, OA ditemukan dalam berbagai tanaman, yang tersimpan di vakuo1a (<;:alis~an 2000)
Dalam penelitian ini, somaklon kacang tanah yang resisten terhadap S. rolfsii yang didapat merupakan zuriat dari galur-galur RO yang diregenerasikan dari ES yang insensitif terhadap filtrat kultur cendawan yang diasumsikan mengandung ~A. Filtrat tersebut tidak mengandung enzim pendegradasi dinding sel, karena biakan cendawan telah disterilkan dengan autoklaf. Dengan demikian, diduga mekanisme pertahanan yang terjadi di dalam somaklon resisten S rolftii tentunya disebabkan oleh detoksifikasi ~A.
Strategi yang umum digunakan untuk menjadikan suatu tanaman resisten terhadap cendawan patogen yang mensekresikan OA adalah dengan mendegradasi OA tersebut pada saat awal terjadinya infeksi (Lu 2003). Terdapat tiga kelas enzim yang dikenal dapat berfungsi dalam katabolisme OA, yaitu oxalate oxidase (OXO) (Lane et al. 1991), oxalate decarborylase (Mehta dan Datta 1991), dan oxalyl-CoA decarborylase (Lung et al. 1994). Gen penyandi oxalyl-CoA decarborylase dari bakteri dapat digunakan untuk mendegradasi OA dan merekayasa tanaman transgenik yang resisten Sclerotinia sclerotiorum (Dickman dan Mitra 1992), tetapi enzim oxalate decarboksylase yang berasal dari cendawan maupun bakteri bekerja dengan mengubah OA menjadi CO, dan asam format yang dapat meracuni tanaman (Lu 2003). 01eb karena itu, para
peneliti telah memfokuskan perhatian pada enzim
axo,
yang mengubah OA dan O2 menjadi CO2 dan H202_ Enzim OXO pertama kali ditemukan di barley dan gandum (Lane et al 1993, Kotsira dan Klonis 1997). Enzim OXO yang berasal dari gandum merupakan enzim yang bekerja apoplastik, multimerik dan terglikosilasi dengan ketahanan yang tinggi terhadap panas, degradasi oleh protease dan H,O, (Lane 2000). Sejauh ini. tanaman yang sudab diketahui mempunyai aktivitas enzimaxo
adalah gandum, barley, jagung, oat, padi, rye dan cemara (Dunwell et al. 2000, Lane 2002).Dengan mengasumsikan bahwa tanaman kacang tanah juga mempunyai gen penyandi enzim OXO sebagaimana tanaman lain di atas, maka kemungkinan tanaman somaklon resisten kacang tanah yang didapatkan dalam penelitian ini telah mengalami over ekspresi enzim OXO setelah- mengalarni seleksi in vitro dengan filtrat kultur S. rolfsii yang mengandung OA pada konsentrasi tinggi. Over-ekspresi suatu produk gen tertentu mungkin terjadi pada mutan tanaman tertentu. Hal ini dapat disebabkan oleh tidak aktifnya trans-acting factor yang semula menghambat aktivitas enzim OXO karena telah terjadi mutasi titik atau insertional inactivation pada sekuen gen penyandinya. Kemungkinan yang lain, telah terjadi duplikasi sekuen gen penyandi OXO karena terjadi chromosom rearrangengement. Perubahan genetik spesifik yang menyebabkan variasi somaklonal lelab ditelaah secara mendalam (Larkin dan Scowcroft 1981; Scowcroft et al.1985, HammerscWag 1992). Perubahan genetik yang mendasari terjadinya variasi somaklonal meliputi mutasi titik (single nucleotide changes), perubahan jumlah kopi gen tertentu, aktivasi transposable element, perubahan jumlah kromosom, chromosomal rearrangenements, dan metilasi DNA.
KESIMPULAN
Dari penelitian ini telah didapatkan beberapa .galur somaklon kacang tanah zuriat dari somaklon RO yang resisten terhadap infeksi S. rolftii. Sifat resistensi ini terdapat diantara populasi somaklon RI dan R2. yang bersegregasi antara resisten dan rentan terhadap infeksi S. rolfsii. Somaklon kacang tanah yang resisten terhadap infeksi S. ro!ftii pada awalnya menunjukkan gejala penyakit busuk batang dengan skoT antara 1-3, tetapi dalam perkembangannya tanaman menunjukkan proses penyembuhan luka pada lesio, dan tanaman mampu menghasilkan polong bernas yang sarna alau lebih banyak dibandingkan tanaman kacang tanah awal yang tidak diinokulasi.