www.elsevier.nlrlocateraqua-online
ž
Protein digestion in juvenile turbot Scophthalmus
/
maximus and effects of dietary administration of
Vibrio proteolyticus
R. De Schrijver
a,), F. Ollevier
ba
Laboratory of Nutrition, Faculty of Agriculture and Applied Biological Sciences, Catholic UniÕersity of
LeuÕen, Kardinaal Mercierlaan 92, B-3001, LeuÕen, Belgium
b
Laboratory of Ecology and Aquaculture, Catholic UniÕersity of LeuÕen, LeuÕen, Belgium
Accepted 11 November 1999
Abstract
The trial was made to study the protein digestion in juvenile turbot during passage of digesta from stomach to rectum. Moreover, diet supplementation with the potential probiotic bacteria,
Vibrio proteolyticus, was evaluated with regard to protein digestion. For a 3-week period, fish
Ž25–30 g were fed by oral intubation and received either a liquid mixture consisting of 40%.
nonpurified control diet and 60% water or this mixture supplemented with 1010 viable V.
proteolyticus mly1. Daily dry matter intake was 1.5% body weight. As digesta progressed from
the stomach to the foregut, hindgut and rectum, the increase in apparent nitrogen digestibility was accompanied by higher ammonia contents, suggesting substantial involvement of the microbiota in protein degradation in the distal segments of the gastrointestinal tract. Water-soluble nitrogen contents were significantly higher in the foregut, presumably corresponding with considerable protein digestion by secreted endogenous enzymes in this digestive segment. Over 65% of the soluble protein in all four parts of the tract had a MW-10,000. The amount of soluble protein and peptides with MW-1000 decreased significantly during transit. This was also found for the
Ž .
proportion of the 10,000–20,000 MW proteins, whereas the highest MW category )200,000 increased. Ingestion of V. proteolyticus tended to stimulate apparent nitrogen digestibility
ŽP-0.1 . This effect corresponded with increased protein degradation in the proximal intestine as.
)Corresponding author. Tel.:q32-16-321575; fax:q32-16-321971.
Ž .
E-mail address: [email protected] R. De Schrijver .
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
was shown by the significantly elevated fraction of soluble proteins with MW-1000. q2000 Elsevier Science B.V. All rights reserved.
Keywords: Turbot; Scophthalmus maximus; Protein digestion; Vibrio proteolyticus; Probiotics
1. Introduction
In turbot hatcheries, considerable mortality and consequently economic losses may occur due to vibriosis. This disease is mainly caused by Vibrio anguillarum. Nowadays, the prophylactic and therapeutic control of vibriosis is based on oral administration of antibiotics. However, such treatment may cause the development of resistant bacteria ŽAoki et al., 1985 and yield residues in fish. An alternative treatment could be the. administration of probiotic bacteria that exert inhibitory effects against V. anguillarum and support the natural host microbial defense mechanisms. Preferentially, such bacteria should be of host origin as they must be able to colonize the gastrointestinal tract ŽConway, 1989; Hansen and Olafsen, 1989 . V. proteolyticus appears to fulfill these. conditions as in vitro experiments have shown that this bacterial strain exerted inhibitory
Ž .
effects against V. anguillarum Grisez, 1997 . Moreover, preliminary experiments indicated that these bacteria are able to colonize the gastrointestinal tract of turbot juveniles. The potential role of V. proteolyticus as a prophylactic agent against vibriosis is currently under further investigation. Another practical aspect of the use of probiotics in fish diets that must be clarified is their possible positive effect on feed efficiency. In this context, the present study dealt with the potential of dietary V. proteolyticus supplementation on protein digestion in juvenile turbot. However, the actual knowledge of the extent of protein digestion and absorption in this species, at this age, is greatly inadequate. Thus, an additional aim of this study was to obtain information in this regard. Following the feeding of a diet with or without V. proteolyticus supplementation, the progress of protein digestion was examined as digesta passed through the alimentary tract. Therefore, apparent nitrogen digestibility was measured in the different segments
Ž .
of the gastrointestinal tract. Molecular weight MW profiles of soluble proteins were determined as well. This latter technique has been successfully used in experiments with
Ž .
young poultry and piglets Sklan and Hurwitz, 1980; Asche et al., 1989 . Additional parameters related to protein degradation were also measured such as water-soluble nitrogen, ammonia and pH.
2. Materials and methods
2.1. Fish husbandry
Ž . Ž .
Turbot Scophthalmus maximus juveniles 25–30 g were obtained from a
commer-Ž .
water during the trial was kept between 188C and 208C, salinity was 3.6%, oxygen saturation was between 80% and 90% and nitrite content varied between 0.05 and 0.1 mg ly1. The experimental protocols and procedures were approved by the Animal Use and Care Committee at the Catholic University of Leuven, Belgium.
2.2. Dietary treatments
The basal diet was formulated to contain 570 g kgy1 crude protein and consisted
Ž .
mainly of nonpurified ingredients Table 1 . In the 3-week experimental period, fish were fed individually twice daily by oral intubations of a liquid diet consisting of 40% ground basal diet and 60% water. Daily dry matter intake was set at 1.5% body weight.
Ž .
Two dietary treatments were tested: the basal diet C vs. the basal diet supplemented
10 Ž . y1
with 10 V. proteolyticus VP ml diet–water mixture. Preliminary trials had revealed that under these circumstances, colonization of the digestive tract with this bacterial strain occurred. Diet–water mixtures were freshly prepared at each time of feeding. Each diet was fed to 18 fish. All fish in the same tank were subjected to the
y1 Ž .
same treatment. The diets contained 5 g kg chromic oxide technical grade, 98% to mark the particulate material and to provide a measurement of protein digestibility.
2.3. Sampling
On the sampling day, all fish were fed in the morning and killed 8 h later. In a preliminary experiment, it was found that this time period resulted in the presence of
Ž
digesta in the overall digestive tract. The stomach, foregut from the pyloric caeca to the
Table 1
Ingredients and chemical composition of the basal diet
a y1 b y1
Ingredient g kg Chemical composition g kg
Herring meal 580 Dry matter 920
Dried whey 50 Crude protein 570
Blood meal 100 Crude fat 158
Condensed milk solubles 25 Crude fiber 5
Wheat middlings 63 Crude ash 118
Fish oil 120 N-free extract 69
Molasses 35
Dietary ingredients were obtained from a local supplier.
b Ž .
Proximate analyses were performed using AOAC 1990 procedures. c
Provided per kilogram diet: 75 mg Zn as ZnSO ; 20 mg Mn as MnSO ; 1.6 mg Cu as CuSO ; 10 mg I as4 4 4 KIO .3
d
Provided per kilogram diet: 2.25 mg retinyl acetate; 0.015 mg cholecalciferol; 500 mgDL-a-tocopheryl
. Ž .
middle of the intestine , hindgut distal half of the intestine and rectum were dissected Ž .
according to Koven et al. 1994 . The contents from the digestive segments were gently extruded and collected separately. As the amount of digesta from individual fish was insufficient to carry out the analyses, the samples from three fish were pooled.
2.4. Digesta analyses
Following killing, the pH and ammonia content of the digesta in the different intestinal segments were measured immediately. The pH was determined with a
Ž .
Hamilton Slimtrode Filter Service, Belgium . Ammonia was measured with an
ion-Ž .
selective electrode Orion, 95-1201, Ankersmit, Belgium following extraction of the
y1 Ž .
samples with 0.1 mol l HCl Byrne and Power, 1974 .
The digesta were freeze-dried and subsequently ground using a pestle and mortar. Nitrogen was determined by the Kjeldahl method. Chromic oxide was measured spectrophotometrically following dry destruction of the samples at 5508C and oxidation
3q 6q Ž .
of Cr to Cr with sodium molybdate Fenton and Fenton, 1979 . In order to determine the amount of water-soluble nitrogen in the digesta, a 2% suspension of the freeze-dried sample was prepared. Following homogenization by stirring, the mixture
Ž .
was centrifuged 25,000=g for 30 min at 58C , the supernatant decanted, the residue
Ž .
resuspended in water in the same volume as was used in the first extraction step , again centrifuged and the supernatant added to the previous one; the combined supernatant was filtered and analyzed for nitrogen.
Soluble proteins in the digesta were separated by size exclusion, high-performance Ž . liquid chromatography according to a modified procedure of Asche et al. 1989 .
Ž
Proteins were fractionated on a TSK-SW2000 column 7.5 mm i.d.=60 cm; Toyo Soda,
. y1
Japan . Elution of the fractions was performed with a 0.1 mol l sodium phosphate
Ž . Ž y1.
buffer pH 7.0 containing sodium dodecyl sulphate 1 g l at a flow rate of 0.3 ml miny1. Conditions were isochratic. The pressure varied between 3.5 and 5.5 kg cmy2
Žpump model 600E, Millipore, Belgium . In order to protect the analytical column, an. in-line filter and a 7.5 mm i.d.=7.5 cm pre-column were used; moreover, samples were filtered through a 0.45-mm membrane filter before injection. Eluted proteins were
Ž .
detected spectrophotometrically at 210 nm model 160, Beckman Instruments, Belgium . The reference solution consisted of thyroglobulin, serum albumin, ovalbumin, b -lacto-globulin A, chymotrypsinogen, ribonuclease, insulin chain B, glucagon and bacitracin ŽSigma-Aldrich, Belgium ..
Apparent nitrogen digestibility was calculated from the nitrogen and chromic oxide Ž
contents in the diets and the digesta using the general formula for digestibility Vanhoof .
and De Schrijver, 1996 .
2.5. Statistical analyses
Ž
Data were analyzed by two-way ANOVA General Linear Model program; SAS .
P-0.1 were considered as trends. Values are reported as means"SD. In a few instances, all analyses could not be completed due to insufficient sample size.
3. Results and discussion
Ž .
The extent of apparent protein digestion Table 2 increased in both dietary treatment groups as digesta moved from the stomach down to the rectum. As expected, there was no substantial apparent nitrogen digestibility measured in the stomach. Although gastric proteolytic enzymes may be produced and secreted in juvenile turbot, the stomach is generally not considered as a site for absorption of amino acids and peptides. In light of the large standard errors that were associated with the apparent nitrogen digestibility data in the stomach, conclusions regarding dietary effects should not be drawn from the slightly negative mean value in the control group and the slightly positive mean value in the V. proteolyticus-supplemented group. As compared with the apparent nitrogen digestibility in the hindgut, the digestibility in the proximal gut tended to be lower ŽP-0.1 . This lower digestibility may result from influx of endogenous nitrogen. originating from the intestinal mucosa, the bile and pancreatic secretions. Substantial protein degradation also occurred in the rectum as differences in apparent nitrogen
Ž .
digestibility were observed between hindgut and rectum P-0.1 . To what extent the inherently present gastrointestinal microflora was involved in protein breakdown could not be derived from the present study since no tests on microbial activity were carried out. On the other hand, more concrete information was obtained concerning the role of the bacteria which were added to the diet. The ingestion of V. proteolyticus resulted in increased apparent nitrogen digestibility, mainly when measured at the site of the
Ž .
hindgut and rectum P-0.1 . These effects suggest that the tested bacterial strain was able to stimulate protein degradation in juvenile turbot as soon as pH requirements were fulfilled. In the stomach, pH values were generally lower than 5, whereas pH values in
Ž .
the gut and rectum were higher than 6 and 7, respectively Table 2 . As compared with control group, the V. proteolyticus group showed elevated ammonia content in the
Ž . Ž .
digesta of the hindgut P-0.05 and rectum P-0.1 , indicating that the supple-mented proteolytic bacteria were active in these parts of the digestive tract. As a consequence, the increased ammonia concentrations following ingestion of V. prote-olyticus corresponded with the earlier mentioned higher apparent nitrogen digestibilities in the distal digestive segments. As in other species, it appears that also young turbot shows the highest microbial proteolytic activity in the distal parts of the digestive tract since in both treatment groups, ammonia contents were significantly increased in hindgut and rectum as compared with the upper gut.
With respect to soluble nitrogen, no significant effects due to feeding regimen were Ž .
found Table 2 . In contrast, there were significant differences in relation with the digestive segment. In the stomach, the content of water-soluble nitrogen was, on average, 18% of total nitrogen. This was clearly the lowest value that was measured in the gastrointestinal tract, showing little protein digestion in the stomach as was also concluded from the apparent nitrogen digestibility data. The highest water-soluble
Ž .
()
R.
De
Schrij
Õ
er,
F.
Olle
Õ
ier
r
Aquaculture
186
2000
107
–
116
Table 2
Ž . Ž . Ž y1.
Apparent nitrogen digestibility % , water-soluble nitrogen % , ammonia content mmol g and pH in gastrointestinal digesta of juvenile turbot fed a diet with or without supplemented V. proteolyticus)
a
Ž .
Diet C Diet VP Pooled SD ANOVA P value
Stomach Foregut Hindgut Rectum Stomach Foregut Hindgut Rectum Diet Digestive segment
c b,c a,b b,c a,b a
d d
Apparent N digestibility y3.1 37 53 63 2.0 43 63 77 9 0.09 0.001
c a b b c a b b
Water-soluble N 17 72 43 40 20 68 47 33 8 0.25 0.001
z c b a,b z b,c a a
NH3 nd 3.5 5.7 6.5 nd 4.0 7.8 7.7 1.2 0.04 0.03
c b b a c b b a
PH 4.7 6.8 6.6 7.6 4.3 6.3 6.7 7.8 0.5 0.35 0.01
) Ž .
Values are means, ns5 or 6. Means within a row without common superscript letter are significantly different P-0.05 as determined by Tukey’s contrasts. a
P)0.1 for diet=digestive segment interaction for any measured parameter z
decline until the average of 36% in the rectum. Most probably, the elevated amounts of water-soluble nitrogen in the proximal intestine were related to the release of endoge-nous enzymes by the host and to considerable digestion of dietary and metabolic protein. Apparently, the production of water-soluble nitrogen compounds in the proximal part of the gut occurred too rapidly to be readily absorbed. Conversely, the decline in water-soluble nitrogen in the distal part of the gut was presumably related to less proteolytic activity and a comparatively higher absorption rate. Moreover, the utilization of water-soluble nitrogen for microbial growth may not be excluded in the distal segments of the alimentary tract.
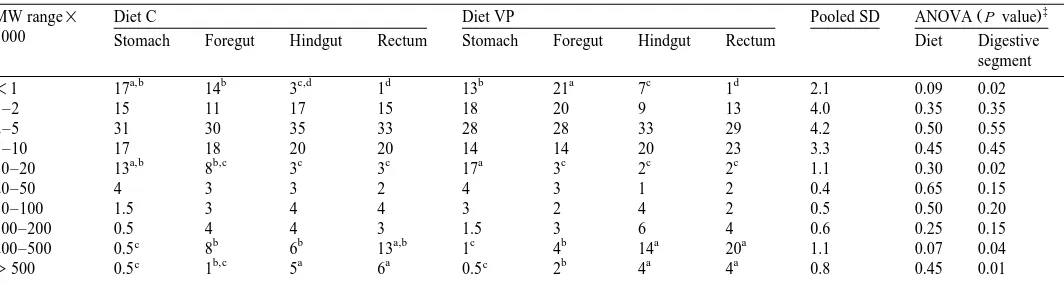
Protein digestion was also examined by measuring the MW profiles of soluble proteins and peptides as digesta passed through the digestive tract. Table 3 gives the percentage partition of total soluble protein in MW categories for each segment of the gastrointestinal tract. Strikingly, the sum of the peptides and proteins, with MW lower than 10,000, represented between 69% and 85% of the total soluble protein in all four segments. Generally, this percentage was not significantly affected by dietary treatment or digestive segment. Characteristically, within the fractions with MWF10,000, the 2000–5000 MW range was predominant in each digestive region and represented between 25% and 35% of total soluble protein. The proportion of the smallest proteins Ž-1000 MW decreased significantly as digesta progressed down the tract, reflecting. absorption by the host and possibly also utilization by the microflora in the distal parts of the tract. Supplementation of the diet with V. proteolyticus resulted in a significantly increased proportion of the lowest MW category in the foregut and in a tendency toward
Ž .
elevated proportions in the hindgut P-0.1 . Thus, ingestion of V. proteolyticus tended to move protein degradation more proximal in the gut. Obviously, these observations were paralleled by enhanced apparent nitrogen digestibility values in the V. proteolyticus group.
The proportion of the 10,000–20,000 MW protein category decreased gradually during the transit of digesta in the tract, but there was no effect from the dietary regimen. The proportion of the 20,000–50,000 MW proteins was generally less than 5% in all parts of the tract, in spite of the fact that the digestive enzymes belong to this MW category. It is not known whether this observation was due to poor production of endogenous enzymes in juvenile turbot as this may be the case in other young animals
Ž .
or to fast hydrolysis after being secreted Bergner et al., 1980 . An additional explana-tion for this apparent discrepancy is the high diluexplana-tion rate of endogenous nitrogen in the
Ž y1.
digestive tract because of the very high protein content in the diet 760 g kg . Asche Ž .
et al. 1989 also reported no substantial proportions of the 20,000–50,000 MW range in the intestine of piglets notwithstanding the feeding of a diet containing only 220 g kgy1 protein. As an explanation, the authors suggested that the endogenous enzymes were rapidly hydrolyzed.
()
R.
De
Schrij
Õ
er,
F.
Olle
Õ
ier
r
Aquaculture
186
2000
107
–
116
Table 3
Ž . Ž .
Molecular weight MW profiles % total protein fraction of the soluble proteins and peptides in gastrointestinal digesta of juvenile turbot fed a diet with or without supplemented Vibrio proteolyticus†
‡
Ž .
MW range= Diet C Diet VP Pooled SD ANOVA P value
1000 Stomach Foregut Hindgut Rectum Stomach Foregut Hindgut Rectum Diet Digestive
segment
a,b b c,d d b a c d
-1 17 14 3 1 13 21 7 1 2.1 0.09 0.02
1–2 15 11 17 15 18 20 9 13 4.0 0.35 0.35
2–5 31 30 35 33 28 28 33 29 4.2 0.50 0.55
5–10 17 18 20 20 14 14 20 23 3.3 0.45 0.45
a,b b,c c c a c c c
10–20 13 8 3 3 17 3 2 2 1.1 0.30 0.02
20–50 4 3 3 2 4 3 1 2 0.4 0.65 0.15
50–100 1.5 3 4 4 3 2 4 2 0.5 0.50 0.20
100–200 0.5 4 4 3 1.5 3 6 4 0.6 0.25 0.15
b b a,b c b a a
c
200–500 0.5 8 6 13 1 4 14 20 1.1 0.07 0.04
b,c a a b a a
c c
)500 0.5 1 5 6 0.5 2 4 4 0.8 0.45 0.01
† Ž .
Values are means, ns5 or 6. Means within a row without common superscript letter are significantly different P-0.05 as determined by Tukey’s contrasts. ‡
microbial utilization as well as to synthesis of microbial protein in the distal segments. This followed, in particular, from the proportions of the proteins with MW)200,000: this protein fraction increased in the control group from 9% in the foregut to 19% in the
Ž .
rectum P-0.1 and in the V. proteolyticus group from 6% in the foregut to 24% in the Ž .
rectum P-0.05 . With respect to the dietary V. proteolyticus treatment, it was found that the 200,000–500,000 MW fraction tended to increase in the hindgut and the rectum, referring to the presence of more microbial protein in the V. proteolyticus group as compared with the control group.
These are the first results that document protein digestion in different parts of the alimentary tract in juvenile turbot. In summary, the observed increase in apparent nitrogen digestibility during the transit down to the rectum was paralleled by a steady
Ž .
decrease in small proteins -1000 MW in the soluble fraction of digesta, whereas the
Ž .
proportion of larger proteins )200,000 increased. Furthermore, the results demon-strated that in the prevailing experimental conditions, dietary fortification with V. proteolyticus moved protein degradation more to the proximal part of the gut and tended to enhance apparent nitrogen digestibility. In animal nutrition, such effects would generally be considered as beneficial. However, as in this study, the observed positive effect on apparent protein digestion was probably highly related to increased proteolytic activity exerted by bacteria and not by fish enzymes; it remains to be elucidated whether the formed protein degradation products contribute to nitrogen retention in the host. Controlled long-term dose–growth response investigations are suggested to further clarify the nutritional significance of the present findings.
Acknowledgements
The authors thank D. Vermeulen and R. Van Houdt for the technical assistance. The present study was supported by grant G.0063.96 from the Fund for Scientific
Research-Ž . Flanders Belgium .
References
AOAC, 1990. Official Methods of Analysis. 15th edn Association of Official Analytical Chemists, Arlington, VA.
Aoki, T., Kanazawa, T., Kitao, T., 1985. Epidemiological surveillance of drug-resistant Vibrio anguillarum strains. Fish Pathol. 20, 199–208.
Asche, G.L., Lewis, A.J., Peo, E.R., 1989. Protein digestion in weanling pigs: effect of feeding regimen and endogenous protein secretion. J. Nutr. 119, 1083–1092.
Bergner, H., Simon, O., Bergner, U., 1980. Endogenous proteins in the process of digestion and absorption. In:
Ž .
Oslage, H.W., Rohr, K. Eds. , Proceedings of the European Association for Animal Production Sympo-sium on Protein Metabolism and Nutrition, Braunschweig, Germany. pp. 198–204.
Byrne, E., Power, T., 1974. Determination of ammonia nitrogen in animal slurries by an ammonia electrode. Commun. Soil Sci. Plant Anal. 5, 51–65.
Ž .
Fenton, T.W., Fenton, M., 1979. An improved procedure for the determination of chromic oxide in feed and feces. Can. J. Anim. Sci. 59, 631–634.
Grisez, L., 1997. The marine fish pathogenic bacterium Vibrio anguillarum: identification, serology, histopathology and selected aspects of virulence. PhD, Thesis, Catholic University of Leuven, B-3000,
Ž .
Leuven, Belgium, Ollevier, F. promoter .
Ž . Ž
Hansen, G.H., Olafsen, J.A., 1989. Bacterial colonization of cod Gadhus morhua and halibut Hippoglossus
.
hippoglossus eggs in marine aquaculture. Appl. Environ. Microbiol. 55, 1435–1446.
Ž .
Koven, W.M., Henderson, R.J., Sargent, J.R., 1994. Lipid digestion in turbot Scophthalmus maximus : I. Lipid class and fatty acid composition of digesta from different segments of the digestive tract. Fish Physiol. Biochem. 13, 69–79.
SAS Institute, 1988. SAS User’s Guide: Statistics. 6.03 edn SAS Institute, Cary, NC.
Sklan, D., Hurwitz, S., 1980. Protein digestion and absorption in young chicks and turkeys. J. Nutr. 110, 139–144.