EKSUDASI ASAM-ASAM ORGANIK PADA BEBERAPA GENOTIPE
TANAMAN PADI TERCEKAM KAHAT HARA FOSFOR
(Organic Acids Exudation in Rice Plant Genotypes under Phosphorus Defficiency) Abd. Aziz Syarif
Balai Pengkajian Teknologi Pertanian Sumatera Barat
ABSTRACT
An experiment to study the effect of plant P status on organic acids exudation among rice plant genotypes differing in field tolerance to low P was carried on in glass house using a Factorial in Compeletely Randomized Block Design with two replicates. The first factor was plant P status (suffi-cient and defi(suffi-cient) and the second factor was rice genotype (three highly tolerant and one highly sensitive). The plant P status was achieved by P starvation and without starvation treatments. Ob-servation was made on organic acids exudation of rice plant using aquadest as collecting medium. The results showed that there were exudations of six organic acids both from starved and unstarved plant, with starved plants exuded higher amount of acids than that from unstarved plants, on plant and on shoot dryweight basis. The contrast result was shown by the amount of exudation on root dryweight basis, where higher exudation was found on unstarved plants. On the shoot dryweight basis, the significantly higher exudation for oxalate was only showed by two of three highly tolerant genotypes (Gadih Ani-2 and O. glumaepatula), and for citrate and format by one highly tolerant genotype (O. glumaepatula). For other acid there was no difference of increase.
Key words : organic acids, exudation, phosphorus defficiency
PENDAHULUAN
umbuhan menunjukkan berbagai
respon terhadap keterbatasan hara fosfor yang dapat dianggap sebagai mekanisme untuk meningkatkan serapan P. Secara garis besar terdapat dua mekanisme pada tumbuhan yang dapat meningkatkan serapan P yakni yang bersifat morfologi akar yang meliputi pertumbuhan, distribusi, dan pembentukan rambut akar, serta fisiologi akar yang meliputi kinetika serapan dan mobilisasi P (Rao et al. 1999). Peningkatan eksudasi asam organik adalah salah satu respon fisiologis tumbuhan terhadap keterbatasan hara P yang dianggap sebagai mekanisme untuk mening-katkan serapan P.
Peningkatan eksudasi asam organik oleh tanaman sebagai respon terhadap keterbatasan P telah banyak dilaporkan. Hoffland et al.
(1989) umpamanya, menemukan peningkatan eksudasi asam malat dan sitrat pada akar
Brassica napus yang mengalami defisiensi P. Peningkatan eksudasi asam sitrat yang hampir dua kali lipat pada tanaman Medicago sativa
defisien P juga dilaporkan oleh Lipton et al.
(1987). Ae et al. (1993) menemukan eksudasi asam piksidat pada kacang gude yang ditanam pada tanah rendah P. Eksudasi asam organik dapat meningkatkan ketersedian P bagi tanam-an karena asam orgtanam-anik dapat melepasktanam-an P dari jerapan baik melalui pertukaran anion maupun pembentukan kompleks dengan Al atau Fe (Jones dan Darrah 1994).
Penelitian tentang peningkatan eksudasi asam organik pada tanaman padi sebagai respon terhadap defisiensi P dirasakan masih terbatas. Percobaan ini bertujuan untuk melihat adanya mekanisme peningkatan eksudasi asam organik pada tanaman padi defisien P dan hubungannya dengan perbedaan ketenggangan genotipe terhadap P rendah di tanah sawah.
BAHAN DAN METODE
Percobaan dilakukan di Rumah Kaca Puslitbang Tanah dan Agroklimat menggu-nakan rancangan percobaan berfaktor dalam acak kelompok dua ulangan. Faktor I adalah status P tanaman (defisien dan cukup). Faktor II adalah empat genotipe yang terdiri
T
atas tiga genotipe sangat tenggang dan satu genotipe sangat rentan.
Benih tanaman padi ditumbuhkan pada larutan hara Yoshida (1976) dengan 10 ppm P selama tiga minggu. Untuk mendapatkan tanaman dengan status P berbeda, pada minggu keempat dilakukan perlakuan pela-paran P dengan cara dua tanaman tiap geno-tipe dipindahkan ke larutan hara Yoshida dengan 10 ppm P dan dua tanaman lainnya tanpa P, dan dipelihara selama empat hari.
Pengumpulan asam organik eksudat akar dilakukan pada hari keempat setelah perlakuan pelaparan P, mengacu pada prosedur yang digunakan Otani et al. (1996) dan Subbarao et al. (1997). Pertama, akar tanaman direndam pada akuades yang diberi chloramphenicol (30 mg/l) selama dua jam untuk mencegah kontaminasi bakteri, dibilas dengan akuades, kemudian dipindahkan ke medium pengumpulan yakni wadah berisi 0,4 L akuades dan dibiarkan selama 24 jam (jam 10 pagi sampai 10 pagi hari berikutnya). Untuk mencegah eksposur pada cahaya maka wadah ditutup dengan kantong plastik hitam. Sampel eksudat (30 ml) dimasukkan ke dalam botol plastik kedap udara, ditambahkan tiga tetes kloroform untuk mencegah dekomposisi oleh bakteri. Metode pengumpulan asam organik di luar larutan hara ini juga digunakan oleh Ohwaki dan Hirata (1990) dan Pellet et al. (1995). Metode ini lebih cocok untuk analisis asam organik yang menggunakan kolom ion-eksklusi. Analisis asam organik dalam larutan hara membutuhkan sistem kroma-tografi ion yang lebih rentan (Pellet et al. 1995). Metode ini juga dapat menghindar-kan pengaruh perbedaan serapan kation/ anion di antara genotipe yang mempenga-ruhi sintesis asam organik pada sitoplasma (Marschner, 1995), sehingga perbedaan eksudasi asam organik yang terjadi hanya akibat perbedaan status P tanaman.
Penentuan jenis dan konsentrasi asam organik eksudat dilakukan di Laboratorium Balai Besar Pascapanen dengan HPLC (High Performance Liquid Chromatography) yang menggunakan kolom analisis eksklusi ion (Waters Associates Model 440) pada fase
mobil 0,05N H2SO4 dengan kecepatan alir 1
ml/menit, tekanan 60 psi, suhu 22 oC, dan
absorban pada 254 nm.
HASIL DAN PEMBAHASAN
Jenis Asam Organik
Terdeteksi eksudasi enam jenis asam organik (oksalat, sitrat, tartarat, format, malat, dan suksinat) dengan jumlah yang berarti pada ke empat genotipe yang digunakan, baik pada perlakuan pelaparan (kahat P) ataupun tanpa pelaparan (cukup P). Hasil ini berbeda dengan hasil penelitian yang dilaporkan Otani et al. (1996) yang hanya mendeteksi satu asam organik (tartarat) dengan jumlah berarti, asam organik lainnya (oksalat, sitrat dan malat) hanya ditemukan dalam jumlah sangat sedikit. Perbedaan ini diduga adalah karena perbedaan perlakuan pelaparan sebelum pengumpulan asam organik. Pada penelitian mereka pela-paran dilakukan dengan menginkubasi tanam-an pada medium pertumbuhtanam-an dengtanam-an P rendah (64 μM), sedangkan pada penelitian ini pelaparan dilakukan dengan medium tanpa P. Hasil penelitian ini juga berbeda dengan hasil penelitian Swasti (2004) yang hanya mendeteksi eksudasi asam oksalat akibat perlakuan cekam-an P rendah dcekam-an Al tinggi. Terbatasnya jenis asam organik yang terdeteksi pada penelitian Otani (1996) dan Swasti (2004) diduga adalah karena penggunaan HPLC dengan eksklusi ion yang kurang sensitif untuk menganalisis asam organik dalam larutan hara (Pellet et al. 1995). Karena itu Pellet et al. (1995) menggunakan pengumpulan eksudat asam organik tanaman dengan media pengumpul akuades.
Jumlah Eksudasi Asam Organik
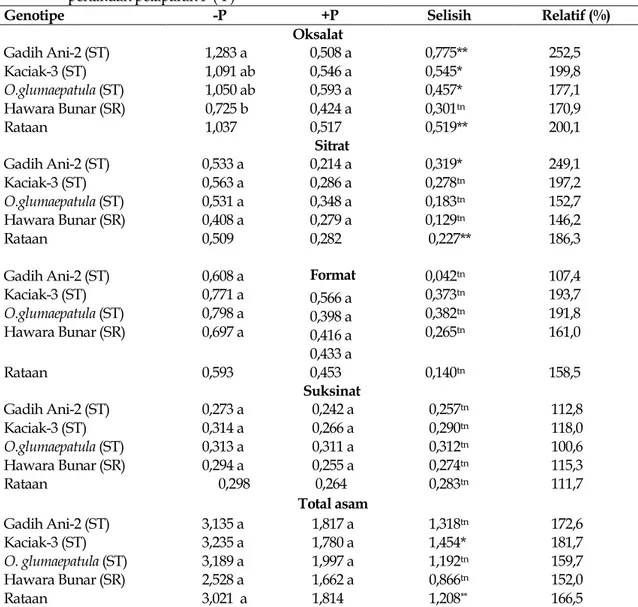
Dari enam asam organik yang terdeteksi, dua asam organik (tartarat dan malat) dihasilkan dalam jumlah yang lebih sedikit dibanding yang lainnya (Tabel 1). Perlakuan pelaparan P meningkatkan dengan nyata eksudasi asam oksalat pada ketiga genotipe sangat tenggang, eksudasi asam sitrat pada genotipe Gadih Ani-2, dan asam format pada genotipe Kaciak-3 (Tabel 2). Walaupun tidak terlihat perbedaan nyata, peningkatan eksudasi sitrat pada Kaciak-3 dan eksudasi format pada
O. glumaepatula cukup tinggi (berturut-turut 97% dan 92%). Tidak terlihat peningkatan eksudasi asam suksinat pada semua genotipe.
Pada kondisi kahat P, perbedaan nyata hanya terlihat pada eksudasi asam oksalat yakni antara genotipe Gadih Ani-2 dengan Hawara Bunar. Namun demikian, secara angka-angka terlihat perbedaan yang cukup besar eksudasi asam oksalat dan format antara Kaciak-3 dan O. glumaepatula dengan Hawara Bunar, serta eksudasi asam sitrat antara geno-tipe sangat tenggang dengan genogeno-tipe sangat rentan. Berdasarkan hal ini dapat disimpulkan,,
peningkatan eksudasi oksalat dan sitrat adalah salah satu mekanisme ketenggangan tanaman padi terhadap P rendah di tanah sawah, se-dangkan format hanya pada genotipe tertentu.
Total eksudasi asam organik (termasuk malat dan tartarat) hanya meningkat dengan nyata pada genotipe Kaciak-3. Pada genotipe lainnya, walaupun tidak nyata secara statistik, juga terdapat peningkatan sebanyak 52 sampai 73%. Pada kondisi kahat P, ketiga genotipe sangat tenggang menunjukkan eksudasi asam organik total yang lebih tinggi dibanding genotipe sangat rentan.
Tabel 1. Eksudasi asam organik per tanaman (μmol) empat genotipe padi empat hari setelah perlakuan pelaparan P (-P)
Genotipe -P +P Selisih Relatif (%)
Oksalat Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 1,283 a 1,091 ab 1,050 ab 0,725 b 0,508 a 0,546 a 0,593 a 0,424 a 0,775** 0,545* 0,457* 0,301tn 252,5 199,8 177,1 170,9 Rataan 1,037 0,517 0,519** 200,1 Sitrat Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 0,533 a 0,563 a 0,531 a 0,408 a 0,214 a 0,286 a 0,348 a 0,279 a 0,319* 0,278tn 0,183tn 0,129tn 249,1 197,2 152,7 146,2 Rataan 0,509 0,282 0,227** 186,3 Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 0,608 a 0,771 a 0,798 a 0,697 a Format 0,566 a 0,398 a 0,416 a 0,433 a 0,042tn 0,373tn 0,382tn 0,265tn 107,4 193,7 191,8 161,0 Rataan 0,593 0,453 0,140tn 158,5 Suksinat Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 0,273 a 0,314 a 0,313 a 0,294 a 0,242 a 0,266 a 0,311 a 0,255 a 0,257tn 0,290tn 0,312tn 0,274tn 112,8 118,0 100,6 115,3 Rataan 0,298 0,264 0,283tn 111,7 Total asam Gadih Ani-2 (ST) Kaciak-3 (ST) O. glumaepatula (ST) Hawara Bunar (SR) Rataan 3,135 a 3,235 a 3,189 a 2,528 a 3,021 a 1,817 a 1,780 a 1,997 a 1,662 a 1,814 1,318tn 1,454* 1,192tn 0,866tn 1,208** 172,6 181,7 159,7 152,0 166,5
Angka pada kolom dengan huruf kecil sama, tidak berbeda nyata menurut DMRT 0,05 -P, +P: dengan dan tanpa pelaparan P. ST, SP= sangat tenggang dan sangat rentan attermasuk malat dan tartarat
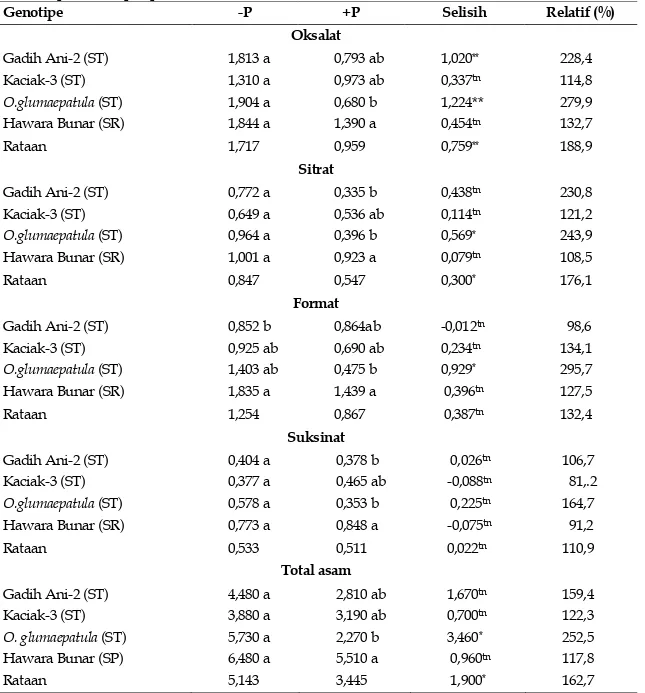
Eksudasi Asam Organik per Bobot Kering Tajuk
Kahat P meningkatkan dengan nyata eksudasi asam oksalat, sitrat, format, dan total
asam per bobot kering tajuk pada genotipe O. glumaepatula. Genotipe ini juga memperlihatkan kenaikan asam suksinat yang cukup tinggi (65%) (Tabel 2).
Genotipe Gadih Ani-2 hanya menunjukkan kenaikan nyata pada asam oksalat dan kenaikan cukup tinggi (131%) pada asam sitrat.
Genotipe Kaciak-3 dan Hawara Bunar tidak memperlihatkan kenaikan nyata pada semua asam, bahkan ada yang menurun.
Pada kondisi kahat P terlihat
kecenderungan yang berbeda dengan
kecenderungan pada peubah eksudasi asam
organik per tanaman. Kecuali pada asam oksalat, ternyata genotipe sangat rentan menunjukkan angka yang lebih tinggi dibanding genotipe sangat tenggang. Dengan demikian, eksudasi pertanaman yang tinggi pada genotipe sangat tenggang dalam kondisi kurang P lebih disebabkan oleh tingginya bobot kering tanaman.
Tabel 2. Eksudasi asam organik per g tajuk (μmol/g) empat genotipe padi empat hari setelah perlakuan pelaparan P
Genotipe -P +P Selisih Relatif (%)
Oksalat Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 1,813 a 1,310 a 1,904 a 1,844 a 0,793 ab 0,973 ab 0,680 b 1,390 a 1,020** 0,337tn 1,224** 0,454tn 228,4 114,8 279,9 132,7 Rataan 1,717 0,959 0,759** 188,9 Sitrat Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 0,772 a 0,649 a 0,964 a 1,001 a 0,335 b 0,536 ab 0,396 b 0,923 a 0,438tn 0,114tn 0,569* 0,079tn 230,8 121,2 243,9 108,5 Rataan 0,847 0,547 0,300* 176,1 Format Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 0,852 b 0,925 ab 1,403 ab 1,835 a 0,864ab 0,690 ab 0,475 b 1,439 a -0,012tn 0,234tn 0,929* 0,396tn 98,6 134,1 295,7 127,5 Rataan 1,254 0,867 0,387tn 132,4 Suksinat Gadih Ani-2 (ST) Kaciak-3 (ST) O.glumaepatula (ST) Hawara Bunar (SR) 0,404 a 0,377 a 0,578 a 0,773 a 0,378 b 0,465 ab 0,353 b 0,848 a 0,026tn -0,088tn 0,225tn -0,075tn 106,7 81,.2 164,7 91,2 Rataan 0,533 0,511 0,022tn 110,9 Total asam Gadih Ani-2 (ST) Kaciak-3 (ST) O. glumaepatula (ST) Hawara Bunar (SP) Rataan 4,480 a 3,880 a 5,730 a 6,480 a 5,143 2,810 ab 3,190 ab 2,270 b 5,510 a 3,445 1,670tn 0,700tn 3,460* 0,960tn 1,900* 159,4 122,3 252,5 117,8 162,7
Angka pada kolom dengan huruf kecil sama tidak berbeda nyata menurut DMRT 0,05 ; **, *, tn sangat nyata, nyata, dan tidak nyata; -P, +P dengan dan tanpa pelaparan P
Total asam hanya memperlihatkan kenaikan nyata (150%) pada O. glumaepatula, kenaikan cukup tinggi (59%) pada Gadih Ani-2, dan
kenaikan lebih rendah (22%) pada Kaciak-3.
Kanaikan asam total ini kurang
genotipe. Hal ini menunjukkan bahwa yang menentukan bukan hanya jumlah asam tetapi juga jenisnya, karena kemampuan tiap jenis asam organik dalam desorbsi ataupun solubilisasi bebeda (Bar-Yosef 1991)
Menurut Hoffland et al. (1990) peningkatan eksudasi asam organik pada tanaman yang
mengalami cekaman P adalah karena peningkatan aktivitas fosfoenol piruvat karboksilase (PEPC) pada tajuk akibat penurunan kadar P. Pada penelitian ini terlihat penurunan yang tajam kadar P baik pada tajuk maupun akar, dengan penurunan yang lebih besar terjadi pada kadar P tajuk (Tabel 3).
Tabel 3. Kadar P tajuk dan akar (%) empat genotipe padi tanpa perlakuan pelaparan P (+P) dan dengan perlakuan pelaparan (-P)
Genotipe +P -P Penurunan (%) +P -P Penurunan (%)
Tajuk Akar Gadih Ani-2 Kaciak-3 O. glumaepatula Hawara Bunar 0,735 1,085 0,897 1,180 0,080 0,067 0,068 0,100 89,1 93,8 92,4 91,5 0,465 0,569 0,510 0,669 0,077 0,083 0,095 0,094 83,4 85,4 81,4 85,9 Rataan 0,959 0,079 91,7 0,553 0,087 84,0
Eksudasi per Bobot Kering Akar
Kondisi kahat P menurunkan dengan signifikan jumlah eksudasi semua asam organik per bobot kering akar pada semua genotipe, kecuali pada O. glumaepatula dimana hampir
tidak terlihat penurunan. Pengaruh status P terhadap jumlah eksudasi asam organik total per bobot kering akar dapat dilihat pada Gambar 1.
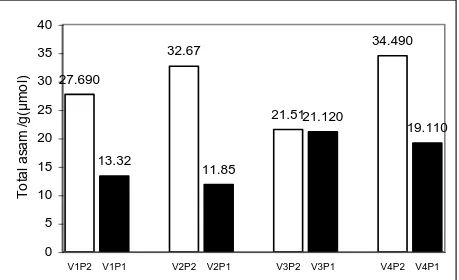
Gambar 1. Pengaruh pelaparan P terhadap eksudasi asam organik total per bobot kering akar. V1, V2, V3, V4 berturut-turut Gadih Ani-2, Kaciak-3, O. glumaepatula, dan Hawara Bunar. P1, P2 kondisi kahat dan cukup P
Turunnya jumlah eksudasi asam organik per bobot kering akar pada kondisi kahat P adalah akibat terjadinya peningkatan bobot kering akar yang jauh lebih tinggi (216%) (Tabel 4) dibanding peningkatan eksudasi per tanaman yang hanya 66% (Tabel 1). Penurunan kadar P akar (Tabel 3) ternyata tidak meningkatkan
eksudasi yang seimbang dengan peningkatan bobot akar.
Menurut Hoffland et al. (1990) pada akar
Brassica napus penurunan kadar P tidak meningkatan aktivitas PEPC dan sintesis asam organik. Peningkatan eksudasi asam organik juga tidak berhubungan dengan kebocoran akar. Peningkatan perakaran akibat pelaparan P
27.690 13.32 32.67 11.85 21.5121.120 34.490 19.110 0 5 10 15 20 25 30 35 40 V1P2 V1P1 V2P2 V2P1 V3P2 V3P1 V4P2 V4P1 T o ta l a sa m /g (μ mo l)
pada tanaman padi yang sebelumnya ditumbuhkan pada medium P cukup juga dilaporkan Ishizuka cit. Takenaga (1995). Hasil ini berbeda dengan hasil penelitian Lipton
et al.(1985) yang menemukan peningkatan eksudasi asam organik per bobot kering akar pada tanaman jagung kahat P serta penelitian Jones dan Darrah (1995) pada tanaman jagung
kahat P dan hara mikro. Perbedaan ini diduga adalah akibat perbedaan praperlakuan sebelum pengumpulan asam organik. Praperlakuan tanaman yang sebelumnya ditumbuhkan pada medium P cukup pada medium tanpa P untuk menginduksi kondisi kahat P mengakibatkan terjadinya peningkatan pertumbuhan akar. Tabel 4. Bobot kering akar (g) empat genotipe padi dengan dan tanpa pelaparan P di larutan hara
Genotipe -P +P Selisih Relatif (%)
Gadih Ani-2 Kaciak-3 O. glumaepatula Hawar Bunar 0,238 a 0,273 a 0,147 b 0,133 b 0,066 ab 0,058 ab 0,093 a 0,049 b 0,173** 0,215** 0,054* 0,084** 360,6 470,7 157,5 273,2 Rataan 0,198 0,066 0,131 315,8
Angka pada kolom yang diikuti huruf kecil sama tidak berbeda nyata dengan uji DMRT. *, ** nyata dan sangat nyata; -P, +P kondisi kahat dan cukup P
KESIMPULAN
1. Teridentifikasi eksudasi enam jenis asam organik (oksalat, sitrat, tartarat, format, malat, dan suksinat) pada tanaman padi dengan status P rendah mapun cukup.
2. Status P rendah pada tanaman meningkatkan eksudasi asam organik per tanaman dan per bobot kering tanaman dengan peningkatan ter-tinggi (2 x lipat) terlihat pada oksalat dan sitrat. 3. Peningkatan eksudasi oksalat per bobot kering tajuk akibat pelaparan P terlihat pada dua dari tiga genotipe sangat tenggang yakni Gadih Ani-2 dan O. glumaepatula, dan untuk sitrat dan format pada O. Glumaepatula; sehingga disim-pulkan bahwa peningkatan eksudasi asam organik merupakan salah satu mekanisme ketenggangan terhadap P rendah, namun tidak terdapat pada semua genotipe tenggang.
DAFTAR PUSTAKA
Ae N, Arihara J, Okada K, Yoshihara T, Otani T, and Johansen C. 1993. The role of piscidic acid secreted by pigeonpea roots grown in an Alfisol with low-P fertility. Dalam Genetic Aspects of Plant Mineral Nutrition. Randall PJ, Delhaize E, Richards RA, and Munns R (eds) p 270-288. Kluwer Academic Publishers, Dordrecht.
Bar-Yosef B. 1991. Root excretions and their environmental effects. Dalam Plant Roots, the Hidden Half. Marcel Dekker.
Hoffland E, Findenegg GR, and Nelemans JA. 1989 Solubilization of rock phosphate by rape. II. Local root exudation of organic acids as a response to P-starvation. Plant and Soil 113: 161-165.
Lipton DS, Blnchar RW, and Blevins DG. 1987. Citrate, malate, succinate concentration in exudates from P-sufficient and P-stressed
Medicago sativa L. seedlings. Plant Physiol 85: 315-317.
Marschner H. 1995. Mineral Nutrition of Higher Plants. London, UK. Asscociated Press. Ohwaki Y and Hirata H. 1990. Phosphorus
absorption by chickpea (Cicer arietinum) as affected by VA mycorrrhiza and carboxylic acids in root exudates. Dalam Beusichem ML
Plant Nutrition – Physiology and application. 171-177. Kluwer Academic Publishers.
Otani T, Ae N, and Tanaka H. 1996. Phosphorus uptake mechanism of crops grown in soils with low P status. II. Significant of organic acid in root exudates of pigeonpea. Soil Science and Plant Nutrition 42 : 553-560.
Pellet DM, Grunes DL, and Kochian LV. 1995. Organic acid exudation as an aluminum-tolerance mechanism in maize (Zea mays L.).
Rao IM, Friesen DK and Osaki M. 1999. Plant adaptation to phosphorus limited tropical soils. Dalam Pessarakli M (ed) Handbook of Plant and Crop Stress. New York. Marcel Dekker.
Subbarao GV, Ae N, and Otani T. 1997. Genetic variation in acquisition and utilization of phosphorus from iron-bound P in pigeonpea.
Soil Science and Plant Nutrition 43: 511-519. Swasti E. 2004. Fisiologi dan pewarisan sifat
efi-siensi fosfor pada padi gogo dalam keadaan tercekam aluminium [disertasi]. Bogor. Sekolah Pascasarjana Institut Pertanian Bogor.
Takenaga H. 1995. Nutrient absorbtion in relation to environment. Dalam Science of the Rice Plant. Vol II. Physiology. 278-294.
Yoshida S, Forno DA, Cock JH, Gomez KA. (1976).
Laboratory Manual for Physiological Studies of Rice. International Rice Research Institute. Los Banos, Philippines.