EMBRYO CULTURE
Embryo maturity plays an important role for the successful
cryopreservation of coconut (Cocos nucifera)
Sisunandar&Hengky Novarianto&Nurhaini Mashud& Yohannes M. S. Samosir&Steve W. Adkins
Received: 22 January 2014 / Accepted: 1 July 2014 / Published online: 7 August 2014 / Editor: John Forster #The Society for In Vitro Biology 2014
Abstract Genetic diversity of coconut (Cocos nuciferaL.) is being lost due to a combination of pest and disease attack, urban encroachment, natural disasters, as well as introgression with exotic genetic types. Consequently, there is a need to undertake germplasm conservation before further loss occurs. Since coconut has a large, recalcitrant seed (sensitive to des-iccation), it cannot be stored in traditional ways in a seed bank. Cryopreservation of zygotic embryos is now seen as an im-portant storage approach although published techniques are still not reliable. Given the importance of embryo maturity to the success of cryopreservation in other species, the effect of coconut embryo maturity on cryopreservation success was investigated using four cultivars (‘Nias Yellow Dwarf’, ‘Tebing Tinggi Dwarf’, ‘Takome Tall’, and ‘Bali Tall’). After cryopreservation, using a new four-step protocol (rapid desiccation, rapid freezing, rapid thawing, and recovery and acclimatization for 4 mo in the glasshouse), we found that the embryos isolated from an 11-mo-old fruit gave the highest number of normal seedlings (~28%) when compared to coun-terparts excised from younger fruits. In addition, the results
showed that fruit could be stored for up to 3 wk prior to embryo isolation before their performance in cryopreservation was compromised.
Keywords Embryo maturity . Cryopreservation . Coconut . Cocos nucifera
Introduction
Coconut (Cocos nuciferaL.) is one of the most important tree crops in the tropics. It is grown by more than 50 million people on 12 million ha of land and in over 90 countries. Crop productivity is continuously being threatened due to a combination of pest and disease attack and natural disasters. As a consequence, the loss of locally well-adapted cultivars has become a major problem, andex vitromethods to con-serve coconut germplasm have been urgently sought. Traditionally, seed gardens have been used to conserve coco-nut germplasm (Batugal and Jayashree2005). However, more recently, a new approach for the long-term storage of coconut germplasm has been developed using cryopreservation (Sisunandar et al. 2010b). This new approach uses zygotic embryos as the explant material and has been shown to be efficient in producing a relatively large number of seedlings ready for field planting after cryopreservation. Furthermore, no significant differences in their genetic and epigenetic char-acteristics prior to and after cryopreservation have been ob-served using this approach (Sisunandaret al. 2010a). In its present form, the approach uses embryos isolated from an 11-mo-old fruit; however, the effect of embryo maturity upon the success of the approach has not been determined.
A number of protocols have been developed for the suc-cessful cryopreservation of plants using zygotic embryos (Pritchard and Prendergast 1986; Abdelnour-Esquivel et al. 1992; Abdelnour-Esquivel and Engelmann 2002; Walters Sisunandar:Y. M. S. Samosir
:
S. W. Adkins (*)School of Agriculture and Food Sciences, The University of Queensland, St Lucia, Brisbane, Queensland 4072, Australia e-mail: [email protected]
H. Novarianto:N. Mashud
Indonesian Coconut and Other Palm Research Institute, Mapanget, Manado, North Sulawesi, Indonesia
Present Address: Sisunandar
Biology Education Department, The University of Muhammadiyah Purwokerto, Kampus Dukuhwaluh, Purwokerto 53182, Indonesia
Present Address: Y. M. S. Samosir
et al.2002; Nadarajanet al.2007; Steinmacheret al.2007). In most of these protocols, the stage of embryo maturity (development) was a factor in determining the degree of success. For orthodox species, fully mature embryos are those that respond best to cryopreservation (Kenmode and Finch-Savage2002; Wen and Song2007); however, for recalcitrant species like coconut, the best embryo maturity stage seems to be species dependent. For example, for coffee (Coffeaspp.; Abdelnour-Esquivelet al. 1992) and jackfruit (Artocarpus heterophyllusLamk.; Chandel et al. 1995; Chaudhury and Malik2004), an intermediate stage of embryo maturity was the best, while for tea (Camellia sinensisL.; Kimet al.2002), the fully mature embryo stage was the best. In earlier studies where embryo maturity was studied in the cryopreservation process of coconut, Chinet al.(1989) reported that immature embryos survived better after cryopreservation, while Assy-Bah and Engelmann (1992a,b) reported fully mature embryos to give the better survival after cryopreservation.
In determining the best embryo maturity stage for cryo-preservation, it is important to take into account the age of the fruit at harvest and the duration of storage before the embryo is removed. This post-harvest time may include the time taken in transporting the fruit from the field to the laboratory or the time the fruit remains in storage in the laboratory prior to the cryopreservation of the embryo (Berjaket al.1992; Barbedo et al. 1999; Demir and Okcu 2005). On many occasions, coconut fruits have to be harvested from remote areas, and the time in transport can be several weeks. On the other hand, some freshly harvested fruits may have to sit in storage for some time before staff can undertake embryo isolation and the cryopreservation step can take place. Thus, this present study evaluates embryo maturity status and post-harvest storage time on the success of the newly developed cryopreservation protocol for coconut (Sisunandaret al.2010a).
Materials and Methods
Plant material. The four coconut cultivars used in this study (viz., ‘Nias Yellow Dwarf’ (NYD); ‘Tebing Tinggi Dwarf’
(TTD); ‘Takome Tall’ (TKT); and ‘Bali Tall’ (BAT)) representing two tall and two dwarf cultivars were harvested from the Mapanget Research Seed Garden (The Indonesian Coconut and Other Palm Research Institute, Manado, Indonesia). From each cultivar, four different embryo maturity stages (viz., 8, 9, 10, or 11 mo after pollination) were harvest-ed, based on a leaf counting method. This method assumes that a new coconut leaf and inflorescence is produced every 4 wk (Pereraet al.2008). Thus, the 8th fruit bunch down from the newly opened inflorescence contains embryos that are 8 mo old. Twelve-mo-old embryos were not used in this present study as such embryos usually start to germinate at this age on at least one cultivar under study.
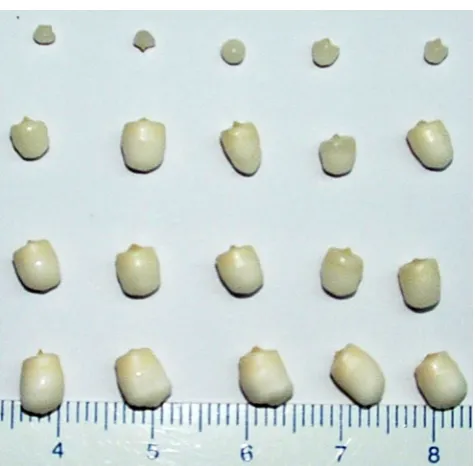
Embryo characteristics(experiment 1). Six hundred and forty embryos (40 from each of the four cultivars and from each of the four different maturity stages) were aseptically isolated from the fruit using the previously described method of Rillo (2004). For the isolation of embryos from very young (8-mo old) fruit, a small metal spoon was used to scoop them from the soft endosperm tissue. All embryos were then surface sterilized using a sodium hypochlorite (2.6%; v/v) solution for 15 min, followed by several rinses in sterile water, and then blotted dry on sterile filter paper. From each of the 40 embryos from each of the four cultivars and four maturity stages, 10 embryos (in two replicates of five) were photographed, and their length and cross-sectional area determined using a free-to-access computer software package Image J 1.37v (Rasband 2006). The fresh weight of these same embryos was then determined using a Precise XT220A analytical balance, and dry weights were determined gravimetrically following oven drying at 103±2°C for 24 h. The remaining 30 embryos (in three replicates of 10) from each batch were germinated as previously described (Rillo2004). Briefly, the embryos were placed into a liquid embryo culture medium for 1 mo, then transferred to a solid medium for a further 2 mo, then to a second liquid culture medium for 2 mo. The seedlings after this final stage were then acclimatized and grown in a glass-house (Sisunandar et al. 2010a). The percentage of viable (embryos showing either root or shoot production), germinat-ing (embryos showgerminat-ing both root and shoot production), and embryos producing seedlings of normal morphology in soil in the glasshouse were determined.
Maturity and cryopreservation(experiment 2). One day fol-lowing fruit harvest, a further 1,120 embryos (70 from each of the four cultivars and from each of the four different maturity stages) were aseptically isolated using the previously de-scribed method (Rillo 2004; with modification for the very young embryos) and were allocated to the following treat-ments. Ten embryos (in two replicates of five) were then rapidly dehydrated for 4, then 6, then 8, and finally 10 h in a rapid drying chamber as previously described (Sisunandar et al. 2010b, a; 2012) with their moisture content being determined at each time period. A further 30 embryos (in three replicates of 10) from each batch were dehydrated for 8 h (Sisunandaret al. 2010a) and then germinated as described above (Rillo2004). The remaining 30 embryos (in three replicates of 10 embryos) were dehydrated for 8 h and then plunged into liquid nitrogen where they remained for 24 h (Sisunandar et al. 2010a). Upon recovery, their viability, germination, and ability to produce seedlings of normal morphology in soil in the glasshouse were determined (Sisunandar et al. 2010a).
NYD and stored at 29±3°C and a relative humidity of 79±5% for either 1, 2, or 3 wk. Every week, the embryos from 60 randomly selected fruits were isolated and (in three replicates of 20 embryos) were rapidly dehydrated for 8 h and then plunged into liquid nitrogen where they remained for 24 h (Sisunandaret al.2010a). Upon recovery, their viability, ger-mination, and ability to become seedlings of normal morphol-ogy in soil in the glasshouse were determined (Sisunandar et al.2010a).
Experimental design and statistical analysis. All experiments were undertaken using a completely randomized design with 5, 20, or 30 embryos per replication, as reported above. An analysis of variance (ANOVA) was undertaken using the Minitab™ statistical software package (Minitab Inc., Belmont, CA). Some of the data sets (those violating the ANOVA principles such as homogeneity of variances) were square-root transformed prior to statistical analysis. However, only untransformed data sets are presented here. All experi-ments were repeated with similar results to those reported here.
The present studies were undertaken (1) to determine the best time for zygotic embryo isolation from fruit to optimize of the production of normal seedlings after cryopreservation and (2) to determine if a short post-harvest storage period of up to 3 wk will affect the cryopreservation process.
Results
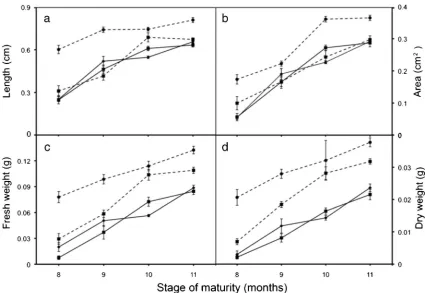
Embryo characteristics(experiment 1). In visual terms, em-bryos of the four cultivars exhibited a similar pattern of maturation over time (Fig. 1). Embryo sizes (length and cross-sectional area) increased rapidly in 8- to 10-mo-old fruits, then to a lesser extent over the next 2 mo (Fig. 2). Similar patterns of development were observed for their fresh and dry weight gain (Fig.2). One minor difference noticed was that the embryos coming from the two tall cultivars (i.e., TKT and BAT) were larger than those of the two dwarf cultivars (i.e., NYD and TTD) when at the same time of maturity.
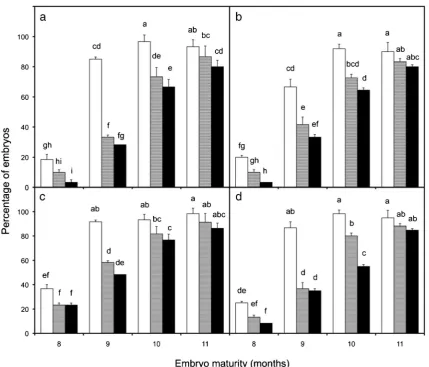
The time-related stage at which the embryos were isolated from the fruit had a significant impact upon their viability (growth observed for either root or shoot) and their ability to germinate (growth observed from both root and shoot). Embryos of 11 mo old were more viable and germinated at a much higher percentage than those that were 8 mo old (Fig.3). The older embryos not only gave the highest viability and germination percentages but also gave the highest number of normal seedlings (up to 90%) growing in soil. Meanwhile, the 8-mo-old embryos gave the lowest percentage of normal
seedlings (up to 20%) growing in soil. No differences in the ability to produce normal seedlings from 11-mo-old embryos were noticed between the four cultivars.
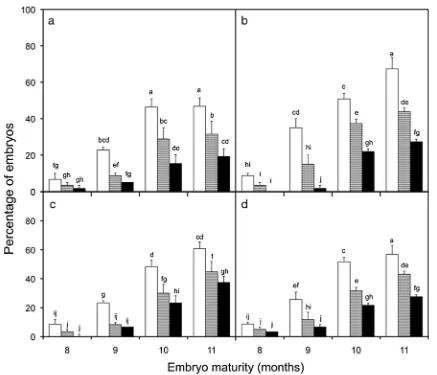
Maturity and cryopreservation (experiment 2). Irrespective of maturity status, all embryos of NYD underwent a similar rate of dehydration when placed in the rapid drying chamber (Fig. 4). During the first 6 h of dehy-dration, water loss followed a simple exponential func-tion and declined from around 80% to around 30%, on a fresh weight basis. The rate of water loss then de-clined and after 8 h reached the optimum level of 20% for cryopreservation (Sisunandar et al. 2010b).
of 21% being produced for NYD and a high value of 36% being produced for TKT (Fig.5).
Storage and cryopreservation(experiment 3). Post-harvest storage of fruit initially containing 11-mo-old NYD embryos for either 1, 2, or 3 wk caused no significant changes to the embryo moisture content (data not shown). Moreover, these storage treatments did not affect embryo viability (~65%) or their ability to germinate (~35%) and produce normal seed-lings (~25%) after cryopreservation (Fig.6). Thus, no signif-icant differences in the ability to cryopreserve the embryos were detected following fruit storage for 1, 2, or 3 wk.
Discussion
The success of cryopreservation in many species relies upon using tissues of an appropriate physiological age as has been demonstrated for a number of species including coffee (Abdelnour-Esquivelet al.1992), jackfruit (Chaudhury and Malik2004), and tea (Chandelet al.1995; Kimet al.2002). For coconut, both immature and mature embryos have been used in the past for cryopreservation but with different out-comes (Bajaj 1984; Chin et al. 1989; Assy-Bah and
Engelmann1992b). Thus, it was necessary to re-evaluate the effect of embryo maturity on the success of cryopreservation and to do this for a range of coconut cultivars, including both tall and dwarf cultivars.
A similar pattern of embryo development was observed for all of the four cultivars of coconut studied. This similar devel-opment was observed in length and cross-sectional area at-tainment (Fig. 1) and fresh and dry weight accumulation (Fig.2). In general terms, the two tall cultivars produced larger embryos than the dwarf cultivars, and this has been reported before (Thampan1981; Foale2003), with BAT being signif-icantly (P>0.05) bigger in all measured variables than all other cultivars (Fig.3). There was also a tendency for greater growth in the early period of development but with this decreasing in embryos greater than 9 mo old. This embryo maturity pattern is similar to that seen in other species such as jackfruit (Farant and Walters1998), wheat (Triticum aestivum; Lehner et al. 2006), and maize (Zea mays; Wen and Song 2007) where maturity was rapid in young embryos, but then declined as embryos approached physiological maturity. The percentage moisture content of embryos found throughout all stages of maturity was about 80% and was similar for all four cultivars (Fig.4). Such a high level of moisture content within its embryos and the lack of any significant moisture loss during maturation indicate that coconut is a recalcitrant Figure 2. Changes in embryo attributes measured at four maturity stages
and for four cultivars: NYD ( ), TTD ( ), TKT ( ), and BAT ( ).aThe longitudinal length,barea,cfresh weight, anddembryo
species (Engelmann 1999). Similar high embryo moisture content has been observed in other recalcitrant species such as landolphia (Landolphia kirkiiDyer; Berjaket al.1992) and jackfruit (Chaudhury and Malik 2004). Moisture retention during maturation in recalcitrant species is in contrast to that seen in orthodox species in which the embryo moisture con-tent decreases dramatically as physical maturity approaches, as seen in wheat (Lehneret al.2006) and maize (Wen and Song2007).
As might be predicted, the germination capacity of embry-os from all four cultivars increased as they matured. Embryembry-os isolated from 11-mo-old fruit produced the highest germina-tion (~90%) and gave the highest percentage of normal seed-lings (~80%), while those isolated from 8-mo-old fruit pro-duced the lowest germination (~10%) and gave the lowest percentage of normal seedlings (~10%; Fig.3). Similar obser-vations have been made on other species, such as in areca palm (Chrysalidocarpus lutescens; Broschat and Donselman 1986), pigmy date palm (Phoenix roebelenii), royal palm Figure 3. The re-growth characteristics of embryos taken from fruit of
cultivars,aNYD,bTTD,cTKT, anddBAT embryos at four different maturity stages (8, 9, 10, or 11 mo old) and placed onto a recovery medium for 8 wk. The data show the percentage of viable (□),
germinating ( ), and embryos producing normal seedlings (■).Bars below different lettersare significantly different atP≤0.05. Thebars represent means±SE of three replications of 10 embryos.
Figure 5. The viability and germination of embryos taken from fruit of cultivars,aNYD,bTTD,cTKT, anddBAT at four different maturity stages (8, 9, 10, and 11 mo old) that had been dehydrated, cryopreserved, and placed onto a recovery medium for 8 wk. The data show the
percentage of viable (□), germinating ( ), and embryos producing normal seedlings (■).Bars below different lettersare significantly differ-ent at P≤0.05. The bars are means±SE of three replications of 20 embryos.
Figure 6. The re-growth attributes of embryos taken from fruit of cultivar ‘Nias Yellow Dwarf’isolated from 11-mo-old fruit that had been stored for either 0, 1, 2, or 3 wk after harvest. Embryos had been dehydrated for 8 h using a rapid dehydration apparatus, cryopreserved, and placed onto a
(Roystonea regia; Broschat and Donselman 1987), bitter vetch (Vicia ervilia; Samarah et al. 2003), and jackfruit (Chaudhury and Malik2004).
When embryos from one cultivar (NYD) and at different stages of maturity were dehydrated, their water loss profiles were similar (Fig.4), reaching about 20% moisture content after 8 h of drying. This observation is similar to that of an earlier study (Chaudhury and Malik2004) where jackfruit embryos, at different stages of maturity, all dried at a similar rate.
Embryos isolated from 11-mo-old fruit gave the highest germination percentage (~40%) and the highest number of normal seedlings (~30%) after cryopreservation (Fig.5). This finding is similar to that of Assy-Bah and Engelmann (1992a,b), who found that mature coconut embryos gave the best survival percentage after cryopreservation than did immature embryos. Thus, mature embryos, isolated from 11-mo-old fruit, are those recommended for coconut cryopreservation. This is similar to jackfruit where ~30% of the mature embryos survived cryopres-ervation, while no immature embryos survived (Chaudhury and Malik 2004). These results indicate an increase in embryo maturity correlates with an increase in the tolerance of embryos to dehydration and cryopreservation, which is a trend that has been reported for many orthodox species (Hong and Ellis1990; Wen and Song2007).
The four cultivars used in this experiment all had similar responses to cryopreservation when the percentage of viable embryos and the percentage of germinating embryos were taken into account. However, the percentage of embryos that went on to produce normal seedlings did show significant differences between cultivars, NYD giving the lowest (21%) and TKT giving the highest (36%; Fig. 5) percentage of normal seedlings. This is a similar result to that reported before (Sisunandaret al.2010a).
During transport of fruit from the field to the laboratory, or during storage at the laboratory, the embryos contained within the fruit will continue to develop (Berjaket al.1992), espe-cially if this time becomes greater than 1 wk. In recalcitrant species such as coconut, which can germinate soon after harvest (i.e., within 30 to 40 d after harvest; Foale 2003), significant delays in post-harvest handling, prior to cryopres-ervation, may result in lower embryo germination or losses in embryo quality. Often, delay is unavoidable as germplasm collection takes place in very remote regions, several days away from the laboratory, and the processing of large numbers of fruit in the laboratory takes time to do. The quality changes that go on inside embryos during this transport and storage period may lead to changes in the tolerance of embryos to dehydration and/or cryopreservation success (Berjak et al. 1992). However, in the present study, storage of fruit in a typical tropical storage at ambient temperature for up to 3 wk did not influence embryo moisture content (data not shown), or their viability, germination, and normal seedling production
capacity after cryopreservation (Fig.6). Such storage condi-tions used are not identical to those found with long-distance transport to a laboratory, but do indicate that transport times of up to 3-wk duration should not affect the cryopreservation outcomes of the embryos contained within the fruit.
Acknowledgments The project was partly funded by the Australian Agency for International Development (AusAID), the Endeavour Re-search Fellowships Australia 2010, and the Australian Centre for Inter-national Agricultural Research,viaproject HORT/1998/061.
References
Abdelnour-Esquivel A, Engelmann F (2002) Cryopreservation of chayo-te (Sechium eduleJacq. SW) zygotic embryos and shoot tips from in vitroplantlets. CryoLetters 23:299–308
Abdelnour-Esquivel A, Villalobos V, Engelmann F (1992) Cryopreservation of zygotic embryos ofCoffeaspp. CryoLetters 13:297–302
Assy-Bah B, Engelmann F (1992a) Cryopreservation of immature em-bryos of coconut (Cocos nuciferaL.). CryoLetters 13:67–74 Assy-Bah B, Engelmann F (1992b) Cryopreservation of mature embryos
of coconut (Cocos nuciferaL.) and subsequent regeneration of plantlets. CryoLetters 13:117–126
Bajaj YPS (1984) Induction of growth in frozen embryos of coconut and ovules of citrus. Current Sci 53:1215–1216
Barbedo C, Barbedo A, Nakagawa J, Sato O (1999) The effect of fruit age and post-harvest period of cucumber on stored seeds. Pesq Agrop Brasileira 34:839–847 (in Portuguese)
Batugal P, Jayashree J (2005) COGENT's multi-site international coconut genebank. In: Batugal P, Ramanatha Rao V, Oliver J (eds) Coconut genetic resources. International Plant Genetic Resources Institute-Regional Office for Asia, the Pacific and Oceania (IPGRI-APO), Serdang, Selangor DE, Malaysia, pp 106–114
Berjak P, Pammenter NW, Vertucci CW (1992) Homiohydrous (recalcitrant) seed: developmental status, desiccation sensitivity and state of water in axes ofLandolphia kirkiiDyer. Planta 186: 249–261
Broschat TK, Donselman H (1986) Factors affecting storage and germi-nation ofChysalidocarpus lutescensseeds. J Am Soc Hort Sci 111: 872–877
Broschat TK, Donselman H (1987) Effect of maturity, storage, presoaking and seed cleaning on germination in three species of palms. J Environ Hort 5:6–9
Chandel KPS, Chudhury R, Radhamani J, Malik SK (1995) Desiccation and freezing sensitivity in recalcitrant seeds of tea, cocoa and jack-fruit. Ann Bot 76:443–450
Chaudhury R, Malik SK (2004) Desiccation and freezing sensitivity during seed development in jackfruit. Seed Sci Tech 32:785–795 Chin HF, Krishnapillay B, Hor YL (1989) A note on the cryopreservation
of embryos from young coconut (Cocos nuciferavar MAWA). Pertanika 12:183–186
Demir I, Okcu G (2005) Effect of post-harvest maturation treatment on germination and potential longevity of pepper (Capsicum annuum) seed. Indian J Agric Sci 75:19–22
Engelmann F (1999) Cryopreservation of coconut germplasm. In: Oropeza C, Verdeil JL, Ashburner GR, Cardena R, Santamaria JM (eds) Current advances in coconut biotechnology. Kluwer Academic Publishers, Dordrecht, pp 289–296
Foale M (2003) The coconut odyssey: the bounteous possibilities of the tree of life. ACIAR Monograph No. 101, Australian Centre for International Agricultural Research, Canberra, 132 pp
Hong TD, Ellis RH (1990) A comparison of maturation drying, germina-tion and desiccagermina-tion tolerance between developing seeds of Acer pseudoplatanusL. andAcer platanoidesL. New Phytol 116:589–596 Kenmode AR, Finch-Savage WE (2002) Desiccation sensitivity in ortho-dox and recalcitrant seed in relation to development. In: Black MJ, Pritchard HW (eds) Desiccation and survival in plants: drying without dying. CABI Publishing, Wallingford, pp 149–184 Kim HH, Cha YS, Baek HJ, Cho EG, Chae TA, Engelmann F (2002)
Cryopreservation of tea (Camellia sinensisL.) seeds and embryonic axes. CryoLetters 23:209–216
Lehner A, Bailey C, Flechel B, Poels P, Come D, Corbineau F (2006) Changes in wheat seed germination ability, soluble carbohydrate and antioxidant enzyme activities in the embryo during the desicca-tion phase of maturadesicca-tion. J Cereal Sci 43:175–182
Nadarajan J, Staines HJ, Benson EE, Marzalina M, Krishnapillay B, Harding K (2007) Optimization of cryopreservation forSterculia cordatazygotic embryos using vitrification technique. J Trop Forest Sci 19:79–85
Perera PIP, Hocher V, Verdeil JL, Bandupriya HDD, Yakandawala DMD, Weerakoon LK (2008) Androgenic potential in coconut (Cocos nuciferaL.). Plant Cell Tiss Organ Cult 92:293–302
Pritchard HW, Prendergast G (1986) Effect of desiccation and cryopres-ervation on thein vitroviability of embryos of the recalcitrant seed speciesAraucaria hunsteiniiK. Schum J Ex Bot 37:1388–1397 Rasband WS (2006) ImageJ [Online]. Available by US National Institutes
of Health. Retrieved fromhttp://rsb.info.nih.gov/ij/. Accessed 2 Nov 2006
Rillo EP (2004) Importing and growing embryos for the coconut genebank. In: Ikin R, Batugal P (eds) Germplasm health manage-ment for COGENT’s multi-site international coconut genebank. International Plant Genetic Resources Institute-Regional Office for Asia, the Pacific and Oceania (IPGRI-APO), Serdang, Selangor DE, Malaysia, pp 62–68
Samarah N, Allataifeh N, Turk M, Tawaha AR (2003) Effect of maturity stage on germination and dormancy of fresh and air-dried seeds of bitter vetch (Vicia erviliaL.). NZ J Agric Res 46:347–354
Sisunandar, Sopade PA, Samosir Y, Rival A, Adkins SW (2010a) Dehydration improves cryopreservation of coconut (Cocos nucifera L.). Cryobiology 61:289–296
Sisunandar RA, Turquay P, Samosir Y, Adkins SW (2010b) Cryopreservation of coconut (Cocos nuciferaL.) zygotic embryos does not induce morphological, cytological or molecular changes in recovered seedlings. Planta 232:435–447
Sopade PA, Sopade PA, Samosir Y, Rival A, Adkins SW (2012) Conservation of coconut (Cocos nuciferaL.) germplasm at sub-zero temperature. CryoLetters 33:465–475
Steinmacher DA, Saldanha CW, Clement CR, Guerra MP (2007) Cryopreservation of peach palm zygotic embryos. CryoLetters 28: 13–22
Thampan PK (1981) Handbook on coconut palm. Oxford and IBH Publishing Co, New Delhi, 302 pp
Walters C, Touchell DH, Power P, Wesley-Smith J, Antolin MF (2002) A cryopreservation protocol for embryos of the endangered species Zizania texana. CryoLetters 23:291–298