MAKALAH BIOLOGI SEL DAN MOLEKULER
STRUKTUR DAN FUNGSI MIKROFILAMEN
Disusun oleh :
Kelompok 5
Setya Ambar Palupi
(13304144037)
Artika Anindyani Nur S. (13304144051)
Fauzia Latifa P.
(13304144054)
Dwi Astuti
(13304244017)
Triajeng Nur Amalia(13304244021)
Sebagian dari protein yang berada di sitosol, berbentuk jejala atau rerangka yang disebut sitoskelet atau rerangka sel. Sitoskelet ini memberi bentuk pada sel, mengatur dan menimbulkan gerakan sitoplasma yang beruntun dan berkaitan serta membentuk jaring-jaring kerja yang membantu mengatur reaksi-reaksi enzimatik (Issoegianti, 1993: 53).
Menurut Koolman (2005: 204), sitoplasma dilintasi oleh tiga struktur perancah dimensi yang didalamnya adalah filamen (jaring protein) yang bersama-sama membentuk sitoskeleton. Filament-filamen ini dibagi menjadi tiga bagian berdasarkan diameternya, yakni mikrofilamen (6-8 nm), intermediate filament (10 nm), dan mikrotubulus (25 nm). Seluruh filament ini adalah berupa polimer yang terbentuk dari komponen-komponen protein.
1. Mikrofilamen
Mikrofilamen ditujukan kepada semua elemen fibrosa yang memiliki garis tengah 60 angstrom dan terdiri dari molekul protein aktin. Aktin merupakan protein globular dengan BM 42000 dalton. Apabila berada dalam bentuk monomer disebut aktin G, yang dapat dirakit menjadi filamen beruntai rangkap dan disebut aktin F. (Reksoatmodjo, 1993: 61)
Dengan meningkatnya kekuatan ionik, kumpulan aktin G membentuk aktin F, homopolimer heliks. Aktin G membawa molekul ATP yang perlahan dihidrolisis untuk membentuk ADP (Koolman, 2005: 204). Menurut Cooper, (2006:474) tiap monomer aktin G tersusun menjadi 2 rantai yang membentuk filamen berotasi 1660 sehinggga mengikat satu
sama lain membentuk susunan double helix. Monomer-monomer aktin berorientasi atau mengarah pada arah yang sama, sehingga kedua ujung filamen aktin (ujung positif dan ujung negatif) mempunyai polaritas yang jelas di kedua ujungnya sehingga dapat dibedakan satu sama yang lain.
Aktin seperti halnya mikrotubula, merupakan filamen polar. Akibat dari polaritas ini terlihat bahwa kecepatan polimerisasi berbeda dari ujung ke ujung. Ujung positif tumbuh lebih cepat daripada ujung negatif. Pertumbuhan aktin murni, 5 sampai 10 kali lebih cepat di daerah positif daripada di daerah negatif. Dengan demikian aktin selalu tumbuh dari daerah positif. Pertumbuhan atau perakitan aktin selalu diawali di daerah selaput plasma terutama selaput sel. Perbedaan kecepatan pertumbuhan ini ditandai dengan banyaknya konsentrasi monomer aktin yang berikatan di kedua ujung filamen (Reksoatmodjo, 1993: 66).
Gerakan sel, seperti fagositosis atau perpindahan sel tergantung pada keseimbangan dinamis antara molekul aktin dan filamen aktin , yaitu terjadinya polimerasi dan depolimerasi pada aktin. Polimerasi sendiri diinduksi oleh ion Mg, ion K, atau Na. Konsentrasi ion yang rendah akan mengakibatkan terjadinya depolimerasi F-aktin menjadi monomer-monomer G-aktin dan akan terpolimerasi secara spontan jika konsetrasi ion kembali tinggi. Tahapan-tahapan polimerasi tersebut adalah sebagai berikut,
1. Monomer aktin G mengikat ATP, dan membentuk kompleks ATP-aktin G
2. Nukleasi. Kompleks ATP-aktin, kemudian membentuk kumpulan kecil yang terdiri dari 3 monomer ATP-aktin.
3. Elongasi. Nukleus yang yang terbentuk kemudian tumbuh memanjang membentuk filamen aktin dengan penambahan monomer pada masing-masing ujung.
Proses pembentukan filamen aktin membutuhkan energi sehingga terjadi proses hidrolisis ATP menjadi ADP. (Cooper & Hausman, 2006: 474-475).
Menurut Karp (2005: 369) selama konsentrasi monomer ATP-aktin masih tinggi di dalam sel, ujung positif akan melanjutkan penambahan monomer (berpolimerisasi). Ketika
dan minus. Pada kondisi ini, terjadi pemisahan monomer (berikatan dengan ADP) dari ujung negatif, dan diseimbangkan dengan penambahan monomer (berikatan dengan ATP) pada ujung positif. Di dalam sel, “actin-binding protein” meregulasi pembentukan dan pemisahan dari filamen aktin (Cooper & Hausman, 2007:476). Menurut Karp (2005: 370) keseimbangan di antara dua aktivitas yang berbeda ini merupakan contoh keadaan yang terus menerus dan terjadi ketika konsentrasi ATP-aktin kira-kira 0,1 μ M.
Pentingnya proses polimerisasi dan depolimerisasi dalam gerakan sel ini ditunjukan dengan menggunakan pengaruh obat-obatan yang mencegah perubahan-perubahan polimerisasi aktin dan karenanya mengganggu gerakan sel seperti perpindahan sel, sitokinesis, dan fagositosis. Akibat pemberian sitokalasin dan obat-obatan sejenisnya adalah hambataan pertumbuhan yang terjadi pada ujung positif filamen aktin.
Senyawa lain yang berpengaruh terhadap mikrofilamen aktin yaitu faloidin. Senyawa ini merupakan alkaloid yang sangat beracun yang dihasilkan oleh Amanita phalloides (cendawan). Berbeda dengan sitokalasin, senyawa ini justru menghambat terjadinya depolimersasi. Demikian pula, senyawa ini tidak mudah melewati selaput sel, oleh karena itu untuk memperoleh hasil yang baik perlu disuntikkan ke dalam sel. Faloidin dapat menghentikan gerakan amoeba sel-sel vertebrata di dalam kultur, sehingga memberi kesan bahwa perakitan dan penguraian mikrofilamen aktin sangat penting bagi gerakan ini (Reksoatmodjo, 1993: 67).
Selain aktin terdapat pula mikrofilamen yang disebut miosin dan tropomiosin yang banyak dijumpai di sel otot. Menurut Subowo (1995: 91), bersama dengan miosin, aktin dapat menyebabkan kontraksi otot melalui mekanisme peluncuran antara kedua jenis protein fibriler tersebut (aktin dan miosin). Miosin sendiri termasuk kategori protein asesori. Filamen aktin yang terdapat dalam sel bukan termasuk otot, paling sedikit mempunyai 2 fungsi, yaitu apabila tersusun bersilang-silang membentuk kerangka sel dan apabila bersama dengan miosin berfungsi dalam sistem kontraktil sehingga bertanggung jawab pada pergerakan sel atau bagian dari sel.
Setiap molekul miosin terdiri dari enam pasang rantai polipeptida, satu rantai berat dan dua pasang rantai ringan yang diatur sedemikian rupa untuk mengahsilkan protein yang asimetris. Molekul miosin terdiri dari:
a) Sepasang kepala bulat yang berisi situs katalik dari molekul
b) Sepasang leher yang masing-masing terdiri dari sebuah α helix yang terputus dan dua rantai ringan
c) Sesuatu yang tunggal, panjang , ekor berbentuk batang yang terbentuk oleh terjalinnya bagian α helix yang panjang dari dua rantai berat.
Struktur molekul miosin II (a) Mikrograf elektron dari molekul miosin. Kedua kepala dan ekor jelas terlihat. (b) Sebuah gambar molekul miosin II yang sangat skematis (massa molekul 52 kDa). Molekul ini terdiri dari sepasang rantai berat (biru) dan dua pasag rantai ringan (Karp, 2005: 271).
Fungsi Mikrofilamen
Pada banyak sel, sitoplasmanya bergerak-gerak dan fenomena ini dinamakan aliran sitoplasmik. Geraknya bergantung apda adanya mikrofilamen. Mikrofilamen ini juga merupakan ciri yang sangat penting dalam sel yang berpindah-pindah dan berubah-ubah bentuknya. Hal ini tidak saja bagi sel yang bergerak bebas seperti amoeba, tetapi juga bagi kebanyakan sel hewan selama pembentukan embrio. Endositosis pada membran sel juga bergantung pada daya kontraktil mikrofilamen.
Filamen aktin yang terdapat dalam sel yang bukan sel otot berfungsi untuk membentuk kerangka sel (tersusun bersilang-silang) dan apabila bersama dengan miosin berfungsi dalam sistem kontraktil sehingga bertanggung jawab pada gerakan sel atau bagian dari sel (Subowo, 1995:91).
Fungsi dari mikrofilamen antara lain sebagai berikut 1. Penyusun Kerangka Sel
Filamen aktin dikelompokkan menjadi dua jenis umum struktur yang disebut bundel aktin dan jaring-jaring aktin, yang memainkan peran yang berbeda dalam sel. Pertama jenis bundel, mengandung filamen aktin spasi berkaitan erat secara paralel mendukung proyeksi membran plasma, seperti mikrovili. Sebuah contoh protein bundling yang terlibat dalam pembentukan struktur adalah fimbrin, yang pertama kali diisolasi dari mikrovili usus dan kemudian ditemukan dalam proyeksi permukaan berbagai jenis sel. di mana filamen dipisahkan banyak spasi, yang memungkinkan bundel untuk berkontraksi. Sedang jaring-jaring aktin ini disebut korteks sel, yang memberi gaya mekanis kepada permukaan sel dan memungkinkan sel dapat bergerak dan berubah bentuk. Di beberapa sel, korteks sel merupakan anyaman trimata yang tebal dari aktin yang berikatan-silang sehingga
membentuk kerangka sel dibawah membran plasma. Umumnya di darah ini tidak ditemui organela dan disebut ektoplasma (Cooper & Hausman, 2007:485).
Tonjolan pada permukaan sel berbasis aktin yaitu mikrovili, dari membran plasma yang terutama berlimpah pada permukaan sel yang terlibat dalam penyerapan, seperti sel-sel epitel yang melapisi usus. Panjangnya sekitar satu mikrometer dengan garis tengah 80 nm, akibatnya luas permukaan penyerapan menjadi 20 kali lebih besar daripada bila tanpa mikrofili. Selaput plasma di daerah mikrofili memiliki selubung ekstraselular yang terdiri dari polisakarida dan enzim-enzim pencernaan. Pada bagian tengah mikrofili terdapat seberkas filamen aktin yang tersusun sejajar satu dengan yang lain. Filamen-filamen tersebut ada di beberapa tempat, dihubungkan antara satu dengan yang lain oleh protein-protein pengikat-aktin yakni fimbrin dan vilin.
2. Sitokinesis Sel Hewan
Mikrofilamen yang terlihat banyak di daerah alur pembelahan sitokinesis pada sel hewan. Tepat di bawah membran plasma di daerah kontriksi terdapat kumpulan mikrofibril kontriksi terdapat kumpulan mikrofibril disebut cincin kontraktil. Sebelum dan sesudah kontriksi mikrofilamen/cincin kontraktril tidak terdapat. Dasar molekular dari kontriksi yang merupakan ciri sitokinesis dalam sel hewan masih masih belum jelas, namun adanya aktin dan miosin menunjukkan bahwa mekanisme kontriksi melibatkan pergeseran filamen seperti pada kontraksi otot. (Siregar, 1990: 173)
3. Kontraksi Sel
Mikrofilamen peranannya dalam pergerakan sel, khususnya sebagai bagian alat kontraksi sel otot. Kontraksi sel otot terjadi akibat filament aktin dan myosin yang saling meluncur melewati yang lain, yang akan memperpendek selnya.
Mikrofilamen berperan sebagai alat kontraksi sel. Sepanjang sel otot ribuan filamen aktin disusun sejajar satu sama lain. Filamen-filamen sejajar tersebut kemudian diselingi dengan filamen yang lebih tebal. Filamen yang lebih tebal inilah yang disebut miosin. Terjadinya kontraksi sel otot akibat filamen aktin dan miosin yang meluncur dan saling melewati satu sama lain. Proses ini akan memperpendek masing-masing sel. Pergerakan kedua protein baik aktin maupun miosin membantu aliran sitoplasma pada sel tumbuhan seperti gerak ameoeboid pada sel protozoa.
4. Viskositas Sel
a. Gerakan Amoeboid
Gerakan amoeboid merupakan gerakan sel bukan sel otot yang mengakibatkan terjadinya perpindahan. Pada dasarnya gerakan amoeboid ini berlandaskan pada perubahan fisik dari sitoplasma yaitu perubahan dari keadaan kental (gel) ke cair (sol) (Issoegianti dan Wibisono, 1993: 80)
Pergerakan amoeboid dibantu otot aktin dan miosin. Aktin berperan dalam pembentukan filamen-filamen. Hal ini berbeda dengan miosin yang tidak membentuk filamen. Ketika aktin bekerja membentuk filamen, lama-kelamaan filamen ini akan menjadi banyak yang kemudian mengubah beberapa bagian sel amoeba dari bentuk sol menjadi gel. Ketika miosin dan filamen bersentuhan maka akan terjadi kontraksi, dan menyebabkan bagian sitoplasma yang berbentuk gel mendorong ke bagian sitoplasma yang berbentuk sol dan maju memasuki pseudopodia. Ketika pseupodia terisi maka akan terjadi gerakan amoeboid (Campbell, 2008).
b. Aliran Sitoplasma
Dalam aliran sitoplasma mikrofilamen memiliki peranan yang penting terutama aktin. Aktinin dan filamin merupakan suatu protein yang dapat mengubah keadaan dari sitoplasma yaitu dari keadaan encer (sol) menjadi kental (gel). Dalam sitoplasma juga terdapat gelsolin dan vilin yang kerjanya berkebalikan dengan aktinin dan filamin. Jadi, aktinin dan filamin mengubah keadaan sitoplasma dari kental ke encer dan dipengaruhi oleh kadar ion Ca2+. Ion
Ca2+ disini memacu terjadinya interaksi anta aktin dan miosin yang mendorong
2. Filamen Intermediet
Filamen intermedia merupakan filamen yang liat dan tahan lama yang terdapat di dalam sel eukariota. Selain itu, filamen intermedia terdiri dari molekul-molekul protein
fibrosa. Filamen ini memiliki daya rentang sangat tinggi. Filamen intermedia berukuran lebih
kecil daripada mikrotubula tetapi lebih besar dari mikrofilamen. Diameternya berkisar antara 8 sampai 10 nm. Filamen intermedia merupakan benang berongga terdiri dari lima buah protofilamen, sejajar satu terhadap yang lain dan membentuk sebuah lingkaran sehingga penampang melintangnya nampak seperti berikut.
Filamen intermedia terdapat di seluruh sitoplasma dengan pusatnya di sekeliling nukleus menyerupai suatu keranjang. Dari sini menjulur ke segala arah menuju daerah perifer. Filamen intermediet memiliki diameter 8-10 nm, yang berada diantara diameter filamen aktin (5-7 nm) dan mikrotubul (25 nm). Merupakan bentuk sitoskeleton yang paling stabil, berfungsi memberikan dukungan struktural bagi sel dan jaringan. Berbeda dengan filamen aktin dan mikrotubul yang merupakan polimer dari protein tunggal (aktin dan tubulin), filamen intermediet tersusun dari berbagai proetin. Lebih dari 50 protein filamen intermediet telah diidentifikasi ke dalam 4 grup berdasarkan kesamaan sekuen asam aminonya. Meskipun berbeda sekuen asam aminonya, filamen intermediet memiliki central rod domain yang terdiri dari sekitar 310-359 asam amino.
Filamen-filamen ini banyak dijumpai dalam sel yang sering mendapat tekanan mekanis, seperti halnya sel epitelium, akson sel saraf atau sel-sel otot polos. Apabila suatu sel diperlakukan dengan detergen yang tidak mengion, filamen-filamen intermedia tetap tidak terlarut. Sesungguhnya istilah sitoskelet, tepat untuk mencandra filamen yang sangat stabil ini. Sifat/tanda Mikrotubula Filamen intermedia Mikrofilamen Struktur Garis tengah (nanometer) Kesatuan monomer Aktivitas ATP-ase Fungsi Berongga dengan dinding terdiri dari 13 protofilamen 24 Tubulin α & β Terletak di dinein a. Kemampuan pergerakan pada eukariota b. Gerakan kromosoma Berongga dengan dinding terdiri 4-5 protofilamen 10 5 jenis protein -Menggabungkan kesatuan kontraktil di dalam sel otot
Mampat (padat) terdiri polimer aktin (Aktin-F) 7 Aktin-G Berperan dalam kontraksi otot perubahan bentuk sek aliran protoplasma
Filamen intermedia diklasifikasikan berdasarkan urutan asam amino penyusunnya. Terdapat empat kelompok filamen intermedia
Macam filamen Polipeptida penyusun Tempat di dalam sel FI tipe I
FI tipe II
FI tipe III FI tipe IV
Keratin bersifat asam Keratin bersifat basa dan netral
Vimentin Desmin
Protein fibrilar yang bersifat asam
Protein-protein penyusun neurofilamen
Protein lamina nukleus
Sel epitelium dan turunan (derivat) epidermis: rambut, kuku
Sel-sel mesenkima Sel-sel dalam kultur Sel-sel otot
Astrosit dan sel Schwan Neuron
Lamina nukleus dari semua sel eukariota
Dari tabel tersebut terlihat bahwa FI (filamen intermedia) tipe I tersusun dari kelompok keratin, subunit keratin asam dan subunit keratin basa atau netral. Filamen keratin merupakan heteropolimer yang terdiri dari kedua subunit tersebut dengan perbandingan 1:1. Keratin ini merupakan protein yang sangat banyak macamnya. Bentuk dan fungsi yang bervariasi dari sel epitelium dapat dibedakan dengan landasan macam keratin yang dibuatnya.
FI tipe II mencakup vimentin, desmin, dan protein fibrilar sel-sel glia. Hampir semua sel derivat sel mesenkima memiliki vimentin, sedangkan desmin terutama dijumpai pada semua sel otot. Protein fibrilar terdapat di sel glia dan sel Schwan.
Protein-protein FI tipe III terakit menjadi neurofilamen yang merupakan penyusun akson dan dendrit. Filamen-filamen ini disebut neurofilamen. Protein FI tipe IV merupakan penyusun lamina nukleus. Protein ini terdiri dari tiga macam lamin yaitu lamin A, B, dan C. Lamin-lamin ini dalam berbagai hal berbeda dari filamen intermedia sitoplasma. Perbedaan yang paling mencolok yaitu bahwa, lamin ini teranyam membentuk suatu lembaran dwimatra. Lamina nukleus merupakan struktur yang dinamis mudah terurai dan terakit kembali. Salah satu contoh yaitu pada stadium tertentu dari mitosis. Semua sel eukariota mensintesis lamin dan biasanya sekurang-kurangnya salah satu di antata keempat tipe filamen intermedia.
daripada mikrofilamen dan mikrotubula, yang sering dibongkar pasang dalam berbagai macam bagian sel. Perlakuan kimiawi yang memindahkan mikrofilamen dan mikrotubula dari sitoplasma meninggalkan jalinan filamen intermediet yang mempertahankan bentuk aslinya. Eksperimen seperti ini menyarankan bahwa filamen intermediet sangat penting dalam memperkuat bentuk sel dan menetapkan posisi organel tertentu (Campbell, 1999). Penyakit degeneratif utama dari kulit, otot dan neuron yang disebabkan oleh gangguan dari filamen intermediet sitoskeleton (Intermediate Filaments.pdf).
3. Mikrotubula
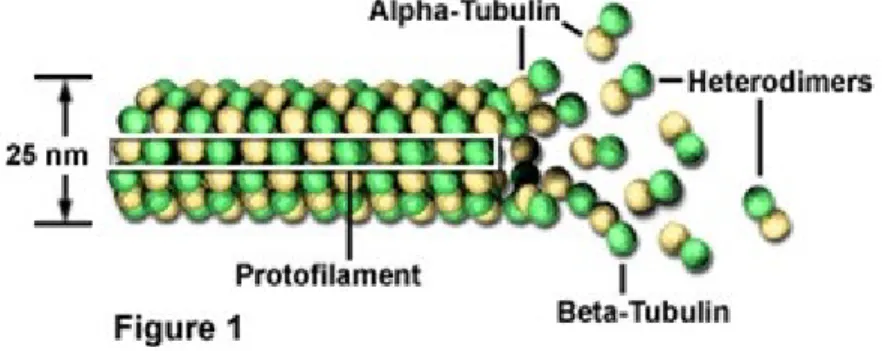
Mikrotubula merupakan salah satu anggota sitoskeleton yang ditemukan dalam sitoplasma semua sel eukariotik. Mikrotubula berupa tabung berongga yang dibangun oleh dua tipe protein, yaitu α tubulin dan β tubulin. Struktur dasar mikrotubul adalah suatu dimer yang terdiri dari α tubulin dan β tubulin. Kedua tubulin ini mengumpul membentuk suatu silinder dengan diameter kira-kira 25 nm dengan diameter lumen atau lubang 15 nm. Kumpulan dimer yang berupa α tubulin dan β tubulin tersusun dalam heliks kemudian tersusun memanjang disebut protofilamen dan membentuk struktur tabung berongga yang disebut mikrotubula (Siregar, 1990: 175).
Gambar struktur helix mikrotubula
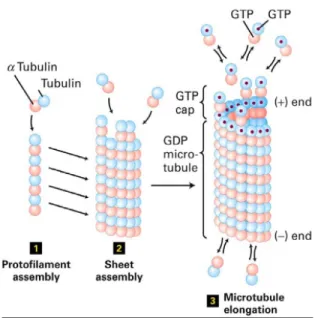
Sebelum molekul-molekul tubulin terakit menjadi mikrotubula, tubulin membentuk protofilamen dengan jalan subunit β dari sebuah molekul tubulin berikatan dengan tubulin subunit α dari molekul tubulin lain yang berada disampingnya (Reksoatmodjo, 1993 : 55)
Gambar pembentukan mikrotubula
Sebuah mikrotubula juga disebut singlet yang terdiri dari 13 protofilamen yang tersusun membentuk suatu lingkaran, pembentukan mikrotubula membutuhkan GTP. Sedangkan 2 buah mikrotubulus disebut dublet, dengan 3 protofilamen ‘milik bersama’ dari kedua mikrotubula. Protofilamen yang dibentuk dari kesatuan dimer tersusun secara paralel. Konsekuensinya, mikrotubula berupas truktur polar dengan 2 ujung yang jelas, yaitu ujung positif dan negatif. Ujung positif merupakan ujung pembentukan yang cepat sedangkan ujung negative merupakan ujung yang lambat pertumbuhannya. Kepolaran ini sangat penting untuk menentukan pergerakan protein motor sepanjang mikrotubulus (Cooper, 2007: 380)
Gambar polarisasi dan depolarisasi pada mikrotubula
Terdapat dua kelompok mikrotubula, yaitu mikrotubula stabil dan mikrotubula labil. Mikrotubula stabil yaitu mikrotubula yang dapat diawetkan dengan larutan fiksatif seperti OsO4, MnO4 atau aldehida pada suhu berapapun. Mikrotubula labil
merupakan mikrotubula yang hanya dapat diawetkan dengan larutan fiksatif aldehida pada suhu sekitar 40 C. Mikrotubula labil dapat dijumpai dalam sitoplasma, tersusun
sejajar satu sama lain seperti yang terdapat dalam aksoplasma sel saraf, dapat pula terlihat terpancar dari satu pusat seperti pada sel yang sedang membelah. Mikrotubula sitoplasma dapat memberikan polaritas kepada sel dan membantu mengatur bentuk sel, gerakan sel dan menentukan bidang pembelahan sel (Reksoatmodjo, 1993 : 59)
Pada mikrotubula labil memiliki ujung positif dan negatif. Ujung positif adalah tempat dimer atau tubulin-tubulin bersatu membentuk heterodimer, sedangkan ujung negatif adalah ujung lepasnya dimer tubulin dari ikatan heterodimer mikrotubula, sehingga strukturnya labil atau bergerak (Sumadi, 2007).
Kegiatan dan fungsi mikrotubula sebagian besar berlandaskan kelabilannya, contoh pada saat pembelahan sel terbentuknya gelendong mitosis dan apparatus mitosis setelah terurainya mikrotubula sitoplasma di awal mitosis.
Mikrotubula gelendong mitosis umumnya sangat labil, cepat terakit maupun terurai. Hal ini yang mengakibatkan gelendong mitosis dengan obat-obatan. Salah satunya adalah kolkisin, yang akan terikat kuat pada tubulin bebas dan mencegah terbentuknya mikrotubula. Akibatnya sel yang sedang mengalami pembelahan akan menghentikan aktivitas mitosis dan menghilangnya gelendong mitosis selama beberapa menit. Senyawa sepeti kolkisin yang mampu menghambat proses pembelahan mitosis disebut sebagai senyawa antimitosis. Pengaruh dari zat ini bersifat timbal balik, sehingga apabila obat-obatan dihilangkan, gelendong mitosis akan terbentuk kembali dan pembelahan kembali berlanjut (Reksoatmodjo, 1993).
Beberapa senyawa pengikat tubulin (senyawa antimitosis)
Senyawa Pengaruh Terhadap Mikroba
Kolkisine, Kolesmid, Nokadazole Menghambat penambahan molekul tubulin ke mikrotubula, menyebabkan depolimerisas imikrotubula
parakristalin dari tubulin, sehingga terjadi depolimerisasi
Taksol Memacu perakitan mikrotubula,
menstabilkan mikrotubula (Reksoatmodjo, 1993 :59)
Tubulin dimer dapat mengalami laju depolimerisasi sama seperti polimerisasi, pada mikrotubula dapat terjadi proses perakitan sekaligus pembongkaran tubulin α dan tubulin β yang mengadakan ikatan dengan GTP yang fungsinya analog dengan pengikatan ATP pada mikrofilamen. GTP yang terikat denganβ tubulin dihidrolisis membentuk GDP selama polimerisasi. Hidrolisis GTP ini melemahkan ikatan afinitas dari tubulin yang berdekatan. Proses ini akan mengakibatkan depolimerisasi dan menimbulkan ketidakstabilan dinamis pada mikrotubulus. Pada proses pengikatan GTP yang tinggi maka polimerisasi akan berlangsung cepat, selama terdapat tubulin bebas yang cukup banyak, namun apabila jumlah monomer yang bebas lebih sedikit dari monomer yang terikat (kondisikritis) hidrolisis GTP akan memacu proses depolimerisasi tubulin pada ujung (-), laju pembentukan dan pembongkaran sama, proses ini disebut treadmilling.
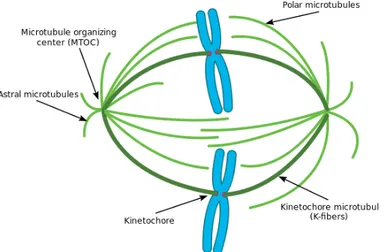
Mikrotubula sitoplasma dapat dilihat pada stadium interfase, dengan teknik immunofluoresensi. Mikrotubula terlihat paling banyak di dekat inti, di daerah ini terpancar bentuk anyaman benang halus ke arah perifer. Untuk mengetahui asal dari mikrotubula, maka dilakukan depolimerisasi kemudian dibiakkan kembali. Awalnya pembentukan mikrotubula diawali dengan adanya titik kecil yang berbentuk bintang sehingga disebut aster yang terletak di dekat inti. Pancaran benang-benang halus memanjang ke arah tepi sel sampai penyebaran awal terbentuk kembali. Daerah pembentukan aster disebut MTOC (Microtubule Organizing Center). Kutub negative mikrotubula berada di daerah MTOC sedangkan kutub positifnya menjauhi MTOC. MTOC merupakan pusat pertumbuhan mikrotubula, berarti tempat polimerisasi tubulin terbentuk berada pada bagian prosimal (menjauhi MTOC) yaitu kutub + (Choirul: 258-259).
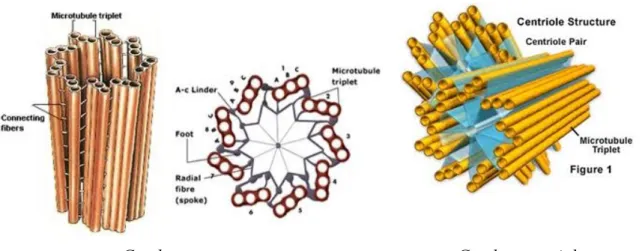
Gambarletak MTOC
Sebagian besar sel hewan memiliki MTOC utama yang disebut pusat sel atau sentrosoma yang terletak pada salah satu sisi inti dan terdapat sepasang sentriola yang tersusun tegak lurus satu terhadap yang lain. Tiap sentrosoma tersusun atas 9 pasang triplet mikrotubula yang tersusun ke dalam suatu cincin. Apabila sel membelah, sentriol ini bereplikasi. Pada tumbuhan MTOC tidak memiliki sentriola, mikrotubula aster muncul dari sentrosoma yang hanya terdiri dari materi padat elektron. Begitu pula pada oositmencit, sentriola tidak dijumpai pada gelendong mitosis (Reksoatmodjo, 1993).
Mikrotubula sitoplasma pada sel hewan cenderung memancar ke segala arah dari sentrosoma. Proses perakitan mikrotubula dipantau sedemikian rupa sehingga mikrotubula yang terbentuk menjulur ke arah tertentu dari sel. Pemasakan mikrotubula ditentukan sebagian oleh modifikasi pascatranslasi dari molekul tubulin dan sebagian lagi oleh interaksi antara mikrotubula dengan protein khusus pengikat mikrotubula. Modifikasi pascatranslasi dari tubulin menunjukkan bahwa mikrotubula telah mantap, namun modifikasi mikrotubula tersepat diduga karena adanya protein yang lain, yang disebut MAPs (Microtubule associated Proteins). Protein ini berfungsi untuk merintangi penguraian mikrotubula dan memacu terjadinya interaksi antara mikrotubula dan komponen sel lainnya.
fosfat dari molekul energi yaitu ATP. Protein motor mampu merubah energy kimiawi menjadi energi mekanik sehingga proses pergerakan organela atau protein dapat berlangsung. Untuk memenuhi energi yang dibutuhkan ,terdapat dua proses yang berkelanjutan untuk membantu protein motor mampu menjalankan fungsinya. Kedua siklus tersebut adalah siklus kimiawi dan mekanik. Tahap proses kimiawi dimulai dengan terjadinya ikatan ATP pada protein motor yang disusul dengan hidrolisis ATP, melepaskan ADP dan Pi dari motor serta terikatnya kembali satu molekul ATP baru ke protein motor. Hidrolisis ATP akan menyediakan energi yang diperlukan untuk pergerakan protein motor tersebut (Rudijanto, 2010).
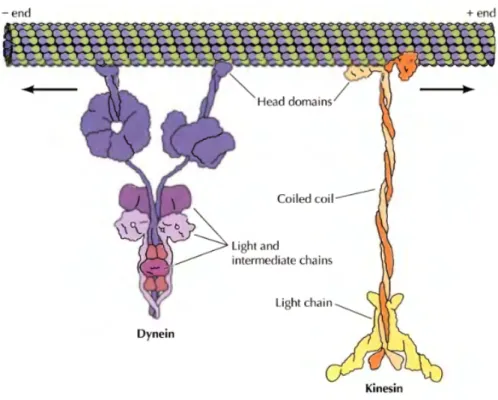
Keduaprotein motor tersebut memiliki arah gerakan yang berbeda. Dinein bergerak dari kutub positif ke negative sedangan kinesin dari arah sebaliknya, kutub negative ke kutub positif. Kinesin berperan dalam proses penarikan sentromer ke arah kutub oleh benang spindel yang tidak lain merupakan mikrotubulus. Pada transpor vesikel, bagian kepala merupakan bagian yang mengikat mikrotubula, sedangkan bagian ekor yang mengikat vesikel. Beberapa fungsi lain dari 2 protein motor ini akan dijelaskan dalam bagian selanjutnya (Cooper, 2007).
Gambar dinein dan kinein
Fungsi mikrotubula :
1. Memberi bentuk sel, beberapa mikrotubula menyusun jala serat yang menyebar dari sentromer (MTOC, pusat pengorganisasian mikrotubula).
mikrotubula berujung di dua polaritas. Ujung positif (+) berdekatan melekat pada sentromer kromosom dan ujung negatif berdekatan dengan sentriol. Jadi, pada ujung sentriola mikrotubula diperpendek untuk menggerakkan kromosom menuju ke kutub pembelahan (Reksoatmodjo, 1993).
Gambarbenangspindelpadapembelahansel
Mikrotubula dapat dirakit, diurai, dan dirakit kembali, pada saat mitosis, di awal metaphase, timbul gelendong mitosis. Gelendong ini terdiri dari dua jenis serabut fungsional, yang dua-duanya adalah mikrotubula. Jenis yang pertama terpancar dari kinetokor yang terdapat di kromosom ke kutub selatan sentriola. Jenis yang lain terentang dari sentriol kesentriola. Mikrotubula penyusun gelendong mitosis ini dirakit tepat sebelum mitosis dan diurai setelah kromosom terpisah. (Wibisono, 1993:147).
Gambar penarikan kinetokor
3. Transpor Vesikel
Pada gerak vesikula badan golgi ke plasma membran, mikrotubula dibantu oleh protein motor kinesin. Antara kinesin dan vesikula terdapat reseptor kinesin. Gerakan
vesikula seolah-olah dibawa oleh kinesin yang berjalan di atas mikrotubula stasioner dengan bantuan energi ATP. Arah gerakan dari kutub negatif ke kutub positif pada mikrotubula.
Gambarpergerakankinesin
4. Untuk medium transport aksonal pada sel saraf terjadi di sepanjang mikrotubula
Sintesis protein hanya terjadi pada badan sel dan dendrit sel saraf, maka bagian akson dan terminal sinaptik sel saraf tidak mengandung kegiatan sintesis protein. Oleh karena itu, protein dan organel bermembran harus disintesis pada badan sel dan ditransportasikan ke akson ke arah ujung sinapsis. Transpor material sepanjang akson ke ujung sinapsis ini dinamakan transpor aksonal.
Menurut Karp (2005: 356-359), struktur dasar cillia dan flagellata adalah sama.
Gambar pembelahan cilia dan flagellata secara vertikal
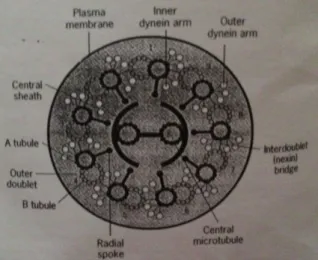
Inti dari cilium disebut sebagai anoxeme yang mengandung kumpulan mikrotubulus yang berjajar secara longitudinal menembus seluruh organel. Dengan pengecualian yang jarang, anoxeme daricillium atau flagellate yang bergerak secara cepat mengandung sembilan doublet mikrotubulus yang periferal mengelilingi sepasang mikrotubulus single yang berada di tengah. Struktur mikrotubulus ini dikenal sebagai “susunan 9 + 2”. Seluruh anoxeme mikrotubulus memiliki kepolaran (muatan kutub) yang sama, bagian positif ada pada ujung atas dan bagian negatif ada pada ujung dasar. Setiap doublet periferal mengandung satu microtubulus yang lengkap.
Mekanisme pergerakan cilia dan flagellata
Karp (2005: 360-363) menyatakan bahwa mesin penggerak dari cilia dan flagellate terletak dalam anoxeme. Dalam bagian supernatant dari anoxeme terdapat ATPase yang mana memiliki kemampuan untuk mengikat ATP, bagian ini dinamakan dinein (dyne = kekuatan). Mekanisme pergerakan ini dipengaruhi oleh kontraksi dari otot yang disebut sliding. Kekuatan sliding ini seperti roda bergigi searah yang terletak di bagian kepala dari myosin. Dengan system otot sebagai model, bisa dibayangkan bahwa mekanisme sliding dari doublet mikrotubul relatif dari satu kelainnya. Dalam hal ini, tangan-tangan dinein berayun dan gerakan ayunan ini dibutuhkan oleh cilia dan flagellate. Bagian akar dari molekul dinein terikat dengan kuat pada permukaan luar tubulus A dengan kepala globular dan batangnya pada tubulus B. tangan-tangan
B.Molekul dinein kemudian melalui gerakan yang menyebabkan bagian bawah doublet meluncur ke ujung dasar dari doublet atas. Kemudian tangan-tangan ini terikat kembali ke bagian atas doublet sehingga siklus berikutnya bisa berjalan.
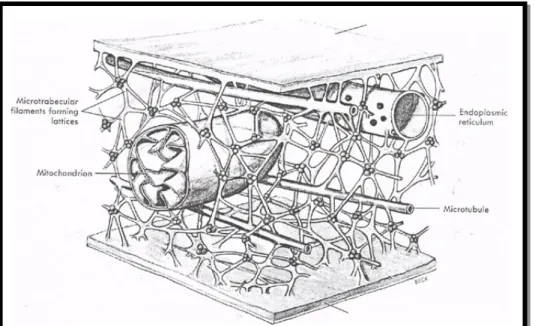
4 Mikrotrabekula
Serabut yang terdapat pada sitoplasma tersusun dari mikrotubula, mikrofilamen, dan filamen menengah. Di samping itu, terdapat filamen lain yang turut membangun kisi-kisi ini. Filamen yang terakhir ini merupakan filamen halus dan tipis dengan dimensi: panjang 30-300 nanometer dengan garis tengah 2-3 nanometer. Filamen ini disebut filamen mikrotrabekula. Mikrotrabekula berperan dalam menghubungkan serabut-serabut yang ada di sitoplasma dan dikaitkan ke
organel sel. Kisi-kisi mikrotrabekula bersama-sama dengan filamen-filamaen sitoplasma dan mikrotubul berperan dalam memelihara bentuk dan gerakan sel. (Siregar, 1990: 170)
Ruang antar trabekula itu berair dan berperan melarutkan atau mensuspensikan berbagai molekul kecil yang terlibat dalam metabolisme sel (misal glukosa, asam amino, oksigen, dan garam organik). (Siregar, 1990: 168)
Mikrotrabekula ini menyangga semua organela supaya tetap di tempatnya dan menambatkan satu dengan yang lain, penyusun sistem serabut di dalam sel serta mengaitkannya dengan korteks sel. (Issoegianti dan Wibisono, 1993: 153)
Daftar Pustaka
Alberts B, et.al.. 2002. Molecular Biology of the Cell. Ney York: Garland Science diakses dari id.wikipedia.org
Campbell, Neil A. 1999. Biologi Jilid 1 Edisi Kelima. Jakarta: Erlangga Campbell, Neil A. 2001. Biologi Jilid 2 Edisi 5. Jakarta: Erlangga
Campbell, Neil A, & Reece, Jane B. 2008. Biologi 1 Ed. 8. Jakarta: Erlangga
Cooper, G.M. & R.E. Hausman . 2007. The Cell: A Molecular Approach. 4th ed. Sunderland: Sinauer Associates, Inc.
Cooper, Geoffrey M. Dan Robert E. Haussman. 2006. The Cell a Mollecular Approach 4th
Edition. New York: ASM Press
Cooper, M Geoffrey. 2007. The Cell A molecular Approach 4th Edition. Washington : ASM Press.
Intermediate filaments.pdf diakses pada 24 November 2014 11:39.
Issoegianti dan Wibisono. 1993. Persiapan Perkuliahan Program Lanjutan MIPA Lembaga
Pendidikan Tenaga Kependidikan (Program B) Bidang Biologi Sel. Yogyakarta: Fakultas
Biologi Universitas Gadjah Mada.
Karp, Gerald. 2005. Cell an Moleculer Biology: Concept and Experiments 4th Edition. USA:
Macmillan Magazines Ltd.
Lodish H, Berk A, Zipursky SL, et al. 2000. Molecular Cell Biology 4th Edition. New York: W.
H. Freeman
Lodish H, Berk A, Zipursky SL, et al. 2000. The Dynamic of Actin Assembly. Diunduh pada tanggal 27 November 2014 dari
http://www.ncbi.nlm.nih.gov/books/NBK21594/figure/A5147/?report=objectonly Reksoatmodjo, Issoegiyanti. 1993. Biologi Sel. Yogyakarta :UniversitasGadjahMada.