STUDI PENGARUH MUTASI GEN rpoB PADA KODON 513:
ANALISIS PADA ISOLAT PAPUA
Richardo Ubyaan1,*)Agnes E. Maryuni,2)dan Alvian Sroyer3) 1)Program Studi Kimia, FKIP, Universitas Cenderawasih, Jayapura, Papua
2)Jurusan Kimia, FMIPA, Universitas Cenderawasih 3)Jurusan Matematika, FMIPA, Universitas Cenderawasih
Disajikan 29-30 Nop 2012
ABSTRAK
Resistensi terhadap RIF disebabkan oleh mutasi pada gen rpoB pengkode RNA polimerase (RNAP) subunit β, terutama pada daerah sepanjang 81 pasang basa (pb) sebagai penentu sifat resistensi RIF, dengan frekuensi yang paling tinggi pada kodon 526 dan 531. Mutasi ini menyebabkan RIF tidak berfungsi dalam menghambat proses inisiasi transkripsi. Pada salah satu galur isolat klinis M. tuberculosis MDR asal Jayapura, Papua yang ada di kelompok penelitian M. tuberculosis Laboratorium Biokimia, Universitas Cenderawasih, terdapat isolat yang memiliki mutasi pada kodon 315 gen katG penyebab resistensi isoniazid, tetapi tidak memiliki mutasi pada kodon rpoB526 dan rpoB531. Fenotipe resistensi terhadap RIF yang dimilikinya diduga disebabkan oleh adanya mutasi pada posisi selain kedua kodon tersebut. Pada penelitian ini dilakukan penentuan penyebab resistensi RIF tingkat genotipe pada isolat klinis M. tuberculosis MDR dan juga dicari penjelasan mengenai hubungan mutasi dengan sifat resistensi tersebut. Tahapan penelitian meliputi PCR multipleks spesifik alel rpoB dan elektroforesis, penentuan urutan nukleotida, dan analisis in silico. Hasil sekuensing dianalisis secara in silico, yaitu disejajarkan dengan urutan nukleotida galur standar M. tuberculosis H37Rv. Selain itu juga, dilakukan pemodelan protein menggunakan program PyMOL versi open source untuk melihat pengaruh mutasi pada interaksi RIF dengan RNAP. Hasil analisis translasi in silico menunjukkan bahwa kodon CAA yang mengkode asam amino glutamin termutasi menjadi CTA yang mengkode leusin. Hasil pemodelan protein menggunakan aplikasi tersebut menunjukkan bahwa perubahan Gln513Leu merubah jarak antara rantai samping residu tersebut dengan gugus hidroksil RIF dari 2,63 Å menjadi 3,71 Å. Hasil sekuensing dan penjajaran di atas menunjukkan adanya mutasi pada posisi lain gen rpoB isolat Papua 1, yaitu pada kodon 513. Mutasi ini diduga kuat merupakan penyebab resistensi RIF. Mutasi yang ada telah merubah residu tersebut menjadi Leu yang memiliki rantai samping non-polar dan membuat jarak rantai samping residu tersebut dengan gugus hidroksil RIF menjadi lebih jauh. Hal ini dapat menyebabkan ikatan hidrogen tidak dapat terbentuk, sehingga mengurangi afinitas pengikatan RIF pada RNAP. Diharapkan pengetahuan tentang mekanisme resistensi ini dapat digunakan sebagai dasar desain obat baru untuk mengatasi masalah MDR-TB.
Kata Kunci: Mtb, mutasi, rpoB, rifampin, isolat Papua
I. PENDAHULUAN
Penyakit tuberkulosis (TB) merupakan suatu penyakit infeksi menular pada manusia yang disebabkan oleh bakteri
Mycobacterium tuberculosis. Sampai saat ini, terdapat banyak
obat anti-TB (OAT) yang merupakan antibiotik seperti rifampin, isoniazid, pirazinamid, etambutol, streptomisin, senyawa-senyawa golongan fluorokuinolon, dan lain-lain. Akan tetapi, walaupun sudah terdapat banyak OAT, TB tetap merupakan penyakit yang sulit di atasi. Hal ini terutama diakibatkan oleh sifat resistensi yang dimiliki TB terhadap antibiotik. Resistensi TB terbagi menjadi dua macam, yaitu resistensi terhadap satu jenis antibiotik, dan resistensi terhadap lebih dari satu jenis antibiotik. WHO telah mendefinisikan TB yang resisten terhadap paling tidak dua jenis antibiotik sekaligus yaitu rifampin (RIF) dan
isoniazid (INH) sebagai multidrug-resistant TB (MDR-TB). Tentunya MDR-TB disebabkan oleh galur M. tuberculosis yang memiliki sifat tersebut. Munculnya kasus MDR-TB merupakan suatu masalah global yang harus di atasi untuk memberantas penyakit TB.
Resistensi M. tuberculosis terhadap antibiotik diakibatkan adanya mutasi pada kromosom bakteri tersebut. Hal ini menyebabkan sensitivitas M. tuberculosis terhadap OAT berkurang. Mutasi ini terjadi pada gen yang mengkode target antibiotik atau gen yang berperan dalam interaksi antibiotik dengan targetnya pada M. tuberculosis. Resistensi terhadap INH sebagian besar terjadi akibat mutasi pada gen
katG pengkode katalase-peroksidase yang berperan dalam
mengubah INH menjadi bentuk aktifnya di dalam sel [1-2]. Resistensi terhadap RIF terjadi akibat mutasi pada gen rpoB
▸ Baca selengkapnya: lkpd mutasi gen
(2)pengkode RNA polimerase (RNAP) subunit β, yang menyebabkan RIF tidak dapat menjalankan fungsinya dalam menghambat inisiasi transkripsi [1-3].
Mutasi utama penyebab resistensi INH pada gen katG adalah mutasi pada kodon 315, sedangkan mutasi utama penyebab resistensi RIF terletak pada daerah sepanjang 81 pasang basa (pb) pada gen rpoB, yang disebut daerah penentu sifat resistensi RIF, yaitu kodon 507-533, dengan frekuensi mutasi paling tinggi pada kodon 526 dan 531 [1-2]. Adapun sistem penomoran kodon tersebut menggunakan nomor kodon rpoB Escherichia coli, bukan nomor kodon M.
tuberculosis sebenarnya [3]. Mutasi penyebab kedua jenis
resistensi di atas telah dapat dideteksi secara sederhana dan cepat menggunakan metode Polymerase Chain Reaction (PCR) multipleks spesifik alel [4].
Pada koleksi 20 galur isolat klinis M. tuberculosis MDR Papua, Indonesia, yang ada di kelompok penelitian M.
tuberculosis Biokimia, FMIPA, Universitas Cenderawasih,
terdapat isolat yang melalui uji genotipe menggunakan PCR multipleks diketahui termutasi pada kodon katG315 tetapi tidak memiliki mutasi pada kodon rpoB526 dan rpoB531. Oleh karena itu, fenotipe resistensi RIF yang dimilikinya haruslah disebabkan oleh faktor lain. Diduga sifat itu diakibatkan oleh adanya mutasi pada posisi selain kedua kodon di atas [5].
Berdasarkan permasalahan yang ada, penelitian ini dilakukan untuk menentukan penyebab resistensi RIF tingkat genotipe pada isolat klinis M. tuberculosis MDR di atas, selain mutasi pada kodon rpoB526 dan rpoB531. Selain itu juga dicari penjelasan mengenai hubungan mutasi dengan sifat resistensi RIF yang dimilikinya.
II. METODOLOGI
Isolat klinis M. tuberculosis MDR yang digunakan pada penelitian ini adalah isolat P1, P2, P3, dan P4, yang berasal dari Laboratorium Biokimia, Universitas Cenderawasih, Jayapura, Papua. Metode yang digunakan merupakan modifikasi metode pengujian PCR multipleks spesifik alel
rpoB untuk deteksi M. tuberculosis resisten RIF (RIFr) yang
dilakukan oleh Mokrousov dan koleganya [4].
Pada penelitian ini pertama kali dilakukan konfirmasi ulang PCR multipleks rpoB526 dan rpoB531 di atas yang telah dilakukan pada keempat isolat tersebut. Untuk tiap jenis PCR multipleks digunakan sepasang primer-luar, yaitu primer forward RF (5’-GTCGCCGCGATCAAGGA) dan primer reverse RR (5’-TGACCCGCGCGTACAC), dan satu primer-dalam R526 (5’-GTCGGGGTTGACCCA) atau R531 (5’-ACAAGCGCCGACTGTC). Templat DNA berupa DNA hasil lisis M. tuberculosis isolat klinis Papua (5 μL) ditambahkan pada campuran reaksi PCR (volume akhir 20 μL untuk PCR-rpoB526 dan 15 μL untuk PCR-rpoB531) yang mengandung bufer PCR 10x tanpa MgCl2 (2,5 μL untuk PCR-rpoB526 dan 2 μL untuk PCR-rpoB531), MgCl2 (3 mM
untuk PCR-rpoB526 dan 4 mM untuk PCRrpoB531), 0,5 U
Taq DNA polimerase, 200 μM masing-masing dNTP, 1 pmol
primer-luar RF, 20 pmol primer-luar RR, dan salah satu primer-dalam spesifik alel R526 (30 pmol) atau R531 (35 pmol). Keempat primer yang digunakan disintesis oleh Biogen. Sebagai kontrol positif digunakan campuran reaksi PCR yang sama, dengan templat DNA berupa hasil lisis galur standar M. tuberculosis H37Rv, sedangkan kontrol negatif tidak mengandung templat DNA (diganti dengan ddH2O dengan volume yang sama).
Kedua reaksi PCR dilakukan pada alat GeneAmp®PCR
System 2700, Perkin Elmer, dengan kondisi sebagai berikut: denaturasi awal pada 96°C selama 3 menit; 5 siklus yang terdiri atas 95°C selama 45 detik, 60°C selama 1 menit, dan 72°C selama 30 detik; 5 siklus yang terdiri atas 95°C selama 40 detik, 59°C selama 50 detik, dan 72°C selama 30 detik; 25 siklus yang terdiri atas 94°C selama 50 detik, 55°C selama 40 detik, dan 70°C selama 30 detik; dan elongasi akhir pada 72°C selama 3 menit. Hasil PCR kemudian dianalisis dengan elektroforesis gel agarosa 1,5% b/v (0,6 g agarosa dalam 40 mL bufer TAE 1X yang mengandung 2 μL EtBr 10 mg/mL). Sampel hasil PCR (5 μL) ditambah dengan 2 μL loading buffer dan dielektroforesis dengan running buffer TAE 1X, pada 75 Volt selama 50 menit. Hasil elektroforesis kemudian divisualisasikan dengan bantuan sinar UV. Sebagai DNA standar (marker) digunakan pUC19/HinfI yang mengandung fragmen-fragmen DNA berukuran 1419 pb, 517 pb, 396 pb, 214 pb, 75 pb, dan 65 pb.
Untuk mengkonfirmasi hasil PCR multipleks di atas, dilakukan penentuan urutan nukleotida (sekuensing) fragmen 249 pb yang mengandung daerah penentu sifat resistensi RIF. Keempat isolat klinis di atas masing-masing terlebih dahulu diamplifikasi menggunakan sepasang primer-luar saja (RF dan RR), dengan kondisi reaksi PCR yang sama dengan PCR multipleks, sehingga dihasilkan fragmen berukuran 249 pb, dengan kuantitas sekitar 800 ng. Hal yang sama juga dilakukan pada galur M. tuberculosis H37Rv, yang kemudian akan digunakan sebagai standar. Fragmen 249 pb hasil PCR keempat isolat klinis dan standar dikirim ke Macrogen Inc., Seoul, Korea Selatan, untuk ditentukan urutan nukleotidanya menggunakan primer RR sebagai primer sekuensing.
Analisis in silico
Analisis in silico pertama yang dilakukan adalah penjajaran (alignment) sekuen isolat klinis dan sekuens galur standar M. tuberculosis H37Rv menggunakan program DNASTAR. Analisis in silico yang dilakukan berikutnya adalah pemodelan protein menggunakan program PyMOL versi Open-Source. Sebagai model interaksi RNAP-RIF digunakan struktur kristal kompleks RNAP inti (core) T.
aquaticus (Taq) dan RIF dengan kode ID Protein Data Bank
tuberculosis, dan E. coli, urutan nukleotida daerah-daerah
yang mengandung mutasi penyebab RIFr pada ketiga organisme tersebut identik sebesar 91% [6]. Pada pemodelan ini, akan dilakukan perubahan asam amino pada residu yang termutasi.
III.
HASIL DAN PEMBAHASAN
A. PCR multipleks Spesifik Alel rpoB
PCR multipleks ditujukan untuk mendeteksi mutasi pada kodon rpoB526 dan rpoB531. Isolat P1 menghasilkan dua pita DNA berukuran 0,25 kb dan 0,18 kb untuk PCR-rpoB526 dan 0,25 kb dan 0,17 kb untuk PCR-rpoB531. Sementara isolat lainnya, P2, P3 dan P4, juga memberi hasil dua pita DNA berukuran 0,25 kb dan 0,18 kb untuk
PCR-rpoB526, tetapi hanya memberikan hasil satu pita DNA
berukuran 0,25 kb untuk PCR-rpoB531.
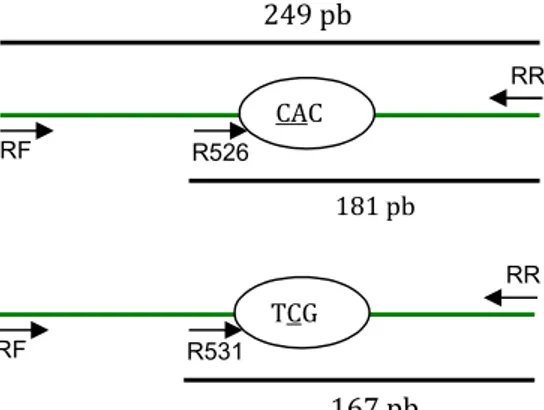
Hasil PCR tersebut divalidasi dengan kontrol positif dan kontrol negatif yang digunakan. Kedua kontrol berjalan dengan baik, ditunjukkan dengan tidak adanya pita DNA pada hasil PCR kontrol negatif dan adanya dua pita DNA berukuran 0,25 dan 0,18 kb atau 0,25 dan 0,17 kb (sesuai jenis PCR multipleks yang dilakukan) pada hasil PCR kontrol positif. Makna terbentuknya satu atau dua pita pada hasil PCR multipleks dan elektroforesis di atas dapat dijelaskan dengan melihat skema target PCR multipleks (Gambar 1). Primer-luar RF dan RR akan mengamplifikasi fragmen tetap 249 pb, baik pada PCR-rpoB526 maupun PCR-rpoB531. Primer dalam (R526 dan R531), masing-masing telah dirancang agar ujung 3’-nya menempel pada basa kedua kodon rpoB526 dan rpoB531 alel wild type [4].
Gambar 1. Skema target PCR multipleks spesifik alel rpoB526 dan rpoB531. (A) PCR-rpoB526. (B) PCR-rpoB531. Panah pendek merepresentasikan primer, panah panjang merepresentasikan fragmen PCR yang tetap (249 pb) dan fragmen spesifik-alel wild type (181 dan 167 pb). Jika kodon 526 dan 531 termutasi, hanya dihasilkan fragmen 249 pb. (Catatan: gambar tidak menggunakan skala.)
Pita DNA hasil PCR multipleks dan elektroforesis yang berukuran 0,25 kb menunjukkan terbentuknya fragmen tetap 249 pb, sedangkan pita berukuran 0,18 dan 0,17 kb berturut-turut menunjukkan terbentuknya fragmen spesifik-alel wild
type 181 pb (PCR-rpoB526) dan 167 pb (PCR-rpoB531). Oleh
karena itu, hasil PCR multipleks berupa dua pita DNA 0,25 dan 0,18 kb atau 0,25 dan 0,17 kb, berturut-turut menunjukkan bahwa kodon rpoB526 atau rpoB531 merupakan alel wild type. Sedangkan hasil PCR multipleks berupa satu pita DNA saja (0,25 kb) menunjukkan adanya mutasi pada kodon yang bersangkutan.
Rangkuman pada data berikut menunjukkan bahwa keempat isolat memiliki kodon rpoB526 alel wild type (tidak termutasi) dan hanya isolat P1 yang memiliki kodon rpoB531 alel wild type, sedangkan pada tiga isolat lainnya (P2, P3, dan P4) kodon tersebut termutasi. Atas dasar data hasil PCR multipleks keempat isolat tersebut, hanya isolat P1 yang
terkonfirmasi tidak memiliki mutasi-mutasi utama penyebab resistensi RIF, sedangkan isolat P2, P3, dan P4 ternyata memiliki mutasi pada kodon rpoB531 yang merupakan penyebab utama resistensi RIF [1-2].
Dengan tidak termutasinya kodon rpoB526 dan rpoB531 pada isolat P1, maka resistensi RIF yang dimiliki isolat tersebut haruslah disebabkan oleh faktor yang lain, misalnya mutasi gen rpoB pada posisi selain kedua kodon di atas. Untuk mengetahuinya, telah dilakukan sekuensing fragmen 249 pb gen rpoB isolat P1, yang mengandung daerah penentu resistensi RIF. Sekuensing tersebut juga dilakukan pada tiga isolat lainnya untuk mengkonfirmasi mutasi rpoB531 di atas.
B. Analisis Penjajaran (Alignment)
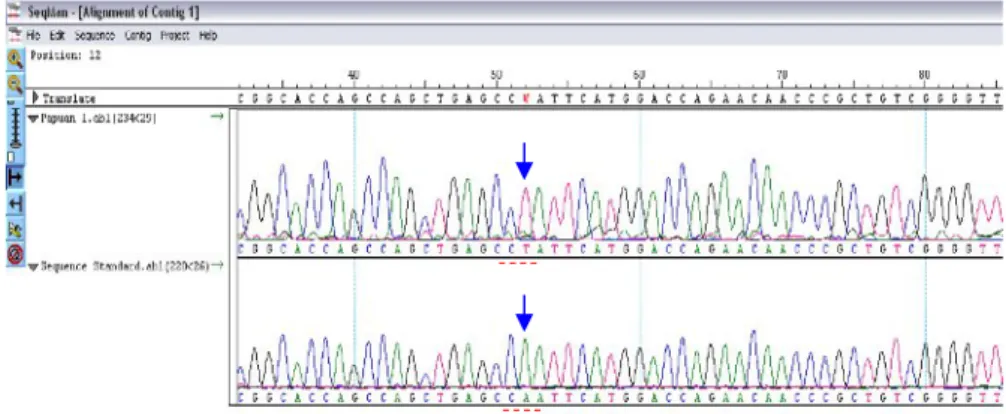
Program SeqManTMdapat menjajarkan urutan nukleotida
dengan menampilkan data elektroforegram. Penjajaran komplemen sekuens isolat P1 (P1-RR) dan sekuens H37Rv
249 pb CAC RF R526 RR 181 pb TCG RF R531 RR 167 pb
(H37R-RR) diperlihatkan pada Gambar berikut secara berurutan (Gambar 2). Hasil penjajaran menunjukkan adanya satu mutasi subtitusi adenin (A) menjadi timin (T), yang merupakan nukleotida nomor 1295 gen rpoB atau
nukleotida nomor 761101 genom M. tuberculosis H37Rv. Nukleotida pada posisi ini merupakan basa kedua kodon
rpoB513, sehingga CAA termutasi menjadi CTA.
Gambar 2. Penjajaran (alignment) SeqManTMDNASTAR. Tampak pada gambar komplemen urutan nukleotida isolat P1 (P1-RR)
disejajarkan dengan komplemen urutan nukleotida galur standar M. tuberculosis H37Rv.
Hasil sekuensing dan penjajaran yang telah dilakukan mengkonfirmasi data hasil PCR multipleks spesifik alel
rpoB526 dan rpoB531. Data yang diperoleh menunjukkan
konsistensi dengan hasil PCR multipleks tersebut, yaitu bahwa pada isolat P1, kodon rpoB526 dan rpoB531 adalah alel wild type, sementara pada isolat P2, P3, dan P4, kodon
rpoB526 merupakan alel wild type sedangkan kodon rpoB531
termutasi. Mutasi pada kodon rpoB531 menjelaskan sifat RIFr pada ketiga isolat tersebut.
Selain dengan SeqManTM, penjajaran sekuens isolat P1
(P1-RR) dengan sekuens standar H37Rv (H37R-RR) juga dilakukan dengan MegAlignTM DNASTAR. Sebelumnya,
urutan nukleotida isolat klinis dan hasil sekuensing disimpan dalam bentuk file EditSeqTMterlebih dahulu (file
dengan ektensi SEQ). Hasil penjajaran MegAlignTM
menunjukkan mutasi yang sama pada isolat P1, yaitu A1295T. Menggunakan program ini, dapat dilakukan secara
langsung penjajaran urutan asam amino hasil translasi urutan nukleotida yang telah disejajarkan sebelumnya. Hasil penjajaran urutan asam amino hasil translasi sekuens isolat P1 dan standar H37Rv menunjukkan mutasi Q (glutamin, Gln) menjadi L (leusin, L). Perubahan asam amino ini terletak pada kodon rpoB513, dihasilkan dari mutasi A1295T yang telah disebutkan sebelumnya, yang merubah CAA yang mengkode Gln menjadi CTA yang mengkode Leu. Analisis penjajaran yang telah dilakukan pada isolat M.
tuberculosis MDR P1, baik menggunakan program SeqManTM
maupun MegAlignTMDNASTAR telah menunjukkan adanya
mutasi pada basa kedua kodon rpoB513, yaitu A1295T. Pada galur standar M. tuberculosis H37Rv, kodon rpoB513 memiliki urutan nukleotida CAA yang mengkode Gln, sedangkan pada isolat P1, kodon ini termutasi menjadi CTA yang mengkode asam amino Leu (Gambar 3).
Gambar 3. Penjajaran (alignment) MegAlignTMDNASTAR. Tampak pada gambar penjajaran urutan asam amino isolat P1 dengan
urutan asam amino galur standar M. tuberculosis H37Rv. Posisi yang ditandai panah menunjukkan mutasi Q (glutamin) pada H37Rv menjadi L (leusin) pada isolat P1.
Apabila mutasi kodon 513 tersebut merupakan satu-satunya mutasi pada gen rpoB isolat M. tuberculosis MDR P1, maka sifat resistensi RIF isolat tersebut diduga kuat
disebabkan oleh perubahan Gln513Leu. Mutasi pada posisi ini sesuai dengan hasil penelitian yang telah dilakukan di berbagai negara [3, 7-12] dan juga diduga menyebabkan H37R
resistensi RIF tingkat tinggi, baik pada M. tuberculosis maupun E. coli [9]. Penjelasan hubungan antara mutasi ini dengan fenotipe resistensi RIF dicari pada tingkat protein.
C. Interaksi RNA Polimerase–Rifampin Berdasarkan Pemodelan Protein
Untuk dapat menjelaskan hubungan mutasi Gln513Leu di atas dengan fenotipe resistensi RIF yang dimiliki isolat M.
tuberculosis MDR P1, dipelajari pengaruh mutasi tersebut
pada interaksi RNAP dan RIF di tingkat protein. Hal tersebut dilakukan dengan pemodelan protein menggunakan struktur kristal kompleks RNAP inti (core) Taq dan RIF (ID PDB 1I6V) sebagai model struktur interaksi RNAP-RIF pada M. tuberculosis [6]. Struktur RNAP-RIF Taq tersebut dipilih sebagai model karena urutan nukleotida daerah-daerah yang mengandung mutasi penyebab RIFr
pada organisme Taq, M. tuberculosis, dan E. coli memiliki tingkat kesamaan yang tinggi, sebesar 91% [6]. Informasi mengenai struktur dan interaksi RNAP-RIF Taq digunakan untuk menjelaskan interaksi RNAP-RIF pada M. tuberculosis. Pemodelan protein ini dilakukan dengan bantuan program PyMOL versi Open-Source, DeLano Scientific, Amerika Serikat.
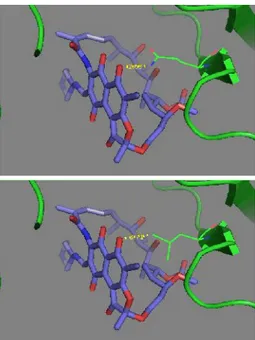
Residu Gln513 RNAP subunit β M. tuberculosis (residu 432 untuk penomoran M. tuberculosis), yang termutasi menjadi Leu pada isolat P1, merupakan homolog residu Gln393 RNAP subunit β Taq. Residu Gln pada posisi tersebut diketahui sebagai residu yang berperan dalam pengikatan
RIF pada RNAP subunit β dengan membentuk ikatan hidrogen antara rantai samping Gln yang bersifat polar dengan gugus hidroksil RIF [6]. Menggunakan program PyMOL, dilakukan mutasi residu Gln393, Taq menjadi Leu yang memiliki rantai samping non polar, dan dihitung jarak antar rantai samping residu tersebut dengan gugus hidroksil RIF, sebelum dan sesudah mutasi. Hasil pemodelan tersebut menunjukkan bahwa perubahan Gln menjadi Leu mengakibatkan jarak antara rantai samping residu tersebut dengan gugus hidroksil RIF menjadi lebih jauh, yaitu dari 2,63 Å menjadi 3,71 Å.
Rantai samping Gln yang pada awalnya membentuk ikatan hidrogen, pada atom nitrogennya, dengan gugus hidroksil RIF yang berperan penting untuk aktivitas RIF [6], digantikan oleh rantai samping Leu yang bersifat non-polar dan tidak dapat membentuk ikatan hidrogen. Pemodelan menggunakan PyMOL dengan model struktur RIF-RNAP
Taq juga menunjukkan bahwa mutasi Gln513Leu
mengakibatkan jarak antar rantai samping residu itu dengan gugus hidroksil RIF menjadi lebih jauh (3,71 Å) (Gambar 4). Perubahan-perubahan akibat mutasi Gln513Leu tersebut diduga kuat dapat menyebabkan ikatan hidrogen di atas tidak terbentuk. Telah diketahui bahwa ikatan hidrogen terjadi antara atom yang sangat elektronegatif dengan atom hidrogen yang terikat pada atom elektronegatif lainnya, dengan jarak kurang dari 3,5 Å [13-17]. Adapun atom yang memiliki keelektronegatifan tinggi adalah fluor, oksigen,
dan nitrogen.
Gambar 6. Interaksi RIF-RNAP subunit β hasil pemodelan PyMOL. (atas) Interaksi Gln393 RNAP wild type-RIF. (bawah) Interaksi Leu393 RNAP mutan-RIF. Gln393 Taq homolog dengan Gln513 M. tuberculosis. Mutasi mengakibatkan rantai samping residu tersebut menjadi non-polar dan merubah jarak antara rantai samping dengan gugus hidroksil RIF dari 2,63 Å menjadi 3,71 Å (ditunjukkan dengan angka berwarna kuning). Perubahan ini dapat mengakibatkan
hilangnya ikatan hidrogen yang pada awalnya terbentuk. Backbone dan atom karbon RNAP ditunjukkan dengan warna hijau, atom karbon RIF berwarna biru keunguan, atom nitrogen berwarna biru tua, dan oksigen berwarna merah.
Pada beberapa isolat M. tuberculosis yang terlebih dahulu diketahui memiliki mutasi penyebab RIFr, juga terdapat
perubahan residu asam amino dengan rantai samping polar menjadi residu dengan rantai samping non polar seperti hal di atas. Contohnya adalah mutasi Asp516 menjadi Val dan Gly [3,7,10-12]. Residu ini homolog dengan Asp396 Taq yang berikatan hidrogen dengan gugus hidroksil RIF. Termutasinya residu ini menjadi Val dan Gly dapat menyebabkan ikatan hidrogen tidak terbentuk. Kecenderungan tidak terbentuknya ikatan hidrogen di atas merupakan penjelasan yang dapat diberikan untuk sifat resistensi RIF isolat P1 yang disebabkan oleh mutasi Gln513Leu. Hilangnya ikatan hidrogen yang pada awalnya terbentuk dapat mengurangi afinitas pengikatan RIF sehingga RIF terikat lebih lemah pada RNAP subunit β. Hal di atas akan mengakibatkan RIF tidak dapat bekerja secara efektif, aktivitas RIF lebih bergantung pada kemampuannya untuk berikatan dengan RNAP. Selain hal di atas, konformasi RIF yang kaku diduga mengakibatkan RIF tidak dapat beradaptasi terhadap mutasi yang merubah bentuk dan lingkungan kimia kantung pengikatannya, sehingga pengikatan RIF pada RNAP sangat sensitif terhadap adanya mutasi di atas [6]. RIF yang terikat lemah pada RNAP diduga dapat mengakibatkan perubahan posisi RIF, terutama pada kondisi enzim tersebut di dalam sel, yang tentunya bersifat dinamis dan juga dipengaruhi oleh adanya molekul air. Bila posisi RIF berubah sedemikian rupa sehingga tidak lagi menghalangi jalur perpanjangan RNA, maka kemungkinan proses transkripsi akan terus berjalan dan bakteri akan resisten terhadap RIF.
IV.
KESIMPULAN
Salah satu isolat (P1) dari 20 isolat klinis M. tuberculosis
multidrug-resistant (MDR) asal Papua, yang tidak memiliki
mutasi-mutasi utama penyebab sifat resistensi rifampin (RIF), telah berhasil diketahui memiliki mutasi Gln513Leu yang diduga kuat menyebabkan fenotipe resistensi RIF yang dimilikinya. Hasil ini didasarkan atas adanya mutasi tingkat genotipe dan hasil analisis in silico yang menunjukkan perubahan sifat rantai samping residu polar menjadi non-polar serta perubahan jarak rantai samping tersebut dengan gugus hidroksil RIF. Mutasi tersebut dapat menyebabkan afinitas pengikatan RIF pada RNA polimerase (RNAP) berkurang sehingga RIF tidak dapat menginhibisi kerja RNAP pada proses transkripsi dan M. tuberculosis menjadi resisten terhadap antibiotik tersebut. Untuk memperkuat kesimpulan di atas, disarankan penelitian ini dilanjutkan dengan percobaan in vitro, misalnya dengan melakukan kloning dan ekspresi gen rpoB mutan, atau dengan pemodelan protein lebih lanjut untuk melihat kestabilan
protein mutan dibandingkan dengan protein wild type. Diharapkan pengetahuan mengenai mekanisme resistensi yang diperoleh pada penelitian ini dapat digunakan sebagai dasar desain obat baru untuk mengatasi masalah resistensi RIF dan MDR-TB pada umumnya.
DAFTAR PUSTAKA
[1] Musser, J.M. (1995), Antimicrobial agent resistance in mycobacteria: molecular genetic insights. Clin. Microb.
Rev., 8 (4), 496-514.
[2] Ramaswamy, S., Musser, J. M. (1998), Molecular genetic basis of antimicrobial agent resistance in
Mycobacterium tuberculosis: 1998 update, Tuberc. Lung Dis., 79, 3–29.
[3] Telenti, A., Imboden, P, Marchesi, F., Lowrie, D., Cole, S., Colston, M.J., Matter, L., Schopfer, K., Bodmer, T. (1993), Detection of rifampicin-resistant mutations in
Mycobacterium tuberculosis. Lancet. 341, 647–650.
[4] Mokrousov, I., Otten, T., Vyshnevskiy, B., Narvskaya, O. (2003), Allele-specificrpoB PCR assays for detection of rifampin-resistant Mycobacterium tuberculosis in sputum smears, Antimicrob. Agents Chemother., 47 (7), 2231-2235.
[5] Ubyaan, R., Maryuni, A.E., Sroyer, A., Palit, E.I.Y., Jukwati, Suaka, I.Y., and Ngili. (2012), Molecular analysis of rifampin-resistant Mycobacterium tuberculosis strains isolates from Papua, Indonesia, Int J. Pharmtech res, 4 (4), 1803-1811.
[6] Campbell, E.A., Korzheva, N., Mustaev, A., Murakami, K., Nair, S., Goldfarb, A., Darst, S.A. (2001), Structural mechanism for rifampicin inhibition of bacterial RNA polymerase, Cell, 104, 901-912. [7] Hirano, K., Abe, C., Takahashi, M. (1999), Mutations
in the rpoB gene of rifampin-resistant Mycobacterium
tuberculosis strains isolated mostly in asian countries
and their rapid detection by line probe assay, J. Clin.
Microbiol., 37 (8), 2663-2666.
[8] Moghazeh, S.L., Pan, X., Arain, T., Stover, C.K., Musser, J.M., Kreiswirth, B.N. (1996), Comparative antimycobacterial activities of rifampin, rifapentine, and KRM-1648 against a collection of rifampin-resistant Mycobacterium tuberculosis isolates with known rpoB mutations, Antimicrob. Agents Chemother., 40 (11), 2655-2657.
[9] Taniguchi, H., Aramaki, H., Nikaido, Y., Mizuguchi, Y., Nakamura, M., Koga, T., Yoshida, S., (1996), Rifampicin resistance and mutation of the rpoB gene in Mycobacterium tuberculosis, FEMS Microbiology
[10] Valim, A.R.M., Rossetti, M.L.R., Ribeiro, M.O., Zaha, A. (2000), Mutations in the rpoB gene of multidrug-resistant Mycobacterium tuberculosis isolates from Brazil, J. Clin. Microbiol., 38 (8), 3119-3122.
[11] Williams, D.L., Waguespack, C., Eisenach, K., Crawford, J.T., Portaels, F., Salfinger, M., Nolan, C.M., Abe, C., Sticht-Groh, V., Gillis, T.P. (1994), Characterization of rifampin resistance in pathogenic mycobacteria, Antimicrob.Agents Chemother., 38 (10), 2380-2386.
[12] Yue, J., Shi, W., Xie, J., Li, Y., Zeng, E., Wang, H. (2003), Mutations in the rpoB gene of multidrug-resistant Mycobacterium tuberculosis isolates from China, J.Clin. Microbiol., 41 (5), 2209-2212.
[13] Nachega, J.B., Chaisson, R.E. (2003), Tuberculosis drug resistance: a global threat, Clinical Infectious
Diseases, 36 (Suppl 1), S24–30.
[14] Kasper, D.L., Editor, (2005), Harrison’s Manual of
Medicine, McGraw-Hill Medical Publishing Division,
USA, 495-499.
[15] Levinson, W., (2006), Review of Medical Microbiology
and Immunology, 9th edition, The McGraw-Hill
Companies, Inc., San Fransisco, 161-162.
[16] Viedma, D.G. (2003), Rapid detection of resistance in
Mycobacterium tuberculosis: a review discussing
molecular approaches, Clin. Microbiol. Infect., 9, 349– 359.
[17] Zhang, G., Campbell, E.A., Minakhin, L., Richter, C., Severinov, K., Darst, S.A. (1999), Crystal structure of
Thermus aquaticus core RNA polymerase at 3.3 Å