Summary We studied the effects of five thinning treatments (T1 = 5.5, T2 = 11, T3 = 16.5, T4 = 22.5 and T5 = 28.5 m2 ha−1 basal area under bark) × two fertilizer treatments (F0 = unfer-tilized and F1 = ferunfer-tilized with 400 kg ha−1 N plus 229 kg ha−1 P) on growth and water relations of pole-sized Eucalyptus marginata J. Donn ex Sm. trees growing in southwestern Australia. Thinning reduced leaf area index (LAI) from 2.1 in the T4 and T5 treatments to 0.8 in the T1F0 treatment. Fertilizer had no effect on LAI in the T2, T4 or T5 treatments, but increased LAI by 45 and 20% in the T1 and T3 treatments, respectively. Thinning plus fertilizing increased diameter growth most in the fastest growing trees, from 0.4 cm year−1 for trees in the T5F0 and T5F1 treatments to 0.7 and 1.2 cm year−1 for trees in the T1F0 and T1F1 treatments, respectively. In both fertilizer treatments, stand basal area and volume growth increased with increasing stand density up to 15 m2 ha−1, and thereafter declined with increasing stand density, such that the growth rate of trees in the T5 treatment was only half of that at a stand density of 15 m2 ha−1. In response to fertilizer, growth rates of the slowest and fastest-growing trees increased from 0.35 and 3.5 m2 ha−1 year−1 (F0) to 0.56 and 5.4 m3 ha−1 year−1 (F1), respectively. Stand growth efficiency (growth per unit LAI) increased in response to thinning, and fertilizer increased stand growth efficiency at all stand densi-ties. Throughout the dry season, T5 trees had lower predawn shoot water potentials (Ψpd) (minimum of −1.5 MPa) than T1 or T2 trees (minimum of −0.7 MPa). Fertilizer decreased Ψpd in T5 trees (by −0.9 and −1.5 MPa, respectively, in F0 and F1), but not in T1 or T2 trees. Stand growth rate was closely related to cumulative midday water stress (CMWS) over the dry sea-son, and volume growth rate declined sharply from 6 m3 ha−1 year−1 at a CMWS of 130 MPa days, to zero at a CMWS of 220 MPa days. Application of fertilizer to thinned stands increased LAI, stand growth efficiency and stand growth. In unthinned stands, fertilizer increased stand growth efficiency and stand growth; however, it also increased tree water stress, which limited the fertilizer-induced increases in LAI and growth. We attribute the increase in tree and stand growth in response to application of fertilizer to increased photosynthetic
rates, increased allocation to stem wood, and in thinned stands also to higher LAIs.
Keywords: drought, growth efficiency, leaf area index, shoot water potential.
Introduction
Water and nutrients are two of the most important factors controlling the growth of forest trees (Nambiar et al. 1990). The relative importance of each factor in controlling growth varies with species, soil and climatic conditions. Under-standing how these factors control forest growth in a range of forest conditions (see Landsberg 1986, Landsberg 1989, Pereira et al. 1989, Gower et al. 1992, Raison and Myers 1992) is fundamental for the development of physiologically based forest growth models (Kimmins et al. 1990, McMurtrie and Landsberg 1992).
We have investigated the effects of thinning and application of fertilizer on Eucalyptus marginata J. Donn ex Sm. tree and stand growth. The specific objectives of the study were to quantify the effects of thinning and fertilization on tree and stand growth, leaf area index (LAI) and plant water stress, and to determine the effect of water stress on stand growth.
Methods
Study area
The experiment was located in an E. marginata stand at Ingle-hope Forest Block (32°45′ S, 116°11′ E), 9 km southeast of Dwellingup, Western Australia. Soil on the site is 0.5 to 1 m of yellow sandy loam overlying ferruginous concretionary grav-els in a sandy loam matrix up to 4 m thick. Beneath this, kaolinitic clay extends to granite bedrock at depths between 18 m and 57 m, with groundwater tables at between 20 and 32 m. The soils are of low fertility (Leeper 1970, Hingston et al. 1981). The climate of the area is typically Mediterranean. During the study, average annual rainfall at Dwellingup was
Growth and water relations of
Eucalyptus marginata
(jarrah) stands in
response to thinning and fertilization
G. L. STONEMAN,
1,2D. S. CROMBIE,
1K. WHITFORD,
1F. J. HINGSTON,
3R. GILES,
1C. C. PORTLOCK,
1J. H. GALBRAITH
3and G. M. DIMMOCK
31
Department of Conservation and Land Management, Research Centre, Dwellingup, WA 6213, Australia 2
Present address: Department of Conservation and Land Management, Research Centre, Como, WA 6152, Australia 3
CSIRO, Division of Forest Research, Private Bag, PO Wembley, WA 6014, Australia
Received June 2, 1994
1217 mm, average annual pan evaporation was 1277 mm, average annual maximum temperature was 21.6 °C and aver-age annual minimum temperature was 9.8 °C (Figure 1). Long-term average annual rainfall at the site is 1100 mm, 200 mm less than at Dwellingup. Average monthly maximum tempera-tures varied from about 30 °C in January and February to 14 °C in July.
Experimental design
Plots (40 m by 40 m with an 8 m buffer on all sides) were first thinned in 1964 (at age 40 years) to stand densities of about 7 (T1), 11 (T2), 15 (T3), and 18 (T4) m2 ha−1 basal area under bark, and stand density of the unthinned plots was about 22 m2 ha−1 (T5). In 1986, a fertilizer treatment and a second thinning were imposed. The experimental design comprised 10 treat-ments of five thinning treattreat-ments (stand basal area under bark of 5.5 (T1), 10.9 (T2), 16.4 (T3), 22.4 (T4) or 28.5 (T5) m2 ha−1 (unthinned) in 1986) × two fertilizer treatments (F0 = unfertil-ized and F1 = fertilunfertil-ized with 400 kg ha−1 N and 229 kg ha−1 P)
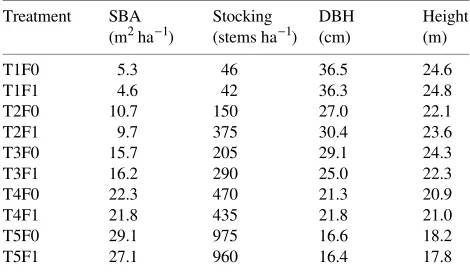
× three replicate plots. For the F1 plots, 200 kg ha−1 N and 229 kg ha−1 P were applied in autumn 1987 as 1145 kg ha−1 of diammonium phosphate, and 200 kg ha−1 N was applied in autumn 1988 as 588 kg ha−1 of ammonium nitrate. This rate of fertilizer application was chosen to give an optimum growth response (Stoneman et al. 1989). In both the first thinning in 1964 and the second thinning in 1986, the smaller, slower-growing trees were removed (Smith 1962). Table 1 shows the effect of the treatments on stand structure.
Growth and leaf area measurements
To estimate growth in diameter, basal area and bole volume, tree diameter over bark and bark thickness were measured on all trees in all plots in 1987 and 1991, and bole height was measured in 1985. Volume was calculated for each tree based on the equation of Pearce et al. (1992). To estimate the seasonal diameter growth pattern, stem diamter was measured monthly from dendrometer bands fitted at breast height on six trees per plot with diameter of about 30 cm. Leaf area index (LAI) of each plot was estimated based on allometric equations and
hemispherical photographs (Whitford 1991, Whitford unpub-lished data). The mean LAI for each plot was based on up to six separate estimates of LAI taken between spring 1987 and autumn 1990. Stand growth efficiency (Waring 1983) is de-fined as the basal area growth per plot per unit of LAI (m2 ha−1 year−1/LAI).
Water relations
Predawn shoot water potential (Ψpd) and midday shoot water potential (Ψmd) were measured with a pressure chamber (Scholander et al. 1965, Turner 1988) at 2- to 8-week intervals, from May 1988 to May 1989, and at about 2-week intervals from November 1989 to May 1990. These measurements were made in one plot of each of the T1F0, T1F1, T2F0, T2F1, T5F0 and T5F1 treatments. Additionally, shoot water potential (Ψs) was measured throughout the day on several occasions from November to April. These measurements were taken in one plot of each of the T1F0 and T1F1, or T2F0 and T2F1 treat-ments as well as in one plot of each of theT5F0 and T5F1 treatments. Shoot water potential was measured on two twigs on each of four dominant or codominant trees in each plot on each occasion for the predawn, midday and diurnal sets of measurements. Cumulative water stress (CWS) was calculated separately from both the Ψpd and Ψmd data for the 1988--1989 and 1989--1990 seasons as described by Ritchie and Hinckley (1975) and Myers (1988).
Results
Diameter growth
Diameter growth rate increased with decreasing stand density from 0.02 cm year−1 for trees on T5F0 plots to about 0.75 cm year−1 for trees on T1F0 plots (Figure 2). Application of fertilizer increased average diameter growth in all thinning treatments, with the greatest increase in rate occurring in trees growing at the lowest stand density (1.2 cm year−1 for trees in the T1F1 plots).
Analysis of average growth rates biased the data because the slower growing trees had been removed from the heavily thinned plots, whereas they remained in the unthinned and Figure 1. Average monthly values for maximum and minimum air
temperature, rainfall and pan evaporation at Dwellingup over the study period.
Table 1. Average stand basal area under bark (SBA), average stocking, average diameter at breast height under bark (DBH), and average tree height for all trees in each of the thinning and fertilizer treatments.
Treatment SBA Stocking DBH Height
lightly thinned plots and depressed the average growth rate (cf. Figures 2 and 3a). The fastest growing 200 stems ha−1 in the unthinned and lightly thinned plots exhibited the greatest re-sponse to both thinning and fertilizer, whereas the remaining trees exhibited little response to either treatment. In the unfer-tilized plots, diameter growth of the fastest growing 50 stems ha−1 increased from 0.35 cm year−1 for T5 plots to 0.75 cm year−1 for T1 plots, and the corresponding values for the fertilized plots were 0.53 and 1.25 cm year−1, respectively. Fertilizer caused only a small increase in diameter growth at high stand densities, but there was an interaction between
thinning and fertilizing resulting in a larger response to fertil-izer at the low stand densities.
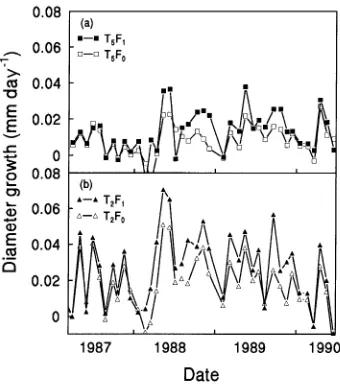
Diameter growth rates of trees in all thinning treatments were not significantly affected by fertilizer application in 1987, the first year after fertilizer was applied (Figure 4); however, in the second and third years, fertilizer increased diameter growth rates of trees in all thinning treatments. Di-ameter growth occurred mainly during the late autumn, winter and spring periods. The high growth rates recorded when the trees were water stressed during the February to April period were caused by bark swelling following rainfall events, and not to diameter growth (see Figure 7a).
Stand growth
Trees in the F0 treatment exhibited maximum basal area growth and volume growth rates at a stand basal area under bark of about 15 m2 ha−1 (Figures 5). Basal area growth rates of trees in the T1 and T5 treatments were about 58 and 50%, respectively, of that of trees growing at a stand density of 15 m2 ha−1. The percentage increase in basal area growth rate in response to fertilizer application was similar over the range of stand densities studied, with increases of 60, 70 and 60% at 5, 15 and 30 m2 ha−1, respectively. The largest absolute increases in basal area growth rate and volume growth rate in response to fertilizing were at a stand basal area of about 15 m2 ha−1, where basal area growth increased from 0.35 to 0.56 m2 ha−1 year−1 and volume growth increased from 3.5 to 5.4 m3 ha−1 year−1.
Leaf area index and stand growth efficiency
Thinning reduced LAI from 2.1 in the T4 and T5 treatments to 0.8 in the T1F0 treatment (Figure 6a). Fertilizing did not increase LAI in the T2, T4 or T5 treatments, but it increased LAI in the T1 and T3 treatments by 45 and 20%, respectively. Figure 2. Average diameter growth in relation to stand basal area for
the fertilized (F1) and unfertilized (F0) treatments. Error bars repre-sent one standard error. The fitted equations are F1: y = 2.250 − 0.6545 lnx, r2 = 0.98, n = 15, F0: y = 1.384 − 0.4034lnx, r2 = 0.90, n = 15.
Figure 3. Diameter growth in relation to stand basal area for the fertilized (F1) and unfertilized (F0) treatments for (a) the fastest growing 50 stems per hectare, (b) the second fastest growing 50 stems per hectare, (c) the second fastest growing 100 stems per hectare, and (d) the third fastest growing 100 stems per hectare. Error bars repre-sent one standard error. The fitted equations are for (a) F1: y = 1.443 − 0.0359x, r2 = 0.86, F0: y = 0.839 − 0.0171x, r2 = 0.66, (b) F1: y = 1.00 − 0.0242x, r2 = 0.70, F0: y = 0.437 − 0.0063x, r2 = 0.33, (c) F1: y = 0.569 − 0.0111x, r2 = 0.35, F0: y = 0.213−0.0012x, r2 = 0.03, (d) F1: y = 0.417 − 0.0092x, r2 = 0.31, F0: y = 0.229 − 0.0045x, r2 = 0.09.
Stand growth efficiency increased with decreasing stand density (Figure 6b). Fertilizing increased stand growth effi-ciency at all stand densities, from 0.08 to 0.17 m2 ha−1 year−1
in the T5 treatment and from 0.27 to 0.43 m2 ha−1 year−1 in the T1 treatment.
Water relations
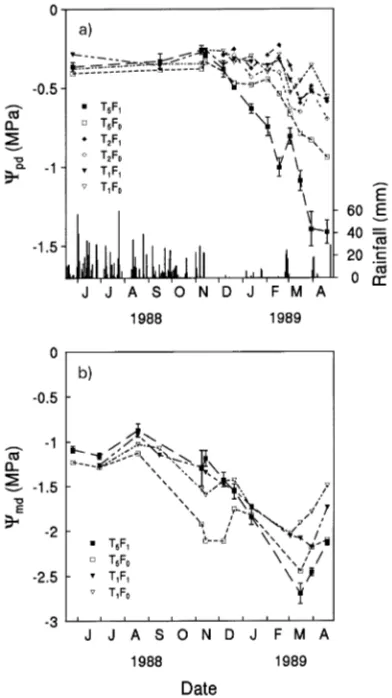
Predawn shoot water potential Trees in thinned plots had higher Ψpd than trees in unthinned plots, and fertilization tended to lower the Ψpd of trees in all T treatments (Figure 7a). In T5 and T2 trees, Ψpd fell below −0.4 MPa in mid-December 1988 and in mid- to late January 1989, respectively, whereas T1 trees maintained Ψpd above −0.4 MPa until after mid-Feb-ruary 1989 (Figure 7a). The differences between the thinning treatments increased as the Ψpd of all trees continued to fall during the remainder of the dry season to reach minimum values in late April when the Ψpd values of trees in the T5F0, T2F0 and T1F0 treatments were −0.94, −0.69 and −0.55 MPa, respectively.
Midday shoot water potential The Ψmd values were about
−1.2 MPa during the wet season and they started to decline at the end of August (Figure 7b). Trees in the T5F0 plot had significantly lower Ψmd (−2 MPa) during November--Decem-ber 1988 than trees in the other plots. The lowest Ψmd values Figure 5. (a) Stand basal area growth for fertilized (F1) and
unfertil-ized (F0) treatments in relation to stand basal area. Error bars represent one standard error. The fitted equations are Weibull curves of the type y = a(c/b)((x/b)^(c−1))(exp(−),(x/b)^c)) for F1, where a = 14.88, b = 21.76, c = 1.885, r2 = 0.59, n = 15, and for F0, where a = 9.028, b = 22.19, c = 1.878, r2 = 0.33 and n = 15. (b) Stand volume growth for fertilized (F1) and unfertilized (F0) treatments in relation to stand basal area. Error bars are for one standard error. The fitted equations are Weibull curves of the type y = a(c/b)((x/b)^(c−1))(exp(−(x/b)^c)) for F1where a = 137.5, b = 21.09, c = 1.924, r2 = 0.71, n = 15, and for F0 where a = 80.73, b = 21.15, c = 2.042, r2 = 0.46 and n = 15.
Figure 6. (a) Stand leaf area index (LAI) for fertilized (F1) and unfertilized (F0) treatments in relation to stand basal area. Error bars represent one standard error. The fitted equations are F1: y = 2.432e^(−4.119/x), r2 = 0.83, F0: y = 2.503e^(6.175/x), r2 = 0.90. (b) Stand growth efficiency for fertilized (F1) and unfertilized (F0) treat-ments in relation to stand basal area. Error bars are for one standard error. The fitted equations are F1: y = 0.488 − 0.0121x, r2 = 0.43, n = 15, F0: y = 0.316 − 0.0079x, r2 = 0.70, n = 15.
occurred in March 1989, when Ψmd of trees in the T1F0, T1F1 and T5F0 plots reached −2, −2.2 and −2.7 MPa, respectively.
Diurnal course of shoot water potential Early in the dry sea-son (November 14, 1988), there was little difference in Ψs among the treatments during the day, and the minimum Ψs was about −1.2 to −1.5 MPa (Figure 8a). By mid-January, the T5 trees had lower Ψs over most of the day, except around midday, than trees in the other treatments (Figure 8b). The difference between T5 trees and trees in the other treatments was still evident late in the dry season (April 19, 1989) (Figure 8d). Fertilizer decreased Ψs of T5 trees but not of T2 trees.
Effect of water deficits on growth Stand growth rate was in-versely related (r2 = 0.91) to cumulative midday water stress (CMWS) over the dry season, such that volume growth rate declined from 6 m3 ha−1 year−1 at a CMWS of 130 MPa days, to zero at a CMWS of 220 MPa days (Figure 9a). Stand growth was less closely related (r2 = 0.59) to cumulative predawn water stress (CPWS) over the dry season, although the curve indi-cated a sharp decline in volume growth as CPWS increased from 10 to 30 MPa days (Figure 9b).
Discussion
Both thinning and fertilizer application increased the growth rate of E. marginata trees and stands; however, the increase was limited to the fastest growing 200 stems per hectare. Thinning reduced LAI, resulting in less water stress and in-creased stand growth efficiency. Fertilizer inin-creased stand growth efficiency, and increased LAI of moderately to heavily thinned stands, but did not significantly increase LAI of lightly thinned or unthinned stands. Application of fertilizer had no
effect on predawn water stress in heavily thinned stands, but led to greater water stress in the unthinned stand.
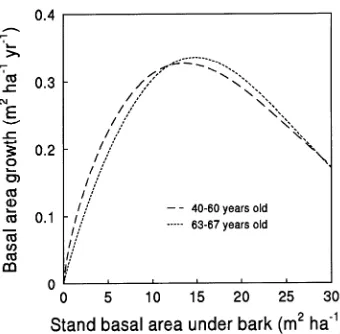
When contrasted with previous studies on this site (Abbott and Loneragan 1983, Stoneman et al. 1989), it seems that as the stand has aged the optimum stand density for growth has increased (Figure 10) (cf. Smith 1962, Shepherd and Forrest 1973). For trees aged 40--60 years, basal area growth was > 90% of maximum growth over the range of stand basal areas
Figure 8. Diurnal courses of shoot water potential (Ψs) for fertilized (F1) and unfertilized (F0) trees in the unthinned treatment (T5), the treatment thinned to 11 m2 ha−1 (T2), and the treatment thinned to 5 m2 ha−1 (T1) during the 1988--1989 dry season. Error bars represent one standard error.
Figure 9. Stand volume growth in relation to (a) cumulative midday water stress (CMWS), and (b) cumulative predawn water stress (CPWS). Data from the T1 plots are not included because there are too few trees on these plots to fully occupy the site. The fitted equations are: (a) y = 15379x^(−0.012x), r2 = 0.91, n = 4, and (b) y = −0.342+65.216/x, r2 = 0.59, n = 4.
under bark from 8.5 to 19.5 m2 ha−1, with maximum growth at 13.5 m2 ha−1. In the present study, which covered a growth period from age 63 to 67 years, basal area growth was > 90% of maximum growth over the range of stand basal areas of 10 to 20 m2 ha−1, with maximum growth at 15 m2 ha−1.
The increase in tree growth rate with thinning was associ-ated with a reduction in shoot water stress. Based on the observed relationship between photosynthesis and Ψpd for
E. marginata seedlings (Stoneman et al. 1994), we conclude that the reduced shoot water stress of trees in the thinned stands enabled greater rates of photosynthesis than in the unthinned stand. Additionally, because it usually favors allocation of photosynthates to shoots in preference to roots (Begg and Turner 1976, Axelson and Axelson 1986, Pereira and Pallardy 1989, Gower et al. 1992), reduced shoot water stress would have further increased bole growth rates. Reductions in stand density also result in increased light availability to remaining tree crowns and increased soil temperatures (Jenkins and Chambers 1989, Chen et al. 1993, Kinal 1993, Stoneman and Dell 1993). However, Stoneman and Dell (1993) found that these factors had less effect on E. marginata seedling growth than shoot water stress. Thinning had no effect on leaf nutrient concentrations (Hingston unpublished data), indicating that nutrition has little effect on the response of tree growth rates to thinning.
Stand density had a major influence on the pattern of E. mar-ginata stand growth. At low stand densities, even though tree growth rates were high, there were too few trees to occupy the site fully, and stand growth was less than maximal. Thus, there was a range of stand densities over which growth in an increas-ing number of trees counteracted reductions in tree growth rates with increasing stand density. At high stand densities, stand growth declined in E. marginata stands. Although this relationship between stand growth and stand density was hy-pothesized by Langsaeter (1941) (as quoted by Smith 1962 and Assmann 1970) there has been little published evidence to support it. We attribute the decline in stand growth at high stand densities to the low rate of self-thinning of this species. Thus at high stand densities, a higher proportion of assimilates was used for respiration and leaf formation by the large num-ber of surviving trees and so a lower proportion of assimilates was available for stem growth than at lower stand densities.
The application of fertilizer increases leaf nitrogen and phosphorus concentrations (Hingston unpublished data), and photosynthetic rates have been found to increase with increas-ing leaf nitrogen or phosphorus concentrations in E. marginata
(Stoneman unpublished data) and other Eucalyptus seedlings (Kirschbaum and Tompkins 1990, Sheriff and Nambiar 1991). Application of fertilizer containing nitrogen or phosphorus also reduces the root/shoot ratio of E. marginata seedlings (Stoneman unpublished data), other eucalypt seedlings (Cromer et al. 1984, Cromer and Jarvis 1990, Kirschbaum et al. 1992) and other forest tree species (Axelsson and Axelsson 1986, Gower et al. 1992), which could result in increased bole growth. Furthermore, the fertilizer treatment resulted in an increase in LAI of moderately and heavily thinned stands. Because an increase in LAI will increase
radia-tion intercepradia-tion (Monteith 1977, Legg et al. 1979, Linder 1985), an increase in LAI will generally also lead to an in-crease in stand growth. Based on this information, we attribute the increase in tree and stand growth in response to application of fertilizer to increased photosynthetic rates, increased alloca-tion to stem wood and higher LAIs.
The fertilizer-induced increase in plant water stress in the unthinned stands may be associated with an increased alloca-tion of assimilates to aboveground tissues relative to root tissues, which in the unthinned plots, with the highest LAI, resulted in an imbalance between water use by leaves and water supply by roots. This response is unusual because fertil-izer usually results in less plant water stress (Hillerdal-Hag-stromer et al. 1982, Brix and Mitchell 1986, Myers and Talsma 1992), as was observed in the thinned plots (Figure 10).
Increased stand growth efficiency in response to application of fertilizer was a reflection of the increased rates of photosyn-thesis per unit leaf area and increased allocation to stem wood. Increases in growth efficiency in response to fertilization have also been observed for Pseudotsuga menziesii (Mirb.) Franco (Binkley and Reid 1984), Pinus sylvestris L. (Waring and Schlesinger 1985), Pinus taeda L. and Pinus elliottii var.
elliottii Engelm. (Colbert et al. 1990). However, Vose and Allen (1988) found no increase in growth efficiency of Pinus taeda following fertilization. It seems likely that application of fertilizer will increase growth efficiency to a greater extent on nutrient-deficient soils and for species that require a high nutrient status.
Increased stand growth efficiency in response to thinning was partly a reflection of the improved water status of trees in thinned stands, which resulted in increased photosynthetic rates, increased allocation to stem wood and decreased alloca-tion to respiraalloca-tion and leaf formaalloca-tion. The increased stand growth efficiency in response to thinning was also partly a result of the selective removal of the slower growing trees (Stoneman unpublished data). Increases in growth efficiency in response to thinning have also been observed for Pseudot-suga menziesii (Waring et al. 1981, Binkley and Reid 1984, Velazquez-Martinez et al. 1992) and Pinus resinosa Ait. (Law et al. 1992). However, thinning did not increase the growth efficiency of Pinus contorta var. latifolia Engelm ex S. Wats. (Amman et al. 1988) or Pseudotsuga menziesii in the experi-ment of O’Hara (1989). Thinning may increase growth effi-ciency to a greater extent for species and sites where thinning causes large reductions in plant water stress, or in unthinned stands with a high LAI where thinning increases light avail-ability to leaves that would otherwise have been in deep shade.
Acknowledgments
We thank S. Bellgard, I. Freeman and M. Reynolds for technical assistance and I. Abbott, F.J. Bradshaw, P. Kimber, J. McGrath and M. Rayner for reviewing the manuscript.
References
Amman, G.D., G.D. Lessard, L.A. Rasmussen and C.G. O’Neil. 1988. Lodgepole pine vigor, regeneration, and infestation by mountain pine beetle following partial cutting on the Shoshone National Forest, Wyoming. U.S. For. Serv. Res. Paper INT-396.
Assmann, E. 1970. The principles of forest yield study. Pergamon Press, Oxford.
Axelsson, E. and B. Axelsson. 1986. Changes in carbon allocation patterns in spruce and pine trees following irrigation and fertiliza-tion. Tree Physiol. 2:189--204.
Begg, J.E. and N.C. Turner. 1976. Crop water deficits. Adv. Agron. 28:161--217.
Binkley, D. and P. Reid. 1984. Long-term responses of stem growth and leaf area to thinning and fertilization in a Douglas-fir planta-tion. Can. J. For. Res. 14:656--660.
Brix, H. and A.K. Mitchell. 1986. Thinning and nitrogen fertilization effects on soil and tree water stress in a Douglas-fir stand. Can. J. For. Res. 16:1334--1338.
Chen, J., J.F. Franklin and T.A. Spies. 1993. Contrasting microcli-mates among clearcut, edge, and interior of old-growth Douglas-fir forest. Agric. For. Meteorol. 63:219--237.
Colbert, S.R., E.J. Jokela and D.G. Neary. 1990. Effects of annual fertilization and sustained weed control on dry matter partitioning, leaf area, and growth efficiency of juvenile loblolly and slash pine. For. Sci. 36:995--1014.
Cromer, R.N. and P.G. Jarvis. 1990. Growth and biomass partitioning in Eucalyptus grandis seedlings in response to nitrogen supply. Aust. J. Plant Physiol. 17:503--515.
Cromer, R.N., A.M. Wheeler and N.J. Barr. 1984. Mineral nutrition and growth of Eucalyptus seedlings. N.Z. J. For. Sci. 14:229--239. Gower, S.T., K.A. Vogt and C.C. Grier. 1992. Carbon dynamics of Rocky Mountain Douglas-fir: influence of water and nutrient avail-ability. Ecol. Monogr. 62:43--65.
Hillerdal-Hagströmer, K., E. Matton-Djos and J. Hellkvist. 1982. Field studies of water relations and photosynthesis in Scots pine. 2. Influence of irrigation and fertilization on needle water potential of young pine trees. Physiol. Plant. 54:295--301.
Hingston, F.J., G.M. Dimmock and A.G. Turton. 1981. Nutrient distri-bution in a jarrah (Eucalyptus marginata) ecosystem in south-west-ern Australia. For. Ecol. Manage. 3:183--207.
Jenkins, M.W. and J.L. Chambers. 1989. Understorey light levels in mature hardwood stands after partial overstorey removal. For. Ecol. Manage. 26:247--256.
Kimmins, J.P., P.G. Comeau and W. Kurz. 1990. Modelling the inter-actions between moisture and nutrients in the control of forest growth. For. Ecol. Manage. 30:361--379.
Kinal, J. 1993. Soil temperature in the northern jarrah forest in relation to plant and litter cover and implications for the periods and depths at which temperatures permit Phytophthora cinnamomi activity. PhD Thesis, Murdoch Univ., Murdoch, Western Australia, 137 p.
Kirschbaum, M.U.F. and D. Tompkins. 1990. Photosynthetic re-sponses to phosphorus nutrition in Eucalyptus grandis seedlings. Aust. J. Plant Physiol. 17:527--535.
Kirschbaum, M.U.F., D.W. Bellingham and R.N. Cromer. 1992. Growth analysis of the effect of phosphorus nutrition on seedlings of Eucalyptus grandis. Aust. J. Plant Physiol. 19:55--66.
Landsberg, J.J. 1986. Physiological ecology of forest production. Academic Press, New York, 198 p.
Landsberg, J.J. 1989. Solar energy conversion efficiency project. In Biomass Production by Fast-Growing Trees. Eds. J.S. Pereira and J.J. Landsberg. Kluwer Academic Publishers, Dordrecht, pp 281--284.
Law, B.E., K.H. Riitters and L.F. Ohmann. 1992. Growth in relation to canopy light interception in a red pine (Pinus resinosa) thinning study. For. Sci. 38:199--202.
Leeper, G.W. 1970. Soils. In The Australian Environment, 4th Edn. Ed. G.W. Leeper. Melbourne Univ. Press, Melbourne, Victoria, pp 21--31.
Legg, B.J., W. Day, D.W. Lawlor and K.J. Parkinson. 1979. The effects of drought on barley growth: models and measurements showing the relative importance of leaf area and photosynthetic rate. J. Agric. Sci. 92:703--716.
Linder, S. 1985. Potential and actual production in Australian forest stands. In Research For Forest Management. Eds. J.J. Landsberg and W. Parsons. CSIRO Forest Research, Canberra, Australia, pp 11--35.
McMurtrie, R.E. and J.J. Landsberg. 1992. Using a simulation model to evaluate the effects of water and nutrients on the growth and carbon partitioning of Pinus radiata. For. Ecol. Manage. 52:243--260.
Monteith, J.L. 1977. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond., B Biol. Sci. 281:277--294. Myers, B.J. 1988. Water stress integral----a link between short-term
stress and long-term growth. Tree Physiol. 4:315--323.
Myers, B.J. and T. Talsma. 1992. Site water balance and tree water status in irrigated and fertilised stands of Pinus radiata. For. Ecol. Manage. 52:17--42.
Nambiar, E.K.S., R. Squire, R. Cromer, J. Turner and R. Boardman. 1990. Management of water and nutrient relations to increase forest growth. For. Ecol. Manage. 30:1--476.
O’Hara, K.L. 1989. Stand growth efficiency in a Douglas-fir thinning trial. Forestry 62:409--418.
Pearce, C.J., W.J. Boardman and M.A. Green. 1992. Data integration and analysis. In Application of Modern Inventory Techniques in the Forests of Western Australia. Ed. R.D. Spencer. Western Australian Department of Conservation and Land Management, Perth, WA, pp 44--55.
Pereira, J.S., S. Linder, M.C. Araujo, H. Pereira, T. Ericsson, N. Bor-ralho and L.C. Leal. 1989. Optimization of biomass production in Eucalyptus globulus plantations----a case study. In Biomass Produc-tion by Fast Growing Trees. Eds. J.S. Pereira and J.J. Landsberg. Kluwer Academic Publishers, Dordrecht, pp 101--121.
Pereira, J.S. and S. Pallardy. 1989. Water stress limitations to tree productivity. In Biomass Production by Fast-Growing Trees. Eds. J.S. Pereira and J.J. Landsberg. Kluwer Academic Publishers, Dor-drecht, pp 37--56.
Raison, R.J. and B.J. Myers. 1992. The Biology of Forest Growth experiment: linking water and nitrogen availability to the growth of Pinus radiata. For. Ecol. Manage. 52:279--308.
Ritchie, G.A. and T.M. Hinckley. 1975. The pressure chamber as an instrument for ecological research. Adv. Ecol. Res. 9:165--254. Scholander, P.F., H.T. Hammel, E.D. Bradstreet and E.A.
Hemming-sen. 1965. Sap pressure in vascular plants. Science 148:338--345. Shepherd, K.R. and W.G. Forrest. 1973. Growth of radiata pine
fol-lowing thinning. Commonw. For. Rev. 52:133--142.
Sheriff, D.W. and E.K.S. Nambiar. 1991. Nitrogen nutrition, growth and gas exchange in Eucalyptus globulus Labill. seedlings. Aust. J. Plant Physiol. 18:37--52.
Smith, D.M. 1962. The practice of silviculture, 7th Edn. John Wiley & Sons, New York, 578 p.
Stoneman, G.L. and B. Dell. 1993. Growth of Eucalyptus marginata (jarrah) seedlings in a greenhouse in response to shade and soil temperature. Tree Physiol. 13:239--252.
Stoneman, G.L., N.C. Turner and B. Dell. 1994. Leaf growth, photo-synthesis and tissue water relations of greenhouse-grown Eucalyp-tus marginata seedlings in response to water deficits. Tree Physiol. 14:633--647.
Turner, N.C. 1988. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 9:289--308.
Velazquez-Martinez, A., D.A. Perry and T.E. Bell. 1992. Response of aboveground biomass increment, growth efficiency, and foliar nu-trients to thinning, fertilization, and pruning in young Douglas-fir plantations in the central Oregon Cascades. Can. J. For. Res. 22:1278--1289.
Vose, J.M. and H.L. Allen. 1988. Leaf area, stemwood growth, and nutrient relationships in loblolly pine. For. Sci. 34:547--563. Waring, R.H. 1983. Estimating forest growth and efficiency in relation
to canopy leaf area. Adv. Ecol. Res. 13:327--354.
Waring, R.H., K. Newman and J. Bell. 1981. Efficiency of tree crowns and stemwood production at different canopy leaf densities. For-estry 54:129--137.
Waring, R.H. and W.H. Schlesinger. 1985. Forest ecosystems concepts and management. Academic Press, Orlando, 340 p.