256 (2001) 241–251
www.elsevier.nl / locate / jembe
Anoxic survival of Macoma balthica: the effect of
antibiotics, molybdate and sulphide
* ´
Albertus de Zwaan , Bartholomeus E.M. Schaub, Jose M.F. Babarro
Netherlands Institute of Ecology, Centre for Estuarine and Coastal Ecology, P.O. Box 140, 4400 AC Yerseke, The Netherlands
Received 7 July 2000; received in revised form 13 September 2000; accepted 26 October 2000
Abstract
In anoxic semi-closed systems, the survival time of the clam Macoma balthica was compared to clams which were incubated in the presence of several antibiotics (chloramphenicol, 5-oxy-tetracycline hydrochloride, penicillin, streptomycin, a mix of penicillin and streptomycin and a mix of chloramphenicol, polymyxin, neomycin and penicillin), sulphide and chloramphenicol at pH 6.8 and 8.2 and molybdate (specific inhibitor of the process of sulphate reduction). The aim was to detect maximum survival times of this clam and indications for the cause of mortality under the conditions tested. Median survival time (LT ) of the clam was 4.8 days (at 1950 8C) in incubations without any addition. Added sulphide (200mM) decreased survival time. At pH 8.2, LT50decreased by 20.8% and at pH 6.8 by 35.2%. However, added molybdate, which suppressed biotic sulphide formation, did not improve survival time (LT5054.4 days). Biotic sulphide probably did not speed up mortality rate, but indicated excessive growth of sulphate reducing bacteria once mortality started. The presence of different antibiotics increased significantly survival time (LT ) from 8.9 to 14.9 days. Qualitative estimations were made of the numbers of50
bacteria present in the systems. Compared to a seawater control, highest numbers were observed in the incubation of clams without additions and in the presence of molybdate. Nevertheless, due to the presence of molybdate, bacteria numbers were significantly lower. However, very low numbers of bacteria were observed in the incubations of clams in the presence of chloramphenicol. These data demonstrated that the presence and proliferation of bacteria was probably the cause of death of the clams. 2001 Elsevier Science B.V. All rights reserved.
Keywords: Anoxia; Antibiotics; Bivalves; Molybdate; Sulphide; Mortality
*Corresponding author. Tel.: 131-113-577-495; fax: 131-113-573-616. E-mail address: [email protected] (A. de Zwaan).
1. Introduction
Benthic macrofaunal species show great variability in anoxic tolerance, even within closely related taxonomic groups. A comparison of studies is often complicated by differences in experimental set-up and conditions used to estimate this tolerance (Groenendaal, 1980). In a study of four bivalve species from the Mediterranean zone we showed that the experimental set-up had a great impact on anoxic survival time (De Zwaan and Eertman, 1996). In closed systems, a decrease in pH of the incubation medium and accumulation of ammonium and sulphide occurred. Both a high dose of cadmium and the broad-spectrum antibiotic chloramphenicol increased survival time. Also a postponed shell blackening and biotic sulphide accumulation was observed.
In the present survey we extend our study to anoxic survival of Macoma balthica, a bivalve from temperate Baltic and Atlantic coastal area. Past reports of mortality rate of this clam show wide variability in anoxic survival. Brafield (1963) observed that this clam only tolerated anoxia for 2 or 3 days (temperature not given). Dries and Theede (1974) reported LT50 values of specimens collected in the Western Baltic Sea of 22 and 20 days at 10 and 158C, respectively. More recent, Jahn and Theede (1997) reported median mortality values of 8–12 days at 108C for clams from the same location. The large variation in data for M. balthica raises the question whether the results obtained in closed systems indeed reflect artefacts or tolerance to anoxia. Understanding of the actual mechanisms causing death may help explain the inconsistency in literature data. Clams (Macoma balthica) were collected from a mudflat of the Dutch coastal waters. The role of biotic sulphide and the influence of exogenous sulphide were studied in semi-static systems. We also compared the effect of chloramphenicol and a number of other antibacterial agents applied in aquaculture suppressing bacterial activity. The activity of bacteria was followed indirectly by estimating the release of sulphide and ammonium and the recording of pH, as well as by counting bacterial numbers in the systems.
2. Material and methods
2.1. Individuals
Infaunal specimens of M. balthica were sampled at Paulinapolder, a tidal flat of the river Scheldt estuary in the southwestern part of the Netherlands. Average shell length of the clams was 16.862.1 mm (n550). The average weight was 0.47 g and the volume 0.41 ml. Salinity at the sampling site was 2762 psu and the temperature 198C. The clams were kept in an aquarium with sterilised sandy sediment and well-aerated unfiltered running seawater of 198C and 31 psu, pumped in from an inlet in the Eastern Scheldt in front of the institute at Yerseke. After 4 days the animals were purged with filtered seawater. The next day the clams were used in the experiments.
2.2. Protocol of experiments
vigorously bubbling for 2 h with N . The pH of the anoxic seawater was about 8.2.2 From this reservoir under continuous flow of N , seawater was siphoned into ten 1-l jars.2 The oxygen concentration after filling was less than 0.15 mg / l (Winkler method). To each jar the following antibiotics were added: 5 mg chloramphenicol, 10 mg 5-oxytetracycline hydrochloride, 100 000 IU penicillin-G (sodium salt) and 100 mg streptomycin sulphate, 100 000 IU penicillin-G, 100 mg streptomycin sulphate, 20 ml Provasoli’s antibiotic concentrated solution containing 240 000 IU penicillin-G, 1 mg chloramphenicol, 6000 IU polymyxin B sulphate and 1.2 mg neomycin (Sigma). As an inhibitor of the process of sulphate reduction, 5 g sodium molybdate (20.8 mM final concentration) was added to one flask. In order to study the effect of molybdate separately on survival time also a flask was used in which 5 g sodium molybdate and 5 mg chloramphenicol were both added. Under these conditions bacterial growth should be inhibited by the activity of chloramphenicol. Also two flasks contained anoxic seawater without additions: one without and one with animals.
In a similar way, four flasks were filled from a reservoir to which 25 mM Tris was added. Two of these flasks were adjusted with 1 M HCl to a pH of 6.8 and two to a pH of 8.2. From each of these two flasks, to one flask 5 mg chloramphenicol was added and to the other 5 mg chloramphenicol and 200 mM sulphide. Sulphide was added from a stock solution (5 ml of 40 mM) of washed crystals of Na S2 ?6–9H O, neutralised with2 HCl.
After addition of 50 individuals the bottles were sealed with rubber stoppers and incubated at 198C. Daily aliquots were taken for the determination of pH, sulphide, ammonium and bacterial count.
All incubation media were exchanged every 5 days in order to prevent decomposition of compounds, especially antibiotics. In such a manner a semi-static system was obtained. No food was added during anoxic incubations. The clams occupied the bottom of the flasks in one layer. Mortality was controlled daily. Mortality was assessed by the failure of constriction after the mantle edge of gaping animals was touched. Dead animals were removed from the incubation containers. The experiment was conducted in autumn 1999.
2.3. Determination of sulphide, ammonium and pH
Sulphide was determined after Svenson (1980) and ammonium after Truesdale (1971). The pH was measured using a Radiometer PHM 82 pH meter with a sulphide insensitive electrode.
2.4. Bacterial count
2.5. Statistics
The non-parametric Kaplan–Meier test was used to estimate log-rank and Wilcoxon values for comparing the survival curves (Kaplan and Meier, 1958). A confidence limit of 95% was used to test the significance of differences between groups. LT50 values (median survival times) were estimated using the trimmed Spearman–Karber method (a 510%) (Hamilton et al., 1997).
Throughout the manuscript the term sulphide refers to total sulphide, and the term Tris to the buffer, tris(hydroxymethyl)aminomethane.
3. Results
In a preliminary experiment we compared controls with clams treated with strep-tomycin sulphate by incubating three replicates (n550) for each condition. LT50 values were 4.7060.12 and 9.3160.21, respectively. Since variability between replicates of 50 clams each was small, we have not used replicates in our studies to the effect of different chemicals on survival time.
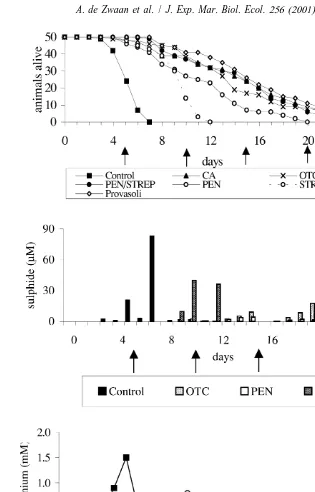
Fig. 1A shows the anoxic survival curves of clams incubated in anoxic seawater compared to the incubations in the presence of different antibiotics. Median survival time (LT ) of Macoma in the flask without addition of antibiotics was 4.8 days. A50 positive effect on survival time was observed due to the presence of all (combinations of) antibiotics. Most favourable was Provasoli’s antibiotic solution that increased LT50 from 4.8 to 14.9 days, closely followed by penicillin plus streptomycin (LT50513.4 days), chloramphenicol (LT50513.3 days) and 5-oxytetracycline (LT50513.0 days). The curves of these antibiotics were mutually not significantly different (all comparisons
1
P.0.05). Except for 5-oxytetracycline (sulphide #17mM, NH4 #0.3 mM) there was in these incubations virtually no biotic formation of sulphide (#2 mM) and small
1
increases of ammonium (NH4 range: 0.02–0.12 mM) in the medium (Fig. 1B and C). In the medium with penicillin there was some sulphide (#4 mM) and ammonium
1
formation (NH4 #0.14 mM), but much less as compared to the medium with 5-oxytetracycline. However, the survival time was significantly shorter as compared to this and previous media (P,0.001). Least favourable on survival time was addition of
1
streptomycin. Accumulation of sulphide (#36mM) and ammonium (NH4 #0.80 mM) was in this incubation medium by far highest of all media containing antibiotics (Fig. 1B and C). The LT50 value of medium with penicillin and streptomycin was 10.2 and 8.9 days, respectively (Fig. 1A), and the survival curves differed significantly (P,0.05). In
1
the control, sulphide (#83mM) and ammonium (NH4 #1.5 mM) accumulation was highest (Fig. 2B). In general ammonium preceded sulphide accumulation (see e.g. 5-oxytetracycline and control; Fig. 1B and C).
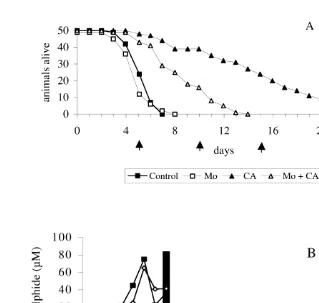
Fig. 2. Survival curves (A) and changes in concentration of sulphide and ammonium (B) under semi-static anoxic incubation of Macoma balthica without additions (Control) and in the presence of chloramphenicol (CA), molybdate (Mo) and molybdate1chloramphenicol (Mo1CA). Arrows indicate when incubation medium was exchanged. Sulphide values in the presence of chloramphenicol and in the presence of molybdate with and without chloramphenicol were below the detection limit (,1mM).
1
sulphide formation, but ammonium accumulation (NH4 #1.3 mM) appeared hardly to be affected by molybdate (Fig. 2B).
In the Fig. 3, the results for the bacterial counts are shown. Compared to the flask with seawater without animals, roughly on average 14 times more bacteria were observed in the water of the flask with animals. In the medium containing molybdate a nine-fold increase of bacteria was observed. In the presence of chloramphenicol the bacterial count were much lower compared to flask with animals without any addition, and only 3–5 times more than in the flask with only seawater.
Fig. 3. Bacterial counts in anoxic seawater without clams (SW) and anoxic incubations of Macoma balthica without additions (control) and in the presence of chloramphenicol (CA), molybdate (Mo) and molybdate1
chloramphenicol (Mo1CA). Bacterial counts represent means6S.D. (n53) of the average value of all daily counts until complete mortality.
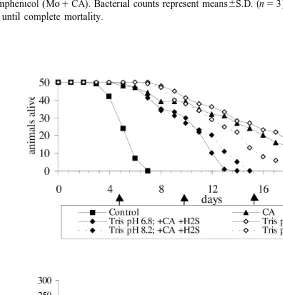
mM) decreased survival time. The effect was more pronounced at a lower pH. At pH 8.2, the LT50decreased by 20.8% (from 12.4 to 9.9 days) and at pH 6.8 by 35.2% (from 14.8 to 9.6 days) (Fig. 4). However, the relative decreases were an order of magnitude smaller than the positive effect of the simultaneously added chloramphenicol. Therefore, survival in the presence of sulphide was still much longer (about 200% overall increase) as for the control without antibiotic (LT50 4.8 days).
During the first 5 days between medium exchange, the added sulphide concentration decreased considerably. In the second interval of 5 days this occurred at a lower extend and from day 10 onward the sulphide concentration maintained at the initial value (Fig. 4B). Fortunately, the changes in sulphide concentrations at both pH values were identical.
4. Discussion
Knowledge of the survival time of bivalves under anoxia is important for understand-ing the impact of poor water quality. Without accurate estimates of how long anoxia must persist to kill benthic organisms, the impact of eutrophication on ecosystems cannot be modelled adequately and incorporated in water quality management decisions. The antibiotics used can be grouped on their mode of action. Penicillin-G is active primarily against gram-positive bacteria. It interferes with the synthesis of the bacterial cell wall. Chloramphenicol, neomycin and 5-oxytetracycline are so-called broad-spec-trum antibiotics, because they act both on gram-positive and gram-negative bacteria. They interfere with bacterial protein synthesis by binding to 50S (chloramphenicol) or 30S subunit (others) of the ribosome. Streptomycin, like neomycin, is an amino-glycoside; both are used clinically against gram-negative bacteria. Polymyxin B binds to and interferes with the permeability of the bacterial cytoplasmic membrane. Provasoli’s antibiotic concentrated solution is composed of antibiotics acting on the cell wall, biosynthetic processes of protein and the cytoplasmic membrane. The beneficial effect of the antibiotics indicates that the growth of opportunistic, partially pathogenic, bacteria to the bivalve is strongly reduced or blocked. Although we have not made dose response curves, we assume from the fact that the broad spectrum antibiotics and the combination of penicillin (gram positive) and streptomycin (gram negative) showed small in-significant differences that all applied concentrations were appropriate to give maximal effects (Fig. 1). Moreover, in a pilot experiment in which we used half the con-centrations for all antibiotics, the positive effects were slightly less (75–90%).
compared to 5-oxytetracycline. This indicates that mortality is not tightly linked to deterioration of the quality of the medium.
In all cases endogenous production of these metabolites is not gradual from the onset, but starts just before (ammonium), or is initiated (sulphide) by mortality of the bivalves and subsequently proceeds progressively. It therefore reflects excessive growth of bacteria in this stage. They possibly indicate necrosis of tissue which may serve as substrate for heterotrophic bacteria. From the fact that the broad band antibiotics and combinations of antibiotics never fully suppressed ammonium formation in contrast to sulphide, we assume that its formation in these incubations (in the range of 0.02–0.12 mM) may arise largely from catabolism of the bivalve.
The observation that molybdate, like most broad spectrum antibiotics, completely blocked sulphide formation, but did not increase survival time, indicates that biotic sulphide does not affect the velocity of mortality in our semi-static system. Despite the proliferation of sulphate reducing bacteria in the control and accumulation of sulphide as opposed to the molybdate incubation, the survival times were 4.8 and 4.4 (P.0.05), respectively. The small, not significant, negative effect by molybdate must have been due to intrinsic properties of the chemical. When the uptake time of molybdate in the tissues of the clams is enlarged by the addition of chloramphenicol, the negative effect became much more pronounced (Fig. 2).
Our particle counts represent bacteria that proliferate in association with the bivalves. The number did not increase in the anoxic seawater control.
The bacterial numbers in medium with chloramphenicol probably represents the relative density of bacteria associated with the clams when introduced to anoxic media, because there was no increase with incubation time. Taking this as a reference, the increase in the controls was about 1.5 times that of the increase in the presence of molybdate. This indicates that about one third of the increase in bacterial numbers of the control was accounted for by sulphate reducing bacteria and the remainder by others. In contrast to the incubations with antibiotics, molybdate did not clearly decrease ammonium accumulation (Fig. 2), although bacterial growth (of sulphate reducers) was inhibited (Fig. 3). Other bacteria than sulphate reducing bacteria (e.g. heterotrophic bacteria) must have been responsible for ammonium production. That sulphide and ammonium production occurs independently follows from the fact that both accumulate not synchronous (Fig. 1A, B and Fig. 2B).
In contrast to its biotic formation addition of sulphide to the incubation medium at the onset of anoxic incubation had a negative effect on survival. Adding 200mM at pH 8.2 and 6.8 (Fig. 4) assessed toxicity of external sulphide. At both pH values there was an important decrease in survival time. However, an almost twice as strong reduction was observed at the lower pH. A same pH dependence has been reported for fish (Longwell and Pentelow, 1935; Broderius et al., 1977) and crustacea (Vismann, 1996). The
2 22
Especially in the time interval between the start and the first water exchange there was a strong decrease of the added sulphide concentration. Some oxidation by residual oxygen in the medium and tissue water may have occurred, but also the system could have buffered a part by precipitation. This was for instance apparent from the prompt blackening of the shell surface.
Also, Jahn and Theede (1997) exposed specimens of M. balthica under hypoxic and hypoxic–sulphidic conditions. They concluded that there were no differences between both groups, although LT50 values at 108C ranged from 8 to 12 days. Our results obtained at 198C are in a comparable LT50range of 9.6–14.8 (pH 6.8) and 9.9–12.4 (pH 8.2) with and without sulphide, respectively. The non-parameter Kaplan–Meier test gave highly significant differences for both incubation media. That our values are close to those of Jahn and Theede (1997) despite a much higher incubation temperature is probably due to the fact that we added chloramphenicol to both media. In the control spontaneous generation of sulphide may mask the negative effects of added sulphide, especially when the applied concentration is low.
Two congeneric clam species Macoma secta and Macoma nasuta showed a lower tolerance to anoxia in the presence of 100mM sulphide (Levitt and Arp, 1991). To the experimental group 50 000 IU penicillin-G and 50 mg streptomycin was added to avoid the growth of sulfur-utilizing bacteria, but not to the control group. Despite the added antibiotics, the experimental group for both species showed about a 2.5-fold shorter survival. Both antibiotics together in these concentrations were also tested by us in a pilot experiment and give about 90% of the positive effect on survival with the (double) doses or broad band antibiotics reported in this study. We observed also a negative effect of 200 mM sulphide on survival (Fig. 4), but due to the simultaneous presence of chloramphenicol, the overall effect was a 2-fold higher survival time than for the control without antibiotic. Since Levitt and Arp (1991) used the experimental clams 1–2 days after they were collected from a tidal mudflat it is not likely that they were not associated with bacteria which strongly must have reduced survival of the controls during anoxic treatment. A control group containing the antibiotics of the experimental group too, would have been desirable.
In conclusion, it is obvious that studies of anoxic resistance in closed systems, even with periodical water exchanges, result in strongly underestimated survival times of M. balthica and other bivalves. This is likely due to bacterial proliferation which are associated with the bivalve. Our studies do not yet answer the question whether the causes of death by bacteria is mainly water quality deterioration by release of noxious products and / or infection and tissue injury.
Ammonium accumulation precedes sulphide accumulation and depends on bacteria which cause reduced survival time. The biotic formation of sulphide has no effect on survival time in contrast to added sulphide at the onset of the anoxic incubation.
Acknowledgements
work was partially financed by the EC project ROBUST (ENV4-CT96-0218, ELOISE publication no. 168). J.M.F. Babarro acknowledges the receipt of a post-doctoral grant
´
from the Ministerio de Educacion y Ciencia (1999–2001). This is publication no. 2687 of the Netherlands Institute of Ecology, Centre for Estuarine and Coastal Ecology, Yerseke. [SS]
References
Brafield, A.E., 1963. The effects of oxygen deficiency on the behaviour of Macoma balthica. Anim. Behav. 11, 345–346.
Broderius, S.J., Smith, Jr. L.L., Lind, D.T., 1977. Relative toxicity of free cyanide and dissolved sulfide forms to the fathead minnow (Pimephales promelas). J. Fish. Res. Board Can. 34, 2323–2332.
De Zwaan, A., Eertman, R.H.M., 1996. Anoxic or aerial survival of bivalves and other euryoxic invertebrates as a useful response to environmental stress. A comprehensive review. Comp. Biochem. Physiol. 113C, 299–312.
Dries, R.-R., Theede, H., 1974. Sauerstoffmangelresistenz mariner Bodenevertebraten aus der Westlichen Ostsee. Mar. Biol. 25, 327–333.
Groenendaal, M., 1980. Tolerance of the lugworm (Arenicola marina) to sulphide. Neth. J. Sea Res. 14, 200–207.
Hamilton, M.A., Russo, R.C., Thurston, R.V., 1997. Trimmed Spearmen–Karber method for estimating median lethal concentrations in toxicity bioassays. Environ. Sci. Technol. 11, 714–719.
Jahn, A., Theede, H., 1997. Different degrees of tolerance to hydrogen sulphide in populations of Macoma balthica (Bivalvia, Tellinidae). Mar. Ecol. Prog. Ser. 154, 185–196.
Kaplan, E.L., Meier, P., 1958. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 53, 457–481.
Levitt, J.M., Arp, A.J., 1991. The effect of sulfide on the anaerobic metabolism of two congeneric species of mudflat clams. Comp. Biochem. Physiol. 98B, 339–347.
Longwell, J., Pentelow, F.T.K., 1935. The effect of sewage on brown trout (Salmo trutta). J. Exp. Biol. 12, 1–12.
Millero, F.J., Plese, T., Fernandez, M., 1988. The dissociation of hydrogen sulfide in seawater. Limnol. Oceanogr. 33, 269–274.
Svenson, A., 1980. A rapid and sensitive spectrophotometric method for determination of hydrogen sulfide with 2,29-dipyridyl disulfide. Anal. Biochem. 107, 51–55.
Truesdale, V.H., 1971. A modified spectrophotometric method for the determination of ammonia (and amino acids) in natural waters, with particular reference to sea water. Analyst 96, 584–590.
Vetter, R.D., Powell, M.A., Somero, G.N., 1991. Metazoan adaptations to hydrogen sulphide. In: Bryant), C. (Ed.), Metazoan Life Without Oxygen. Chapman and Hall, London, pp. 109–128, Chapter 6.