Oseana, Volume XIV, Nomor 2 : 55 – 63, 1989 ISSN 0216–1877
BEBERAPA SEGI BIOLOGI CREPIDULA MOLLUSCA : GASTROPODA
oleh
Subagjo Soemodihardjo 1)
ABSTRACT
SOME ASPECTS OF BIOLOGY OF CREPIDULA. Members of the genus Crepidula are well known for their two unique characters. Firstly, they belong cate-gorically to the Class Gastropoda and yet their feeding habit follows that of the Bivalves, i.e. by way of a mechanism referred to as "ciliary feeding". Strangely enough, these gastropods retain their radular teeth which is absent in the bivalves. It appears that in Crepidula the radula has been functionally transformed from a rasper to a grasper. Secondly, in term of sexuality, these gastropods belong to pro-tandric hermaphrodite, which means sexually male when young and female as they grow older. To some degree, environmental factors may modify the transformation process, but the sequence of the sexual phases is irreversible.
PENDAHULUAN
Barangkali tidak berlebihan bila di-katakan bahwa genus Crepidula termasuk salah satu marga dari Gastropoda yang ba-nyak dijadikan obyek penelitian dan di-bahas. Tampaknya keistimewaan ini adalah berkat perilaku seksual dan perikehidupan-nya yang unik. Marga Crepidula terkenal karena sifat seksualnya yang dikategorikan sebagai hermaprodit protandrik yang dalam dunia binatang berarti berkelamin jantan sewaktu muda dan berubah menjadi betina menjelang usia tua. Faktor apa yang memacu terjadinya perubahan itu masih belum jelas benar.
Sampai beberapa tahun terakhir telah banyak karya-karya ilmiah yang membahas mengenai marga Crepidula ini. Sebagian besar kegiatan penelitian dan karya tulis dipusatkan pada segi perilaku seksualnya. Dalam satu seri tulisan yang cukup men-dalam, COE membahas berbagai segi kehi-dupan siput ini, meliputi antara lain : fase-fase seksual (COE 1936), pengaruh ke-lompok terhadap seksualitas (COE 1938b) dan pengaruh nutrisi terhadap seksualitas (COE 1948).
GOULD juga telah banyak mencurahkan waktunya untuk mempelajari marga Crepi-dula. Tulisan-tulisannya mencakup antara
lain : siklus seksual (GOULD 1917a), pe-ngaruh lingkungan terhadap seks (GOULD 1917b), penyebaran stimulan seks jantan melalui air (GOULD 1919) dan faktor-faktor dalam serta luar yang mempengaruhi pertumbuhan dan seksualitas (GOULD 1952). Cara makan Crepidula yang unik di-gambarkan oleh ORTON (1912a) dan YONGE (1938). MORITZ (1938) meleng-kapi pengetahuan mengenai siput ini dengan menyajikan informasi lengkap tentang ana-tomi Crepidula adunca dan YIPP (1985) mengenai anatomi organ reproduksi Crepi-dula walshi dari Hong Kong.
Pada dasarnya Crepidula tersebar di daerah dingin dan sub-tropik. Di laut tropik seperti di Indonesia hanya dijumpai kerabat dekatnya misalnya Siphopatella dan Cheilea (ROBERTS et al. 1982). Namun sifat-sifat biologinya yang unik sangat menarik dan bermanfaat untuk diketahui para peminat Moluska. Atas dasar pertimbangan itulah tulisan ini disusun dengan memanfaatkan sumber-sumber informasi yang berhasil di-peroleh.
MORFOLOGI
Cangkang Crepidula berbentuk keru-cut pendek dan melebar bagai caping bapak tani di sawah atau seperti selop (slipper) priyayi Solo. Oleh karena itu disebut "sli-pper limpet" oleh orang Inggris. Walaupun ada perbedaan bentuk antar jenis, namun sebagai kelompok marga ini mudah dikenal. Variasi intraspesifik bentuk cangkang juga teijadi, akibat dari cara hidup yang menetap sehingga harus menyesuaikan bentuk cang-kangnya dengan tempat di mana binatang tersebut melekat (ROGERS 1951). Satu ciri khas dari cangkang Crepidula ialah adanya sekat serambi (shelf) yang menutup bagian belakang lubang cangkang (shell aperture).
Bentuk spiral dari cangkangnya, yang me-rupakan ciri umum Gastropoda, sudah tidak nampak pada binatang dewasa, sehingga sepintas lalu cangkangnya kelihatan seperti simetri bilateral. Perbedaan lain ialah cang-kang Crepidula tidak dilengkapi dengan tutup cangkang (operculum).
Gambar 1 memperlihatkan cangkang Crepidula fornicata, yang merupakan jenis terbesar dari marga ini. Panjang cangkangnya berkisar antara 2 cm sampai 5 cm. Bentuk-nya agak meninggi, puncak cangkang sedikit condong ke samping-belakang. Sekat serambi berwarna putih, bentuknya sedikit cekung dengan pinggiran menggelombang (ROGERS 1951). Jenis-jenis lain mempunyai ukuran yang lebih kecil dengan warna-warna yang berbeda. Crepidula convexa, misalnya, ber-cangkang agak melengkung, berwarna gelap kemerahan, kecuali dinding-dalam cangkang dan sekat serambi yang berwarna kebiruan. C. plana, sebaliknya, mempunyai cangkang yang sedikit pipih. Kebiasaannya melekat pada dinding-dalam cangkang Gastropoda lain tidak jarang menyebabkan cangkangnya menjadi cekung (ROGERS 1951). Cangkang Crepidula adunca mencuat relatif tinggi dan puncaknya membentuk tekukan tajam.
Gambar l a. Morfologi Cangkang Crepidula forni- cata dilihat dari sisi ventral. l=puncak cangkang; 2= sekat serambi 3=lubang cangkang (shell fperture).
Gambar l b. Cangkang Crepidula fornicata dilihat dari sisi ventral.
Gambar l c. Cangkang Crepidula fornicata dilihat dari sisi dorsal.
Dinding-luar cangkang berwarna kecoklatan, sedangkan dinding-dalam, termasuk sekat serambi, berwarna putih (ROGERS 1951).
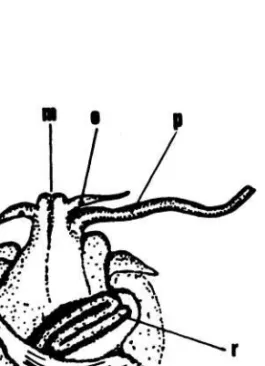
Tubuh-lunak Crepidula, atau biasanya disebut "visceral mass", menempati rongga cangkang di atas sekat serambi. Kepalanya dapat dibedakan dengan jelas dari bagian tubuh yang lain. Di kepala terdapat mon-cong (mulut), dua buah sungut (tentacles) dan mata semu (ocellus) di dasar tentakel. Pada binatang jantan, sebuah penis (phallus) yang relatif besar dan panjang mencuat keluar dari pangkal tentakel kanan (Gambar 2).
Crepidula memperoleh makanannya dengan menyaring partikel-partikel organik dari air di sekitarnya. Dengan kata lain Crepidula termasuk "ciliary feeder", Oleh karena itu binatang ini dilengkapi dengan insang yang cukup besar yang berfungsi ganda, yaitu untuk bernafas dan untuk makan. Yang dipandang ganjil ialah bahwa siput ini masih memiliki lidah parut (radula) walaupun telah menjadi "ciliary feeder". Rupanya fungsi radula pada marga ini telah berubah dari alat penggaruk menjadi alat pemungut makanannya.
Gambar 2. Tubuh-lunak Crepidula adunca (dari
MORITZ 1938).
s=sungut; m=moncong; o=ocelusi; p=penis; r=usus; i=ingsang; t=testis (gonad).
PERILAKU
Seperti telah disinggung di muka, marga Crepidula terdiri dari beberapa jenis. Mereka tersebar luas di seluruh dunia, ter-utama di daerah ugahari dan daerah dingin; menghuni daerah pasang surut sampai ke-dalaman lebih dari 30 m, dan menduduki hampir semua tipe habitat. Misalnya Crepi-dula fornicata dan C. onyx memanfaatkan habitat lumpur sebagai tempat tinggalnya. Jenis lain menempel pada batu karang, pada dinding-dalam rumah kelomang, dan sebagainya. Diduga ada kecenderungan untuk memilih tuan rumah tertentu, misal-nya C. plana umummisal-nya menempel pada rumah kelomang jenis Eupagurus bernhardus (GOULD 1917a). Crepidula muda aktif bergerak kian-kemari, tetapi yang sudali berumur dapat dikatakan menetap di satu tempat. Sering terjadi sejumlah Crepidula hidup bersama dalam satu ruangan yang sempit sehingga tidak dapat tumbuh men-capai ukurannya yang normal. Apabila dipindahkan ke ruangan yang lebih luas, mereka cepat berkembang melebihi besar semula. GOULD (1917a) menyebut gejala tersebut sebagai polimorfi lingkungan (envi-ronmental polymorphism).
Sangat menarik adalah kebiasaan me-reka untuk hidup saling melekat sehingga terjadi rangkaian siput antara 3 sampai 16 ekor (COE 1938a) (Gambar 3). Perilaku ini lebih menonjol pada jenis yang hidup di atas substrat lumpur, seperti Crepidula fornicata dan C. onyx. Siput yang paling bawah selalu merupakan yang tertua, ter-besar dan berkelamin betina; sedangkan yang ada di puncak adalah yang termuda, kecil dan berkelamin jantan. Tidak jarang antara siput terbawah dan teratas terdapat pula siput-siput muda lain yang belum ber-kembang jenis kelaminnya, ataupun yang kelaminnya dalam fase transisi.
ORTON (1912a) menegaskan bahwa rangkaian Crepidula tersebut merupakan kelompok yang tetap atau permanen. Kesimpulan itu didasarkan pada kenyataan bahwa siput yang berada paling bawah telah menjadi binatang menetap (sedentary), dalam arti tidak pernah bergerak/berpindah tempat sama sekali. Dalam satu koloni yang telah berumur cukup lama, siput yang ter-bawah mungkin bahkan telah mencengke-ramkan pinggiran cangkangnya ke dalam substrat di mana binatang itu melekat. Kesesuaian bentuk antara pinggiran cang-kang seekor siput dengan cangcang-kang siput di bawahnya, tidak dapat diartikan lain kecuali bahwa mereka telah hidup dan ber-kembang bersama-sama dalam posisi demi-kian untuk waktu yang lama. Diduga rang-kaian Crepidula seperti itu dapat berlanjut terus dari tahun ke tahun; binatang muda yang datang bergabung akan mengambil posisi di puncak, sedangkan siput-siput betina yang sudali uzur akan mati di dasar. Pemeriksaan seksama rangkaian siput akan memperlihatkan bahwa susunannya tidak berdiri tegak tetapi melengkung (Gambar 3). Hal ini adalah akibat dari keadaan bahwa
Gambar 3. Koloni Crepidula fornicata (dari ORTON 1912 b). Panah menunjuk- kan arah aliian air sewaktu koloni tersebut aktif menyaring makanan dari air di sekitarnya.
siput yang satu tidak melekat tepat di atas siput di bawahnya tetapi sedikit miring ke kanan sehingga pinggir kanan cangkang masing-masing saling berdekatan.
Semula para peneliti menduga bahwa untaian kelompok Crepidula semacam itu ada kaitannya dengan aktivitas reproduksi. Kemudian ORTON (1912b) mengemukakan pendapat yang berbeda, yaitu untaian itu lebih condong berkaitan dengan upaya mencari makan dari pada upaya reproduksi. Dasar pertimbangannya adalah bahwa siput jantan di puncak terlalu jauh untuk melak-sanakan fertilisasi. Tugas ini dilakukan oleh pejantan lain yang dapat bergerak kesana-kemari. Tumpukan Crepidula yang meleng-kung ke kanan tersebut diduga menguntung-kan semua anggota koloni. Waktu mamenguntung-kan, air masuk dari sisi kiri dan keluar melalui sisi kanan. Tumpukan cangkang yang miring ke kanan ini akan memberikan ruangan yang lebih luas bagi aliran air memasuki koloni siput-siput ini. Sebaliknya, pinggiran kanan yang saling berdekatan mempersatu-kan aliran air keluar sehingga menjadi lebih kuat dan mampu mengangkut kotoran dan sisa-sisa metabolisme koloni tersebut.
Dalam kelas Bivalvia, cara makan dengan bantuan getaran silia (ciliary feeding) adalah hal yang umum. Akan tetapi pada kelas Gastropoda ciliary feeding justru me-rupakan kekecualian. Dan Crepidula ter-masuk dalam perkecualian itu. Getaran silia pada insang dan mantel menggerakkan air masuk dan keluar rongga mantel melalui insang. Partikel-partikel yang tersaring insang dibalut dengan lendir dan oleh gerakan silia didorong ke arah kepala dan dijatuhkan di dekat mulut. Dengan radulanya yang telah berubah fungsi itu butiran lendir berisi par-tikel itu dipungut dan ditarik ke mulut.
SEKSUALTTAS
Aspek paling populer dalam per ke-hidupan Crepidula adalah keke-hidupan seksual-nya. Sebagaimana telah disinggung di muka, jenis-jenis Crepidula termasuk dalam kelom-pok organisme yang disebut hermaprodit protandrik. Ada tiga hal yang menarik untuk dibahas yaitu fase seksual, pengaruh asosiasi terhadap seksualitas dan kecenderungan sek-sual.
Fase Seksual
Pada Crepidula yang masih belia, gonadnya bersifat biseksual (COE 1936), dalam arti gonad tersebut berisi baik bibit gamet jantan maupun betina. Menurut GOULD (1952) Crepidula muda yang panjangnya antara 4 mm – 5 mm masih belum mempunyai jenis kelamin atau netral (neuter). Dengan bertambahnya umur, gatra gonad (primary gonad) jantan mulai berkembang, sedangkan gatra gonad betina tetap tinggal diam (inert). Dalam fase lanjut usia, sistem reproduksi jantan akan meng-alami transformasi total menjadi sistem reproduksi betina. Pada saat itu gatra gonad betina yang semula diam, mulai aktif ber-kembang.
Dari uraian di atas dapat disimpulkan bahwa Crepidula muda pertama-tama akan berkembang menjadi siput jantan dan ber-fungsi sebagai pejantan sampai beberapa lama, biasanya hampir setahun. Kemudian setelah melalui masa transisi, siput tadi berubah menjadi betina dan tetap menjadi betina selama sisa hidupnya (COE 1947). Sebelum mencapai fase jantan, Crepidula betul-betul tidak berjenis kelamin dan tidak memperlihatkan tanda-tanda seksual sekun-der. Akan tetapi fase jantan biasanya dicapai
dalam waktu tidak lama setelah binatang itu menetas dari telur. Tidak berapa lama setelah menetas alat-alat kelengkapan seksual sekunder jantan mulai berkembang dan binatang tersebut menjadi jantan yang fungsional. Proses perkembangan semacam itu menyebabkan siput jantan selalu ber-ukuran kecil dibandingkan dengan yang betina. Menurut ROGERS (1951) Crepidula betina dapat 15 kali lebih besar dari yang jantan.
Fase jantan sangat mudah dikenal secara visual dari adanya alat kelamin jantan (phalus) yang besar dan panjang, men-cuat dari pangkal sungut kanan (Gambar 2). Pada fase transisi, sisa-sisa gonad jantan dan isinya diserap kembali dan palusnya meng-alami degenerasi. Gatra gonad betina segera mengalami proliferasi dan berkembang men-jadi gonad betina menggantikan gonad jantan. Saluran telur (oviduct), uterus dan asesori sistem reproduksi lainnya pun terbentuk (COE 1947). Pertumbuhan yang cepat umumnya menyertai proses transfor-masi. Sejalan dengan perubahan tubuhnya, berubah pula proses fisiologinya. Yang disebut terakhir ini perlu untuk mengatur perilaku dan naluri seksualnya. Sampai batas-batas tertentu faktor lingkungan mem-punyai peran dalam proses metamorfose seksual tersebut, namun urutan fasenya tidak dapat dibalikkan (COE 1947).
Pengaruh Asosiasi terhadap Fase Seksual. GOULD (1919) berpendapat bahwa asosiasi dengan siput betina diperlukan untuk merangsang siput muda berkembang menjadi jantan. Disebutkan bahwa fase jantan tidak akan terjadi pada Crepidula muda yang dipisahkan dari koloninya. Dengan kata lain ketiadaan siput betina menyebabkan siput muda yang masih ne-tral berkembang langsung menjadi siput
betina. Selanjutnya GOULD berhipotesa bahwa siput betina mengeluarkan zat yang dapat merangsang siput netral menjadi jantan.
Hasil percobaan kemudian memlihatkan bahwa pendapat GOULD yang per-tama tidak seluruhnya benar. Mengenai hipotesa kedua mula-mula diragukan kebe-narannya, tetapi kemudian didukung oleh COE (dalam FRETTER & GRAHAM 1961).
COE (1938a) mengisolasi sejumlah Crepidula muda dan memeliharanya di akuarium secara terpisah-pisah. Kondisi ling-kungan dalam akuarium diusahakan agar sebanyak mungkin mirip dengan lingkungan alami. Sebagai kontrol, sejumlah Crepidula lainnya yang dipisahkan dalam tabung-tabung dimasukkan dalam laut. Setelah 6 pekan, hasilnya memperlihatkan bahwa 30 % dari binatang percobaan telah berkembang menjadi siput jantan, sedang pada kelompok kontrol 46 %. GOULD (1952) melakukan percobaan yang sama, tetapi binatang per-cobaan tidak dipisah-pisahkan. Hasilnya ter-nyata hampir sama dengan eksperimen yang dilakukan oleh COE di atas. Atas dasar hasil percobaan tersebut, GOULD me-nyimpulkan bahwa komunitas Crepidula muda yang terdiri dari individu-individu yang masih netral, pada akhirnya (asalkan diberi waktu yang cukup) akan membentuk komunitas yang terdiri dari jantan, betina dan fase transisi dalam rasio yang hampir sama. Menurut WILCZYNSKI (1955), rasio tersebut mendekati 60 % jantan, 35 % betina dan 5 % transisi jantan-betina.
Hasil percobaan di atas memperlihat-kan bahwa kehadiran siput betina tidak mut-lak untuk perkembangan siput jantan, namun berdampak positif. Di lain pihak hasil percobaan menunjukkan bahwa dalam keadaan tertentu fase jantan dapat ditinggal-kan. Bahwa kehadiran siput betina
mendo-rong perkembangan siput jantan, hal ini juga ditunjukkan oleh percobaan GOULD (1952). GOULD memisahkan Crepidula jan-tan dari koloninya dan mengisolasikannya sehingga penis mengalami degenerasi. Ketika siput yang bersangkutan dimasukkan kem-bali ke koloninya, penisnya segera tumbuh kembali.
Kecenderungan Seks.
Kendatipun dalam kondisi alami semua individu mengalami ketiga fase seksual tersebut, namun masih juga terlihat sifat-sifat individualistis untuk cenderung ke arah jantan atau betina, sehingga dalam satu koloni dapat dibedakan individu tipe jantan dan individu tipe betina (COE 1936). Tipe
betina adalah individu-individu yang fase jantannya sangat singkat, sedangkan tipe jantan adalah mereka yang fase jantannya panjang (satu tahun atau lebih) walaupun pada akhirnya juga berubah menjadi betina. Gambar 4 memperlihatkan tanggapan sek-sual (sexual response) dari kedua tipe seks tersebut dalam situasi lingkungan yang ber-beda-beda.
Menurut GOULD (1952), kebanyakan binatang muda tipe betina cepat tanggap terhadap rangsangan jantan, tetapi masih lebih lambat dari tanggapan tipe jantan terhadap rangsangan betina. Di antara tipe jantan terdapat pula yang termasuk jantan sejati, dalam arti cenderung tetap berkelamin jantan (COE 1948). Akan tetapi Crepidula tipe ini jumlahnya kecil.
REPRODUKSI
Musim memijah Crepidula bervariasi dari jenis yang satu ke jenis lainnya. Jenis-jenis dari pantai Amerika Serikat memijah mulai bulan April atau Mei dan berlanjut sampai Nopember (ORTON 1912a). Bebe-rapa jenis lainnya seperti C. adunca dan C. norrisiarum, diduga memijah sepanjang tahun (MORITZ 1939; MAC GINITIE dan MAC GINITIE 1964).
Fertilisasi pada jenis-jenis Crepidula terjadi secara internal. Waktu berkopulasi binatang jantan menempel di atas pinggir kanan cangkang yang betina. Sering terjadi lebih dari seekor jantan bersama-sama membuahi seekor betina.
Pembentukan Gamet
GAULD (1917a) menyatakan ada dua macam bentuk cikal-bakal sel gamet pada Crepidula muda, yaitu yang disebutnya sel gamet primordial tipe A dan sel gamet primordial tipe B. Yang disebut pertama adalah primordial sel telur, sedangkan yang disebut kedua primordial sel sperma. Per-bedaannya ialah sel tipe A mempunyai inti transparan dengan bahan kromatin yang menyerap warna dengan kuat, sedangkan sel tipe B mempunyai inti retikular dan jaringan kromatinnya tidak kentara.
Pada permulaan fase jantan, sel-sel tipe B memperbanyak diri dengan cepat melalui pembelahan mitosis, menghasilkan sejumlah besar sel-sel bibit yang lepas dan berkumpul di rongga-rongga khusus (lumen). Sel-sel bibit ini merupakan cikal-bakal sperma, oleh karena itu disebut spermato-gonia. Spermatogonia terus memperbanyak diri sehingga lumen-lumen menjadi penuh dan membengkak. Kemudian pembelahan mitosis berhenti dan diganti pembelahan meiosis yang menghasilkan sel-sel haploid spermatosit primer.
Ada dua spermatosit primer yaitu yang berinti besar dengan sitoplasma tipis dan yang berinti kecil dengan sitoplasma tebal. Hanya yang disebut pertama yang akan menjadi sperma fungsional. Apa tugas spermatosit bentuk kedua, belum diketahui dengan pasti.
Pembentukan sel gamet betina tidak banyak berbeda dengan pembentukan sel gamet jantan, kecuali bahwa ovogonia tidak melepaskan diri dan memperbanyak diri dalam lumen, tetapi tetap melekat pada dinding gonad.
PERKEMBANGAN LARVA Segera setelah kopulasi, Crepidula be-tina mulai bertelur. Telurnya disimpan dalam kapsul-kapsul bulat, masing-masing berisi sampai 250 butir telur. Seekor betina dapat menghasilkan 50 sampai 70 kapsul (ORTON 1912a). Kapsul-kapsul tersebut dilekatkan dengan tangkai pada substrat dan dierami di bawah kaki induknya sampai menetas.
Semua jenis Crepidula, kecuali C. adunca, mengembangkan larva veliger plank-tonik. Anak C adunca langsung merayap seperti siput dewasa waktu menetas (MO-RITZ 1939). Cangkang larva veliger plank-tonik sudah berbentuk spiral dan velumnya besar membentuk dua belahan (bilobed). Masa inkubasi telur dapat sampai 4 pekan. Setelah menetas larva veliger hidup sebagai plankton selama 14 hari, kemudian turun ke dasar dan bermetamorfose menjadi Crepidula kecil.
DAFTAR PUSTAKA
COE, W.R. 1936. Sexual phases in Crepi-dula. J. Exp. Zool. Philadelphia : 72 : 455 – 477.
COE, W.R. 1938a. Influence of association on the sexual phases of Gastropods having protandric consecutive sexuality. Biol. Bull. Woods Hole : 75 : 274 – 285. COE, W.R. 1938b. Condition influencing
change of sex in mollusks of the genus Crepidula. J. Exp. Zool. Philadelphia : 77 : 401 – 424.
COE, E.R. 1947. Biology of Crepidula williamsi, a new species of prosobranch gastropod from the Pacific coast. J. Morph. Philadelphia : 81 : 241 – 248. COE. W.R. 1948. Nutrition, environmental
conditions and growth of marine bivalve mollusks. J. Mar. Res. New Haven : 7 ( 3 ) : 5 8 6 – 6 0 1 .
FRETTER, V. and A. GRAHAM 1962. British prosobranch molluscs. Their functional anatomy and ecology. Ray Society, London. GOULD, H.N. 1917a. Studies on sex in the
hermaphrodite mollusc Crepidula plana I. History of the sexual cycle. J. Exp. Zool. Philadelphia : 23 : 1 - 68.
GOULD, H.N. 1917b. Studies on sex in the hermaphrodite mollusc Crepidula plana II. Influence of environment on sex. J. Exp. Zool. Philadelphia : 23 : 225 – 250.
GOULD, H.N. 1919. Studies on sex in the hermaphrodite mollusc Crepidula plana III. Transferens of the male-producing stimulus through sea water. J. Exp. Zool. Philadelphia : 29 : 113 – 120. GOULD, H.N. 1952. Studies on sex in the
hermaphrodite mollusk Crepidula plana TV. Internal and external factors influencing growth and sex development. J. Exp. Zool. Philadelphia : 119 : 93 - 163. MAC GINITIE, N and G.E. MAC GINITIE
1964. Habitats and breeding seasons of the shelf limpet Crepidula norrisiarum Williamson. Veliger 7 : 34.
MORITZ, C.E. 1938. The anatomy of the gastropod Crepidula adunca Sowerby. Univ. Calif. Publ. Zool. 435 : 83 – 92.
MORITZ, C.E. 1939. Organogenesis in the gastropod Crepidula adunca Sowerby. Univ. Calif. Publ. Zool. 43 : 217 – 248.
ORTON, J.H. 1912a. The mode of feeding of Crepidula with an account of the current-producing mechanism in the man- tle cavity, and some remarks on the mode of feeding in gastropods and lamelli-branchs. J. Mar. Biol. Ass. Plymouth 9 : 444 – 478.
ORTON, J.H. 1912b. An account of the natural history of the slipper limpet (Crepidula fornicata) with some remarks on its occurrence on the oyster grounds of the Essex coasts. J. Mar. Biol. Ass. Plymouth 9 : 437 – 443.
ROBERTS, D., S. SOEMODIHARDJO and W. KASTORO 1982. Shallow water marine molluscs of North–West Java. Lembaga Oseanologi Nasional–LIPI, Jakarta.
ROGERS, J.E. 1951. The shell book. A popular guide to a knowledge of the families of living mollusks and an aid to identification of shells native and foreign. Boston, Mass. C.T. Bradford Co.
YEPP, M.W. 1985. The anatomy of the organs of reproduction of Crepidula walshi (Mollusca : Gastropoda). In The Malacofauna of Hong Kong and Southern China. 2. (Morton and Dudgeon, Eds) Proceedings of the second International Workshop on the Malacofauna of Hong Kong. University Press, Hong Kong : 245 – 256.
YONGE, CM. 1938. Evaluation of ciliary feeding in the Prosobranchia, with an account of feeding in Capulus ungaricus. J. Mar. Biol. Ass. Plymouth : 222 : 453 – 468.
WILCZYNSKI, J.C. 1955. On sex behaviour and sex determination in Crepidula for-nicata. Biol. Bull. Woods Hole 109 : 353 – 354.