Penyakit Akar Gada Gejala
Gejala yang nampak di atas perrnukaan tanah adalah daun-daun tanaman yang terinfeksi P. brassicae layu pada hari panas dan kering
,
pulih kembali selama malam hari, dan kelihatan normal dan segar pada pagi hari. Selanjutnya jika penyakit berkembang terus, daun-daun menjadi kuning, tanaman kerdil dan mungkin mati atau hidup dalam keadaan merana selama musim tanam (Karling 1968). Menurut Agrios (1997) bibit cruciferae yang terinfeksi ketika masih muda dapat mati dalam beberapa minggu, bahkan mungkin mati segera setelah infeksi patogen ini. Infeksi pada tanaman yang lebih tua kemungkinan masih aktif tetapi tanaman menjadi kerdil dan gaga1 membentuk krop sehingga produksi menurun atau tidak berproduksi sama sekali.Pembengkakan akar merupakan ciri khas penyakit akar gada (Gambar 1). Bentuk dan letak tergantung pada spesies inang dan tingkat infeksi. Pada Brassica oleracea mula-mula pembengkakan berbentuk "spindel" (kurus panjang) yang sangat kecil pada akar-akar utama dan lateral. Dengan pertumbuhan jaringan inang yang tidak terkendali, akar-akar menjadi sangat besar dan berubah bentuk, dan akhirnya bersatu membentuk gada (Channon dan Maude 197 1). Makin lama akar yang membengkak makin besar dan biasanya hancur sebelum akhir musim tanam karena serangan bakteri dan cendawan lain (Agrios 1997). Pada akar kubis, pembengkakan dapat mencapai ukuran kepalan tinju manusia dan warnanya nampak kelabu dan kuning pucat (Karling 1968). Warna akar yang bengkak kekuning-kuningan berbeda dengan akar-akar yang sehat berwarna putih
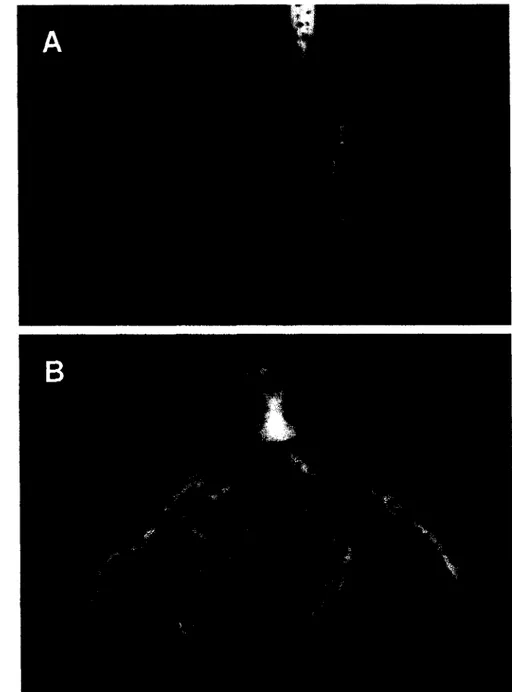
Gambar 1.
Akar
tanaman caisin yang sehat atau tanpa gejala penyakit akar gadaP. brassicae
(A) dan yang sakit karena infeksiP. brassicae
(B),
umur 53 hari setelah semai.(Graveland et al. 1992). Apabila infeksi terjadi pada akhir musim tanam, ukuran gada biasanya kecil dan tanaman dapat bertahan hidup (Karling 1968).
Patogen
Penyakit akar gada disebabkan oleh Plasmodiophora brassicae Woronin. Alexopoulos et al. (1996) menggolongkan patogen ini ke dalam:
Kingdom : Protista Phylum : Plasmodiophoromycota Kelas : Plasmodiophoromycetes Ordo : Plasmodiophorales Famili : Plasmodiophoraceae. Genus : Plasmodiophora
Spesies : Plasmodiophora brassicae Wor.
Menurut Agrios (1997), P. brassicae dianggap sebagai "Pseudofungi" atau organisme yang menyerupai fungi dan diklasifikasikan sebagai berikut:
Kingdom : Protozoa Phylum : P1asmodiophoromycota Kelas : Plasmodiophoromycetes Ordo : Plasmodiophorales Famili : Plasmodiophoraceae Genus : Plasmodiophora
Spesies : Plasmodiophora brassicae Wor.
Plasmodiophora brassicae merupakan endoparasit obligat pada jaringan tanaman dan dapat membentuk struktur bertahan berupa spora rehat yang dapat terlepas masuk ke dalam tanah (Agrios 1997).
8 Siklus penyakit dimulai dengan perkecambahan satu zoospora primer dari satu spora rehat haploid di dalam tanah. Zoospora primer ini mempenetrasi rambut akar dan menginjeksi isi sel masuk ke dalam sel inang (Aist dan Williams 1971 dalam Voorrips 1995). Setelah penetrasi rambut akar atau sel epidermis inang oleh zoospora primer, protoplas yang berinti satu terbawa masuk ke dalam sel inang. Pembelahan mitotik terjadi dan protoplas membentuk plasmodium primer. Setelah plasmodium primer mencapai ukusan tertentu, tergantung pada ukuran sel epidermis inang, membelah menjadi beberapa bagian yang berkembang menjadi zoosporangia (Alexopoulos et al. 1996). Setiap zoosporangium mengandung 4 atau 8 zoospora sekunder yang dapat terlepas melalui lubang atau pori-pori pada dinding sel inang (Agrios 1997), apakah langsung masuk ke dalam sel inang yang lain atau keluar akar (Alexopoulos et al. 1996). Naiki et al. (1984) dalam Voorrips (1995) menunjukkan bahwa zoospora sekunder dapat menginfeksi kembali rambut akar, menyebabkan perkembangan aseksual patogen yang cepat. Menurut Agrios (1997) beberapa zoospora sekunder bergabung berpasangan menghasilkan zigot yang dapat menyebabkan infeksi baru dan menghasilkan plasmodium baru. Plasmodium ini mempenetrasi langsung jaringan akar yang muda, sedangkan akar-akar yang tua, akar-akar yang tebal, dan pangkal batang dipenetrasi melalui luka-luka. Dari tempat infeksi plasmodium menyebar ke korteks dan kambium dengan penetrasi langsung, kemudian plasmodium menyebar ke seluruh bagian kambium dan menuju xilem. Plasmodium berada di dalam beberapa sel menstimulir pembelahan dan pembesaran sel secara tidak normal. Menurut Dekhuijzen dan Overeen (1971), Dekhuijzen (1980) dalarn Voorrips (1995) penyebaran patogen terjadi terutama
karena distimulasi dengan pembelahan sel inang yang diduga distimulir oleh konsentrasi sitokinin dan auksin yang tinggi. Perkembangan selanjutnya, inti yang haploid dalam plasmodia yang berinti banyak bergabung berpasangan (Tommerup dan Ingram 197 1 dalam Voorrips 1995). Setelah miosis terbentuk inti diploid yang baru yang kemudian berkembang menjadi spora rehat haploid yang terlepas masuk ke dalam tanah ketika akar yang sakit rusak (Voorrips 1995).
Penyebaran Penyakit dan Virulensi Patogen
Penyakit akar gada (clubroot) yang disebabkan oleh P. brassicae menyebar hampir di seluruh pertanaman cruciferae yang ada di dunia (Karling 1968). Di Indonesia penyakit ini telah tersebar di Sumatera Utara, Jawa Tengah, dan terutama di Jawa Barat (Djatnika 1984).
Akar-akar yang busuk dari tanaman sakit sebelumnya mengandung sejumlah besar spora rehat P. brassicae yang merupakan sumber inokulum satu- satunya untuk penyakit akar gada (Orihara dan Yamamoto 1998). Apabila akar- akar yang sakit hancur di lahan
,
maka spora-spora terlepas masuk ke dalam tanah dan bertahan hidup dalam waktu yang lama (Karling 1968; Voorrips 1995). Kemampuan bergerak bentuk amuba dan zoospora patogen merupakan faktor penting dalam penyebaran penyakit, walaupun jarang berpindah lebih dari 5 inci secara horizontal selama satu musim (Chupp dalam Karling 1968). Spora- spora yang berkecambah dan menginfeksi akar-akar muda dari cruciferae kemudian tersebar melalui akar-akar yang sakit pada saat pemindahan tanaman tersebut ke lapangan (Walker 1975).Penyakit akar gada dapat terpencar di alam melalui tanah dengan berbagai cara atau perantara, misalnya transportasi tanah yang terinfestasi P. brassicae
10 melalui perlengkapan usahatani, sepatu pekerja, kuda dan kaki-kaki ternak merupakan penyebaran penyakit yang efektif (Karling 1968); melalui butiran tanah yang terbawa hasil panen (Walker 1975); melalui air permukaan (Stakman dan Harrar 1957); penyebaran penyakit melalui angin juga efektif walaupun tidak universal (Chupp dalam Karling 1968); melalui pupuk kotoran hewan (Karling 1968; Suryaningsih 198 1). Patogen juga dapat ditularkan oleh biji melalui kontaminasi permukaan dengan tanah yang terinfeksi. Selain itu sejumlah tanaman cruciferae liar dan beberapa tanaman inang lain yang rentan terhadap penyakit akar gada dapat menjadi tempat bertahan hidup patogen pada saat tanaman budidaya tidak ada (Karling 1968).
Menurut Karling (1968) dari 61 genera cruciferae, lebih dari 300 spesies dan varietas rentan terhadap P. brassicae dan 9 spesies dari genera bukan cruciferae ditemukan mengandung zoosporangia P. brassicae pada rambut- rambut akarnya. Reyes et al. (1974) melaporkan 9 jenis gulma dari cruciferae rentan terhadap P. brassicae ras 6. Di beberapa lahan pertanaman kubis-kubisan di Jawa Barat ditemukan 4 ras P. brassicae (Djatnika 1989).
Pengaruh Faktor Lingkungan
Penyakit akar gada berkembang dengan baik pada pH tanah 5,7, menurun dengan tajam pada pH tanah antara 5,7 dan 6,2 dan gaga1 berkembang pada pH tanah 7,8 (Chupp dalam Stakrnan dan Harrar 1957). Menurut Karling (1968) perkecambahan spora P. brassicae terjadi pada pH 5,4 - 7,5 dan tidak berkecambah pada pH 8, tetapi pH tanah yang rendah tidak menjamin te rjadinya infeksi untuk semua keadaan (Mattusch 1977).
Kisaran temperatur optimum bagi perkembangan P. brassicae adalah 17,8 - 25 OC dengan temperatur minimum dan maksimum 12,2 OC dan 27,2 OC (Agrios 1997; Walker 1957; Channan dan Maude 1971). Menurut Chupp dalam Stakman dan Harrar (1 957), spora-spora P. brassicae berkecambah pada temperatur kamar dalam media agar yang mengandung bibit kubis yang muda, tetapi spora-spora tersebut tidak berkecambah atau berkecambah sangat buruk pada kondisi yang sama tanpa bibit kubis.
Kelembaban optimum selama 18 - 24 jam memungkinkan perkecambahan dan penetrasi patogen ke dalam jaringan inang (Wellman dalam Stakman dan Harrar 1957), kemudian infeksi hanya terjadi jika kelembaban tanah di atas 45 % (Monteith dalam Stakman dan Harrar 1957) dan kelembaban tanah 50 % atau lebih menyebabkan perkembangan penyakit bertambah cepat, sedangkan kelembaban tanah 45 % dapat menghambat infeksi (Mattusch 1977).
Tanaman inang yang berada pada lingkungan yang intensitas cahayanya rendah lebih tahan terhadap serangan P. brassicae dibanding dengan tanaman inang yang berada pada lingkungan yang intensitas cbhayanya tinggi (Garret 1970 dalaln Djatnika 1989).
Jumlah spora rehat akan menentukan tingkat infeksi pada inangnya. Suspensi yang mengandung paling sedikit lo6 - lo8 sel spora setiap ml sangat efektif untuk mengadakan infeksi (Mattusch 1977), sedangkan berdasarkan penelitian Djatnika (1989) lo4 sel spora masih mampu menginfeksi tanaman. Disamping itu kondisi inang turut mempengaruhi perkembangan P. brassicae, seperti kisaran inang, inang yang rentan, dan morfologi dari sistem perakaran (Mattusch 1977), serta peranan mikrob yang lain seperti cendawan, bakteri, virus
dan nematoda. Mikrob tersebut dapat bersifat antagonistik, sinergistik atau tidak mempengaruhi aktivitas patogen.
Pengendalian
Beberapa upaya pengendalian yang telah dilakukan, baik di Indonesia maupun di luar negeri, antara lain pengapuran tanah (Karling 1968; Dobson et al. 1983; Djatnika 1989; Herdian 2000); rotasi tanaman (Karling 1968); perendaman lahan (Djatnika 1989); penggunaan fungisida (Lambe dan McCart 1975; Rowe dan Farley 1979; Djatnika 1989); penggunaan mulsa jagung (Djatnika 1989; Herdian 2000); penggunaan ekstrak bawang putih (Djatnika 1989); pemanfaatan mikrob antagonis (Murtafingah 1987; Djatnika 1989; Djatnika 1990; Widodo et al. 1993); varietas resisten (Rowe 1980; Yamagishi 1987); dan solarisasi tanah (Horiuchi et al. 1982; Widodo dan Suheri 1995).
Pengaturan kemasaman tanah dengan kapur hingga mencapai pH di atas netral (pH 7,2) pada beberapa kasus memberikan pengendalian yang baik tetapi pada kasus yang lain tidak efektif (Karling 1968). Menurut Colhoun (1953) tingkat inokulum P. brassicae yang sangat tinggi mendukung te rjadinya infeksi di dalam tanah dengan pH 8. Myers et al. (1981) dan Horiuchi dan Hori (1980) melaporkan bahwa pengapuran pada tipe tanah yang berbeda memberikan tingkat pengendalian penyakit yang berbeda. Pengapuran tanah dipengaruhi oleh distribusi atau redistribusi kapur di dalam tanah (Dobson et al. 1983), juga ada kemungkinan ras-ras P. brassicae yang berbeda bereaksi secara berbeda terhadap pH tanah (Stakman dan Harrar 1957).
Upaya pemuliaan tanaman untuk memperoleh varietas yang resisten perkembangannya lambat (Dobson et al. 1983). Selain itu varietas resisten dapat
kehilangan sifat resistensinya atau varietas tersebut dipatahkan resistensinya oleh perkembangan ras-ras fisiologi patogen (Reyes et al. 1974; Seaman et al. 1963; Yoshikawa dan Buczacki 1978). Rotasi tanaman dengan tanaman selain famili Brassicaceae memerlukan waktu yang lama karena spora rehat patogen dapat bertahan di dalam tanah lebih dari 7 tahun (Karling 1968; Channon dan Maude
1971).
Perbaikan drainase tanah dapat mengurangi kehilangan hasil tetapi tidak memberikan pengendalian yang dapat diandalkan, khususnya selama periode curah hujan yang berlebihan (Colhoun 1958; Rowe dan Farley 1979). Pengendalian dengan cara perendaman lahan hanya dapat dilakukan pada lahan sawah (Djatnika 1989).
Penggunaan pestisida seperti Benlate tidak memberikan pengendalian yang cukup memuaskan apabila diaplikasikan hanya satu kali dan Vorlex tidak praktis digunakan untuk pengendalian di lapangan (Reyes et al. 1974). Menurut Horiuchi et al. (1982) penggunaan pestisida sulit diterapkan pada lahan yang ditanami dengan famili Brassicaceae stcara terus-menerus. Walaupun Flusulfamida dapat menghambat perkecambahan spora rehat P. brassicae, tetapi tidak efektif mengendalikan P. brassicae yang sudah ada dalam sel korteks (Tanaka et al. 1999).
Penggunaan mikrob antagonis dapat menekan serangan P. brassibcae tetapi tidak menunjukkan pengaruh yang nyata terhadap bobot basah tanaman (Widodo et al. 1993). Solarisasi tanah selama 5 - 7 minggu dapat menekan tingkat kejadian dan indeks penyakit akar gada serta meningkatkan produksi tanaman kubis. Namun demikian solarisasi tanah masih perlu dikaji karena untuk
14 penerapan di lapangan biayanya cukup mahal sehingga kemungkinan penggunaannya tidak efisien (Widodo dan Suheri 1995).
Peran Tanaman Perangkap
Tanaman perangkap telah banyak digunakan dalam pengelolaan serangga hama tanaman. Tanaman perangkap juga digunakan dalam pengelolaan penyakit tanaman walaupun masih sangat sedikit dan dalam cara yang berbeda (Agrios
1997).
Tanaman perangkap yang bukan inang sesungguhnya ditanam dengan tujuan membuat patogen-patogen tular tanah menjadi lemah potensi infeksinya (Palti 198 1). Ciri khas dari tanaman perangkap adalah mampu menstimulir perkecambahan biji tanaman parasit, dapat menyebabkan perkecambahan propagul cendawan yang dorman, atau dapat menstimulir penetasan telur-telur nematoda dalam bentuk kiste (Parbery dan Morgan 1980). Pengendalian nematoda puru akar pada perkebunan nenas berhasil dilakukan dengan menanam tanaman tomat sebagai tanaman perangkap dan kemudian tanaman tersebut dimusnahkan sebelum nematoda ini dapat memproduksi telur (Stevens 1960
dalarn Palti 1981).
Pada prinsipnya penggunaan tanaman perangkap dapat merangsang perkecambahan spora rehat patogen di dalam tanah dan kemudian tanaman tersebut dipanenldimusnahkan sebelum siklus hidup patogen sempurna (Parbery dan Morgan 1980). Dengan demikian dapat mengurangi populasi patogen di dalam tanah sebelum tanaman utama ditanam.
Peran Bahan Organik
Bahan organik tanah dapat mempengaruhi sifat-sifat tanah seperti laju infiltrasi, kepadatan, stabilitas agregat, kapasitas tukar kation dan aktivitas biologi, dimana sifat-sifat ini berhubungan dengan fungsi-fungsi tanah yang penting (Sikora dan Stott 1996 dalam Gaskell et al. 2000). Bahan organik menstimulir pertumbuhan akar tanaman, meningkatkan ketersediaan hara, mengurangi evaporasi tanah, meningkatkan kapasitas menahan air, mengurangi aliran permukaan, memperbaiki drainase, mengatur temperatur tanah, dan menyediakan substrat yang berlimpah bagi mikrob tanah (Chen et al. 1987; Chen et al. 1988).
Penambahan berbagai jenis bahan organik (pupuk hijau, pupuk kandang, sisa-sisa tanaman atau sampah organik) yang dapat menyebabkan perubahan keseimbangan mikroflora tanah telah diakui sebagai suatu pendekatan biologi yang memberikan harapan dalam perbaikan pengelolaan penyakit tular tanah (William 1970). Hal yang sama dikemukakan oleh Gaskell et al. (2000) bahwa meningkatnya bahan organik di dalam tanah dapat menyebabkan populasi organisme tanah menjadi lebih banyak dan lebih beragam dan selanjutnya diduga dapat meningkatkan pengendalian hama dan penyakit secara biologi. Jenis substrat yang spesifik yang terdapat di dalam tanah secara nyata mempengaruhi jenis dan jumlah organisme yang akan menetap dan berkembang di dalam tanah
tersebut (William 1970).
Peran Solarisasi
Solarisasi tanah, sebagai proses peningkatan panas di dalam tanah di bawah lembaran plastik transparan hingga temperatur yang merugikan patogen, telah berhasil mengendalikan berbagai jenis penyakit (Stapleton dan DeVay
16 1986). Greenberger et al. (1987) membuktikan bahwa tanah-tanah yang disolarisasi lebih bersifat supresif terhadap patogen-patogen tular tanah tertentu dari pada tanah yang tidak disolarisasi. Menurut Horiuchi et al. (1982) solarisasi tanah menunjukkan efek penekanan terhadap penyakit akar gada. Walaupun mereka menduga bahwa tidak ada kaitannya antara penekanan penyakit akar gada dengan peranan mikroorganisme antagonis karena solarisasi. Peneliti lain melaporkan adanya korelasi positip antara solarisasi tanah dengan peranan mikroorganisme antagonis (Gamliel dan Katan 199 1). Widodo dan Suheri (1 995) menduga bahwa tejadinya penekanan penyakit akar gada pada kubis dengan solarisasi tanah bukan merupakan pengaruh langsung dari peningkatan suhu akibat solarisasi, melainkan karena adanya perubahan populasi mikrob tanah terutama aktinomisetes yang umumnya berpotensi sebagai antagonis terhadap patogen tanaman. Disamping itu, solarisasi tanah mempunyai efek jangka panjang (Katan et al. 1983).