ISOLASI, IDENTIFIKASI DAN KARAKTERISASI
MOLEKULAR VIRUS HEPATITIS B PADA
OWA JAWA (Hylobates moloch) DI INDONESIA
RACHMITASARI NOVIANA
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2012
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Isolasi, Identifikasi dan Karakterisasi
Molekular Virus Hepatitis B pada Owa Jawa (Hylobates moloch) di Indonesia adalah karya saya dengan arahan dari komisi pembimbing dan belum
diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Februari 2012
Rachmitasari Noviana NRP. P053090031
ABSTRACT
RACHMITASARI NOVIANA. Isolation, Identification and Molecular Characterization of Hepatitis B Virus in Javan Gibbon in Indonesia. Under direction of JOKO PAMUNGKAS and DIAH ISKANDRIATI
Hepatitis B virus (HBV) was reported not only able to infect human being, but also non-human primates, especially the great apes group such as orangutans, gorillas, chimpanzees, as well as the lesser apes from the family of Hylobatidae. Among other species within the genus, Hylobates moloch (owa jawa or silvery/ javan gibbons), which is considered as one of Indonesia’s endemic endangered species, has been reported to harbor HBV of their own strain. Analyses of HBV isolated from infected gibbons were done in this study to reveal their relatedness to the published data. Plasma samples from nine javan gibbons were obtained as part of diagnostic purpose. Those animals were previously tested positive for the presence of hepatisis B-surface antigen (HBsAg) by enzyme-linked immunosorbent assay (ELISA). DNA samples were extracted from these nine plasma samples to be used as templates for amplification of the Pre-S1 region by polymerase chain reactions (PCR) technique using semi degenerate primers that could amplify several strains of HBVs. The PCR products of 457 - 459 base pairs were then subject to restriction fragment length polymorphism (RFLP) analyses using Bst2UI enzyme to evaluate the pattern of cleaved fragments. The PCR products were sequenced for further homology analyses and to create phylogenetic tree of the sequenced data obtained from the study in comparison with published data. Two plasma samples were used as templates for whole region amplification using sets of 4 primer. The PCR products of 3192 base pairs were sequenced for further homology analyses and to create phylogenetic tree. Phylogenetic tree analyses for Pre-S1 nucleotide region and whole region showed that the javan gibbon isolates formed their own clusters, separate from the other nonhuman primate isolates.
Keywords : Hepatitis B virus, Pre-S1, whole genome, javan gibbon, PCR, Phylogenetic analysis
RINGKASAN
RACHMITASARI NOVIANA. Isolasi, Identifikasi dan Karakterisasi Molekular Virus Hepatitis B pada Owa Jawa (Hylobates moloch) di Indonesia. Dibimbing oleh JOKO PAMUNGKAS dan DIAH ISKANDRIATI.
Hepatitis atau peradangan hati adalah suatu kondisi klinis akibat terjadinya peradangan atau inflamasi pada organ ataupun jaringan hati yang ditunjukkan dengan ditemukannya sel-sel inflamatori pada jaringan hati tersebut. Infeksi virus hepatitis merupakan salah satu penyebab terjadinya peradangan hati selain adanya penyebab non infeksius seperti penggunaan obat-obatan terlarang dan minuman beralkohol. Menurut data organisasi kesehatan dunia (WHO, 2011), satu dari tiga penduduk dunia telah terinfeksi virus ini sementara satu dari 20 penduduk dunia hidup dengan infeksi kronis. Infeksi virus hepatitis yang bersifat kronis dapat mengakibatkan kematian karena menyebabkan sirosis hati.
Virus hepatitis B, salah satu dari 5 virus penyebab penyakit hepatitis, dapat ditularkan melalui darah, hubungan kelamin (sexual intercourse) dan perinatal (mother to child) pada saat melahirkan atau menyusui. Transmisi perinatal dan infeksi carrier yang bersifat jangka panjang menyebabkan adanya endemisitas dan prevalensi tinggi di benua Asia terutama bagian selatan dan timur. Selain dapat menyerang manusia, virus hepatitis B juga diketahui dapat menyerang satwa primata terutama dari golongan kera. Infeksi virus hepatitis B pada satwa primata terdeteksi melalui uji serologis maupun uji viral. Deteksi viral virus hepatitis B dilakukan melalui polymerase chain reaction (PCR).Secara eksperimental telah dilakukan transmisi VHB manusia melalui inokulasi cairan saliva dari manusia yang menderita hepatitis B ke satwa primata keluarga Hylobatidae. Replikasi virus yang terjadi pada satwa tersebut mengindikasikan adanya hubungan kekerabatan yang dekat antara kedua inang (manusia dan Hylobatidae).
Penelitian mengenai infeksi virus hepatitis B pada owa jawa (Hylobates
moloch), salah satu spesies dari keluarga Hylobatidae, belum banyak dilakukan di
Indonesia meskipun spesies ini merupakan spesies endemik Indonesia. Penelitian ini bertujuan mengeksplorasi informasi mengenai infeksi virus hepatitis B (VHB) melalui penyidikan isolasi dan identifikasi VHB dari spesies owa jawa (Hylobates
moloch) yang berasal dari pusat rehabilitasi dan lembaga konservasi eks-situ di
Indonesia, dilanjutkan dengan melakukan pengkarakterisasian virus hepatitis B asal spesies tersebut.
Penelitian ini dilakukan dari bulan Januari 2011 sampai Juni 2011. Metode yang dilakukan untuk memperoleh isolat VHB yaitu melalui uji PCR yang dilakukan terhadap sembilan sampel owa jawa yang telah berstatus positif antigen hepatitis B permukaan (HbsAg). Menggunakan kit ekstraksi DNA (QIAmp DNA
Mini Blood Kit,Qiagen) didapatkan ekstrak DNA dari sediaan plasma owa jawa.
Amplifikasi DNA dilakukan dengan memanfaatkan primer yang dirancang untuk mengamplifikasi daerah Pre-S1 yang merupakan daerah variabel dan karakteristik untuk VHB yang berasal dari spesies yang berbeda. Untuk mengamplifikasi VHB regio Pre-S1 digunakan pasangan primer hepB-SF1 dengan sekuens 5’-TGYGGGTCACCWTATTCTTGGG-3’ dan hepB-SRout yang memiliki sekuens 5’-CACTGTTCCTGAACTGGAGC-3’. Pasangan primer
tersebut memiliki target produk kurang lebih 455 pasang basa. Pasangan primer ini sebelumnya telah diketahui dapat mengamplifikasi VHB daerah Pre-S1 dari isolat orangutan. Hasil amplifikasi menunjukkan bahwa semua isolat hasil studi memperlihatkan pita DNA VHB pada elektroforesis horizontal menggunakan gel agarosa.
Penggunaan enzim restriksi BST2UI mampu menunjukkan bahwa VHB yang menginfeksi owa jawa berbeda dengan VHB yang menginfeksi orangutan maupun manusia. Hal ini terlihat dari visualisasi gel elektroforesis produk PCR yang terpotong dengan menggunakan enzim tersebut. Produk PCR dari owa jawa maupun dari kontrol positif VHB dari orangutan, dapat terfragmentasi menggunakan enzim restriksi BST2UI, namun produk PCR kontrol positif VHB manusia tidak dapat terpotong. Posisi pemotongan enzim restriksi yaitu pada urutan spesifik CC(A/T)GG, dari hasil sekuensing, terlihat bahwa tiap amplikon mempunyai posisi dan jumlah situs pemotongan yang berbeda. Dengan demikian enzim BST2UI dapat digunakan sebagai deteksi awal yang membedakan infeksi VHB berasal dari manusia atau bukan manusia.
Untuk mendapatkan sekuens lengkap dari genom VHBGi dilakukan amplifikasi menggunakan empat pasang primer yang didisain berasal dari daerah yang conserved sehingga masing-masing amplikon yang dihasilkan mempunyai fragmen kontagius yang saling overlapped. Pasangan primer pertama adalah PreS1F (5’-GGGTCACCATATTCTTGGGAAC-3’) dan R5(5’- AGCCCAAA AGACCCACAATTC-3’). Target produk PCR yang diharapkan sebesar 1840 pasang basa. Set primer kedua adalah F6 (5’-ATATGGATGATGTGGTA TTGGG-3’) dan X102 (5’-ACCTTTAACCTAATCTCC-3’). Target produk PCR yang diharapkan sebesar 1027 pasang basa. Primer forward X101 (5’-TCTGTGCCTTCTCATCTG-3’) dengan primer reverse CORE2 (5’-CCCAC CTTATGAGTCCAAGG-3’). Target produk PCR yang diharapkan sebesar 924 pasang basa. Set primer keempat adalah CORE1 (5’-GAGTGTGGATTC GCACTCCTCC-3’) dan R1 (5’-TGTAACACGAGCAGGGGTCCTA-3’) dengan target produk PCR sebesar 2109 pasang basa. Dua dari 9 isolat VHB dapat teramplifikasi dengan baik menggunakan empat pasang primer tersebut. Hal ini ditunjukkan dengan terlihatnya pita DNA hasil amplifikasi dari masing-masing pasangan primer.
Produk PCR yang didapatkan baik menggunakan primer untuk daerah Pre-S1 maupun genom lengkap VHB kemudian dilakukan purifikasi menggunakan kit ekstraksi gel QiaQuick (QiaGen, USA), sebelum dilakukan sekuensing untuk mendapatkan data sekuens VHBGi dari isolat hasil studi, baik untuk regio Pre-S1 ataupun sekuens genom lengkap VHB Gi.
Hasil analisa filogenetik VHBGi daerah Pre-S1 (459 nukleotida) menggunakan program MEGA memperlihatkan bahwa terjadi keragaman di antara isolat owa jawa hasil studi, namun isolat owa jawa tersebut berada pada percabangan tersendiri terpisah dari kelompok satwa primata lainnya yang data sekuensya didapat dari GenBank.
Untuk sekuens genom lengkap, pohon filogenetik memperlihatkan bahwa isolat hasil studi berada pada satu cluster tersendiri. Hubungan kekerabatan yang dekat dengan orangutan diperlihatkan dengan adanya percabangan yang lebih dekat antara isolat owa jawa hasil studi dengan isolat orangutan dibandingkan dengan isolat manusia.
@Hak Cipta Milik IPB, tahun 2012 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk laporan apapun tanpa izin IPB
ISOLASI, IDENTIFIKASI DAN KARAKTERISASI
MOLEKULAR VIRUS HEPATITIS B
DI OWA JAWA (Hylobates moloch) DI INDONESIA
RACHMITASARI NOVIANA
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Primatologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012
Penguji Luar Komisi pada Ujian Tesis: Dr. Ir. Dedi Duryadi Solihin, DEA
Judul Tesis : Isolasi, Identifikasi dan Karakterisasi Molekular Virus Hepatitis B pada Owa Jawa (Hylobates moloch) di Indonesia
Nama : Rachmitasari Noviana
NIM : P053090031
Disetujui Komisi Pembimbing
Dr. drh. Joko Pamungkas, M.Sc Dr. drh. Diah Iskandriati
Ketua Anggota
Diketahui
Ketua Mayor Primatologi Dekan Sekolah Pascasarjana
Prof. drh. Dondin Sajuthi, MST. PhD Dr. Ir. Dahrul Syah, MSc.Agr
Judul Tesis : Isolasi, Identifikasi dan Karakterisasi Molekular Virus Hepatitis B pada Owa Jawa (Hylobates moloch) di Indonesia
Nama : Rachmitasari Noviana
NIM : P053090031
Disetujui Komisi Pembimbing
Dr. drh. Joko Pamungkas, M.Sc Dr. drh. Diah Iskandriati
Ketua Anggota
Diketahui
Ketua Mayor Primatologi Dekan Sekolah Pascasarjana
Prof. drh. Dondin Sajuthi, MST. PhD Dr. Ir. Dahrul Syah, MSc.Agr
PRAKATA
Segala puji dan syukur penulis haturkan ke hadirat Allah SWT atas segala limpahan rahmatNya pada saat melakukan penelitian hingga dapat diselesaikannya tesis yang bertema virus hepatitis B dengan judul tesis Isolasi,
Identifikasi dan Karakterisasi Molekular Virus Hepatitis B pada Owa Jawa (Hylobates moloch) di Indonesia.
Penulis menyampaikan penghargaan setinggi-tingginya kepada Dr. drh. Joko Pamungkas, M.Sc, selaku ketua komisi pembimbing sekaligus sebagai kepala Pusat Studi Satwa Primata, LPPM-IPB dan Dr. drh. Diah Iskandriati selaku anggota komisi pembimbing dan kepala Laboratorium Mikrobiologi dan Imunologi PSSP, LPPM IPB atas segala bimbingan, arahan, pengertian dan dukungan fasilitas serta dana penelitian sejak perencanaan penelitian sampai penulisan tesis dapat diselesaikan. Ucapan terima kasih penulis sampaikan pula kepada Dr. Ir. Dedi Duryadi Solihin, DEA sebagai penguji luar komisi yang tidak hanya mengenalkan penulis dalam dunia analisa molekular namun juga memberikan tambahan wawasan pengetahuan kepada penulis dan masukan dalam penulisan tesis ini.
Rasa terima kasih dan penghargaan yang tulus penulis sampaikan kepada Prof. drh. Dondin Sajuthi, MST, Ph.D; selaku Ketua Program Studi Primatologi Sekolah Pascasarjana IPB yang telah membuka kesempatan kepada penulis untuk melanjutkan pendidikan ke jenjang yang lebih tinggi. Kepada Direktur Taman Safari Indonesia, atas kerjasamanya dalam penelitian ini, rasa terima kasih juga penulis sampaikan.
Kepada teman sejawat di Laboratorium Mikrobiologi dan Imunologi, PSSP LPPM-IPB, Uus Saepuloh, S.Si, M.BioMed; Silmi Mariya, S.Si, MS; dra. Maryati Surya, MS; dra. Isti Kartika Sari; Sela Mariya, S.Si; Iin Indriawati, Tri Faujiani, Dede Juarsa dan Budi Doyo serta teman-teman di Pusat Studi Satwa Primata LPPM IPB atas kerjasamanya dan dukungan moril baik langsung maupun tidak langsung.
Ucapan terima kasih tidak lupa penulis haturkan kepada seluruh staf pengajar Program Studi Primatologi atas kesempatan untuk mendapatkan tambahan wawasan ilmu pengetahuan dan etika keilmuan selama penulis
mengikuti perkuliahan di Program Studi Primatologi, Sekolah Pascasarjana IPB. Ucapan terima kasih juga disampaikan kepada staf administrasi Program Studi Primatologi atas segala bantuannya selama ini.
Kepada ibunda tercinta, Siti Hidajati, yang tak pernah putus berdoa untuk keberhasilan penulis, kepada suami dan anak-anakku terkasih, Dr. Ir. Entang Iskandar, MS; Dienita Aulia dan Tiara Dwina Amany yang dengan penuh kesabaran, pengertian dan kasih mendampingi penulis selama ini, penulis sangat berterima kasih. Almarhum ayahanda Purnomo dan Mohamad Dawami serta ibunda Siti Sadiah yang selama hidupnya selalu mengingatkan untuk selalu belajar dan belajar. Kepada kakak-kakakku dan adik-adikku terima kasih atas doa dan semangatnya selama ini.
Penghargaan dan ucapan terima kasih disampaikan kepada semua pihak yang tidak dapat disebutkan satu persatu sehingga peneliti dapat menyelasaikan studinya di program studi ini.
Semoga hasil penelitian penulis dapat menambah khasanah wawasan dan berguna bagi dunia ilmu pengetahauan dan kesejahteraan manusia serta hewan.
Bogor, Februari 2012
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 14 November 1972 dari ayah Purnomo dan ibu Siti Hidayati. Penulis merupakan putri pertama dari tiga bersaudara.
Penulis menyelesaikan pendidikan dasarnya pada tahun 1985 di SD Hang Tuah VI, Jakarta. Tahun 1988 menyelesaikan Sekolah Menengah Pertama di SMPN 30 Jakarta, dan tahun 1991 menyelesaikan Sekolah Menengah Atas di SMAN 3 Jakarta. Pada tahun yang sama, penulis lulus seleksi masuk IPB melalui Undangan Seleksi Masuk IPB. Tahun 1996 penulis meyelesaikan pendidikan Strata 1 dari Fakultas Kedokteran Hewan IPB.
Tahun 2009, penulis diterima masuk Sekolah Pascasarjana pada Mayor Primatologi, Institut Pertanian Bogor.
DAFTAR ISI
halaman
DAFTAR GAMBAR ………. xi
DAFTAR TABEL …….……… xi
DAFTAR LAMPIRAN ……….. xii
PENDAHULUAN ……… 1 Latar Belakang ………... 1 Tujuan Penelitian ………... 2 Manfaat Penelitian ……….. 3 TINJAUAN PUSTAKA ………..……… 4 Owa jawa ………... 4 Taxonomi ……….………. 4
Morfologi owa jawa ……….………. 5
Status konservasi ……… ……….. 5 Virus Hepatitis B ……… 6 Klasifikasi Virus ……….…….. 6 Genom Virus ………. 6 Replikasi Virus ……….. 7 Transmisi Virus ………. 8 Patogenesa ……….. 9
Virus Hepatitis B pada satwa primata ……….... 9
Identifikasi asam nukleat ……….. 10
METODE PENELITIAN ………... 12
Waktu dan Tempat Penelitian ……… 12
Sampel Penelitian……… 12
Ekstraksi DNA ……….. . 12
Amplifikasi DNA untuk Sekuens Daerah Pre-S1 VHB ……… 12
Restriction Fragment Length Polymorphism ………. 14
Amplifikasi DNA untuk Genom Lengkap VHB ………. 14
Pemurnian Produk PCR ………. 15
Analisa Hasil Sekuensing ……….. 15
Alur penelitian ……… 16
HASIL DAN PEMBAHASAN ………. 17
Virus hepatitis B Gibbon Regio Pre-S1……….. 17
Amplifikasi Virus Hepatitis B Regio Pre-S1 ……… 17
Restriction Fragment Length Polymorphism ……… 19
Pembuatan Pohon Filogenetik ……….. 22
Sekuens Genom Lengkap Virus Hepatitis B Gibbon ………. 25
Amplifikasi Virus Hepatitis B Gibbon ………. 25
SIMPULAN DAN SARAN ………... 31 DAFTAR PUSTAKA ……….. 32 LAMPIRAN ………. 35
DAFTAR GAMBAR
halaman 1 Organisasi genom virus hepatitis B manusia (sirkular) ……… 7 2 Siklus hidup virus hepatitis B ………....………. 8 3 Organisasi virus hepatitis B pada woolly monkey ……….. 10 4 Visualisasi PCR VHB regio Pre-S1 yang menginfeksi spesies owa jawa
di lokasi A ………. 18
5 Visualisasi PCR VHB regio Pre-S1 yang menginfeksi spesies owa jawa
di lokasi B ………. 18
6 Visualisasi pemotongan produk DNA menggunakan enzim restriksi
BSt2UI lokasi A……….. 20 7 Visualisasi pemotongan produk DNA menggunakan enzim restriksi
BSt2UI lokasi B ……….. 21 8 Pohon filogenetik VHB Hylobatidae berdasarkan (a) sekuens nukleotida
VHB regio Pre-S1 (459pb); (b) situs pemotongan sekuens nukleotida VHB region pre-S1 menggunakan ensim restriksi BsT2UI ……….. 22 9 Pohon filogenetik VHB Hylobatidae dan orangutan berdasarkan sekuens nukleotida VHB regio Pre-S1 (459 nuklotida)……… 24 10 Pohon filogenetik VHB Hylobatidae dan orangutan berdasarkan sekuens
asam amino VHB regio Pre-S1 ……… 24 11 Visualisasi hasil amplifikasi genom lengkap VHBGi ………... 26 12 Rekonstruksi genom lengkap VHBGi linear berdasarkan posisi primer
yang digunakan ………. 26 13 Prediksi genom linear sekuens sampel C1 dibandingkan dengan sekuens
H. pileatus dari GenBank ……….….. 28
14 Pohon filogenetik VHB genom lengkap asal satwa primata dan
DAFTAR TABEL
halaman 1 Klasifikasi dan penyebaran genus Hylobates ………. 4 2 Pasangan primer untuk amplifikasi genom lengkap VHBGi ………. 14 3 Hasil pemeriksaan serologis HbSAg (data sekunder) dan hasil PCR atas
Regio Pre-S1 dari virus hepatitis B pada owa jawa ……….. 17 4 Data sekuens produk PCR VHBGi regio Pre-S1 dan situs pemotongan
dari enzim restriksi BSt2UI ... 19 5 Data lokasi gen VHB pada isolat Ttblack (GenBank, AY330916) ….. 27 6 Daerah gen VHBGi berdasarkan pasangan primer yang digunakan ….. 28
DAFTAR LAMPIRAN
1 Pensejajaran berganda virus hepatitis B regio Pre-S1 isolat hasil
studi dan situs pemotongan enzim restriksi BSt2UI halaman ………….. 35 2 Data sekuens GeneBank virus hepatitis B ………. 37 3 Komposisi asam amino VHBGi regio Pre-S1 ……… 38
PENDAHULUAN
Latar BelakangHepatitis merupakan manifestasi klinis dari perubahan jaringan atau organ hati berupa peradangan atau inflamasi yang dikarakterisasikan dengan terdapatnya sel-sel inflamatori pada jaringan hati. Peradangan tersebut dapat diakibatkan oleh agen yang bersifat non-infeksius seperti minuman beralkohol dan penggunaan obat-obatan, sedangkan agen yang bersifat infeksius dapat disebabkan oleh infeksi virus hepatitis atau bakteria. Pada infeksi virus hepatitis penyakit dapat bersifat akut maupun kronis serta dapat berakhir dengan kematian akibat terjadinya sirosis pada hati. Saat ini menurut data WHO satu dari tiga penduduk dunia telah terinfeksi virus ini sementara satu dari 20 penduduk dunia hidup dengan infeksi kronis (WHO, 2011).
Sampai saat ini terdapat lima virus penting yang dilaporkan menjadikan hati sebagai organ target utama infeksi, yaitu virus Hepatitis A (VHA), B (VHB), C (VHC), D (VHD) dan virus Hepatitis E (VHE). Virus hepatitis B (VHB) merupakan satu-satunya virus DNA dalam keluarga virus hepatitis. Tiga cara utama transmisi virus hepatitis B yaitu melalui darah, hubungan kelamin (sexual
intercourse) dan perinatal (mother to child) pada saat melahirkan atau menyusui.
Transmisi perinatal dan infeksi carrier yang bersifat jangka panjang menyebabkan adanya endemisitas dan prevalensi tinggi di benua Asia terutama bagian selatan dan timur.
Selain menginfeksi manusia, VHB dilaporkan dapat pula menginfeksi beberapa spesies satwa primata di fasilitas konservasi eks-situ, terutama dari golongan kera yaitu simpanse (Pan troglodytes), orangutan (Pongo sp), gorilla dan gibbon (Hylobates sp), serta dari golongan monyet yaitu woolly monkey (Lagothrix lagotricha).
Adanya infeksi VHB dapat dideteksi melalui uji serologis untuk antigen permukaan VHB (HBsAg) dan deteksi DNA viral melalui uji polymerase chain
reaction (PCR). Mac Donnald et al. (2000), menemukan kejadian infeksi VHB
pada simpanse yang dilahirkan di alam (wild-born). Isolasi VHB pada satwa primata dari golongan monyet dilaporkan pertama kali dilakukan dari Lagothrix
lagotricha (woolly monkey) asal kebun binatang di Amerika Serikat, yang
mengalami peradangan hati (Lanford et al. 1998).
Analisis pohon filogenetik mengindikasikan bahwa virus yang menginfeksi simpanse dan Hylobates bersifat indigenus pada masing-masing inangnya (Norder
et al. 1996) dan berada pada cabang pohon filogenetik yang berbeda dengan
infeksi VHB pada manusia (Mac Donnald et al. 2000). Secara eksperimental telah dilakukan pula transmisi VHB manusia melalui inokulasi cairan saliva dari manusia yang menderita hepatitis B ke satwa primata keluarga Hylobatidae (Scott
et al. 1980). Replikasi virus yang terjadi pada satwa tersebut mengindikasikan
adanya hubungan kekerabatan yang dekat antara manusia dan Hylobatidae dan kemungkinan adanya transmisi alami dari manusia ke keluarga Hylobatidae.
Penelitian mengenai infeksi virus hepatitis B pada owa jawa di Indonesia belum banyak dilakukan. Informasi kejadian infeksi VHB pada satwa ini banyak berasal dari luar Indonesia, meskipun spesies ini merupakan spesies endemik Indonesia. Owa jawa juga merupakan spesies yang terancam punah menurut International Union for Conservation of Nature (2008). Dengan status ini, telah dilakukan upaya untuk mengatasi kepunahan spesies owa jawa ini melalui beberapa usaha berupa penangkaran, taman satwa dan upaya pengembalian atau pelepasliaran satwa ini ke habitat aslinya. Upaya tersebut sebaiknya diiringi pula dengan pemeriksaan status kesehatan satwa dan orang-orang yang mengalami kontak langsung dengan satwa tersebut. Salah satunya adalah virus hepatitis B yang telah diketahui dapat menginfeksi manusia dan satwa primata.
Tujuan Penelitian
Mengeksplorasi informasi mengenai infeksi virus hepatitis B pada satwa primata dari keluarga Hylobatidae, khususnya melalui penyidikan isolasi dan identifikasi VHB dari spesies owa jawa (Hylobates moloch) yang berasal dari pusat rehabilitasi dan beberapa lembaga konservasi eks-situ di Indonesia, dilanjutkan dengan melakukan pengkarakterisasian virus hepatitis B asal spesies tersebut.
Manfaat Penelitian
Dengan informasi yang diperoleh mengenai infeksi virus VHB dan perbedaan karakter antara virus hepatitis B yang menginfeksi manusia (VHBHu) dengan virus Hepatitis yang menginfeksi owa jawa (VHBGi), diharapkan dapat membantu penapisan status mikrobiologik owa jawa di fasilitas lembaga konservasi eks-situ, serta lebih lanjut dapat dimanfaatkan dalam pengelolaan manajemen kesehatan satwa tersebut.
TINJAUAN PUSTAKA
Owa jawa
Taksonomi
Owa jawa (Hylobates moloch), dikenal pula dengan nama Javan gibbon atau Silvery gibbon, menurut Napier dan Napier (1985), diklasifikasikan sebagai berikut: Ordo : Primate Subordo : Anthropoidea Infra-ordo : Catarrhini Superfamili : Hominoidea Famili : Hylobatidae Genus : Hylobates
Spesies : Hylobates moloch
Menurut Geissmann (1995), genus Hylobates dapat dikelompokkan dalam empat subgenus, yaitu Hylobates, Nomascus, Bunopithecus dan Sympalangus. Pola penyebaran dari masing-masing subgenus disajikan pada tabel di bawah ini.
Tabel 1 Klasifikasi dan Penyebaran genus Hylobates
Genus Subgenus Spesies Penyebaran
Hylobates Hylobates Nomascus Bunopithecus Sympalangus Agilis Lar Moloch Muelleri Pileatus Klosii Concolor Leucogenys Gabriellae Hoolock Syndactylus Sumatera Barat, Kalimantan, Malaysia,Thailand, Burma, Semenanjung Malaysia, Yunan, Sumatera Barat Jawa Barat, Jawa Tengah Kalimantan
Thailand, Kamboja Mentawai
Vietnam, Yunan, Laos Laos, Vietnam
Laos, Vietnam, Kamboja Assam, Bangladesh, Burma
Semenanjung Malaysia, Sumatera
Morfologi owa jawa
Owa jawa adalah satwa primata arboreal, dengan tempat hidupnya adalah kanopi pohon. Mereka tidak mempunyai ekor, mempunyai formulasi gigi yang sama dengan Pongidae. Mempunyai tangan yang panjang, dengan panjang tangan dapat mencapai tanah disaat mereka berdiri dengan dua kaki (bipedal). Pergelangan tangan dan bahu telah mengalami adaptasi sehingga memudahkan pergerakan mereka dalam brakhiasi. Nowak (1999) mendefinisikan Hylobates sebagai penghuni pohon, dan gibbon (owa) sangat sesuai dengan penamaan tersebut. Ketangkasan genus ini dalam melakukan brakhiasi, bergerak dari satu pohon ke pohon lainnya, melebihi satwa lainnya.
Supriatna dan Wahyono (2000) menyatakan bahwa tubuh owa Jawa ditutupi rambut yang berwarna kecoklatan sampai keperakan atau kelabu. Bagian dagu pada beberapa individu berwarna gelap. Rambut di atas kepala hitam dan kulit muka hitam, alis berwarna putih, rambut pada bayi berwarna kelabu terang dibanding dengan dewasa (Rowe 1996).
Adanya pembengkakan pada pada alat kelamin betina, terutama pada
Hylobates moloch, merupakan cirri menonjol pada genus Hylobates, namun
pembengkakan ini tidak begitu nyata terlihat pada Hylobates pileatus (Mootnick 2006).
Status Konservasi
Owa jawa merupakan salah satu spesies endemik Indonesia. Keberadaan spesies ini telah dilindungi sejak tahun 1931 untuk menghindari kepunahan melalui Peraturan Perlindungan Binatang Liar No. 266 yang kemudian diperkuat dengan Undang-undang No. 5 tahun 1990 dan SK Menteri Kehutanan 10 Juni1991 (Supriatna & Wahyono 2000). Pada tahun 1986 – 1990, International
Union for Conservation Nation (IUCN) telah memasukkan owa jawa sebagai
spesies yang terancam punah. Dikatakan sebagai terancam punah karena populasinya di alam diperkirakan kurang dari 2500 individu, kemudian dengan observasi yang berkesinambungan terjadi penurunan jumlah individu dewasa dan tidak ada subpopulasi yang terdiri lebih dari 250 individu dewasa (IUCN
Virus Hepatitis B Klasifikasi Virus
Virus Hepatitis adalah virus yang menjadikan hati sebagai target utama infeksi. Infeksi virus dapat menyebabkan peradangan hati yang ditandai dengan ditemukannnya sel-sel inflamatori pada hati. Terdapat lima virus yang dikenal dapat mengakibatkan hepatitis dan berasal dari keluarga virus yang berbeda. Virus hepatitis A merupakan anggota dari keluarga Picornaviridae. Virus hepatitis B adalah anggota keluarga Hepadnavidae. Virus hepatitis C merupakan anggota dari keluarga Flaviviridae, sedangkan virus hepatitis D dan E masing-masing merupakan anggota dari keluarga Deltaviridae dan Caliciviridae.
Menurut Komite Internasional Taksonomi Virus (International Committee
on Taxonomy of Viruses, ICTV, 2009) keluarga Hepadnaviridae dibagi menjadi
dua genus yaitu:
1. Genus Orthohepadnavirus, yaitu virus hepatitis yang menyerang mamalia, seperti hepatitis B virus (yang menginfeksi ordo primata), woodchuck
hepatitis virus, ground squirrel hepatitis virus dan arctic squirrel hepatitis virus
2. Genus Avihepadnavirus, yaitu virus hepatitis yang menyerang bangsa unggas, seperti duck hepatitis virus, heron hepatitis virus dan goose
hepatitis virus.
Genom Virus
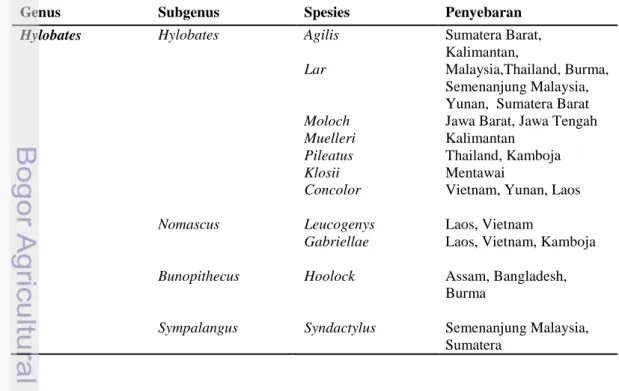
Virus hepatitis B (VHB), sesuai dengan nama keluarga (Hepadnavirus) adalah virus DNA dengan virion beramplop (envelope) berukuran 42-nm, dengan sebagian DNA virion adalah utas ganda (partially double stranded). Virus ini merupakan virus DNA hewan berukuran terkecil dan mempunyai ukuran genom sebesar kurang lebih 3200 pasang basa, terdiri dari empat open reading frame (ORF) untuk gen P, C, S dan X yang masing-masing mengkode DNA polimerase/reverse transcriptase, protein inti (core), protein permukaan (surface) dan protein X. Untuk gen S dibagi menjadi regio pre-S1, pre-S2 dan S. Gen C terbagi menjadi regio pre-C dan C.
Protein permukaan yang berada pada pembungkus virus (envelope) dikenal sebagai antigen permukaan (HbsAg) yang merupakan protein penting dalam pendiagnosaan klinis infeksi dan imunisasi virus ini.
Selain HBsAg terdapat dua antigen penting lainnya yaitu antigen inti hepatitis B (HBcAg) yang membentuk nukleokapsid virion, dan antigen e (HBeAg) adalah antigen yang dikeluarkan ke dalam peredaran darah oleh sel-sel yang terinfeksi virus (Levinson, 2008)
Gambar 1 Organisasi genom virus hepatitis B manusia (sirkular). Sumber: Wands, JR. 2004
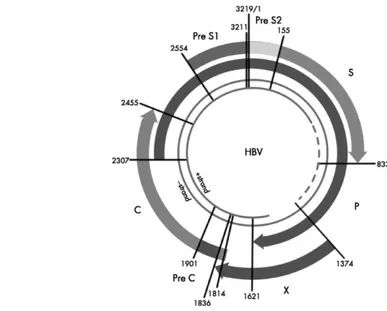
Replikasi Virus
Virus hepatitis B merupakan virus DNA dengan utas ganda sebagian yang
menggunakan enzim transkripsi balik (reverse transcriptase) dalam replikasinya. Proses replikasi virus secara umum terdiri dari beberapa tahap, yaitu perlekatan (attachment), penetrasi (penetration), uncoating, ekspresi gen, replikasi genom,
assembly dan pelepasan (release). Proses transkripsi terjadi di dalam nukleus,
sementara replikasi genom berlangsung di sitoplasma, di dalam protein inti (White dan Fenner, 1994).
Protein permukaan virion dapat menempel (attach) pada permukaan sel inang melalui reseptor spesifik. Situs penempelan virus hepatitis B adalah pada protein L. Virion yang menempel pada permukaan sel inang mengalami endositosis, kemudian nukleokapsid akan dikeluarkan dari endosoma melalui fusi
yang terjadi antara virion dan membran endosoma. Nukleokapsid akan memasuki nukleus sel inang, genom virus akan terlepas dalam nukleus sel inang dan berkonversi menjadi molekul DNA sirkular (Carter dan Sanders, 2007).
Gambar 2 Siklus hidup virus hepatitis B (http://www.natap.org/2004/VHB/100804_02.htm)
Asam deoksiribonukleat rantai ganda sirkular (covalently closed circular, cccDNA) ini kemudian menjadi cetakan untuk sintesa asam ribonukleat
messanger (mRNA) menggunakan enzim polimerase RNA selular. Hepadnavirus
merupakan keluarga DNA virus yang unik karena menggunakan mRNA sebagai cetakan dalam menghasilkan genom DNA melalui transkripsi terbalik (reverse
transcription) (White dan Fenner, 1994).
Transmisi Virus
Infeksi virus ini ditularkan melalui darah, hubungan kelamin dan perinatal (dari ibu ke anak saat melahirkan dan menyusui). Transmisi melalui jarum suntik yang terkontaminasi virus memperlihatkan bahwa transmisi sangat mudah terjadi. Infeksi kronis VHB dapat mengakibatkan sirosis pada hati dan hepatocellular
Patogenesa
Setelah menginfeksi inangnya dan memasuki peredaran darah, VHB akan menginfeksi hepatosit kemudian antigen viral akan berada pada permukaan sel inang. Sel T sitotoksik akan memediasi sistem pertahanan tubuh untuk melawan masuknya antigen viral berupa adanya inflamasi dan nekrosis. Virus ini tidak menghasilkan efek sitopatik, sehingga diduga patogenesa virus ini merupakan hasil dari pertahanan tubuh bermediasi sel (White dan Fenner, 1994). Penderita dapat menjadi chronic carrier, bila antigen permukaan VHB (HBsAg) terdeteksi lebih dari 6 bulan. Pada penderita chronic carrier, terjadi kasus hepatocellular
carcinoma dengan prevalensi tinggi (Levinson, 2008)
Virus Hepatis B Pada Satwa Primata
Warren et al. (1999) menemukan adanya infeksi VHB di lapangan secara alami pada orangutan yang berada di Pusat Rehabilitasi Orangutan Wanariset, Kalimantan Timur. Sebanyak 195 sampel serum diujikan untuk mendeteksi anti-hepatitis B inti (HBcAb), anti-HB permukaan (HBsAb) dan antigen permukaan hepatitis B (HBsAg) serta uji PCR. Ditemukan bahwa 55 individu adalah HBsAg positif, 28 HBsAb positif, uji PCR yang dilakukan pada individu HbsAg positif diperoleh 32 sampel adalah positif VHB.
Vaudin et al. (1988) menemukan adanya infeksi VHB pada simpanse (Pan
troglodytes), VHB juga terbukti dapat menginfeksi genus Hylobates dan Nomascus (Noppornpanth et al. 2003), Gorilla gorilla (Grethe et al. 2000) dan Lagothrix lagothricha (Lanford et al. 1998).
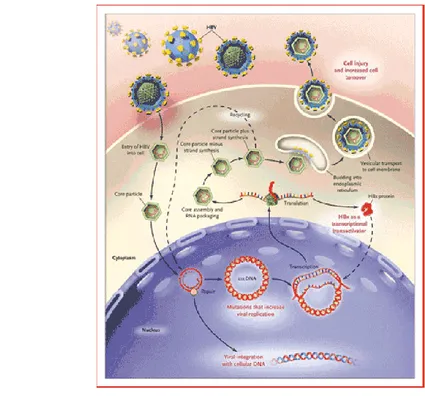
Infeksi VHB yang terjadi pada woolly monkey menjadi acuan awal penelitian hepatitis B pada satwa primata. Lanford et al. (1998) menemukan bahwa hepadnavirus yang diisolasi dari woolly monkey mempunyai perbedaan dari VHB yang berasal dari isolat manusia. Analisa filogenetik terhadap sekuens nukleotida dilakukan pada bagian gen inti dan permukaan. Ditemukan bahwa sekuens tersebut merupakan basal atau ancestral dari grup VHB pada manusia, sehingga diperkirakan bahwa virus hepatitis B yang menginfeksi woolly monkey merupakan progenitor dari virus hepatitis B manusia.
Gambar 3 Organisasi virus VHB pada woolly monkey (WMHBV) (Lanford et al. 1998)
Transmisi VHB juga terjadi pada genus Hylobates. Analisa filogenetik dari isolat genus tersebut menyatakan bahwa sekuens nukleotida gen permukaan VHB yang menginfeksi Hylobates yang berada dalam lembaga konservasi berada pada
cluster yang berbeda dengan VHB yang berasal dari inang lainnya (Noppornpanth et al. 2003). Virus hepatitis B ditemukan tidak hanya pada sediaan darah namun
juga dari cairan saliva Hylobates pileatus, H. lar, dan H. concolar. Dari analisa lanjutan menggunakan enzim restriksi (analisis RFLP) dari isolat gibbon dan isolat manusia terlihat bahwa VHB yang menginfeksi keduanya merupakan VHB yang mempunyai karakterisasi molekular yang berbeda (Noppornpanth et al. 2003).
Identifikasi Asam Nukleat Virus
Identifikasi agen virus dapat dilakukan melalui analisa genom virus. Penggunaan reaksi enzim Taq DNA polimerase dalam tehnik PCR (Polymerase
Chain Raction) memungkinkan identifikasi secara molekular yang memiliki
sensitifitas tinggi dengan mengamplifikasi hanya dari satu molekul DNA tunggal dan kopi gen tunggal dapat diekstraksi dari campuran genomik yang kompleks. Dengan kata lain, PCR merupakan suatu reaksi in vitro untuk menggandakan (mengamplifikasikan) jumlah molekul DNA pada target tersebut dengan bantuan enzim Taq DNA polimerase dan oligonukleotida sebagai primer dalam sebuah mesin thermocycler (Ubaidillah dan Sutrisno, 2009).
Tehnik amplifikasi DNA berbasis pada siklus termal berupa pemanasan dan pendinginan secara berulang yang terdiri dari tiga tahap yaitu pemecahan
(denaturation), penempelan (annealing) dan pemanjangan (elongation). Primer
yang digunakan berisi sekuens komplementari yang didisain untuk mengamplifikasi region target tertentu . Primer yang berada sebelum daerah target disebut primer forward dan yang berada setelah target disebut primer reverse (Ubaidillah dan Sutrisno, 2009). Hasil amplifikasi DNA dengan tehnik PCR kemudian dapat divisualisasikan sebagai pita-pita DNA pada gel agarosa. Teknik ini sangat efisien untuk mengamplifikasi urutan DNA VHB.
Enzim endonuklease restriksi adalah enzim bakteri yang digunakan dalam tehnik molekular untuk mengenali sekuens spesifik dalam DNA dan kemudian melakukan pemotongan DNA tersebut untuk mendapatkan fragmen-fragmen spesifik yang dikenal sebagai fragmen restriksi (Ubaidillah dan Sutrisno, 2009). Ensim restriksi memainkan peranan penting dalam konstruksi molekul DNA rekombinan dan mapping lokasi dari situs restriksi pada DNA. Selain itu, situs spesifik dari enzim restriksi ini pada fragmen gen tertentu dapat dijadikan sebagai alat genotiping (karakteristik genotipe) dari individu pada spesies tersebut (Ubaidillah dan Sutrisno, 2009).
METODE PENELITIAN
Waktu dan Tempat PenelitianPenelitian ini dilakukan dari bulan Januari sampai dengan bulan Juni 2011, bertempat di Laboratorium Mikrobiologi dan Imunologi, Pusat Studi Satwa Primata, Lembaga Penelitian dan Pengabdian kepada Masyarakat, Institut Pertanian Bogor (PSSP LPPM-IPB), Jalan Lodaya II/5, Bogor 16151.
Sampel Penelitian
Sampel yang dimanfaatkan dalam penelitian ini adalah plasma owa jawa yang merupakan koleksi sampel Laboratorium Mikrobiologi dan Imunologi PSSP LPPM-IPB, sebagai bagian dari pemeriksaan rutin kesehatan satwa dari beberapa fasilitas konservasi eks-situ satwa primata. Semua sampel yang digunakan dalam penelitian ini, berasal dari 9 ekor satwa owa jawa yang memiliki status positif atas pemeriksaan terhadap antigen permukaan virus hepatitis B (HBsAg) melalui uji ELISA (data sekunder).
Ekstraksi DNA
Pemurnian DNA virus dilakukan dari sampel plasma owa jawa menggunakan kit QIAmp DNA Mini Blood Kit (Qiagen, USA) sesuai dengan petunjuk dari pedoman penggunaan dari perusahaan. Sebanyak 200µl sampel plasma ditambahkan ke dalam tabung mikro yang telah berisi 20µl (20mg/ml)
proteinase K. Larutan penyangga pelisis (lisis buffer) ditambahkan sebanyak
200µl ke dalam masing-masing tabung mikro. Untuk menghomogenkan campuran tersebut dilakukan homogenisasi menggunakan vortex dan dilanjutkan dengan inkubasi selama 10 menit pada suhu 560 C. Prosedur selanjutnya dilakukan sentrifugasi, pencucian dan elusi sesuai dengan prosedur baku dari kit ekstraksi DNA QiAmp DNA Miniblood Kit.
Amplifikasi DNA untuk Sekuens Daerah Pre-S1 VHB
Amplifikasi DNA dilakukan menggunakan metode polymerase chain
mengamplifikasi daerah Pre-S1 yang merupakan daerah variabel dan karakteristik untuk VHB yang berasal dari spesies yang berbeda. Set primer forward dan
reverse disintesa dari sekuens bagian paling conserved di daerah yang variabel di
antara berbagai strain VHB.
Sebanyak 50 µl reagen PCR yang terdiri dari, masing-masing 1µl primer
forward dan reverse (10 pmol/µl), 4 µl MgCl2 (25mM), 5 µl dNTPs (10 mM), 0,5µl Taq Gold Polymerase (5 U/µl), 5 µl PCR Buffer 10X (500mM KCl, 100mM Tris-HCl (pH 8,3), sampel DNA (10 ul) dan ddH2O (23,5 ul) dimasukkan ke dalam tabung mikro 200µl dan dihomogenkan menggunakan vortex.
Merujuk kepada penelitian yang dilakukan oleh Warren et al. (1999) yang telah berhasil mengamplifikasi VHB daerah Pre-S1 dari isolat orangutan, digunakan pasangan primer yang sama untuk mengamplifikasi VHB daerah Pre-S1 dari isolat DNA owajawa yaitu hepB-SF1 dengan sekuens 5’-TGYGGGTCACCWTATTCTTGGG-3’ dan hepB-SRout yang memiliki sekuens 5’-CACTGTTCCTGAACTGGAGC-3’. Pasangan primer tersebut memiliki target produk kurang lebih 455 pasang basa.
Amplifikasi DNA dilakukan menggunakan mesin PCR (Perkin Elmer, Model 9700), melalui beberapa tahapan. Pada tahap awal dilakukan pre-PCR untuk mengaktifkan enzim polymerase pada suhu 940C selama 10 menit. Tahapan selanjutnya adalah amplifikasi PCR yang terdiri atas denaturasi sampel pada suhu 940C selama 30 detik, annealing pada suhu 620C selama 30 detik, dan tahap elongasi pada suhu 720C selama 1 menit. Tahapan ini dilakukan selama 30 kali dengan siklus yang berulang. Tahap akhir adalah post-PCR dengan suhu 720C selama 10 menit.
Produk PCR yang telah diamplifikasi tersebut dijalankan pada gel agarosa 2% yang mengandung ethidium bromida 1 µg/ml dalam bufer TAE menggunakan elektroforesis horizontal. Penanda DNA 1 kb (Invitrogen, USA) dan produk PCR yang telah ditambahkan pewarna (loading dye) dimasukkan ke dalam sumur gel. Alat dokumentasi Gel Doc 2000 (BioRad, USA) digunakan untuk memvisualisasikan hasil elektroforesis. Sebagai kontrol positif PCR digunakan DNA positif VHB gibbon (VHBGi), VHB manusia (VHBHu) dan VHB orangutan (VHBOu).
Restriction Fragment Length Polymorphism (RFLP)
Terhadap produk PCR yang memberikan hasil positif pada uji PCR dilakukan digesti menggunakan enzim restriksi BSt2UI (1 U/µl) yang bekerja pada sekuens spesifik yaitu CC(A/T)GG dari sekuens nukleotida sampel. Enzim restriksi ini telah diketahui dapat memotong sekuens nukelotida dari VHBOu namun tidak dapat memotong sekuens nukleotida dari VHBHu. Sebanyak 20µl campuran reagensia yang terdiri dari 1µl enzim BSt2UI (1IU/ul), buffer pereaksi 10 x sebanyak 1,5µl, dan produk PCR sebanyak 3,5µl. Ditambahkan air destilasi sampai volume mencapai 20µl. Kemudian dilakukan inkubasi pada suhu 600C selama 1 jam.
Untuk memvisualisasikan hasil restriksi enzim, produk PCR yang telah diinkubasi dengan enzim restriksi tersebut dijalankan melalui gel agarosa menggunakan elektroforesis horizontal dengan berkonsentrasi 2% yang ditambahkan ethidium bromida sebagai pewarna (staining), selama 1,5 jam, 100V. Pembacaan hasil eletroforesis dilakukan melalui alat GelDoc.
Amplifikasi DNA untuk Sekuens Genom Lengkap VHB
Amplifikasi DNA untuk mendapatkan sekuens genom lengkap VHBGi merujuk kepada Sa-Nguanmoo et al. (2008) yang menggunakan empat set primer seperti tertera pada tabel 2 di bawah ini.
Tabel 2 Pasangan primer untuk amplifikasi genom lengkap VHBGi
Primer set
Primer Sekuens primer Posisi nukleotida Target produk (pb) 1 PreS1F 5’-GGGTCACCATATTCTTGGGAAC-3’ 2814 -2835 1840 R5 5’-AGCCCAAAAGACCCACAATTC-3’ 1015 - 995 2 F6 5’-ATATGGATGATGTGGTATTGGG-3’ 737-758 1027 X102 5’-ACCTTTAACCTAATCTCC-3’ 1764 - 1748 3 X101 5’-TCTGTGCCTTCTCATCTG-3’ 1552 - 1569 924 CORE2 5’-CCCACCTTATGAGTCCAAGG-3’ 2476 - 2457 4 CORE1 5’-GAGTGTGGATTCGCACTCCTCC-3’ 2268 - 2289 2109 R1 5’-TGTAACACGAGCAGGGGTCCTA-3’ 201 - 180
Sebanyak 50 µl reagensia PCR yang terdiri dari, masing-masing satu pasang primer forward dan reverse sebanyak 1µl (10 pmol/µl), 4 µl MgCl2 (25mM), 5 µl
dNTPs (10 mM), 1µl Taq Gold Polymerase (5 U/µl), 5 µl PCR Buffer 10X (500mM KCl, 100mM Tris-HCl (pH 8,3), sampel DNA (10 ul) dan ddH2O (23ul) dimasukkan ke dalam tabung mikro 200µl dan dihomogenkan menggunakan vortex.
Amplifikasi DNA dilakukan menggunakan mesin PCR (Perkin Elmer, Model 9700), melalui beberapa tahapan. Pada tahap awal dilakukan pre-PCR untuk mengaktifkan enzim polymerase pada suhu 940C selama 10 menit. Tahapan selanjutnya adalah amplifikasi PCR yang terdiri atas denaturasi sampel pada suhu 940C selama 30 detik, annealing pada suhu 550C selama 30 detik, dan tahap elongasi pada suhu 720C selama 2 menit. Tahapan ini dilakukan selama 40 kali dengan siklus yang berulang. Tahap akhir adalah post-PCR dengan suhu 720C selama 10 menit.
Produk PCR yang telah diamplifikasi tersebut dijalankan pada gel agarosa 2% yang mengandung ethidium bromida 1 µg/ml dalam bufer TAE menggunakan elektroforesis horizontal. Penanda DNA 1 kb dan produk PCR yang telah ditambahkan pewarna (loading dye) dimasukkan ke dalam sumur gel. Alat dokumentasi Gel Doc 2000 (BioRad, USA) digunakan untuk memvisualisasikan hasil elektroforesis. Sebagai kontrol positif digunakan DNA positif VHB gibbon (VHBGi), VHB manusia (VHBHu) dan VHB orangutan (VHBOU).
Pemurnian Produk PCR
Pemotongan gel produk PCR dilakukan dengan memotong gel yaitu tepat pada bagian gel yang memiliki pita yang berpendar saat diradiasi sinar UV. Potongan gel hasil amplifikasi kemudian dilakukan pemurnian menggunakan kit ekstraksi gel QiaQuick (Qiagen, USA). Untuk mendapatkan sekuens nukleotida, hasil pemurnian produk PCR dilakukan di Macrogen Inc, Korea. Sekuensing dilakukan baik terhadap produk PCR VHBGi regio Pre-S1 maupun genom lengkap VHBGi.
Analisa Hasil Sekuensing
Pembacaan hasil sekuensing mengunakan perangkat lunak komputer BioEdit. Pensejajaran urutan nukleotida dianalisa menggunakan program BLAST
2.0 (BLAST, 2011) dan ClustalW2 (Kumar et al. 2011). Pembuatan pohon filogenetik menggunakan perangkat lunak komputer MEGA versi 5.0 (Kumar et
al. 2011). Sebagai pembanding dimasukkan urutan nukleotida virus Hepatitis B
asal spesies Gibbon lainnya di luar Indonesia, orangutan, manusia , woolly
monkey, simpanse dan gorilla dari data GeneBank.
Alur Penelitian
Seleksi owa jawa yang terdeteksi positif dari hasil uji serologi HBsAg akan dilanjutkan dengan uji karakteristik diagnostik cepat melalui PCR dengan primer daerah Pre-S21 VHB.
Isolat yang memberikan karakteristik spesifik dengan sediaan material DNA yang memadai maka akan dilakukan karakteristik nukleotida penyusun virus hepatitis B pada owa jawa secara utuh.
Kemungkinan terjadinya transmisi VHB secara interspesies maupun
intraspesies pada owa jawa
Sampel darah owa jawa penangkaran eksitu
Karakteristik diagnostik cepat melalui PCR dengan primer
spesifik VHB daerah pre-S1dari sampel positif serologi owa jawa yang positif secara
uji serologi HBsAg
Rekonstruksi pohon filogenetik dengan satwa
primata lain
Karakteristik molekular secara utuh VHB pada owa jawa Rekonstruksi pohon
filogenetik dengan manusia dan satwa primata lain
HASIL DAN PEMBAHASAN
Virus Hepatitis B Gibbon Regio Pre-S1 Amplifikasi Virus Hepatitis B Regio Pre-S1Hasil amplifikasi dari 9 sampel DNA owa jawa yang telah berstatus serologis positif terhadap antigen virus hepatitis B menggunakan pasangan primer hepSF-1 dan hepSR-out menghasilkan fragmen pita DNA sekitar 459 pasang basa (Tabel 3). Hal ini menunjukkan bahwa ke-9 individu owa jawa tersebut terinfeksi dengan virus hepatitis B (VHB).
Tabel 3 Hasil pemeriksaan serologis HbsAg (data sekunder) dan hasil PCR atas regio Pre-S1 dari virus hepatitis B pada owa jawa
No. ID HBsAg Hasil PCR VHB regio pre-S1 Tempat Asal Satwa 1 A1 Positif Positif A 2 A2 Positif Positif A 3 A3 Positif Positif A 4 A5 Positif Positif A 5 A6 Positif Positif A 6 A8 Positif Positif A 7 A9 Positif Positif A 8 C1 Positif Positif B 9 C2 Positif Positif B
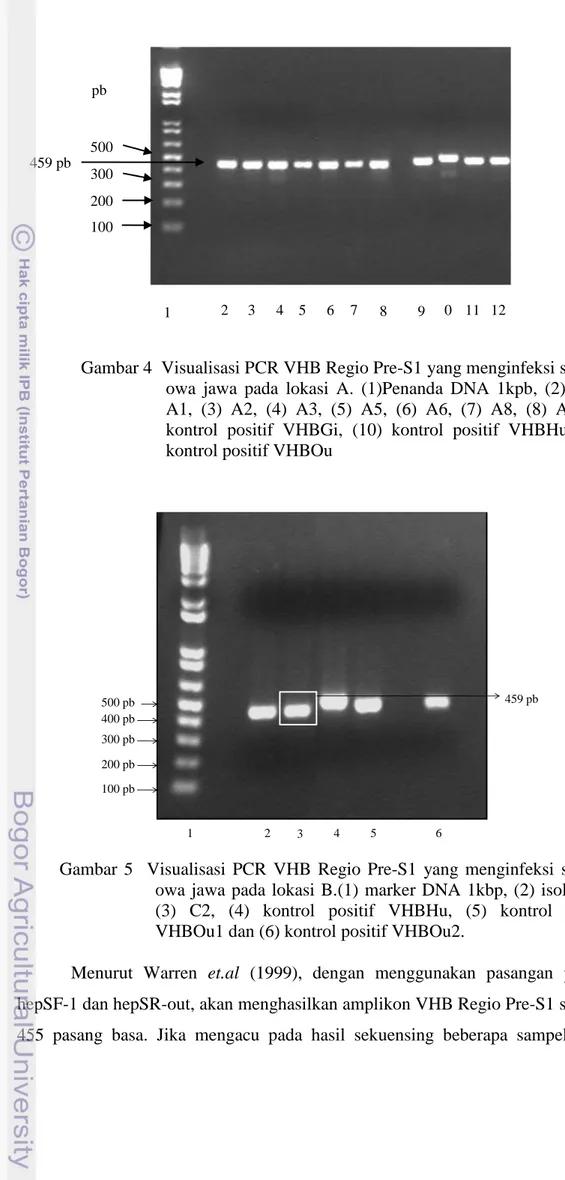
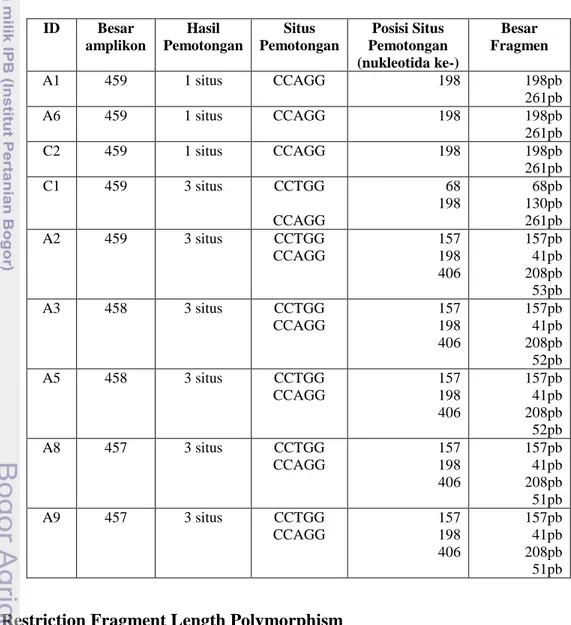
Sebagai pembanding dalam amplifikasi digunakan kontrol positif yang berasal dari pasien manusia dan orangutan penderita hepatitis B. Gambar 4 dan 5 menunjukkan hasil amplifikasi virus hepatitis B gibbon yang dilalukan pada gel agarosa menggunakan elektroforesis horizontal. Pada kontrol positif pasien manusia, terlihat bahwa pita DNA isolat VHB manusia berada di atas pita DNA dari isolat kontrol positif DNA VHB orangutan maupun sampel penelitian. Posisi pita isolat kontrol positif VHB orangutan berada di antara pita isolat VHB sampel dan kontrol positif VHB manusia. Hasil PCR memperlihatkan bahwa ketujuh (Gambar 4) dan kedua (Gambar 5) sampel DNA owa jawa teramplifikasi dengan baik menggunakan pasangan primer untuk regio pre-S1 VHB dengan ukuran sekitar 459 pasang basa.
Gambar 4 Visualisasi PCR VHB Regio Pre-S1 yang menginfeksi spesies owa jawa pada lokasi A. (1)Penanda DNA 1kpb, (2) isolat A1, (3) A2, (4) A3, (5) A5, (6) A6, (7) A8, (8) A9, (9) kontrol positif VHBGi, (10) kontrol positif VHBHu, (11) kontrol positif VHBOu
100 pb 300 pb 200 pb 400 pb 500 pb 1 2 3 4 5 6 459 pb
Gambar 5 Visualisasi PCR VHB Regio Pre-S1 yang menginfeksi spesies owa jawa pada lokasi B.(1) marker DNA 1kbp, (2) isolat C1, (3) C2, (4) kontrol positif VHBHu, (5) kontrol positif VHBOu1 dan (6) kontrol positif VHBOu2.
Menurut Warren et.al (1999), dengan menggunakan pasangan primer hepSF-1 dan hepSR-out, akan menghasilkan amplikon VHB Regio Pre-S1 sebesar 455 pasang basa. Jika mengacu pada hasil sekuensing beberapa sampel yang
3 4 6 1 9 10 1 2 3 5 7 8 11 12 300 200 100 500 pb 459 pb
dilakukan di Macrogen (Korea), didapatkan bahwa besar masing-masing amplikon berbeda-beda (Tabel 4). Dengan melakukan pensejajaran sekuens terhadap semua sekuens isolat sampel VHB menggunakan program ClustalW2 (Larkin MA, et al. 2007) terlihat adanya keragaman pada besar amplikon, hal ini mungkin disebabkan adanya insersi ataupun delesi nukleotida dari masing-masing produk PCR. Data sekuen isolat VHB bervariasi antara 457pb, 458pb dan 459 pb, berbeda dengan target produk yang diharapkan sebesar 455pb.
Tabel 4 Data sekuens produk PCR VHBGi regio Pre-S1 dan situs pemotongan dari enzim restriksi BSt2UI
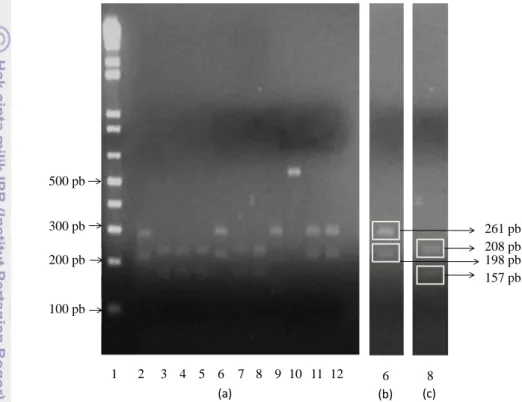
ID Besar amplikon Hasil Pemotongan Situs Pemotongan Posisi Situs Pemotongan (nukleotida ke-) Besar Fragmen A1 459 1 situs CCAGG 198 198pb 261pb A6 459 1 situs CCAGG 198 198pb 261pb C2 459 1 situs CCAGG 198 198pb 261pb C1 459 3 situs CCTGG CCAGG 68 198 68pb 130pb 261pb A2 459 3 situs CCTGG CCAGG 157 198 406 157pb 41pb 208pb 53pb A3 458 3 situs CCTGG CCAGG 157 198 406 157pb 41pb 208pb 52pb A5 458 3 situs CCTGG CCAGG 157 198 406 157pb 41pb 208pb 52pb A8 457 3 situs CCTGG CCAGG 157 198 406 157pb 41pb 208pb 51pb A9 457 3 situs CCTGG CCAGG 157 198 406 157pb 41pb 208pb 51pb
Restriction Fragment Length Polymorphism
Pemotongan produk PCR regio Pre-S1 dari VHB menggunakan enzim restriksi Bst2UI menghasilkan fragmentasi produk PCR dengan situs pemotongan tertentu (CCA/TGG) sehingga dapat diidentifikasi isolat virus hepatitis B dari
spesies tertentu. Hasil potongan enzim restriksi (Gambar 6 dan 7) memperlihatkan bahwa pada 9 isolat owa jawa terjadi fragmentasi DNA, demikian pula pada kontrol positif VHB gibbon dan orangutan. Namun pada kontrol VHB manusia tidak terjadi pemotongan regio Pre-S1.
10 11 500 pb 300 pb 200 pb 100 pb 1 2 3 4 5 6 7 8 9 12 261 pb 208 pb 198 pb 157 pb 8 6 (a) (b) (c)
Gambar 6 Visualisasi produk DNA menggunakan enzim restriksi BSt2UI. (a). Baris 1(penanda DNA 1kpb Invitrogen, USA), Baris 2-8 (isolat hasil studi), Baris 9 kontrol positif VHBGi, Baris 10 kontrol positif VHBHu, Baris 11 kontrol positif VHBOu1, Baris 12 kontrol positif VHBOu2. (b). Isolat A6. (c). Isolat A9
Gambar 6a memperlihatkan adanya keragaman posisi pemotongan produk PCR regio Pre-S1 VHB yang diisolasi. Sampel A1 dan A6, selanjutnya dikelompokkan sebagai kelompok I, memperlihatkan posisi pemotongan yang hampir sama dengan kontrol positif VHB regio Pre-S1orangutan. Sedangkan sampel A2, A3, A5, A8 dan A9, selanjutnya dikelompokkan sebagai kelompok II, mempunyai posisi pemotongan yang berbeda dengan kontrol positif VHB regio pre-S1gibbon maupun orangutan. Kelompok I terlihat terpotong menjadi 2 fragmen yaitu 198 pb dan 261 (Gambar 6b), pada kelompok II juga terlihat
terpotong menjadi 2 fragmen namun dengan besar fragmen yang berbeda dengan kelompok I yaitu 208 pb dan 157 pb (Gambar 6c).
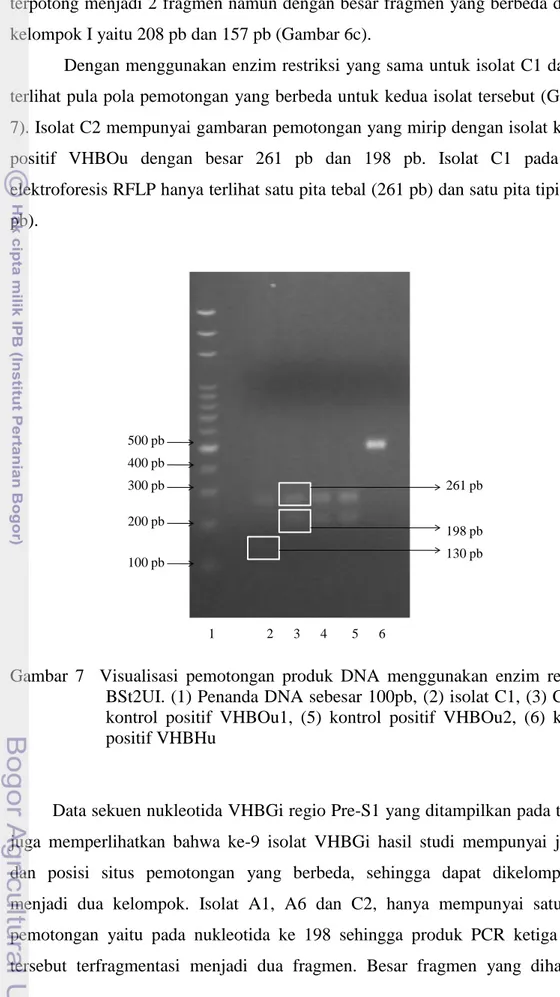
Dengan menggunakan enzim restriksi yang sama untuk isolat C1 dan C2, terlihat pula pola pemotongan yang berbeda untuk kedua isolat tersebut (Gambar 7). Isolat C2 mempunyai gambaran pemotongan yang mirip dengan isolat kontrol positif VHBOu dengan besar 261 pb dan 198 pb. Isolat C1 pada hasil elektroforesis RFLP hanya terlihat satu pita tebal (261 pb) dan satu pita tipis (130 pb). 500 pb 200 pb 400 pb 300 pb 100 pb 261 pb 130 pb 198 pb 2 1 3 4 5 6
Gambar 7 Visualisasi pemotongan produk DNA menggunakan enzim restriksi BSt2UI. (1) Penanda DNA sebesar 100pb, (2) isolat C1, (3) C2, (4) kontrol positif VHBOu1, (5) kontrol positif VHBOu2, (6) kontrol positif VHBHu
Data sekuen nukleotida VHBGi regio Pre-S1 yang ditampilkan pada tabel 3 juga memperlihatkan bahwa ke-9 isolat VHBGi hasil studi mempunyai jumlah dan posisi situs pemotongan yang berbeda, sehingga dapat dikelompokkan menjadi dua kelompok. Isolat A1, A6 dan C2, hanya mempunyai satu situs pemotongan yaitu pada nukleotida ke 198 sehingga produk PCR ketiga isolat tersebut terfragmentasi menjadi dua fragmen. Besar fragmen yang dihasilkan
adalah 198 pb dan 261 pb. Isolat C1 mempunyai 2 situs pemotongan yaitu pada nukleotida ke 68 dan 198, dengan besar fragmen 68 pb, 130 pb dan 261 pb.
Produk PCR isolat A2, A3, A5, A8 dan A9 mempunyai 3 situs pemotongan dan terfragmentasi menjadi 4 fragmen. Hasil pemotongan pada kelima isolat tersebut berada pada situs nukleotida ke 157, 198 dan 406. Besar fragmen yang terbentuk adalah 157 pb, 41 pb, 208 pb dan 53 pb. Hasil elektroforesis RFLP (Gambar 6a) tidak terlihat dua fragmen terkecil dari kelompok ini (41 pb dan 53 pb). Hal ini disebabkan karena dua fragmen tersebut mempunyai ukuran di bawah 100 pb, sehingga dengan menggunakan elektroforesis agarosa diduga mungkin bermigrasi melewati batas bawah gel.
Pembuatan Pohon Filogenetik
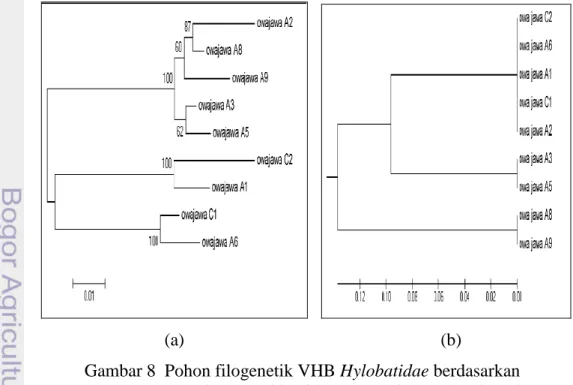
Pensejajaran sekuens nukleotida VHBGi regio Pre-S1 untuk membangun pohon filogenetik menggunakan perangkat lunak ClustalW dan Mega (versi 5) dengan menyandingkan data sekuens nukleotida HBVGi Pre-S1 isolat hasil studi. Menggunakan data sekuens nukleotida VHBGi Pre-S1 isolat hasil studi (459 nukleotida) dan hasil pemotongan sekuens menggunakan enzim restriksi BSt2UI dapat dibentuk dua pohon filogenetik.
(a) (b)
Gambar 8 Pohon filogenetik VHB Hylobatidae berdasarkan (a) sekuens nukleotida VHB regio Pre-S1 (459 pb)
(b) situs pemotongan sekuens nukleotida VHBregio Pre-S1 menggunakan enzim restriksi BsT2UI.
Hubungan kekerabatan virus hepatitis B regio Pre-S1 pada isolat owa jawa hasil studi diperlihatkan pada Gambar 8a. Menggunakan metode rekonstruksi filogeni neighbour-joining dengan model substitusi kimura 2-paramater (MEGA 5, 2011) terbentuk dua cluster besar yang memisahkan isolat-isolat VHBGi Pre-S1 hasil studi. Isolat VHBGi Pre-Pre-S1 owa jawa C2 dan A1 bersama-sama dengan isolat VHBGi Pre-S1 owa jawa C1 serta A6 membentuk kelompok besar terpisah dari isolat VHBGi Pre-S1 owa jawa A5, A3, A9, A8 dan A2. Dari 459 urutan basa nukleotida didapatkan situs conserved sebanyak 375 (81,6%), situs variable 85 (18,4%), situs singleton 7,2 % dan situs parsim-info sebanyak 11,2%.
Menggunakan situs-situs restriksi yang dihasilkan oleh enzim restriksi BsT2UI pada sekuens nukleotida isolat VHBGi hasil studi (lampiran 2) dapat dibentuk pohon filogeni menggunakan konstruksi unweighted pair group method
with arithmetic mean (UPGMA, metode tanpa pembobotan). Pada pohon
filogenetik yang terbentuk (Gambar 8 b), isolat VHBGi Pre-S1 owa jawa A1, C2, A6, C1 dan A2 membentuk cluster tersendiri, diikuti oleh percabangan yang dibentuk oleh isolat VHBGi Pre-S1 owa jawa A3 dan A5. Cabang terluar dibentuk oleh isolat VHBGi Pre-S1 owa jawa A8 dan A9.
Terdapat perbedaan antara gambar 8a dan 8b yaitu pada gambar 8a isolat VHBGi Pre-S1 owa jawa A2 berada pada cluster yang sama dengan A8 sementara pada gambar 8b isolat VHBGi Pre-S1 owa jawa A2 berada pada percabangan bersama isolat VHBGi Pre-S1 owa jawa C1, A6, C2 dan A1 terpisah dari isolat VHBGi Pre-S1 owa jawa A8.
Gambar 9 memperlihatkan pohon filogenetik yang menunjukkan hubungan kekerabatan berdasarkan sekuens nukleotida antara isolat VHBGi regio Pre-S1 owa jawa di Indonesia dengan data isolat satwa primata lainnya dari GeneBank. Isolat owa jawa hasil studi membentuk cluster besar tersendiri, terpisah dari cluster isolat Hylobates lar (no. akses HQ603076), orangutan (no. akses AF193864) dan H. pileatus (no. akses AY781187). Hal ini memperlihatkan bahwa virus hepatitis B yang menginfeksi owa jawa berbeda dengan orangutan dan spesies Hylobates lainnya. Di dalam kelompok isolat hasil studi juga terbentuk pengelompokan, kelompok isolat C1, A6, C2 dan A1 berada pada percabangan yang berbeda dengan kelompok isolat A2, A8, A9, A5 dan A3. Isolat
C1 dan A6 mempunyai kekerabatan sekitar 100% demikian pula antara isolat C2 dan A1. Isolat Hylobates agilis terpisah membentuk cluster terluar pada phon filogentik tersebut.
Gambar 9 Pohon filogenetik VHB Hylobatidae dan orangutan berdasarkan sekuens sekuens nukleotida VHB regio Pre-S1(459 nukleotida). Data sekuens nukleotida VHB gibbon region Pre-S1 isolat hasil studi (459 bp) dan data isolat dari genBank di atas ditranlasikan ke dalam bentuk sekuens asam amino menggunakan program MEGA (Kumar, et al. 2011).
Gambar 10 Pohon filogenetik VHB Hylobatidae dan orangutan berdasarkan sekuens asam amino VHB regio Pre-S1 (459 nukleotida).
Hubungan kekerabatan berdasarkan sekuens asam amino diperlihatkan pada Gambar 10. Pohon filogenetik yang terbentuk sama dengan pohon
filogenetik yang dibentuk oleh sekuens nukleotida (Gambar 9). Isolat owa jawa hasil studi tetap berada pada cluster tersendiri seperti pada gambar 9. Namun
cluster isolat A6 dan C1 berada pada percabangan yang berbeda dengan kelompok
owa jawa hasil studi lainnya. Analisa asam amino menunjukkan bahwa isolat VHBGi owa jawa C1 dan A6 mempunyai kemiripan komposisi asam amino dengan isolat VHBGi dari orangutan, H. lar dan H. agilis. Perbedaan pohon filogenetik dapat terjadi karena perbedaan pembacaan basa nukleotida yang ditranslasikan menjadi asam amino. Perbedaan urutan basa nukleotida akan mempengaruhi komposisi asam amino yang terbentuk.
Dari kedua pohon filogenetik yang terbentuk dapat diperlihatkan bahwa isolat virus hepatitis B yang menginfeksi owa jawa hasil studi berbeda dengan virus hepatitis B yang menginfeksi orangutan dan dapat terdeteksi dini menggunakan primer yang mengamplifikasi regio pre-S1 dari VHB.
Sekuens Genom Lengkap Virus Hepatitis B Gibbon Amplifikasi Virus Hepatitis B Gibbon
Untuk mendapatkan sekuens genom lengkap dari virus hepatitis B Gibbon (VHBGi), dilakukan amplifikasi DNA menggunakan empat pasangan primer. Masing-masing pasangan primer mempunyai daerah overlapped pada ujung 5’ dengan ujung 3’ pada primer lainnya. Dari kesembilan isolat owa jawa hasil studi yang digunakan adalah isolat C1 dan C2.
Mengacu pada sekuens genom lengkap VHBGi (Sa-Nguanmoo et al., 2008), target produk PCR yang diharapkan pasangan primer pertama adalah sebesar 1383 pasang basa, pasangan primer II sebesar 1027 pb, pasangan III sebesar 927 pb dan pasangan primer IV sebesar 1115 pb.
Isolat C1 dan C2 diambil untuk mendapatkan data sekuens genom lengkap VHBGi karena seluruh produk PCR dapat teramplifikasi dengan baik menggunakan keempat pasangan primer tersebut (Gambar 11). Hasil sekuens masing-masing produk PCR dari Macrogen Inc. (Korea) menggunakan primer yang sama dengan yang digunakan dalam amplifikasi kemudian dianalisa menggunakan program BioEdit versi 7 (2005) dan MEGA versi 5 (2011).
R1 1397 pb 1030 pb 1121 pb 921 pb 100 pb 850 pb 1650 pb 300 pb 1 2 3 4 5 6 7 8 9
Gambar 11 Visualisasi hasil amplifikasi genom lengkap VHBGi. Baris1 Ladder DNA1 kb, Baris 2-5 isolat C1 primer set 1, 2, 3, 4. Baris 6-9 isolat C2 primer set 1, 2, 3,4
Data sekuens dari masing-masing primer dipetakan untuk mendapatkan rekonstruksi genom lengkap VHBGi dan dibuat secara linear (Gambar 12). Area hijau menunjukkan posisi hasil amplifikasi set primer I (1397 pb), area tersebut terlihat seperti terpotong dikarenakan rekonstruksi yang dibuat adalah secara linear bukan melingkar yang merupakan bentuk asli genom VHB. Area merah adalah posisi hasil amplifikasi pasangan primer kedua (1030 pb).
Gambar 12 Rekonstruksi genom lengkap VHBGi linear berdasarkan posisi primer yang digunakan.
Posisi pasangan primer ketiga ditunjukkan oleh area kuning (921 pb). Sekuens nukleotida hasil amplifikasi pasangan primer keempat (1121 pb) ditunjukkan oleh warna biru dan terlihat seperti terpotong. Hal ini karena posisi
CORE2 CORE1 X101 R5 1 201 741 1021 1559 1770 2273 2479 2817 3192 F6 X102 PreS1F
primer keempat, seperti halnya primer pertama, dimulai dari nukleotida ke 2273 dan melingkar kembali ke posisi ke 201. Hasil rekonstruksi dan pensejajaran keempat pasang primer tersebut didapatkan ukuran VHB owa jawa sebesar 3192 pb (isolat C1) dan 3188 pb (isolat C2).
Data sekuens dari masing-masing pasangan primer VHBGi (3192 pb) hasil studi disejarkan dengan data yang didapat dari GenBank yaitu isolat Ttblack (3182 pb) dengan nomor akses GenBank AY330916 (Tabel 5). Pada Tabel 5 terlihat lokasi masing-masing gen VHBGi pada genom linearnya. Gen P dan S mempunyai posisi tumpang tindih pada nukleotida ke-1 sampai 835.
Tabel 5 Data lokasi gen VHB pada isolat Ttblack (GenBank, AY330916)
Gen Posisi nukleotida Ukuran gen (bp) P 2307 – 3182 1 – 1623 2499 S 2848 – 3182 1-835 1170 X 1374 – 1838 465 C 1814 – 2542 729
Mengacu pada sekuens isolat Ttblack tersebut didapatkan posisi masing-masing pasangan primer dari urutan nukleotidanya dalam mengkode gen-gen VHBGi seperti yang tertera pada Tabel 6. Masing-masing pasangan primer mempunyai daerah yang tumpang tindih (overlapped) dengan pasangan primer lainnya. Pada primer set pertama, dengan membandingkan posisi isolat hasil studi (174-757) dengan isolat Ttblack (Tabel 5), dapat terlihat bahwa nukleotida pada posisi tersebut berada pada daerah yang mengkode gen P dan sekaligus gen S. Posisi primer set II mengkode sebagian daerah gen P dan S. Demikian pula primer set 3 dan 4, mempunyai daerah yang tumpang tindih dengan gen lainnya.
Potongan-potongan sekuens primer kemudian dirangkaikan untuk mendapatkan daerah-daerah yang mengalami overlapping pada ujung-ujung 5’ dan 3’. Dengan mengacu pada sekuens genom lengkap VHBGi pada Hylobates
pilleatus data GenBank (nomor akses AY781186) dapat dilihat posisi overlapping
1374
2848 1835
Tabel 6 Daerah gen VHBGi berdasarkan pasangan primer yang digunakan
Primer Set Primer Target produk Gen Besar isolat C1 Besar isolat C2 I PreS1F R5 1397 pb S/P 1397 pb 1317 pb II F6 X102 1030 pb P/S 1030 pb 999 pb III X101 CORE2 921 pb C/P X/C 921 pb 891 pb IV CORE1 R1 1121 pb P/S 1121 pb 1071 pb
Gambar 13 memperlihatkan posisi masing-masing open reading frame (ORF) dari genom lengkap VHBGi berdasarkan data sekuens nukleotida dari isolat C1. Area hijau menunjukkan ORF S yang mengkode gen S (pre-S1, pre-S2 dan S), terbagi menjadi dua bagian karena genom VHB yang sebenarnya berbentuk sirkular dibuat menjadi linear, demikian pula area kuning yang menunjukkan ORF P yang mengkode gen P. Warna merah menunjukkan ORF X yang mengkode gen X. Sedangkan posisi pasangan primer ketiga yang mengkode gen C (Core dan pre-Core) ditunjukkan oleh area biru.
S S
C X
P P
Gambar 13 Prediksi genom linier sekuens sampel C1 dibandingkan dengan sekuens H. pileatus dari GenBank (no akses AY781186)
berdasarkan posisi open reading frame.
Hasil penggabungan empat pasang primer menghasilkan genom lengkap VHBGi isolate C1 yang terdiri dari 3192 nukleotida, sedangkan pada isolat C2 dihasilkan genom lengkap VHBGi yang terdiri dari 3188 nukleotida. Dengan menggunakan sekuens genom lengkap VHBGi, isolat C1 dibandingkan dengan C2 mempunyai perbedaan sebesar 1%. Hal ini disebabkan adanya insersi maupun delesi nukleotida baik pada sekuens isolat C1 maupun isolat C2.
1623 1 1000 2000 3182 1 1 1838 1814 2542 3182
Pohon Filogenetik
Menggunakan data sekuens VHBGi genom lengkap yang didapatkan dari GeneBank dan perangkat lunak komputer MEGA dapat dikonstruksi pohon filogenetik yang memperlihatkan hubungan kekerabatan VHB antar spesies. Data GeneBank yang digunakan adalah sekuens VHB genom lengkap dari isolat VHB asal manusia (no. akses GQ 358158, HM 750156, GQ 358157, GQ 358156), orangutan (no. akses AF 193864, AF 193863), Hylobates pileatus (no. akses AY 781187, AY 781187, AB 037928), H. agilis (no. akses FM209513), dan gorila (FJ798097) serta isolat HBV dari woolly monkey (AF 046996). Pohon filogenetik yang digunakan adalah neighbour-joining dengan model substitusi kimura-2
paramater menggunakan bootsrap 1000 kali (MEGA, 2011).
Gambar 14 Pohon filogenetik VHB genom lengkap asal satwa primata dan manusia (3129 nukleotida)
Gambar 14 memperlihatkan bahwa isolat VHBGi hasil studi (H. moloch C1 dan C2) berada pada satu percabangan dan mempunyai kedekatan genetik sebesar 99% di antara keduanya. Kedua isolat tersebut mempunyai kekerabatan yang lebih dekat dengan isolat dari orangutan karena berada pada percabangan pohon yang sama dibandingkan dengan isolat dari spesies Hylobates lainnya. Isolat
orangutan berada pada cluster tersendiri bersama dengan isolat H. agillis. Isolat VHB manusia membentuk cluster tersendiri, terpisah dari cluster VHB pada satwa primata. Hal ini menunjukkan bahwa kekerabatan VHB manusia jauh dari VHB yang berasal dari satwa primata. Isolat Lagothrix lagothrica (woolly
monkey) digunakan sebagai outgroup dalam pembentukan pohon filogenetik ini. Lagothrix lagotricha sampai saat ini diketahui sebagai satu-satunya spesies satwa