Harimau Sumatera (Panthera tigris sumatrae)
Klasifikasi harimau berdasarkan Nowak & Paradiso (1983) adalah sebagai berikut: Kingdom Animal, Filum Chordata, Subfilum Vertebrata, Kelas Mammalia, Subkelas Placentalia, Ordo Carnivora, Familia Falidae, Genus Panthera, Spesies Panthera tigris.
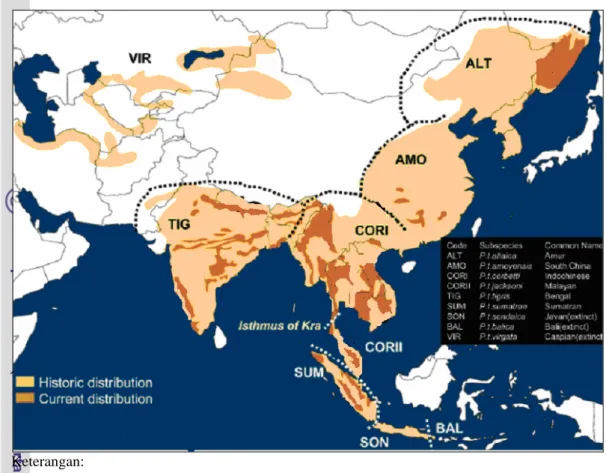
Asal usul harimau berdasarkan bukti fosil yang ditemukan, diduga berasal dari daerah China Utara pada masa Pleistocene awal (± 1,3 juta-2,1 juta tahun yang lalu) yang kemudian menyebar ke berbagai daerah (Kitchener 1999). Lou et al. (2004) menjelaskan bahwa berdasarkan daerah penyebaran dan morfologinya terdapat sembilan subspesies harimau di seluruh dunia. Enam subspesies harimau yang keberadaanya masih ada sampai sekarang adalah (1) Panthera tigris altaica (Harimau Siberia), (2) P. t. amoyensis (Harimau China Selatan), (3) P. t. tigris (Harimau India atau Benggala), (4) P. t. corbetti (Harimau Indo-China), (5) P. t. jacksoni (Harimau Malayan), dan (6) P. t. Sumatrae (Harimau Sumatera). Sedangkan tiga subspesies harimau yang saat ini keberadaannya sudah punah adalah (1) P. t. virgata (Harimau Kaspia), (2) P. t. sondaica (Harimau Jawa), dan (3) P. t. balica (Harimau Bali). Gambar sembilan subspesies harimau di dunia dan daerah penyebarannya ditunjukkan pada Gambar 1 dan 2.
(1) P. t. altaica (2) P. t. amoyensis (3) P. t. tigris (4) P. t. corbetti (5) P. t. jacksoni
(6) P. t. sumatrae (7) P. t. virgata (8) P. t. sondaica (9) P. t. balica
Keterangan:
ALT (P. t. altaica, daerah penyebaran di Rusia, Korea), AMO (P. t. amoyensis, daerah penyebaran di China Selatan), TIG (P. t. tigris, daerah penyebaran di India, Nepal, Bangladesh, Bhutan), COR I (P. t. corbetti, daerah penyebaran di China, Myanmar, Thailand, Vietnam, Laos, Kambodia), COR II (P. t. jacksoni, daerah penyebaran di Semenanjung Malaysia), SUM (P. t. sumatrae,
daerah penyebaran di Pulau Sumatera-Indonesia), VIR (P. t. virgata, daerah penyebarannya di Afghanistan, Iran, Turki, Mongolia), SON (P. t. sondaica, daerah penyebarannya di Pulau Jawa-Indonesia), BAL (P. t. balica, daerah penyebarannya di Pulau Bali-Indonesia).
Daerah warna coklat muda = daerah sejarah distribusi; Daerah warna coklat tua = daerah distribusi saat ini
Gambar 2 Daerah penyebaran sembilan subspesies harimau di dunia (Sumber: Lou et al. 2004)
Selain berdasarkan daerah penyebarannya, harimau di dunia dapat dibedakan berdasarkan perbedaan morfologi harimau seperti pada Tabel 1.
Tabel 1 Perbedaan morfologi sembilan subspesies harimau di dunia
No Subspesies Morfologi Umum
1 P. t. altaica
(Harimau Siberia)
• Ukuran panjang jantan 2,70-3,30 m dan betina 2,40-2,75 m.
• Ukuran berat jantan 180-306 kg dan betina 100-167 kg.
• Merupakan harimau terbesar di antara subspesies harimau. Dengan tubuh yang besar dan berlemak berguna untuk menahan panas tubuh.
• Mempuyai rambut yang panjang dan tebal dan berwarna kuning terang dan putih hal ini merupakan adaptasi terhadap lingkungan yang dingin dan bersalju.
• Terdapat rambut ekstra pada telapak kaki untuk menahan dingin.
• Mempunyai sedikit loreng pada tubuh, berwarna kecoklatan.
• Hampir tidak mempunyai loreng pada kaki depan.
Lanjutan Tabel 1
No Subspesies Morfologi umum
2 P. t. amoyensis
(Harimau China Selatan)
• Ukuran panjang jantan 2,30-2,60 m dan betina 2,20-2,40 m.
• Ukuran berat jantan 130-175 kg dan betina 100-115 kg.
• Rambut pada tubuh berwarna kuning gelap.
• Jarak antar loreng berdekatan.
• Ujung ekor tidak meruncing. 3 P. t. tigris
(Harimau Benggala)
• Ukuran panjang jantan 2,70-3,10 m dan betina 2,40-2,65 m.
• Ukuran berat jantan 180-258 kg dan betina 100-160 kg.
• Ukuran tubuh No. 2 terbesar setelah Harimau Siberia.
• Mempunyai rambut tubuh yang bervariasi warnanya, ada yang kuning dan putih dengan loreng berwarna hitam atau kecoklatan.
• Harimau putih mempunyai mata berwarna biru, hidung merah muda, dan loreng berwarna gelap. 4 P. t. corbetti
(Harimau Indo China)
• Ukuran panjang jantan 2,57-2,84 m dan betina 2,31-2,64 m.
• Ukuran berat jantan 150-195 kg dan betina 100-130 kg.
• Rambut pada tubuh berwarna kuning gelap, tapi lebih terang daripada Harimau Sumatera.
• Mempunyai loreng berwarna kehitaman.
• Kaki bagian dalam, perut, leher, dan pipi mempunyai warna dominan putih.
• Jantan mempunyai kumis lebih banyak daripada betina. 5 P. t. jacksoni
(Harimau Malayan)
• Ukuran panjang jantan 2,20-2,55 m dan betina 2,15-2,30 m.
• Ukuran berat jantan ± 120 kg dan betina ± 110 kg.
• Rambut pada tubuh berwarna kuning gelap, tapi lebih terang daripada Harimau Sumatera.
• Mempunyai loreng berwarna kehitaman.
• Kaki bagian dalam, perut, leher, pipi mempunyai warna dominan putih.
• Jantan mempunyai kumis lebih banyak daripada betina. 6 P. t. sumatrae
(Harimau Sumatera)
• Ukuran panjang jantan 2,20-2,55 m dan betina 2,15-2,30 m.
• Ukuran berat jantan 100-140 kg dan betina 75-110 kg.
• Merupakan harimau terkecil di antara subspesies harimau yang ada saat ini.
• Rambut tubuh berwarna kekuningan dan putih serta mempunyai banyak loreng berwarna hitam yang jaraknya berdekatan.
• Pada bagian kaki mempunyai belang, telapak kaki berambut dan mempunyai bantalan, cakar tajam dapat ditarik masuk.
• Rambut tengkuk pendek.
• Mempuyai kumis yang panjang berfungsi sebagai sensor.
• Mempunyai gigi taring yang besar.
• Mata bulat besar, iris berwarna kuning, penglihatan tajam..
• Panjang ekor ½ dari panjang tubuh berfungsi untuk keseimbangan 7 P. t. virgata
(Harimau Kaspia)
• Ukuran panjang jantan 2,64-2,95 m dan betina 2,41-2,59 m.
• Ukuran berat jantan 169-240 kg dan betina 85-135 kg.
• Mempunyai badan yang besar dan panjang dengan kaki yang kuat dan cakar yang besar.
• Saat musim dingin, rambut pada badan tebal dan panjang, pendek pada bagian leher.
• Rambut berwarna kuning dengan loreng kuning kehitaman.
• Telinga pendek dan kecil tanpa rambut di ujungnya.
• Bagian muka banyak ditumbuhi rambut 8 P. t. sondaica
(Harimau Jawa)
• Ukuran panjang jantan sekitar 2,46 m
• Ukuran berat jantan 100-141 kg dan betina 75-115 kg.
• Morfologi mirip dengan Harimau Sumatra tapi loreng lebih banyak dan mempunyai warna lebih gelap.
• Mempunyai kumis yang sangat panjang dibanding subspesies harimau yang lain. 9 P. t. balica
(Harimau Bali)
• Ukuran panjang jantan 2,20-2,31 m dan betina 1,91-2,11 m.
• Ukuran berat jantan 90-100 kg dan betina 65-80 kg.
• Merupakan harimau yang terkecil dibanding subspesies harimau yang lain.
• Rambut pada tubuh pendek tapi tebal dengan warna kuning gelap dan berloreng hitam.
Indonesia mempunyai tiga subspesies harimau endemik. Harimau Jawa dan Harimau Bali sudah dinyatakan punah, hanya Harimau Sumatera yang keberadaannya masih ada sampai sekarang. Di Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong-Bogor, terdapat koleksi spesimen awetan satu ekor Harimau Bali dan lima ekor Harimau Jawa berupa tengkorak dan kulit.
Harimau Sumatera hidup soliter dan dapat ditemukan pada berbagai tipe habitat mulai ketinggian 0-2000 m dpl (di atas permukaan laut) seperti di dataran pantai berawa, daerah payau, hutan primer, hutan sekunder, padang rumput, lahan pertanian, dan perkebunan masyarakat (O’Brien et al. 2003). Secara umum, harimau memangsa babi hutan (Sus scrofa), rusa sambar (Cervus unicolor), kijang (Muntiacus muntjak), kancil (Tragulus javanicus), dan beruk (Macaca nemestrina) (O’Brien et al. 2003).
Karena semakin terbatasnya habitat maka penyebaran Harimau Sumatera sekarang ini hanya terbatas di daerah Gunung Leuser, Bukit Tiga Puluh, Kerinci-Seblat, Berbak, Way Kambas, dan Bukit Barisan Selatan (STT 2007) (Gambar 3).
Keterangan:
1. Gunung Leuser 2. Bukit Tiga Puluh 3. Kerinci-Seblat 4. Berbak 5. Way Kambas 6. Bukit Barisan Selatan
1 6 4 2 3 5
Gambar 3 Daerah penyebaran Harimau Sumatera di Pulau Sumatera (Sumber: Shepherd & Magnus 2004)
Saat ini Harimau Sumatera semakin menurun jumlahnya sehingga dikhawatirkan mengalami kelangkaan. Tumbelaka (2001) menjelaskan lima faktor yang menyebabkan Harimau Sumatera dan satwa liar lainnya mengalami proses kelangkaan yaitu (1) Penyebaran yang terbatas. Hal ini disebabkan adanya pengurangan lahan hutan dan fragmentasi habitat sebagai hasil dari pembukaan hutan, perladangan dan pemukiman. Dengan demikian terjadi pengkonsentrasian populasi pada wilayah terbatas. Sebagai akibatnya adalah penurunan daya tampung dan peningkatan inbreeding coefficient (perkawinan silang dalam) yang memungkinkan terjadinya penurunan daya reproduksi dan daya tahan terhadap penyakit sehingga kepunahan tidak dapat dihindari; (2) Ukuran tubuh yang besar. Harimau Sumatera dengan tubuh yang besar memerlukan habitat yang relatif luas, sehingga dengan adanya pengurangan tempat hidup maka ancaman kepunahan dapat terjadi; (3) Nilai ekonomis yang tinggi. Perburuan satwa liar akan meningkat apabila satwa itu banyak diminati pasar. Kulit harimau harganya mahal dan diminati untuk koleksi atau pajangan, selain itu banyak anggota tubuh harimau yang dipercaya bermanfaat sebagai obat sehingga Harimau Sumatera banyak diburu dan sekarang menjadi langka; (4) Merupakan rantai makanan. Adanya perburuan terhadap hewan-hewan yang menjadi mangsa Harimau Sumatera oleh manusia akan memberikan dampak terus berkurangnnya sumber makanan bagi Harimau Sumatera; (5) Habitat yang jenuh. Fragmentasi habitat menyebabkan semakin tingginya kepadatan satwa yang ada di dalam wilayah tersebut. Hal ini dikarenakan berkurangnya kawasan tempat hidup dan tempat menyebar, sehingga hanya ada di tempat tertentu saja. Habitat yang jenuh akan berdampak pada peningkatan kompetisi intraspesies untuk memenuhi kebutuhan hidup satwa di dalam kawasan tersebut.
Kegiatan konservasi perlu dilakukan agar kepunahan Harimau Sumatera dapat dicegah karena Harimau Sumatera memiliki beberapa fungsi penting yaitu: (1) Fungsi ekologis. Sebagai top predator yang menempati posisi puncak pada rantai makanan. Harimau berfungsi sebagai pengendali (bioregulator) terhadap satwa-satwa di bawahnya, seperti hewan herbivora, yaitu terutama hewan ungulata yang memiliki daya survivorship dan reproduksi yang sangat tinggi, contohnya babi hutan. Walaupun tidak ada kaitannya langsung dengan fungsi
ekologis, namun dalam konteks konservasi, harimau sering diistilahkan sebagai umbrella species, yaitu jenis satwa yang apabila diselamatkan maka otomatis akan menyelamatkan satwa-satwa lain yang daerah jelajahnya sempit. Harimau memiliki daerah jelajah sangat luas (wide-ranging species atau landscape species) dibandingkan dengan satwa terestrial lain yang ada di Sumatera; (2) Fungsi ekonomis. Di beberapa negara, seperti Nepal, India, dan Buthan, keberadaan harimau sebagai satwa kharismatik di suatu kawasan konservasi yang menjadi tujuan wisata alam. Sehingga keberadaannya akan dapat meningkatkan daya tarik kawasan tersebut bagi turis mancanegara. Harimau juga menjadi salah satu daya pikat utama yang ditawarkan oleh berbagai lembaga konservasi ex-situ (misalnya kebun binatang, taman safari, dan taman nasional), baik nasional maupun internasional; (3) Fungsi sosial. Di Indonesia harimau merupakan salah satu jenis satwa yang paling banyak digunakan secara turun temurun untuk berbagai simbol dan tujuan sosial, seperti simbol perkumpulan (Siliwangi) dan tarian tradisional (reog Ponorogo); (4) Fungsi politis. Harimau merupakan satwa yang dilindungi undang-undang Indonesia dan konvensi internasional seperti IUCN (International Union for Conservation of Nature and Natural Resources) dan CITES. Oleh karena itu, kinerja konservasi Harimau Sumatera sangat mempengaruhi citra bangsa Indonesia di mata dunia internasional. Indonesia sudah memperoleh citra tidak baik secara politis akibat ketidakmampuannya dalam mencegah kepunahan global Harimau Jawa dan Harimau Bali. Oleh karena itu keberhasilan konservasi Harimau Sumatera akan meningkatkan citra Indonesia di mata internasional dalam upaya konservasi keanekaragaman hayati (Azhar 2008a).
Konservasi Harimau Sumatera
Konservasi satwa liar (termasuk Harimau Sumatera) merupakan kegiatan yang meliputi perlindungan, pengawetan, pemeliharaan, rehabilitasi, introduksi, pelestarian, pemanfaatan, dan pengembangan satwa liar. Konservasi satwa liar bertujuan menjamin kelangsungan hidup satwa liar tersebut dan menjamin kebutuhan masyarakat untuk memanfaatkannya baik secara langsung maupun tidak langsung berdasarkan prinsip kelestarian (Alikodra 2002).
Pada tahun 1994 Departemen Kehutanan Indonesia mengeluarkan dokumen
panduan konservasi Harimau Sumatera dengan empat strategi utama yaitu (1) Pengembangan dan pengelolaan konservasi populasi Harimau Sumatera; (2) Pengamanan dan perlindungan populasi Harimau Sumatera yang masih ada di habitatnya; (3) Mengembangkan penangkaran Harimau Sumatera untuk mendukung pemulihan populasi di alam; (4) Membangun network untuk kelestarian Harimau Sumatera di Indonesia (Azhar 2008b). Kemudian pada tahun 2007 dikeluarkan Peraturan Menteri Kehutanan Nomor: P.42/Menhut-II/2007 tentang Strategi dan Rencana Aksi Konservasi Harimau Sumatera 2007-2017 yang bertujuan (1) Populasi Harimau Sumatera beserta bentang alamnya di seluruh Sumatera menjadi pulih kembali dan dapat dipertahankan atau bertambah dengan dukungan berbagai pihak; (2) Terbangunnya infrastruktur dan meningkatnya kapasitas Departemen Kehutanan dalam pemantauan dan evaluasi
terhadap upaya konservasi Harimau Sumatera dan satwa mangsanya; (3) Penguatan pengelolaan Harimau Sumatera di luar kawasan konservasi dan
keterlibatan para pihak dalam mendorong konservasi Harimau Sumatera dan habitatnya baik di tingkat regional maupun nasional; (4) Terbangunnya jejaring kerja dan infrastruktur komunikasi serta terciptanya kelompok masyarakat yang
peduli dan bertanggungjawab terhadap kelestarian Harimau Sumatera; (5) Terbangunnya program konservasi ex-situ yang bermanfaat dan selaras dengan
upaya kelestarian Harimau Sumatera di alam (Dephut 2007).
Sudah lebih dari satu dekade ini, studi genetik digunakan untuk mendapatkan informasi yang lebih mendalam dalam pengambilan keputusan yang berhubungan dengan mempertahankan keberadaan suatu spesies di alam. Dengan studi genetik, informasi tentang keragaman antar individu di dalam dan antar populasi, terutama pada spesies-spesies yang terancam punah dapat diketahui (Hedrick 2001, Sunnuck 2000). Perkembangan teknik molekuler dewasa ini, seperti penemuan teknik Polymerase Chain Reaction (PCR) yang mampu mengamplifikasi untai DNA hingga mencapai konsentrasi tertentu, penggunaan untai DNA lestari (conserved) sebagai marka dalam proses PCR, penemuan lokus mikrosatelit yang hipervariabel dan penemuan metode sekuencing DNA telah menyebabkan ilmu genetik molekuler mempunyai pengaruh yang sangat besar
dalam studi biologi suatu populasi (Sunnuck 2000). Terobosan-terobosan ini, bersamaan dengan berkembangnya teknik pemodelan matematika melalui program-program komputer telah mempermudah para peneliti untuk mendapatkan data genetik suatu populasi yang sangat berguna dalam merancang program konservasi suatu spesies tertentu.
Penerapan studi genetik dalam permasalahan konservasi didasari oleh teori genetika populasi yang mempelajari tentang faktor-faktor yang menentukan komposisi genetik suatu populasi dan bagaimana faktor-faktor tersebut berperan dalam proses evolusi (Halliburton 2004). Terdapat beberapa faktor yang sangat berperan dalam kejadian evolusi pada suatu populasi, yaitu mutasi, rekombinasi, seleksi alam, genetic drift, gene flow, dan perkawinan yang tidak acak. Faktor-faktor tersebut akan memunculkan keragaman genetik pada suatu populasi, dan keragaman genetik merupakan informasi yang paling berguna untuk memahami informasi tentang kekuatan-kekuatan yang menyebabkan evolusi (Cavalli-Sforza 1998). Memahami dan mempertahankan keragaman genetik suatu populasi sangat penting dalam konservasi karena keragaman genetik yang tinggi akan sangat membantu suatu populasi beradaptasi terhadap perubahan-perubahan yang terjadi di lingkungan sekitarnya (Rhymer 1999).
Mutasi didefinisikan sebagai segala perubahan di dalam material genetik yang akan diturunkan ke generasi selanjutnya. Mutasi dapat disebabkan oleh beberapa faktor, diantaranya adalah kesalahan saat proses replikasi DNA, unequal crossing over, chromosome breakage atau meiotic disjunction. Mutasi merupakan sumber utama keragaman genetik. Tanpa mutasi, tidak akan ada keragaman genetik dan berarti tidak akan ada evolusi.
Rekombinasi merupakan penyebab kedua munculnya keragaman genetik. Rekombinasi menyebabkan kombinasi baru suatu pasangan alel, tetapi tidak memunculkan alel baru. Kombinasi baru suatu pasangan alel dapat memicu munculnya fenotip baru pada suatu individu.
Seleksi alam merupakan ide yang dilontarkan oleh Charles Darwin yang menyatakan bahwa suatu individu yang memiliki keragaman yang menguntungkan akan bertahan hidup dan bereproduksi lebih baik dibandingkan dengan individu lain di dalam populasi. Mereka akan memiliki keturunan yang
lebih banyak, dan menurunkan keragaman yang dimilikinya pada keturunannya tersebut. Oleh karena itu, keragaman ini akan semakin banyak pada generasi berikutnya.
Genetic drift adalah perubahan frekuensi alel sepanjang waktu sebagai akibat dari pemilihan gamet dan gen-gennya secara acak pada saat proses pembentukan suatu generasi baru berlangsung. Dalam jangka panjang genetic drift memiliki efek yang bertolak belakang dengan mutasi. Genetic drift yang menyebabkan perubahan frekuensi alel dalam jumlah besar dan berlangsung secara tiba-tiba akan menurunkan keragaman genetik di dalam populasi dan akan memunculkan perbedaan genetik antar populasi.
Gene flow akan terjadi pada saat suatu individu berpindah dari satu area ke area lain, bahkan antar populasi. Jika individu-individu tsb dapat bertahan hidup dan bereproduksi di tempat barunya, maka mereka akan memasukkan gen-gen mereka ke dalam populasi barunya dan pada saat inilah gene flow terjadi. Gene flow menyebabkan populasi-populasi memiliki material genetik yang semakin mirip satu sama lain dan dalam jangka panjang gene flow akan menimbulkan efek yang bertolak belakang dengan genetic drift.
Perkawinan yang tidak acak terjadi jika individu-individu yang melakukan perkawinan merupakan individu-individu yang secara genetik saling berhubungan satu sama lain.
Prinsip dasar dalam genetik populasi adalah prinsip Hardy-Weinberg. Prinsip Hardy-Weinberg menduga bahwa dalam kondisi tertentu, frekuensi alel dan genotipe akan tetap konstan atau seimbang dalam populasi, dan keduanya saling berhubungan satu sama lain. Kondisi-kondisi tertentu yang dimaksud dalam prinsip Hardy-Weinberg meliputi reproduksi antar individu yang dilakukan secara seksual dan acak, tidak ada seleksi alam, kejadian mutasi diabaikan, tidak ada individu yang masuk atau keluar dari suatu populasi dan ukuran populasi yang cukup besar. Jika kondisi-kondisi ini terpenuhi oleh suatu populasi, maka populasi tersebut disebut sebagai populasi yang berada dalam keseimbangan Hardy-Weinberg (Hardy-Hardy-Weinberg Equilibrium). Keseimbangan Hardy-Hardy-Weinberg sangat penting di dalam konservasi dan kejadian evolusi genetik, karena penyimpangan
dari keseimbangan Hardy-Weinberg ini merupakan dasar untuk mendeteksi kejadian inbreeding, fragmentasi populasi, migrasi, dan seleksi (Hartl & Clark 1997).
DNA Mitokondria
Pada umumnya material DNA yang digunakan dalam analisis genetik berasal dari DNA inti tetapi sumber DNA dapat pula diperoleh dari organel-organel sitoplasmik. Salah satu organel-organel yang dapat menjadi sumber bahan genetik adalah mitokondria. mtDNA adalah DNA berbentuk sirkuler dan berutas ganda yang ada di dalam mitokondria yaitu organel sel yang berperan dalam metabolisme sel. Ukuran mtDNA sangat pendek, biasanya kurang dari 17.000 pb (pasang basa) dan tersusun atas gen yang mengontrol metabolisme selular. Jika setengah dari DNA inti diperoleh dari garis maternal dan setengah lagi diperoleh dari garis paternal, maka mtDNA seluruhnya berasal dari garis maternal. Dalam setiap molekul DNA terdapat ratusan kopi empat basa nitrogen yaitu Adenin (A), Guanin (G), Citosin (C) dan Timin (T). Ke empat basa tersebut berasal dari dua kelompok basa yaitu kelompok purin (terdiri dari A dan G) dan kelompok pirimidin (terdiri dari C dan T). Berdasarkan kandungan basa guaninnya, mtDNA dibagi menjadi dua untai yaitu untai yang kaya G disebut untai berat (heavy strand) dan yang mengandung sedikit G disebut untai ringan (light strand) (Reyes et al. 1998).
Berdasarkan jenis gennya, genom mitokondria dibagi menjadi dua bagian, yaitu daerah penyandi (coding region) dan daerah bukan penyandi (non coding region). Daerah penyandi terdiri dari 37 gen yaitu 13 gen penyandi protein yang berperan penting di dalam transport elektron dan fosforilasi oksidatif, dua gen penyandi rRNA (ribosomal Ribonucleic Acid), dan 22 gen penyandi tRNA (transfer RNA). Gen tersebar secara asimetris pada kedua untai DNA (Reyes et al. 1988). Untai berat mtDNA mengandung 28 gen yaitu dua gen penyandi rRNA (12S rRNA dan 16S rRNA), 12 gen penyandi protein yang terdiri dari enam NADH Dehidrogense (ND1, ND2, ND3, ND4L, ND4, ND5,), Cytochrome c Oxydase (COX1, COX2, COX3), sebuah Cytochrome b (Cyt. b), dua ATPase (ATP6, ATP8), dan 14 gen penyandi tRNA yang terdiri dari fenilalanin
(tRNAPhe), valin (tRNAVal), leusin (tRNALeu), isoleusin (tRNAIle), metionin (tRNAmet), triptofan (tRNATrp), asam aspartat (tRNAAsp), lisin (tRNALys), glisin (tRNAGly), arginin (tRNAArg), histidin (tRNAHis), serin (tRNASer), leusin (tRNALeu), dan treonin (tRNAThr). Sedangkan untai ringan mtDNA mengandung sisanya (sembilan gen) yaitu, satu gen penyandi protein yaitu NADH Dehidrogense 6 (ND6) dan delapan gen penyandi tRNA yang terdiri dari asam glutamat (tRNAGlu), prolin (tRNAPro), serin (tRNASer), tirosin (tRNATyr), sistin (tRNACys), asparagin (tRNAAsn), alanin (tRNAAla), dan glutamin (tRNAGlu). Daerah bukan penyandi genom mitokondria hanya terdiri dari daerah kontrol (control region) atau d-loop (displacement loop) (Reyes et al. 1998). Contoh struktur genom mitokondria seperti pada Felis catus (kucing) yang termasuk dalam satu famili dengan Harimau Sumatera (Famili Falidae), dapat dilihat pada Gambar 4.
Gambar 4 Struktur genom mitokondria Felis catus (Lopez et al. 1996)
Peranan Marka DNA Mitokondria dalam Konservasi Genetik
Pengkajian keragaman genetik melalui penandaan molekuler menggunakan DNA didapatkan hasil yang dapat mengungkapkan perbedaan dengan lebih teliti dalam membedakan intra dan interspesies yang menyangkut tentang struktur, komposisi, dan organisasi genom pada tingkat DNA (Duryadi 1994).
Duryadi (1994) menjelaskan beberapa hal yang mendukung penggunaan mtDNA sebagai penanda dalam studi keragaman genetik adalah (1) Kopi mtDNA
terdapat dalam jumlah yang tinggi. Jumlah kopi yang tinggi menjadikannya mudah diisolasi dan dipurifikasi untuk berbagai keperluan analisis genom; (2) Ukuran mtDNA relatif kecil, sekitar 14-39 kb (kilobase), sehingga dapat dipelajari sebagai satu kesatuan yang utuh; (3) Bagian-bagian dari genom mitokondria berevolusi dengan kecepatan yang berbeda. Tingkat evolusi dari suatu bagian DNA merupakan faktor penting yang menentukan penggunaan penanda DNA dalam studi sistematika dan biogeografi. Gen-gen yang terkonservasi dengan baik dapat dijadikan sebagai dasar penelusuran kesamaan asal muasal (ancient taxa) sedangkan bagian yang berubah cepat digunakan untuk mengetahui seberapa cepat divergensi dalam spesies tersebut terjadi; (4) Genom mitokondria hewan berukuran kecil karena mtDNA hewan tidak memiliki intron ataupun spacer yang berukuran besar antar gennya; (5) Sifat khusus mtDNA adalah diturunkan melalui induk betinanya tanpa melalui rekombinasi, sehingga afinitas genetik yang diatur oleh genom mitokondria merupakan refleksi dari Phylogeni matriarcale; (6) Penyusun mtDNA sangat polimorf, baik untuk intrapopulasi maupun interspesies.
Duryadi (1994) juga menerangkan manfaat penggunaan mtDNA antara lain (1) Sebagai penanda genetik dalam studi variabilitas intraspesifik (interpopulasi) telah memberikan informasi secara kualitatif maupun kuantitatif. Penggunaan penanda mDNA pada Harimau menunjukkan strukturisasi geografik (Luo et al. 2004); (2) Karena mtDNA berevolusi sangat cepat maka dapat digunakan untuk melacak kejadian yang relatif baru seperti studi hibridisasi alami antara dua subspesies. (3) Sifat khusus mtDNA yaitu diturunkan melalui induk betina tanpa mengalami rekombinasi. Adanya sifat tersebut dapat digunakan untuk suatu rekonstruksi historik dari genealogi matrilinier suatu spesies maupun antar populasi yang ada (Luo et al. 2004); (4) Studi keragaman genetik intraspesifik berdasarkan perbedaan dan persamaan mtDNA dapat menghasilkan rekonstruksi filogenik dari beberapa spesies yang saling berdekatan. Dengan demikian, berdasarkan penanda genetik dapat diketahui proses terjadinya pemecahan dari spesies yang satu terhadap yang lainnya (Janczewski et al. 1995, Zhang et al. 2006).
Masing-masing gen mitokondria memiliki laju evolusi dengan kecepatan yang berbeda. Ada yang laju perubahannya pelan sehingga daerahnya bersifat conserve (contohnya 16S rRNA dan 12S rRNA), ada yang laju perubahannya sedang sehingga ada daerah yang bersifat conserve dan ada sebagian daerah yang bersifat variatif (sebagai contoh Cyt. b), ada yang laju perubahannya cepat sehingga daerahnya bersift variatif (CO I dan D-loop). Beberapa marka mtDNA telah digunakan dalam analisis genetik suatu populasi. Pemilihan marka genetik DNA disesuaikan dengan tujuan analisis yang akan dicapai.
Gen penyandi Cyt. b mempunyai ukuran 1140 pb. Adanya variasi urutan letak pada Cyt. b menyebabkan gen ini banyak digunakan untuk membandingkan spesies dalam genus yang sama atau famili yang sama (Randi 1996). Protein Cyt. b dibagi dalam tiga domain fungsional, yaitu intermembran, matriks dan transmembran. Domain intermembran terbentang antara membran dalam dan luar mitokondria, mengalami evolusi yang lebih lambat daripada ke dua domain fungsional lainnya. Berdasar jajaran urutan basa gen Cyt b, daerah tersebut dapat memberikan informasi filogenetik pada tingkat intraspesies sampai pada tingkat antar genus.
Daerah bukan penyandi (non coding region) yaitu D-loop merupakan genom yang tidak membawa urutan informasi untuk pembentukan protein maupun RNA, namun daerah ini merupakan situs replikasi dari mtDNA secara umum. D-loop merupakan bagian dari mtDNA yang sangat variatif dalam substitusi nukleotida, insersi atau delesi (indel) dan memiliki VNTRs (Variable Number Tandem Repeat) yang dinamis yang terletak pada bagian yang hipervariatif dan domain yang khusus (Fumagalli et al. 1996). Daerah D-loop dibagi menjadi tiga domain yaitu, CSB (Conserved Sequence Block) dan HVS-II. Domain I yang berbatasan dengan tRNAPro, terdiri dari runutan yang diasosiasikan dengan Termination of H-strand Replication (TAS) yang sering mengandung VNTRs (R1 repeats). Domain II yang terdapat di bagian sentral dan bersifat kekal, terdapat daerah Conserved Sequence Block (CSB B, C, D, E dan F). Domain III yang berbatasan dengan tRNAPhe, terdiri dari runutan yang variabilitasnya tinggi karena substitusi nukleotida, insersi dan delesi (indels) serta VNTRs (R2 repeat) dan runutan
nukleotida bersifat kekal yang merupakan promotor untuk transkripsi untai berat (heavy strand) dan untai ringan (light strand) (Sbisa et al. 1997).
Tingginya angka dari polimorfisme nukleotida atau adanya perbedaan runutan pada kedua bagian hipervariabel dari non coding region digunakan untuk membedakan di antara individu suatu spesies (Melton 1999). Selain itu analisis derah D-loop digunakan juga untuk melihat keragaman antar subspesies ataupun antar populasi (Brown 1986). Daerah D-loop diketahui sangat cepat berkembang dibandingkan dengan bagian mtDNA lainnya. Hal ini karena terjadinya akumulasi substitusi basa, proses insersi dan delesi yang lajunya amat cepat bila dibandingkan dengan DNA inti (Foran et al. 1988).