KARAKTERISASI DAN IDENTIFIKASI MOLEKULER ISOLAT BT951
SERTA EVALUASI POTENSINYA SEBAGAI BAKTERI PROBIOTIK
PADA BUDIDAYA UDANG WINDU (Penaeus monodon)
M uliani, Nur baya, dan Endang Susianingsih
Balai Riset Perikanan Budidaya Air Payau
Jl. Makm ur Dg. Sit akka No. 129, Maros 90512, Sulawesi Selat an E- m ail: [email protected]
(Naskah diterima: 25 Juli 2011; Disetujui publikasi: 29 September 2011)
ABST RAK
Penelitian ini bertujuan untuk m engetahui karakteristik, dan m enentukan posisi relatif i sol at BT9 5 1 m el al u i i d en t i f i k asi m ol ek u l er m en g g u n ak an g en 1 6 S- r RNA, ser t a m engevaluasi pot ensinya sebagai probiot ik pada budidaya udang windu (P. Monodon). Penelitian ini m eliputi tahapan: (1) isolasi dan karakterisasi secara m orfologi; (2) uji fisiologi dan biokim ia; (3) identifikasi isolat BT951 secara m olekuler; (4) uji sensitifitas t erhadap ant ibiot ik ; (5) uj i daya ham bat isolat BT951 t erhadap V. harveyi; (6) uj i patogenisitas isolat BT951 terhadap larva udang; (7) uji tantang isolat BT951 dengan V. harveyi; (8) uji ketahanan isolat BT951 pada suhu ruang dan suhu 4oC. Hasil penelitian m enunjukkan bahwa, secara m orfologi isolat BT951 koloninya berbentuk bulat kecil, berwarna krem kekuningan, t epian licin, dan cem bung, t erm asuk kelom pok bakt eri gram posit if yang berbent uk bat ang, oksidase dan kat alase posit if, indol negat if, dan bersifat m ot il. Bakt eri ini dapat t um buh pada suhu 40oC, salinit as 0- 50 ppt , dan sensit if terhadap beberapa jenis antibiotik seperti rifanpisin, eritrom isin, dan kloram penikol akan t et api resit en t erhadap am pisilin, oksit et rasiklin, gent am isin, dan furazolidon. Berdasarkan hasil analisis gen 16S- rRNA m enunjukkan bahwa isolat BT951 term asuk kelom pok Brevibacillus laterosporus dengan indeks kedekat an sebesar 97,7%. Bakt eri ini t idak bersifat pat ogen t erhadap udang windu, dapat m engham bat pert um buhan bakteri V. harveyi, dan pot ensial dijadikan sebagai bakt eri probiot ik pada budidaya udang windu.
KATA KUNCI: isolat BT 9 5 1 , p r ob iot ik , Br evib acillus lat er osp or us, Penaeus m o n o d o n
ABST RACT : Char act er iz at ion and m olecular id ent if icat ion of BT 9 5 1 and e v a l u a t i o n o f i t s p o t e n t i a l a s p r o b i o t i c b a ct e r i a i n t i g e r shr im p (Penaeus monodon) cult ur e. By: M uliani, Nur baya, and En d a n g Su si a n i n g si h
(7) in vitro and in vivo challenge test of BT951 bacteria against V. harveyi; (8) survival test of BT951 at room and 4oC temperatures. Morphological characterization showed that the colonies of BT951 were small, circular, yellowish-white, has smooth edges, and convex shape. Physiological and biochemical test showed that BT951 isolate was gram positive, short-rod shape, catalase and cytocrome oxidase positive, indole negative, non motile, protelytic, Lysine and arginin Decaroxilase positive. This bacteria can still grow at 40oC, salinity of 0-50 ppt, and sensitive to rifampicine, erythromicine, and chloramphenicol, but resistant to ampicilin, oxytetracyline, gentamicine, dan furazolidone. Base on 16S-rRNA sequencing, BT951 isolate was closely related (97.7%) to DNA sequence of Brevibacillus laterosporus. This bacteria is not pathogenic to tiger shrimp larvae, known to have inhibitory effect on the growth of V. harveyi, and potential as probiotic bacteria in tiger shrimp culture.
KEYWORD S: i so l a t BT 9 5 1 , Pr o b i o t i k , Brevibacillus laterosporus, Penaeus monodon
PENDAHULUAN
Keberhasilan budidaya udang di t am bak masih terkendala oleh serangan penyakit yang m asi h saj a t er u s t er j ad i m esk i p u n t el ah dilakukan berbagai upaya pencegahan dan penanggulangan. Serangan penyakit tersebut t idak hanya t erjadi pada level budidaya akan t et api juga pada level perbenihan. Di t am bak, serangan penyakit umumnya disebabkan oleh serangan virus. Ada beberapa jenis virus yang sering t erident if ikasi m enginf eksi udang di t am bak di ant aranya, MBV, IHHNV, HPV, dan WSSV. Nam u n d i an t ar a j en i s- j en i s vi r u s t ersebut , WSSV m erupakan penyebab ut am a t erjadinya kem at ian udang di t am bak yang beref ek t erhadap m erosot nya indust ri per-t am b ak an u d an g (Leon ar d o eper-t al., 2 0 0 5 ; Balasubram anian et al., 2008). Jenis virus ini t idak hanya m enyerang udang windu t et api j u g a u d an g van am e yan g saat i n i t el ah dibudidayakan secara meluas di hampir seluruh pertambakan di Indonbesia. Serangan penyakit di level perbenihan masih saja disebabkan oleh serangan bakt eri berpendar yait u V. harveyi. Mesk i p u n ser i n g d i t em u k an ad an ya l ar va udang yang positif terinfeksi oleh WSSV, akan t et api kem at ian m assal benur di perbenihan akibat serangan virus ini masih jarang, bahkan ham pir t idak pernah ada laporan akan hal t ersebut .
Ber b ag ai u p ay a p en c eg ah an d an p en an g g u l an g an p en yak i t p ad a b u d i d aya udang windu telah dilakukan, namun hasilnya b el u m m em b er i k an h asi l yan g m ak si m al seh i n g g a ser an g an p en yak i t m asi h t er u s t erjadi. Serangkaian penelit ian m aupun uji cob a t elah d ilak uk an unt uk m end ap at k an m et od e p encegahan d an p enanggulangan
penyakit pada udang windu antara lain dengan m enggunak an obat - obat an dan ant ibiot ik sebagai anti bakteri (Karunasagar et al., 1994). Pengelolaan lim bah budidaya udang dengan m enggunakan t andon dan biofilt er juga t elah banyak dilakukan (Chanratchakool et al., 1995; Muliani et al., 1998a). Upaya lain yang t elah dilakukan unt uk pencegahan penyakit pada b u d i d ay a u d an g w i n d u ad al ah d en g an m erangsang kekebalan non- spesif ik m elalui penggunaan vaksin dan im unostim ulan (Itam i & Takashi, 1991; Sung et al., 1994; Devaraja et al., 1998; Salf ira, 1998; Vargas- Albores et al., 1998), penggunaan ektrak mikroalga untuk merangsang kekebalan dan resistensi terhadap p en y ak i t (Su p am at t ay a et al., 2 0 0 5 ), penggunaan probiot ik (Aust in & Day, 1990; Lavilla- Pit ogo et al., 1998; Rengpipat et al., 1998; Maeda, 1999), serta penggunaan bahan ak t if sponge dan hydrozoan sebagai ant i-bakt eri (Ahm ad et al., 1995; Muliani et al., 1998b; Suryati et al., 2000). Nam un dem ikian usaha tersebut belum membuahkan hasil yang m aksim al dan serangan penyakit m asih saja terus terjadi.
harveyi d ar i ai r l aut , ai r t am b ak , d an ai r p em el i h ar aan l ar va. Hal a (1 9 9 9 ) m en g e-m uk ak an bahwa V. e-metschnicovii Z dan M yang diisolasi dari larva udang sehat ef ekt if m engham bat pert um buhan dan pelekat an V. carchariae YA32.2 (pat ogen t erhadap larva udang) dan dapat m engurangi jum lah larva udang yang m at i. Haryant i et al. (2000) t elah m engisolasi t iga isolat bakt eri yang m am pu m engham bat pert um buhan V. harveyi pada m edium agar dalam cawan pet ri dan sat u di ant ara t iga isolat t ersebut m am pu m enekan p er t u m b u h an V. harveyi p ad a m ed i a pemeliharaan larva udang. Sedangkan Muliani et al. (2003), t elah m engisolasi sedikit nya 15 isolat bakt eri laut , baik dari sedim en, air, dan k ar ang yang p ot ensi al m eng ham b at p er -t um buhan bak-t eri V. harveyi pa-t ogen udang windu. Macey & Coyne (2005) dan Iehat a et al. (2010) m engisolasi probiot ik dari abalon, Merrif ield et al. (2010) m engisolasi probiot ik dari lingkungan budidaya ikan salm on, Avella et al. (2010) m engisolasi probiot ik dari larva g i l t h ead sea b r eam (Sparus aurata, L), s ed an g k an Bj o r n s d o t t i r et al . , (2 0 1 0 ) m enisolasi probiot ik dari larva ikan sebelah (Atlantic halibut).
Pemanfaatan bakteri- bakteri probiotik yang diisolasi dari berbagai sumber belum maksimal karena adanya beberapa kendala di ant ara-nya opt im alisasi pert um buhan, kem am puan b er ad ap t asi, d an k onsist ensi ant ivib r iosis yang dim iliki belum diket ahui. Kem am puan t um buh dan berkem bang suat u jenis bakt eri probiotik sangat dipengaruhi oleh asal bakteri tersebut diisolasi. Untuk lebih mengoptimalkan pert um buhan dan perkem bangan probiot ik yang digunakan pada budidaya udang windu, maka dicoba mengisolasi bakteri dari sedimen t am bak . Menurut Poernom o (2004) bahwa probiotik yang diaplikasikan ke dalam tambak harus mampu hidup di dalam tambak, mampu tumbuh, mampu berkembang biak, dan mampu berfungsi/ bekerja akt if pada bidang m asing-m asing sesuai yang diharapkan. Oleh karena it u, karakt erisasi dan ident if ikasi baik secara f i si ol og i m au p u n b i ok i m i a ser t a eval u asi pot ensinya sebagai probiot ik dari suat u jenis b ak t er i p er l u d i l ak u k an u n t u k l eb i h m e-m ak s i e-m al k an p en g g u n aan n y a s eb ag ai probiot ik di t am bak. Penelit ian bert ujuan ini untuk mengetahui karakteristik, mengevaluasi p o t en s i n y a s eb ag ai p r o b i o t i k , s er t a menentukan posisi relatif isolat BT951 melalui analisis gen 16S- rRNA.
BAHAN DAN METODE
Isolasi dan Karakterisasi Secara M or f ol og i
Sedim en t am bak diam bil m enggunakan bot ol sam pel st eril kurang lebih 10 g, di-m asukkan ke daladi-m coolbox dan selanjut nya dibawa ke laboroat orium Pat ologi, Balai Riset Perikanan Budidaya Air Payau, Maros. Sedimen t am bak dit im bang sebanyak 1 g, kem udian d i en c er k an d en g an l ar u t an f i s i o l o g i s (NaCl8,5%) secara berseri (bisa sam pai 10- 4 tergantung kondisi sedimen tambak, semakin t inggi kadar bahan organik di dasar t am bak m aka t ingkat pengenceran sem akin t inggi, k ar ena b iasanya p op ulasi b ak t er inya j uga sem ak i n t i n g g i ). Sel an j u t n ya d ar i set i ap pengenceran diam bil 100 m L dan disebar pada media Triptic Soy Agar (TSA) dalam cawan pet ri, kem udian diinkubasi pada suhu ruang selam a 1- 2 hari. Koloni bakt eri yang t um buh selanjutnya diidentifikasi berdasarkan bentuk, warna, elevasi, dan ukuran koloni bakteri yang t erbent uk (At las, 1997; Prescot t et al., 2002). Bakteri yang tum buh dim urnikan dan dikultur pada m edia TSA yang dim iringkan, diinkubasi selam a 48 jam dan selanjut nya diuji secara fisiologi dan biokimia.
Karakterisasi Fisiologi dan Biokimia
Karakt erisasi fisiologi dan biokim ia isolat BT951 yang dilakukan adalah pewarnaan gram, oksidase, kat alase, indol, m ot ilit as, akt ivit as am ilolit ik , ak t ivit as prot ease, dan ak t ivit as kitinase (Hadioetomo, 1993; Muir, 1996).
Identifikasi Isolat BT9 5 1 Secara M olek uler
Unt uk m enent ukan ident it as isolat BT951 b er d asar k an sek uen 1 6 S- r RNA, d ilak uk an an al i si s yan g m el i p u t i b eb er ap a t ah ap an sesuai dengan metode yang dikemukakan oleh Marchesi et al. (1998) dan t elah dim odif ikasi oleh Suwant o et al. (2 0 0 0 ) yait u m elip ut i ek st r ak si DNA, am p lif ik asi gen 1 6 S- r RNA dengan PCR, gene clean dengan m etode glass milk, Cycle sequensing d en g an m et o d e BigDye, p r esi p i t asi DNA, d an sek u en si n g dengan m esin Sequenser.
Ekstraksi DNA
24 jam. Sel bakteri dipanen dengan mengambil 1 m L suspensi biakan bakt eri, lalu dim asuk-k an asuk-k e d al am Ep p end or f vol um e 1 ,5 m L. Selanjut nya disent rifugasi dengan kecepat an 6000 rpm selam a 5 m enit dan supernat annya dibuang. Hal ini dilakukan dua kali. Pelet bakteri yan g t er b en t u k k em u d i an d i r esu sp en si dengan 250 mL buffer 1xTE dan 2 mg/ L lisozim, diikubasi 37oC selam a 30 m enit . Selanjut nya dit am bahkan SDS 10% sebanyak 50 m L dan 5 m L Proteinase- K, dibolak- balik sampai bening berlendir. Cam puran ini diinkubasi pada suhu 37oC selam a satu jam (tiap 15 m enit dikocok). Set elah it u dit am bahkan 65 m L NaCl 5 M, di bolak- balik sehingga tercampur dengan baik, k em udian dit am bahk an 80 m L CTAB 10%. Dihom ogenkan sam pai berwarna put ih susu, kem udian diinkubasi pada suhu 65oC selam a 20 m enit . Selanjut nya k e dalam cam puran t er seb u t d i t am b ah k an 6 5 0 m L p h en o l : chloroform:isoamyl alkohol (25:24:1), divortex selama 30 detik, kemudian disentrifugasi pada kecepatan 5.000 rpm pada suhu ruang selama 10 m enit . Supernat an diam bil dengan m ikro p i p et d an d i p i n d ah k an k e d al am t ab u n g Eppendorf st eril yang t elah diisi dengan 600 m L isopropanol dan dibolak balik sehingga timbul benang- benang DNA dan dibiarkan pada suhu r uang selam a 3 0 m enit . Selanj ut nya disent rif ugasi dengan kecepat an 5.000 rpm pada suhu ruang dan set elah it u supernat an d i b u an g . DNA d al am b en t u k p el et d i cu ci dengan 300 m L et hanol 70%, disent rif ugasi selam a 5 m enit pada suhu ruang dan set elah i t u et an o l d i b u an g . Pel et DNA d i k er i n g -udarakan at au dengan m enggunakan pom pa vakum untuk m enguapkan etanol yang m asih t er sisa. Langk ah t er ak hir d alam ek st r ak si DNA adalah penam bahan akuabides bebas nuklease at au buffer 1x TE at au Elut ion Buffer (EB) dan selanjutnya DNA disim pan pada suhu - 20oC untuk keperluan selanjutnya.
Amplifikasi Gen 1 6 S- rRNA dengan PCR
Sebelum dilakukan am plif ikasi gen 16S-rRNA dengan PCR, t erlebih dahulu dilakukan p en g ecek an t er h ad ap D N A y an g t el ah d i ek st r ak m el al u i el ek t r of or esi s g el m i n i (Suwant o et al., 2 0 0 0 ) at au d engan gene Quant . Am plif ik asi gen 1 6 S- r RNA dengan PCR d i g u n ak an Ready to Go PCR Bead s dengan kom ponen reaksi t erdiri at as ddH2O 23 m L; DNA tem plet 0,5 m L; 1 m L 63f prim er; dan 1 m L 1387r prim er. Prim er yang diguna-kan adalah prim er universal unt uk dom ain
b ak t er i a b er u p a For war d p r i m er 6 3 f (5 ’ -CAGGCCTAACACATGCAAGTA- 3’) dan reverse prim er 1387r (GGGCGGWTGGTACAAGGC- 3’ ) ( Marchesi et al. 1998; Suwant o et al., 2000). Sem ua k om p onen r eak si d icam p ur d alam mikrotube dan selanjutnya di-running. Proses running pada PCR dilakukan dalam 30 siklus, di mana masing- masing siklus terdiri atas tahap pre start pada suhu 94oC selama 2 menit, tahap denat urasi DNA ut as ganda pada suhu 92oC selam a 30 det ik, t ahap annealing pada suhu 55oC selama 30 detik, dan tahap perpanjangan pada suhu 75oC selam a 1 m enit , t ahap post PCR pada suhu 75oC selam a 20 m enit , dan t ahap st op PCR pada suhu 4oC. Selanjut nya hasil PCR disimpan pada suhu - 20oC (Suwanto et al., 2000).
Pemurnian DNA
Pad a t ah ap an i n i DNA yan g t el ah d i -am plif ikasi dengan PCR di “running” dal-am elekt rof oresis kem udian dim urnikan dengan m et ode Glass milk.
Cycle Sequensing, Presipitasi, dan Sekuensing DNA
Pada t ahapan ini pereaksi dan langkah k erj a dilak uk an berdasark an m et ode yang dilaporkan oleh Marchesi et al. (1998) yang t elah dim odifikasi oleh Suwant o et al. (2000).
Evaluasi Potensi Isolat BT9 5 1 sebagai Probiot ik
Uji Sensitivitas terhadap Antibiotik
Uji Daya Hambat Isolat BT9 5 1 terhadap
V. harveyi
Bakt eri V. harveyi MR5339 dit um buhkan pada m edium TCBSA (Thiosulfate Citrate Bile Sucrose Agar) selam a 24 jam . Koloni t unggal yang t um buh diam bil dengan jarum Ose dan disuspensikan dalam larutan garam fisiologis (0,85% (w/ v) NaCl). Kem udian disebar pada m ed i a SWC 1 0 0 % d al am c aw an p et r i , selanj ut nya dit aruh paper disk st eril yang berdiameter 7 mm. Paper disk ditetesi dengan b i ak an i so l at BT9 5 1 seb an yak 1 0 - 2 0 L (konsentrasi biakan dalam suspensi sekitar 108 -109 cfu/ m L). Kem udian diinkubasi pada suhu 28oC selam a 2 4 - 4 8 j am , set elah it u zona hambatan yang terbentuk diukur dengan mistar penggaris pada t iga posisi dan selanjut nya diratakan.
Uji Patogenisitas Isolat BT9 5 1 terhadap Larva Udang
Konsent rasi isolat BT951 yang digunakan pada uj i pat ogenisit as adalah 106 CFU/ m L dengan metode perendaman (Hameed, 1995). Wadah yang digunakan adalah st oples kaca b er k ap asit as 3 L yang d iisi air laut st er il dengan kadar garam 28 ppt sebanyak 2 L dan dit ebar i dengan lar va udang sebanyak 2 5 ekor/ wadah.
Unt uk m enj ag a k et er sed i aan ok si g en , wadah pem eliharaan larva dilengkapi dengan aerasi. Pat ogenisit as isolat BT951 t erhadap larva udang windu diam at i m elalui kem at ian l ar va u d an g set el ah 2 4 j am p er en d am an (Rengpipat et al., 1998) dan dibandingkan dengan kontrol.
Uji Tantang Isolat BT951 dengan V. harveyi
Uji Tantang Secara In vitro
Uj i t an t an g secar a In vitro d i l ak u k an dengan menggunakan media SWC kaldu 100% (air laut 750 mL, akuades 250 mL, bakto pepton 5 g, ekst rak kham ir 1 g, gliserol 3 m L) dalam labu erlem eyer. Kepadat an bakt eri V. harveyi dibuat m enjadi 107 CFU/ m L (Rengpipat et al., 1998) dan kepadat an bakt eri BT951 dibuat m enjadi 102, 104, dan 106 CFU/ m L (Tjahjadi, 1994; Hala, 1999; Muliani et al., 2003). Biakan bakteri BT951 dimasukkan ke dalam media SWC kaldu sesuai dengan kepadat an yang t elah ditentukan, dan selanjutnya biakan V. harveyi dim asukkan ke dalam m edia t ersebut . Biakan kedua bakteri ini dikultur selam a 96 jam pada
su h u r u an g d al am i n k u b at o r b er g o yan g . Populasi V. harveyi menggunakan media SWC 100% yang t elah dit am bah rifam pisin sebagai m edia selektif.
Uji Tantang Secara In vivo
Uji t ant ang secara In vivo dilakukan di dalam akuarium kaca yang berkapasit as 3 L. Wadah t ersebut diisi air laut st eril dengan kadar garam 28 ppt sebanyak 2 L, dan ditebari larva udang st adia PL7 sebanyak 25 ekor/ wadah. Perlakuan dalam penelit ian ini t erdiri at as A) Isolat BT951 dengan kepadat an 102 CFU/ m L, B) Isolat BT951 dengan kepadat an 104 CFU/ mL, C) Isolat BT951 dengan kepadatan 106 CFU/ mL, D) Kontrol (V. harveyi 107CFU/ mL t an p a BT9 5 1 ). Masi n g - m asi n g p er l ak u an diulang tiga kali. Pengamatan populasi bakteri d an si n t asan l ar va u d an g wi n d u d i am at i setelah 96 jam.
Uji Ketahanan Isolat BT9 5 1 pada Suhu Ruang dan Suhu 4oC
Bakt eri isolat BT951 dit um buhkan pada m edia biakan m urni at au m edia kaldu m urni (nutrient broth). Wadah yang digunakan untuk m enum b uhk an isolat BT9 5 1 ad alah g elas Er l enm eyer 2 5 0 m L. Bi ak an d i t um b uhk an p ad a su h u r u an g sam b i l d i g o yan g p ad a inkubator bergoyang selama 24 jam kemudian di-sampling sebagai data pertum buhan awal. Untuk menjaga agar tidak terjadi kontaminasi, m aka biakan dibagi ke dalam botol kecil yang steril dan disimpan pada suhu ruang dan suhu 4oC. Jum lah bot ol k ecil yang berisi biak an isolat BT951 disesuaikan dengan f rekuensi sampling sehingga setiap dilakukan sampling b i ak an u n t u k sampling b er i k u t n ya t i d ak t erganggu, dan biakan yang t ersisa set elah di-sampling langsung dimusnahkan.
Pen yi m p an an i so l at BT9 5 1 d i l ak u k an selam a 10 bulan, dan dilak uk an sampling pertumbuhan satu kali dalam sebulan. HASIL DAN BAHASAN
Morf ologi BT9 5 1
Gambar 1. Koloni isolat BT951 setelah 72 jam inkubasi pada media TSA
Figure 1. Colony of BT951 isolate after 72 hours incubation on TSA media
Sif at Fisiologi dan Biok im iaw i Isolat BT951
Beberapa sifat fisiologi dan biokimia isolat BT951 disajikan pada Table 1. Pada Tabel 1 t er l i h at b ah w a i so l at BT 9 5 1 t er m asu k kelompok bakteri gram positif yang berbentuk bat ang, Oksidase dan kat alase posit if , indol negat if , dan bersif at m ot il. Bakt eri ini dapat tum buh pada suhu 40oC, tum buh pada m edia yang dibubuhi NaCl 0- 10%. Bakteri ini sensitif t erhadap beberapa j enis ant ibiot ik sepert i rif anpisin, erit rom isin, dan cloram penik ol, ak an t et ap i r esi t en t er h ad ap am p i si l i n , oksit et rasiklin, gent am isin, dan f urazolidon. Populasi bakt eri ini pada salinit as 0- 30 ppt set elah 24 jam m ecapai 104 CFU / m L, nam un pada salinit as yang lebih t inggi yait u 40- 50 ppt sedikit m enurun yait u 103 CFU/ m L.
Identifikasi Isolat BT9 5 1 Secara M olek uler
Dari hasil analisis gen 16S- rRNA dapat d i k et ah u i b ah w a i so l at BT9 5 1 t er m asu k kelom pok Brevibacillus laterosporus dengan indeks kedekat an sebesar 97,7%. Bakt eri ini t er m asu k d al am k i n g d o m b ak t er i , Di vi si Firm icut es, Klass Bacilli, Ordo Bacillus, Fam ily Paenibacillaceae, Genus Brevibacillus, spe-cies Brevibacillus laterosporus. Jenis bakt eri ini banyak digunak an sebagai biok ont r ol unt uk di bidang pert anian. Menurut Oliveira et al. (2 0 0 4 ), Brevibacillus laterosporus pot ensial sebagai agen biologi kont rol.
Hasil Uji Patogenisitas terhadap Larva Udang Windu
Hasi l u j i p at eg en i si t as i so l at BT9 5 1 t erhadap larva udang windu m enunj uk k an bahwa hingga kepadat an 106 CFU/ m L t idak bersifat patogen terhadap larva udang windu. Hal ini dapat diket ahui dari sint asan udang windu yang dihasilkan m encapai 96% sedikit lebih tinggi dibanding dengan sintasan udang windu pada bak yang t idak m enggunak an probiotik yaitu 92%. Beberapa hasil penelitian sebelumnya menunjukkan hal yang serupa, di m ana bakt eri yang diisolasi dari air laut dan tambak yang potensial sebagai probiotik tidak bersifat patogen pada larva udang windu. Rosa et al. (1997) melaporkan bahwa pada kepadatan 105, 106, 107 dan 108 cf u/ m L selam a 24 jam perendaman semua bakteri kandidat biokontrol yang d iuj i p at ogenisit asnya t id ak b er sif at pat ogen pada larva udang windu. Dem ikian p ul a yang d i hasi l k an ol eh Har yant i et al. (2000), isolat BY- 9 sebagai biokont rol t idak b er sif at p at ogen p ad a lar va ud ang wind u set elah inkubasi 12 jam pada konsent rasi 105 CFU/ m L. Beber apa isolat bak t er i k andidat probiot ik yang disolasi dari sedim en karang (Muliani et al., 2003) dan daun m angrove (Muliani et al., 2004) juga dilaporkan t idak pat ogen pada larva udang windu. Widanarni et al. (2010) m elaporkan bahwa ket iga isolat kandidat probiotik (isolat U, isolat 9, dan isolat P1 7 Bo) yan g d i u j i p at o g en i si t asn ya p ad a konsentrasi 106 CFU/ mL terhadap larva udang windu hingga hari ke- 5 tidak patogen terhadap larva udang windu.
Hasil Uji Daya Hambat Isolat BT9 5 1 terhadap V. harveyi
Hasi l u j i d aya h am b at i so l at BT9 5 1 terhadap V. harveyi disajikan pada Gam bar 2. Pada Gam bar 2 t erlihat bahwa isolat BT951 mampu menghambat pertumbuhan V. harveyi yang dit andai dengan adanya zona bening di sekitar koloni BT951.
Hasil Uji Tantang Secara In vitro
Tabel 1. Karakteristik fisiologi dan biokimia isolat BT951
Table 1. Physiological and biochemical characterization of BT951 isolate
Iso lat BT 951 BT 951 isola t e
+
Ampisilin 50 (mg/ mL) +
Eritromidsin 50 (mg/ mL)
-Rifampisin 50 (mg/ mL)
-Ox itetrasiklin 50 (mg/ mL) +
Gentamisin 50 (mg/ mL) +
Furaz olidon 50 (mg/ mL) +
Chloramphenikol 50 (mg/ mL)
-0% +
0.01% +
0.10% +
1% +
10% +
0 ppt 1.0x 104
10 ppt 2.5x 104
20 ppt 1.0x 104
30 ppt 1.0x 104
40 ppt 1.0x 103
50 ppt 3.1x 103
+ + + -+ -+ -+ +
-Arginin dehidr (Arginine dehydr)
Lisin dekarb (Lysine decarb) Produksi amilase (Amilolytic) Produksi protease (Proteoly tic)
Ketoglutarat (Cetoglutarate) Pertumbuhan pada salinitas y ang berbeda setelah 24 jam
Growth on different salinities after 24 hours
Heptanoat (H eptanoate) Xantin (X anthine)
Amonobutirat (Aminobutirate) Arabinosa (Arabinose) Sellubiosa (Cellubiose) Glukuronata (Glucuronate) Produksi gas (Gas from glucose) Uji O-F (O-F test)
Putrisin (Putriscine) Sukrosa (Sucrose) Indol (Indole) Uji VP (VP-test)
Pertumbuhan pada suhu 40oC (Growth at 40oC)
Pertumbuhan pada etanol 95% (Growth at 95% ethanol)
Peny ebaran (Swarming) Pendaran (Luminescence)
Ornitin dekarb (Ornithine decarb) Serin (Serine)
Pew arnaan gram (Gram staining) Oksidase (Ox idase)
Katalase (Catalase) Motilitas (Motility)
Uji f isio lo g i d an b io kimia Ph ysiolog ica l a n d b ioch em ica l t est s
Uji hambat terhadap V. harveyi (Inhibition test of BT951 against V. harveyii) Pertumbuhan pada media SWC
y ang dibubuhi antibiotik 50 mg/ mL
Growth on SWC media
supplemented with 50 mg/mL of antiobiotic
Pertumbuhan pada beberapa konsentrasi NaCl
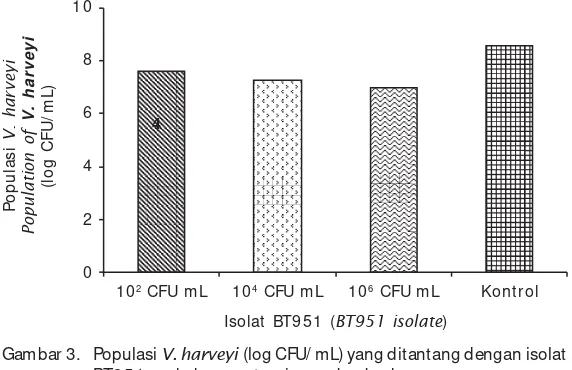
m en g g u n ak an i s o l at BT 9 5 1 d en g an konsentrasi 102 CF/ m L, 104 CFU/ m L, m aupun pada 106 CFU/ mL. Hal ini menunjukkan bahwa isolat Bt951 m am pu m enekan perkem bangan p op ulasi b ak t er i V. harveyi d alam wad ah pem eliharaan udang windu.
Sintasan Udang pada Uji Tantang Secara In vivo
Sint asan udang windu pada uji t ant ang secara In vivo disajikan pada Tabel 2. Pada Tabel 2 t erlihat bahwa sint asan larva udang wi n d u t er t i n g g i p ad a p en g g u n aan BT9 5 1
dengan konsent rasi 102 CFU/ m L. Meskipun demikian hasil uji statistik menunjukkan bahwa t idak ada per bedaan yang nyat a (P> 0 ,0 5 ) terhadap sintasan larva udang windu baik yang diberi isolat BT951 m aupun t anpa pem berian isolat BT951. Hal ini menunjukkan bahwa isolat BT951 dapat dijadikan sebagai probiotik pada budidaya udang windu.
Berdasarkan hasil sekuensing gen 16S-rRNA, isolat BT951 termasuk dalam kelompok Brevibacillus. Poernom o (2004) m elaporkan bahwa bahwa Bacillus sp. m erupakan salah satu jenis bakteri yang telah diproduksi secara Gambar 2. Hasil uji daya ham bat isolat BT951 dengan V. harveyi dengan
m etode goresan
Figure 2. Test result of BT951 inhibition to V. harveyi by scratch method V. harveyi
BT9 5 1
Gambar 3. Populasi V. harveyi (log CFU/ mL) yang ditantang dengan isolat BT951 pada konsent rasi yang berbeda
Figure 3. Population of V. harveyi (log CFU/mL) challenged with BT951 isolate
Isolat BT951 (BT951 isolate)
P
o
p
u
la
s
i
V. harveyi
Population of
V
.
h
a
rv
e
y
i
(l
o
g
C
F
U
/
m
L
)
102 CFU m L 1 0
0 8
6
4
2
4
Tabel 2. Sint asan benur windu (%) dalam uji t ant ang secara In vivo antara bakteri kandidat probiotik dengan V. harveyi
Table 2. Survival rate of tiger shrimp postlarvae (%) in vivo challenge between probiotic candidate bacteria with V. harveyi
Nilai yang diik ut i superscript serupa dalam k olom yang sam a t idak berbeda nyat a (The values followed by the same superscript in the same column were not significantly different) (P> 0.05)
Perlakuan T r ea t m en t s
Sint asan larva ud ang w ind u ( %) Sur viva l r a t e (%) of t ig er sh r im p la r va e
BT951 (102 CFU/ mL) 95.67a
BT951 (104 CFU / mL) 87.78a
BT951 (106 CFU / mL) 86.67a
Kontrol/Control 94.44a
komersil dan diaplikasikan di lapangan sebagai probiotik.
Ketahanan BT9 5 1 pada Penyimpanan dengan Suhu yang Berbeda
Isolat BT951 yang disim pan pada suhu ruang (± 28oC) selama 10 bulan penyimpanan m en g al am i p en u r u n an , n am u n d em i k i an secara keseluruhan tidak berbeda jauh dengan yan g d i si m p an p ad a su h u 4oC. Hal i n i menunjukkan bahwa dalam kondisi tidak ada alat pendingin (kulkas) isolat BT951 sebagai probiot ik m asih bisa disim pan pada kondisi suhu ruangan (± 28oC). Ini berart i jika bakt eri ini diproduksi sebagai bakteri probiotik, maka
b ak t er i i n i d ap at d i k em as d al am b en t u k probiotik cair dan dapat disimpan pada kondisi suhu ruangan, sehingga m udah penanga-nannya dan dapat digunakan di m asyarakat.
Pada um um nya probiot ik k om ersil cair yang beredar di pasaran hanya m encapai 103 CFU/ mL sebelum difermentasi meskipun pada labelnya t er t er a bahwa populasi pr obiot ik t ersebut m encapai 1011- 1012 CFU/ m L. Hal ini m en u n j u k k an b ah w a s el am a p r o s es penyim panan populasi probiot ik m engalam i penurunan. Gunarto et al. (2006) melaporkan bahwa populasi bakt eri probiot ik kom ersil (Super NB) yang dikem as dalam larut an st ok sebelum difermentasi sebesar 103 CFU/ mL dan I
P
o
p
u
la
s
i
is
o
la
t
B
T
9
5
1
Population of BT951 isolate
(l
o
g
C
F
U
/
m
L
)
Awal (Initial) 1 2
1 0 8
6 4
2
0
Wakt u sampling (setiap bulan) Sampling time (each month)
Gambar 4. Populasi isolat BT951 pada penyim panan dengan suhu yang berbeda Figure 4. Population of BT951 isolate stored at different temperatures
II III IV V VI VII VIII IX X
set elah dif erm ent asi m enggunakan t epung ik an, dedak halus, ragi, dan m olase m ak a populasinya meningkat menjadi 1010- 1012 CFU/ mL.
KESIMPULAN
Isolat BT951 t erm asuk kelom pok gram p o si t i f , b er b en t u k b at an g , Brevibacillus laterosporus, tidak bersifat patogen terhadap u d an g w i n d u , d ap at m en g h am b at p er -t um buhan bak-t eri V. harveyi, dan po-t ensial sebagai bakteri probiotik pada budidaya udang windu.
DAFTAR ACUAN
Ahmad, T., Suryati, E. , & Muliani. 1995. Screen-ing sponge for bact ericide t o be use in shrimp culture. Ind. Fish. Res. J., 1: 1- 10. Atlas, R.M. 1997. Hand Book of Microbiologicak
m edia. 2nd edition. CRC Press. Boca Raton. New York. London. Tokyo, 1706 pp. Austin, B. & Day, J.G. 1990. Inhibition of prawn
pat hogenic Vibrio spp. By a com m ercial spray- dried preparat ion of Tetraselmis suecica. Aquaculture, 90: 389- 392. Avella, M.A., Gioacchini, G., Decam p, O., &
M ar k i d i s, P. 2 0 1 0 . A p p l i cat i o n o f m u l t i sp eci es of Baci l l u s i n sea b r eam larviculture. Aquaculture, 305:12- 19. Balasubramanian, G., Sarathi, M., Venkatesan,
C., Thomas, J., & Hameed, A.S.S. 2008. Oral adm inist rat ion of ant ifiral plant ex t ract of cynodon dactylon on a large scale produc-tion against White Spote Syndorom e Virus (WSSV) in Penaeus monodon. Aquaculture, 279: 2- 5.
Bjornsdottir, R., Karadottir, E.G., Johannsdottir, J., Thorarinsdot t ir, E.E.., Sm aradot t ir, H., Sigurgisladot t ir, S., & Gudm undsdot t ir, B. K. 2010. Select ion of Bact eria and t he ef-fects of bacterial treatment of Atlantic hali-but (Hippoglossus hippoglossus L.) eggs and larvae. Aquaculture, 302: 219- 227. Chanratchakool, P., Turnbull, J. F., Smith, S. F., &
Limsuwan, C. 1995. Health management in shrim p ponds. 2nd Edition. Aquatic Anim al Healt h Research Inst it ut e. Depart m ent of Fisheries. Kaset sart Universit y Cam pus. Bangkok, 111 pp.
D evar aj a, T. N. , Ot t a, S. K. , Sh u b h a, G. , Karunasagar, I., Tauro, P., & Karunasagar, I. 1 9 9 8 . Im m u n ost i m u l an t i on of sh r i m p through oral administration of Vibrio bacteri and yeast glucan. In Flegel TW. (Ed.)
Ad-vances in shrim p biot echnology. Nat ional Cent er for Genet ic Engineering and Bio-technology, Bangkok, p. 167- 170. Devaraja, T.N., Yussoff, F. M., & Shariff, M. 2002.
Ch an g es i n b act er i al p op u l at i on s an d shrim p product ion in ponds t reat ed wit h com m ercial m icrobial product s. Aquacul-ture, 206: 245- 256.
Gunarto, Tampangallo, B.R., & Susianingsih, E. 2006. Pemeliharaan udang windu, Penaeus monodon d i l ab o r at o r i u m d en g an penambahan probiotik hasil perbanyakan. Dalam Taufiqurrohm an, M., Prayogi, U., Gim an, Winar no, A. Prosiding Seminar Nasional Kelautan III. Universit as Hang Tuah. 24 April 2006. Surabaya, hlm. 26- 33. Hadioet om o, R.S. 1993. Mikrobiologi dasar dalam praktek: Teknik dan prosedur dasar laboratorium. PT Gramedia, Jakarta, hlm. 62-68.
Hala, Y. 1999. Penggunaan gen penanda mo-lekular untuk deteksi pelekatan dan kolonisasi vibrio harveyi pada larva udang windu (Penaeus monodon) [Disert asi]. Bogor. Inst it ut Pert anian Bogor, Program Pascasarjana, 91 hlm.
Ham eed, A.S.S. 1995. Suscept ibilit y of t hree Penaeus species to a Vibrio campbelli- like bacterium. J. World Aqua. Soc., 26: 315- 319. Haryanti, Sugama, K., Tsamura, S., & Nishijima, T. 2000. Vibriost at ic bact erium isolat ed from seawat er: Pot ent ialit y as probiot ic agent in t he rearing of Penaeus monodon larvae. Ind. Fish. Res. J., 6: 26- 32.
Iehata, S., Inagaki, T., Okunishi, S., Nakano, M., Tanaka, R., & Maeda, H. 2010. Improve gut envirom ent of abalone Haliotis gigantea through Pseudiococcus sp. Ah1. treatment. Aquaculture, 305: 59- 65
It am i, T. & Takahashi, Y. 1991. Survival of larval giant tiger prawns Penaeus monodon af t er addit ion of k illed vibrio cell t o a m icroencapsulat ed diet . J. Aqua Anim. Health, 3:151- 152.
Kar u n asag ar , I., Pai , R., Mal at h i , G.R., & Karunasagar, I. 1994. Mass m ort alit y of Penaeus monodon larvae due to antibiotic-r esi st an t Vibrio harveyi i n f ect i o n . Aguaculture, 128: 203- 209.
Na-tional Center for Genetic Engineering and Biotechnology, Bangkok, p. 185- 192. Leonardo, V.A.D., Bonnichon, V., Roch, F.,
Parrinello, N., & Bonami, J.R. 2005. Compara-t ive WSSV infecCompara-t ion rouCompara-t e in Compara-t he shrim p g en er a Mar su p en aeu s an d Pal aem o n . Aquaculture, 28: 565- 569.
Macey, B.M. & Coyne, V.E. 2 0 0 5 . Im pr ove g r owt h r at e an d r esi st an ce i n f ar m ed Haliotis midae t hrough Probiot ic t reat -ment. Aquaculture, 245: 249- 261.
Maeda, M. 1999. Microbial proses in aquacul-ture. National Research Institute of Aquac-ulture. Nansei, Mie. 516- 0193. Japan, 102 pp.
Marchesi, J., Sato, R.T., Weightman, A.J., Martin, T.A., Fry, J.C., Hiom, S.J., & Wade, W.G. 1998. Design and evaluation of useful bacterium-spesifik PCR prim ers t hat am plify genes co d i n g f o r b act er i al 1 6 S- r RNA. Ap p l . Environ. Microbiol., 64: 795- 799.
Merrifield, D.L., Dimitroglou, A., Foey, A., Davies, J., Baker, R.T.M., Bogwld, J., Cast ex , M., & Ringo, E. 2010. The current status and fu-t ure focus of probiofu-t ic and prebiofu-t ic ap-plication for salmonids. Aquaculture, 302: 1- 18.
Muir, P. 1996. Ident if icat ion of Vibrio and Pseudomonas b act er ia. Dep ar t m ent of Micr ob iology, Biom ed ical and Tr op ical Veterinary Science. Jam es Cook University of North Queensland. Australia, 6 pp. Muliani, Atmomarsono, M., & Madeali, M.I. 1998a.
Pengaruh penggunaan kekerangan sebagai b i o f i l t er t er h ad ap k el i m p ah an d an k om posisi jenis bak t eri pada budidaya udang windu (Penaeus monodon) dengan sistem resirkulasi air. J. Pen. Perik. Indone-sia, 3: 54- 61.
Mu l i an i , Su r yat i , E., & Ah m ad , T. 1 9 9 8 b . Pen g g u n aan ek st r ak sp o n s u n t u k penanggulangan bakt eri Vibrio spp. pada udang windu Penaeus monodon. J. Pen. Perik. Indonesia, 1: 108- 115.
Muliani, Suwant o, A., & Hala, Y. 2003. Isolasi dan karakterisasi bakteri asal laut Sulawesi unt uk biokont rol penyakit vibriosis pada larva udang windu (Penaeus monodon Fab.). Hayati, 10: 6- 11.
Muliani, Nurbaya, Tompo, A., & Atmomarsono, M. 2004. Eksplorasi bakt eri filosfer dari t an am an m an g r o ve seb ag ai b ak t er i pr obiot ik pada budidaya udang windu Penaeus monodon, J. Pen. Perik. Indonesia, 2: 47- 57.
Oclarit, J.M., Ohta, S., Kamimura, K., Yamaoka, Y., & Ikegam i, S. 1994. Product ion of an ant ibact erial agen, o- Am iniphenol, by a bact erium isolat ed from m arine sponge, Adocia sp. Fish. Sci., 60: 559- 562. Oliveira, E.J,D., Rabinovitch, L., Monnerat, R.G.,
Passo s, L. K. J. , & Zah n er , V. 2 0 0 4 . Appl.Environ. Microbiol., 11: 6657- 6664. Prescott, L.M., Harley, J.P., & Klein, D.A. 2002.
Microbiology. 5t h edit ion. Mc Graw Hill. Boston Burr Ridge, IL Dubuque, IA Madison, WI New York , San Fransisco, Bangk ok , Bogota, Caracas, Kualalumpur, Lisbon, Lon-don, Madrid, Mex ico City, Milan, Montreal, Newd el hi , Sant i ag o, Seoul , Si ng ap or e, Sydney, Taipei, Toront o, 1026 pp. Poernom o, A. 2004. Technology of probiotics
to solve the problems in shrimp pond cul-t ure and cul-t he culcul-t ure environm encul-t . Paper presented in the National Symposium on on Development and Scientific and Tech-nology Innovation in Aquaculture, Semarang, January 27- 29, 2004, 24 pp. Ren g p i p at , S. , Ru k p r at an p o r n , S. ,
Piyatiratitivorakul, S., & Menasveta, P. 1998. Probiot ics in Aquacult ure: A case st udy of probiot ics for larvae of black t iger shrim p (Penaeus monodon). In Flegel TW. (Ed). Ad-vances in shrim p biot echnology. Nat ional Cent er for Genet ic Engineering and Bio-technology, Bangkok, p. 177- 181. Rosa, D., Zafran, Taufik, I., & Girsang, M.A. 1997.
Pen g en d al i an Vibrio harveyi secar a biologis pada larva udang windu (Penaeus monodon): I. Isolasi Bakteri Penghambat. J. Penel. Perik. Indonesia, 3: 1- 10.
Sal f i r a. 1 9 9 8 . Pengaruh pemberian LPS (Lipopolisakarida) dari dinding sel bakteri
V i b r i o h a r v eyi terhadap gambaran sistem kekebalan Non-spesifik pada udang windu (Penaeus monodon Fab.) [Tesis]. Program Pascasarjana Inst it ut Pert anian Bogor. Bogor.
Sung, H.H., Kou, G.H., & Song, Y.L. 1994. Vibriosis reVibriosist ance induced by glucan t reat -m ent in t iger shri-m p (Penaeus -monodon). Fish Phatol., 1: 11- 17.
Suryati, E., Rosmiati, Moka, W., & Hala, Y. 2000. Hydrozoan Aglaophenia sp. Bioactive Sub-st ance Analysis for Bact ericide. Ind. Fish. Res. J., 6: 55- 61.
respon and diseaseresist ance in black t i-ger shrim p (Penaeus monodon). Aquacul-ture, 248: 207- 216.
Suwant o, A., Yogiara, Suryant o, D., Tan, I., & Puspit asari, E. 2000. Select ed prot ocols. Training Course on Advances in Molecular Biology Techniques t o Assess Microbial Diversity. Bogor, 28 pp.
Tjahjadi, M.R, Angka, S.L., & Suwanto, A. 1994. Isolation and evaluation of marine bacteria for biocont rol of lum inous bact erial dis-eases i n t i g er shr i m p l ar vae (Penaeus monodon Fab . ). Asp ac. J. Mol. Biol. Biotechnol., 2: 347–352.
Vargas- Albores, F., Hernandez- Lopez, J., Gallas-Galvan, T., Montano- Perez, K., Jimenez- Vega, F., 7 Yepiz- Plascencia, G. 1998. Activation of shrim p cellular defence fungt ions by m icrobial product s. In Flegel TW. (Ed). Advences in shrim p biot echnology. Na-t ional CenNa-t er for GeneNa-t ic Engineering and Biotechnology, Bangkok, p. 161- 166. Widanarni, Rajab, F., Sukenda, & Set iawat i, M.