PRODUKSI PREBIOTIK MANOOLIGOSAKARIDA (MOS)
SERTA PENGUJIANNYA TERHADAP PERTUMBUHAN
Salmonella sp. DAN Pediococcus pentosaceus
LIA ASYARIYAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
BOGOR 2014
PERNYATAAN MENGENAI SKRIPSI
DAN SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA* Dengan ini saya menyatakan bahwa skripsi berjudul Produksi Prebiotik Manooligosakarida (MOS) serta Pengujiannya Terhadap Pertumbuhan Salmonella sp. dan Pediococcus pentosaceus adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2014
Lia Asyariyah
ABSTRAK
LIA ASYARIYAH. Produksi Prebiotik Manooligosakarida (MOS) serta Pengujiannya Terhadap Pertumbuhan Salmonella sp. dan Pediococcus
pentosaceus. Dibimbing oleh ANJA MERYANDINI dan I KOMANG G
WIRYAWAN.
Prebiotik telah direkomendasikan sebagai bahan pengganti antibiotik dalam pakan. Salah satu bahan pangan yang bisa dijadikan sebagai kandidat prebiotik adalah manooligosakarida (MOS). MOS merupakan hasil dari degradasi bakteri mananolitik. Penelitian ini bertujuan menganalisis kemampuan isolat dalam menghasilkan manooligosakarisa (MOS) dari bungkil inti sawit (BIS) serta mengujinya terhadap pertumbuhan bakteri asam laktat dan bakteri patogen. Analisis dilakukan dengan uji zona bening, aktivitas enzim, pemilihan konsentrasi substrat bungkil inti sawit (BIS), serta uji terhadap pertumbuhan bakteri asam laktat dan bakteri patogen. Isolat W6 memiliki indeks mananolitik sebesar 6.5. Hasil dari zona bening menunjukkan warna merah transparan. Isolat W6 memiliki beberapa puncak aktivitas enzim, yaitu pada jam ke 24, 30, 48, 78, dan 96 pada inkubasi kultur. Aktivitas tertinggi enzim mananase isolat W6 yaitu pada pH 5.5 dan suhu 30°C dengan waktu inkubasi 96 jam sebesar 0.0265 U/mL. Isolat ini mampu menghasilkan MOS terbaik dengan konsentrasi BIS 10%. Prebiotik MOS dapat meningkatkan pertumbuhan Pediococcus pentosaceus serta mampu menekan pertumbuhan Salmonella sp.. Dapat disimpulkan bahwa isolat W6 memiliki kemampuan untuk memproduksi MOS dengan derajat polimerasi sebesar 3.3 – 8.6 yang potensial sebagai prebiotik.
Kata kunci: bungkil inti sawit, mananolitik, manooligosakarida, prebiotik
ABSTRACT
LIA ASYARIYAH. Production of Prebiotic Mannooligosaccharide (MOS) and Test it Against the growth of Salmonella sp. dan Pediococcus pentosaceus. Supervised by ANJA MERYANDINI and I KOMANG G WIRYAWAN.
Prebiotic was recomended as a substitute for antibiotics in the feed ingredients. One of prebiotics is mannooligosaccharide (MOS). MOS is the result of bacterial degradation from mannose containing substrates. The aim of this research is to analyze the ability of the W6 to produce mannooligosaccharide prebiotics from palm kernel cake (PKC) and test it against the growth of lactic acid and pathogenic bacteria. The analyses were performed with the clearing zone test, enzyme activity, substrate concentration election of palm kernel cake (PKC), and the growth of lactic acid and pathogenic bacteria tests. Isolate W6 had mannanolytic index of 6.5. The results of the clearing zone showed a transparant red color. Isolate W6 had several peaks of enzyme activity, at 24th, 30th , 48th , 78th , and 96th hours of culture incubation. Highest mannanase activity of isolate W6 at pH 5.5 and 30° C with incubation time of 96 hours was 0.0265 U/mL. The
isolate was able to produce the best MOS with PKC concentration was 10%. The prebiotic MOS increase the growth of Pediococcus pentosaceus and was able to inhibit growth of Salmonella sp. It is concluded that W6 has the degree of polymerization at 3.3-8.6 in production of MOS as prebiotic.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
PEMBUATAN PREBIOTIK MANOOLIGOSAKARIDA (MOS)
SERTA PENGUJIANNYA TERHADAP PERTUMBUHAN
Salmonella sp. DAN Pediococcus pentosaceus
LIA ASYARIYAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Skripsi : Pembuatan Prebiotik Manooligosakarida (MOS) serta pengujiaanya Terhadap Pertumbuhan Salmonella sp. dan Pediococcus pentosaceus
Nama : Lia Asyariyah NIM : G34100025
Disetujui oleh
Prof Dr Anja Meryandini, MS Pembimbing I
Prof Dr Ir I Komang G Wiryawan Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas limpahan berkah, rahmat, dan karunia-Nya sehingga penulis dapat menyelesaikan kegiatan penelitian dan penulisan karya ilmiah ini. Penelitian ini telah dilaksanakan sejak bulan Februari hingga Juli 2014 di Laboratorium Bioteknologi Hewan dan Biomedis, PPSHB, Institut Pertanian Bogor dengan judul Pembuatan Prebiotik Manooligosakarida (MOS) serta Pengujiannya terhadap Pertumbuhan Salmonella sp. dan Pediococcus pentosaceus.
Terima kasih penulis sampaikan kepada dosen pembimbing Ibu Prof Dr Anja Meryandini, MS dan Bapak Prof Dr Ir I Komang G Wiryawan atas segala bimbingan, arahan, ilmu, dan saran selama kegiatan penelitian dan penulisan ini. Terima kasih penulis sampaikan kepada Ibu Puji Rianti, MSi yang telah menjadi penguji sidang dan pemberi saran dalam penyusunan karya ilmiah ini. Ucapan terima kasih penulis sampaikan kepada Ibu Dewi sebagai laboran Bioteknologi Hewan dan Biomedis Pusat Antar Universitas, PPSHB dan Teh Fitri atas bimbingan teknis selama bekerja di Laboratorium; Ika, Kak Tini, Kak Leni, Kak Yeni, Kak Novi, Kak Anik, Kak Rahmi, Mba Debby, Kak Fathin, Kak Ira, dan Kak Deddy sebagai teman seperjuangan atas kesetiaan, dukungan, dan bantuannya selama bekerja di Laboratorium; serta teman-teman Biologi 47 atas kebersamaanya.
Ungkapan terima kasih terbesar disampaikan kepada kedua orang tua, kakak, adik, beserta seluruh keluarga atas doa, kasih sayang, dan dukungannya hingga penulis bisa menyelesaikan strata satu. Terima kasih pula penulis sampaikan kepada program Bidikmisi dari DIKTI atas kesempatan beasiswa hingga penulis bisa menyelesaikan studi ini, serta beberapa pihak yang tidak bisa disebutkan satu persatu.
Penulis menyadari bahwa karya ilmiah ini masih banyak kekurangan. Oleh karenanya, penulis mengharapkan kritik dan saran yang membangun. Semoga karya ilmiah ini bermanfaat bagi kemajuan bidang penelitian dan perkembangan sains.
Bogor, Agustus 2014
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1 Latar Belakang 1 Perumusan Masalah 2 Tujuan Penelitian 2 Manfaat Penelitian 2 METODE 2 Waktu Penelitian 2 Bahan 3 Alat 3 Prosedur 3
Peremajaan dan Uji Zona Bening 3
Pengujian Aktivitas Enzim 3
Produksi dan Pemekatan Prebiotik 4
Uji Pengaruh Prebiotik Terhadap Pertumbuhan Bakteri Asam Laktat dan Bakteri
Patogen 4
HASIL DAN PEMBAHASAN 5
Hasil 5
Pembahasan 7
SIMPULAN DAN SARAN 10
Simpulan 10
Saran 11
DAFTAR PUSTAKA 11
LAMPIRAN 14
DAFTAR TABEL
1 Hasil hidrolisis enzim mananase isolat W6 dengan BIS 1%, 5%, dan
10% 6
2 Nilai gula pereduksi, total gula, dan derajat polimerasi hasil pemekatan
prebiotik dengan konsentrasi BIS 10% 6
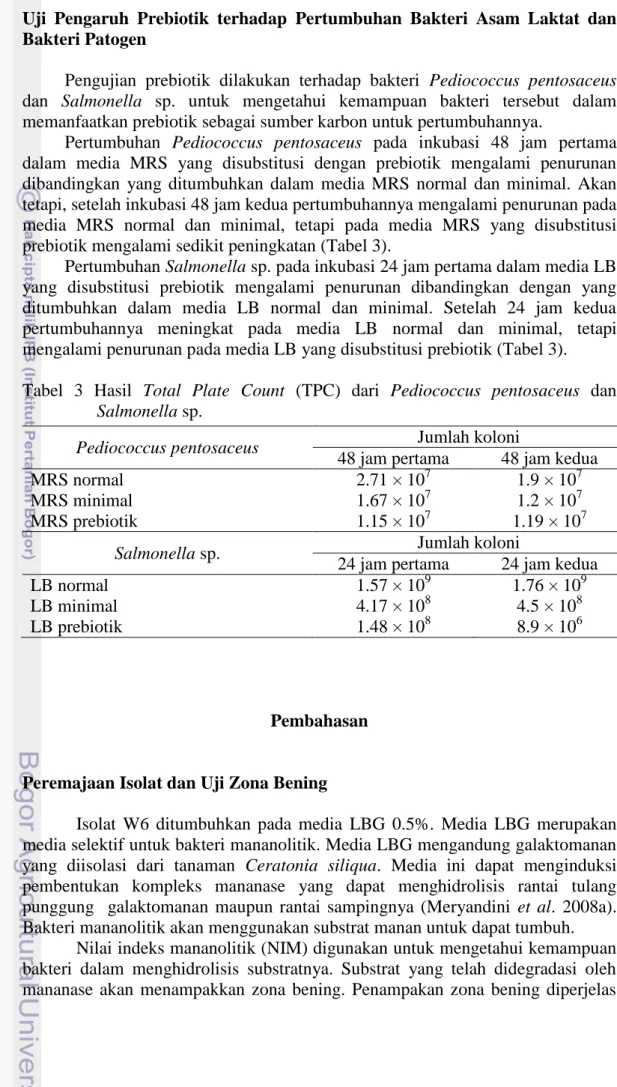
3 Hasil Total Plate Count (TPC) dari Pediococcus pentosaceus dan
Salmonella sp. 7
DAFTAR GAMBAR
1 Pembentukan zona bening isolat W6 pada media padat LBG 0.5% 5 2 Aktivitas enzim mananase isolat W6 pada substrat BIS 1% pH 5.5
dan suhu 30°C dengan inkubasi selama 96 jam 6
DAFTAR LAMPIRAN
1 Komposisi reagen dinitrosalicylic acid (DNS) 14
2 Cara perhitungan aktivitas enzim 15
3 Cara perhitungan total gula dan derajat polimerasi 16
1
PENDAHULUAN
Latar Belakang
Sumber protein dapat berupa protein nabati ataupun protein hewani. Protein hewani dapat dipenuhi oleh daging ayam, daging sapi, daging kambing, telur, dan ikan. Protein hewani tersebut dapat diperoleh dari produk ternak. Pemberian pakan pada ternak dapat mempengaruhi kualitas dari produk ternak tersebut. Banyak cara yang digunakan untuk meningkatkan produksi dan kualitas ternak, salah satunya dengan penambahan antibiotik dalam pakan. Menurut Daud (2005) penambahan antibiotik dalam pakan ternak bertujuan merangsang pertumbuhan, memperbaiki efisiensi pakan, dan menekan tingkat kematian ternak. Penambahan antibiotik tersebut menimbulkan masalah baru yaitu munculnya bakteri yang resisten terhadap antibiotik. Meskipun penggunaan antibiotik diizinkan secara legal, namun di beberapa negara telah dilarang penggunaan antibiotik sebagai bahan aditif dalam pakan ternak. Hal ini disebabkan adanya kemungkinan residu yang tertinggal dari antibiotik yang akan menjadi racun bagi konsumen dan dapat menciptakan mikroorganisme yang resisten dalam tubuh manusia maupun ternak. Oleh karena itu peternak mulai direkomendasikan menggunakan prebiotik sebagai bahan aditif untuk pakan ternak.
Prebiotik merupakan bahan makanan yang tidak dapat dicerna dan dapat meningkatkan kesehatan inangnya dengan cara merangsang pertumbuhan dan atau aktivitas satu atau beberapa bakteri dalam usus besar. Suatu bahan pangan dapat diklasifikasikan sebagai prebiotik jika tidak terhidrolisis atau terserap pada saluran pencernaan bagian atas, menjadi substrat selektif untuk satu atau beberapa jumlah bakteri menguntungkan dalam usus besar, dapat mengubah mikroflora usus besar, serta mampu menginduksi efek sistemik yang bermanfaat bagi kesehatan inang (Gibson dan Roberfroid 1995). Banyak bahan pakan sebagai kandidat yang berpotensi untuk dijadikan prebiotik. Salah satunya adalah manooligosakarida (MOS) (Iskandar et al. 2008).
Berdasarkan hasil penelitian Utami (2010) secara in vitro, prebiotik MOS dapat menekan laju pertumbuhan Escherichia coli dan dapat meningkatkan laju pertumbuhan Lactobacillus casei, sedangkan secara in vivo, MOS berpotensi sebagai antimikroba Salmonella typhimurium untuk ternak unggas (Tafsin et al. 2007). Prebiotik dapat dihasilkan dari manooligosakarida. Manooligosakarida merupakan hasil akhir dari hidrolisis manan. Manan adalah komponen utama dari bungkil inti sawit (BIS).
BIS merupakan salah satu hasil samping dari olahan minyak inti sawit. BIS mengandung air kurang dari 10%, protein 14-17%, lemak 9.5-10.5%, dan serat kasar 12-18%. BIS berpotensi menjadi pakan baik untuk ruminansia maupun nonruminansia (Iskandar et al. 2008). BIS juga memiliki kadar serat dan kadar manan yang tinggi sehingga melindungi molekul protein (Yopi et al. 2006). Hal ini menyebabkan nilai kecernaan protein menurun. Peningkatan daya cerna protein dihasilkan dari hidrolisis manan. Enzim yang berperan dalam hidrolisis manan adalah mananase.
2
Berdasarkan hasil penelitian Rahayu (2013) isolat W6 pendegradasi BIS yang memiliki aktivitas enzim mananase optimum pada pH 5.5 dan suhu 30 °C. Isolat W6 menghasilkan manooligosakarida (MOS) dengan derajat polimerasi yang baik.
Perumusan Masalah
Penelitian mengenai potensi manooligosakarida sebagai kandidat prebiotik pengganti antibiotik dalam pakan masih terbatas. Produksi manooligosakarida perlu dieksploitasi dari isolat yang potensial dalam menghirdolisis substrat BIS.
Tujuan Penelitian
Penelitian ini bertujuan menganalisis kemampuan isolat dalam menghasilkan prebiotik manooligosakarida (MOS) dari bungkil inti sawit (BIS) serta mengujinya terhadap pertumbuhan bakteri asam laktat dan bakteri patogen.
Manfaat Penelitian
Penelitian ini diharapkan dapat menambah koleksi isolat dan data mengenai potensi manooligosakarida sebagai prebiotik, serta mengeksploitasi pemanfaatan limbah pertanian seperti bungkil inti sawit.
METODE
Waktu Penelitian
Penelitian ini dilaksanakan pada bulan Februari–Juli 2014 yang bertempat di Laboratorium Bioteknologi Hewan dan Biomedis PPSHB, IPB.
3
Bahan
Bahan yang digunakan adalah isolat W6 asal Perkebunan Kelapa Sawit Cikasungka Bogor, isolat Salmonella sp., Pediococus pentosaceus, media LBG (Locust bean gum), MRS (de Man Rogosa Sharpe), LB (Luria Broth), BIS (Bungkil Inti Sawit), larutan DNS (Lampiran 1), larutan fenol, dan larutan asam sulfat.
Alat
Alat yang digunakan yaitu autoklaf, laminar air flow cabinet (LAFC), pH meter, sentrifuse, shaker, ruang asam, water bath, dan spektrofotometer.
Prosedur
Peremajaan Isolat dan Uji Zona Bening
Isolat diremajakan pada 100 mL media LBG yang di-shaker dengan kecepatan 120 rpm selama 48 jam. Media tersebut terdiri atas 0.5% LBG, 0.2% KNO3, 0.1% K2HPO4, 0.05% MgSO4, 0.05% NaCl, 0.001% FeSO4, dan 0.3%
CaCO3. Setelah itu isolat tersebut ditumbuhkan pada media padat yang sama dan
diinkubasi selama 48 jam pada suhu ruang. Koloni yang terbentuk pada media padat LBG dimurnikan dengan metode kuadran hingga diperoleh koloni tunggal. Koloni tunggal kemudian dilakukan uji zona bening pada media padat LBG yang diinkubasi selama 48 jam dengan teknik pewarnaan merah kongo 0,1% sehingga diperoleh nilai indeks mananolitik (NIM).
NIM =
Pengujian Aktivitas Enzim
Sebanyak 3 ose bakteri dari media padat LBG diinokulasikan pada 50 mL media BIS 1% untuk menjadi inokulum. Inokulum diukur pada panjang gelombang 620 nm hingga OD 0.6 – 0.8, kemudian ditumbuhkan pada media padat dan diinokulasikan sebanyak 10 mL kedalam media BIS 1%. Mulai dari jam ke 18, setiap 6 jam hingga jam ke 96 dilakukan pengujian aktivitas enzim. Uji ini dilakukan dengan cara mensentrifugasi kultur dengan kecepatan 6000 rpm selama 30 menit hingga diperoleh enzim ekstrak kasar (EEK). Supernatan (EEK) ditentukan aktivitas enzimnya pada substrat LBG 0.5% dalam bufer sitrat pH 5.5
4
yang diinkubasi selama 30 menit dengan menggunakan metode Dinitrosalisilic Acid (DNS) (Miller 1959) (Lampiran 2).
Perlakuan sampel dengan cara 0.5 mL EEK direaksikan dengan 0.5 mL substrat, setelah itu diinkubasi pada suhu 30°C selama 30 menit kemudian ditambahkan 1 mL reagen DNS. Perlakuan kontrol dengan cara 0.5 mL substrat direaksikan dengan 1 mL reagen DNS lalu ditambahkan dengan 0.5 mL EEK. Perlakuan blanko dilakukan dengan cara 0.5 mL substrat direaksikan dengan 1 mL reagen DNS lalu ditambahkan dengan 0.5 mL akuades. Perlakuan sampel, kontrol, dan blanko kemudian dipanaskan pada suhu 100°C selama 15 menit. Setelah dingin larutan diukur absorbansinya pada panjang gelombang 540 nm. Kurva standar gula pereduksi manosa dibuat dengan berbagai konsentrasi manosa dari 0 – 0.30 mg/mL dengan selang 0.05 yang direaksikan dengan 2 mL reagen DNS, lalu dipanaskan pada suhu 100°C selama 15 menit. Setelah dingin larutan diukur absorbansinya pada panjang gelombang 540 nm. Satu unit aktivitas enzim mananase didefinisikan sebagai banyaknya enzim yang dapat memproduksi 1 µmol manosa dalam 1 menit pada kondisi pengujian.
Produksi dan Pemekatan Prebiotik
EEK dari isolat potensial pada waktu produksi tertinggi direaksikan dengan berbagai konsentrasi BIS yaitu 1.0%, 5%, dan 10%. Jumlah total gula, gula perduksi, dan derajat polimerasi (DP) dihitung setelah 5 jam menggunakan metode fenol-asam sulfat (Dubois 1956). Analisis total gula dilakukan dengan cara meraksikan 1 mL sampel dengan 0.5 mL larutan fenol 0.5% dan 2.5 mL larutan asam sulfat pekat. Setelah dingin larutan diukur absorbansinya pada panjang gelombang 490 nm. Kurva standar total gula manosa dibuat dengan berbagai konsentrasi manosa dari 0 – 0.1 mg/mL dengan selang 0.01. Larutan direaksikan seperti pengujian pada sampel. Analisis gula pereduksi dilakukan dengan cara mereaksikan 1 mL sampel dengan 1 mL reagen DNS, lalu dipanaskan pada suhu 100°C selama 15 menit. Setelah dingin larutan diukur absorbansinya pada panjang gelombang 540 nm. Nilai DP diperoleh dengan cara membandingkan nilai total gula dengan gula pereduksi (Lampiran 3).
Prebiotik dengan nilai total gula tertinggi dipekatkan dengan cara diuapkan dalam air mendidih sampai berkurang sekitar setengah dari volumenya. Setelah itu dihitung jumlah total gula, gula pereduksi, dan DP dari larutan tersebut.
Uji Pengaruh Prebiotik Terhadap Pertumbuhan Bakteri Asam Laktat dan Bakteri Patogen
Prebiotik yang telah dipekatkan diuji terhadap bakteri Salmonella sp. dan
Pediococcus pentosaceus. Salmonella sp. ditumbuhkan pada media LB sedangkan Pediococcus pentosaceus ditumbuhkan pada media MRS. Pengujian prebiotik ini
dilakukan menggunakan tiga jenis media, yaitu media LB atau MRS normal, minimal, dan media yang disubstitusi prebiotik untuk memastikan bahwa bakteri menggunakan prebiotik sebagai sumber karbon untuk pertumbuhannya. Media LB atau MRS minimal dibuat dengan cara mengurangi konsentrasi pepton, ekstrak
5
khamir, glukosa, dan tripton sepersepuluh dari konentrasi normalnya (Utami 2010). Media LB atau MRS yang disubstitusi prebiotik dibuat dengan cara mengurangi konsentrasi pepton dan ekstrak khamir serta mengganti tripton pada media LB dan glukosa pada media MRS dengan prebiotik dalam konsentrasi yang sama (Lampiran 4).
Dua koloni bakteri Salmonella sp. dan Pediococcus pentosaceus secara terpisah dikultur dalam 10 mL media cair normalnya, media minimal, dan media yang disubstitusi prebiotik. Salmonella sp. diinkubasi pada suhu 37°C selama 24 jam sedangkan Pediococcus pentosaceus diinkubasi pada suhu ruang (28°C - 30°C) selama 48 jam. Kultur tersebut kemudian dicawankan pada media yang sama. Koloni hasil pencawanan ini kemudian diinokulasikan pada media yang sama dan dicawankan kembali. Jumlah koloni dihitung dengan metode TPC (Total Plate Count).
HASIL DAN PEMBAHASAN
Hasil
Peremajaan Isolat dan Uji Zona Bening
Penelitian diawali dengan meremajakan isolat terpilih, yaitu isolat W6 yang ditumbuhkan pada media LBG 0.5%. Isolat W6 memiliki ciri morfologi koloni berbentuk oval, pigmentasi coklat hilain, dan tepian licin, seperti tercantum dalam hasil penelitian Rahayu (2013).
Nilai indeks mananolitik (NIM) merupakan perbandingan antara diameter koloni dengan diameter zona bening yang dihasilkan setelah inkubasi selama 48 jam pada media padat LBG dengan pewarnaan merah kongo 0.1%. Isolat W6 ini memiliki nilai indeks mananolitik sebesar 6.5 (Gambar 1).
zona bening
Gambar 1 Pembentukan zona bening isolat W6 pada media padat LBG 0.5% setelah diinkubasi selama 48 jam pada suhu ruang dengan teknik pewarnaan merah kongo 0.1%
6
Pengujian Aktivitas Enzim
Isolat W6 memiliki beberapa puncak aktivitas enzim pada pH 5.5 dan suhu 30°C yaitu pada jam ke 24, 30, 48, 78, dan 96 pada inkubasi kultur. Aktivitas mananase optimum dari isolat W6 terdapat pada jam ke 96 sebesar 0.0265 U/mL (Gambar 2).
Gambar 2 Aktivitas enzim mananase isolat W6 pada substrat BIS 1% pH 5.5 dan suhu 30°C dengan inkubasi selama 96 jam
Produksi dan Pemekatan Prebtiok
Enzim ekstrak kasar (EEK) dari hasil produksi aktivitas enzim optimum direaksikan dengan berbagai konsentrasi BIS untuk menghasilkan produk hidrolisis sebagai kandidat prebiotik. Hasil dari reaksi menunjukkan bahwa konsentrasi substrat BIS sebanyak 10% menghasilkan nilai gula pereduksi yang paling tinggi yaitu 3.105 mg/mL (Tabel 1).
Tabel 1 Hasil hidrolisis enzim mananase isolat W6 dengan BIS 1%, 5%, dan 10% Konsentrasi BIS Gula pereduksi (mg/mL) Total gula (mg/mL) Derajat polimerasi (DP) 1% 0.395 0.77 2.0 5% 1.760 2.81 1.6 10% 3.105 5.08 1,6
Prebiotik dengan nilai total gula tertinggi yaitu dengan konsentrasi BIS 10% dipekatkan dengan cara diuapkan dalam air mendidih. Nilai total gula dan derajat polimerasi dari prebiotik setelah dipanaskan mengalami peningkatan (Tabel 2). Tabel 2 Nilai gula pereduksi, total gula, dan derajat polimerasi hasil pemekatan
prebiotik dengan konsentrasi BIS 10% Prebiotik Gula pereduksi
(mg/mL) Total gula (mg/mL) Derajat polimerasi (DP) Produksi pertama 1.4 12 8.6 Produksi kedua 3.0 9.8 3.3 0 0.005 0.01 0.015 0.02 0.025 0.03 0.035 0 18 24 30 36 42 48 54 60 66 72 78 84 90 96 Ak tiv it a s E nzim ( U/m L )
7
Uji Pengaruh Prebiotik terhadap Pertumbuhan Bakteri Asam Laktat dan Bakteri Patogen
Pengujian prebiotik dilakukan terhadap bakteri Pediococcus pentosaceus dan Salmonella sp. untuk mengetahui kemampuan bakteri tersebut dalam memanfaatkan prebiotik sebagai sumber karbon untuk pertumbuhannya.
Pertumbuhan Pediococcus pentosaceus pada inkubasi 48 jam pertama dalam media MRS yang disubstitusi dengan prebiotik mengalami penurunan dibandingkan yang ditumbuhkan dalam media MRS normal dan minimal. Akan tetapi, setelah inkubasi 48 jam kedua pertumbuhannya mengalami penurunan pada media MRS normal dan minimal, tetapi pada media MRS yang disubstitusi prebiotik mengalami sedikit peningkatan (Tabel 3).
Pertumbuhan Salmonella sp. pada inkubasi 24 jam pertama dalam media LB yang disubstitusi prebiotik mengalami penurunan dibandingkan dengan yang ditumbuhkan dalam media LB normal dan minimal. Setelah 24 jam kedua pertumbuhannya meningkat pada media LB normal dan minimal, tetapi mengalami penurunan pada media LB yang disubstitusi prebiotik (Tabel 3). Tabel 3 Hasil Total Plate Count (TPC) dari Pediococcus pentosaceus dan
Salmonella sp.
Pediococcus pentosaceus Jumlah koloni
48 jam pertama 48 jam kedua
MRS normal 2.71 × 107 1.9 × 107
MRS minimal 1.67 × 107 1.2 × 107
MRS prebiotik 1.15 × 107 1.19 × 107
Salmonella sp. Jumlah koloni
24 jam pertama 24 jam kedua
LB normal 1.57 × 109 1.76 × 109
LB minimal 4.17 × 108 4.5 × 108
LB prebiotik 1.48 × 108 8.9 × 106
Pembahasan
Peremajaan Isolat dan Uji Zona Bening
Isolat W6 ditumbuhkan pada media LBG 0.5%. Media LBG merupakan media selektif untuk bakteri mananolitik. Media LBG mengandung galaktomanan yang diisolasi dari tanaman Ceratonia siliqua. Media ini dapat menginduksi pembentukan kompleks mananase yang dapat menghidrolisis rantai tulang punggung galaktomanan maupun rantai sampingnya (Meryandini et al. 2008a). Bakteri mananolitik akan menggunakan substrat manan untuk dapat tumbuh.
Nilai indeks mananolitik (NIM) digunakan untuk mengetahui kemampuan bakteri dalam menghidrolisis substratnya. Substrat yang telah didegradasi oleh mananase akan menampakkan zona bening. Penampakan zona bening diperjelas
8
dengan penambahan Merah Kongo (Downie et al. 1994). Zona bening merupakan daerah yang berwarna bening dan berada disekeliling koloni. Merah Kongo berfungsi dalam mengikat β-1,4-D manopiranosil pada manan. Manooligosakarida (MOS) memiliki sedikit ikatan tersebut sehingga Merah Kongo tidak dapat terikat secara kuat. Pembilasan dengan NaCl 0.2 M mampu melunturkan Merah Kongo sehingga zona bening menjadi terlihat (Sumardi 2005). Hasil dari zona bening menunjukkan warna merah transparan, yang berarti bahwa bakteri ini mampu mendegradasi substrat LBG menjadi MOS.
Pengujian Aktivitas Enzim
Isolat W6 ditumbuhkan pada media cair dengan substrat BIS 1%. Keberadaan substrat dapat memacu bakteri untuk mengeluarkan metabolit selnya. Penentuan produksi enzim mananase optimum dapat dilihat dari nilai aktivitas enzim tertinggi. Satu unit aktivitas enzim mananase didefinisikan sebagai banyaknya enzim yang dapat memproduksi 1 µmol manosa dalam 1 menit pada kondisi pengujian.
Hasil penelitian Meryandini et al. (2008b) isolat RA05 memiliki aktivitas mananase sebesar 9.60 nkat/mL pada substrat bungkil kelapa 0.5%. Enzim mananase dari sumber berbeda kemungkinan memiliki karakteristik dan spesifikasi yang berbeda. Secara umum, enzim mananase menunjukkan aktivitas yang tinggi pada pH 3.0-7.5 dan suhu 45-92°C (Purnawan 2013). Mananase
Streptomyces costarianus 451-3 menunjukkan aktivitas yang tinggi pada pH 6.0
dan suhu 40°C (Meryandini et al. 2008a).
Berdasarkan hasil pengujian aktivitas enzim mananase, isolat W6 memiliki beberapa puncak aktivitas enzim pada kultur inkubasi. Hal ini dapat terjadi karena dimungkinkan enzim yang dimiliki isolat berupa isoenzim. Isoenzim adalah enzim yang mengkatalisis reaksi yang sama tapi memiliki karakteristik yang berbeda (Williams dan Wilkins 1996). Berkurangnya produk berupa manosa atau MOS akan meningkatkan aktivitas mananase karena produk ini digunakan oleh bakteri sebagai sumber karbon dan nutrisi untuk pertumbuhannya (Meryandini et al. 2008a). Aktivitas enzim dipengaruhi oleh beberapa faktor, diantaranya konsentrasi enzim, substrat, produk, senyawa inhibitor atau aktivator, pH, dan suhu (Suhartono 1989). Selain itu karena kandungan BIS yang kompleks diduga menyebabkan bakteri mengeluarkan berbagai macam metabolitnya. Bungkil kelapa mengandung serat kasar dalam bentuk β-manan dan galaktomanan (Sumardi 2005). Berdasarkan penelitian Purnawan (2013) BIS mengandung serat kasar sebanyak 18.35%. BIS mengandung 78% serat hemiselulosa dalam bentuk manan dan 12% dalam bentuk selulosa.
Manan sebagai komponen hemiselulosa dikelompokkan menjadi 4 subfamili yaitu β-manan, galaktoglukomanan, galaktomanan, dan glukomanan. β-manan (hanya terdiri atas manosa), galaktomanan (tersusun dari manosa dan galaktosa), glukomanan (tersusun dari manosa dan glukosa), galaktoglukomanan (tersusun dari manosa, galaktosa, dan glukosa) (Petkowicz et al. 2001). Aktivitas enzim dipengaruhi oleh kelengkapan dari enzim yang dihasilkan oleh bakteri. Fungi dan bakteri harus menghasilkan setidaknya tiga jenis enzim untuk dapat menghidrolisis manan yaitu β-mananase, β-manosidase, dan α-galaktosidase
9
(Hilge et al. 1989). Bakteri mengeluarkan β-1,4-mananase untuk memecah polimer manan menjadi MOS. MOS kemudian dipecah oleh β-1,4-manosidase dan α-1,6-galaktosidase menghasilkan monosakarida (manosa dan galaktosa) sebagai sumber karbon (Sumardi 2005).
Prinsip pengujian aktivitas enzim mananase merupakan reaksi antara enzim dan substrat untuk menghasilkan produk berupa manosa atau MOS. Produk ini akan bereaksi dengan reagen dinitrosalisilic acid (DNS). Reagen dinitrosalisilat yang terdiri atas asam 3,5-dinitrosalisilat yang berwarna kuning akan tereduksi menjadi asam 2-amino-5-salisilat. Gugus nitro yang terjadi pada reaksi reduksi disebabkan karena adanya gula pereduksi hasil hidrolisis substrat oleh mananase (Miller 1959). Reagen DNS berfungsi untuk menghentikan reaksi dan memberikan warna pada larutan sehingga absorbansinya dapat diukur menggunakan spektrofotometer. Warna yang terbentuk bergantung pada banyak sedikitnya gula pereduksi yang dihasilkan. Pemanasan pada saat uji aktivitas enzim bertujuan memaksimalkan penghentian reaksi antara enzim dan substrat. Penggunaan kontrol pada uji aktivitas enzim bertujuan mengetahui bahwa hasil yang diperoleh merupakan hasil reaksi antara enzim dan substrat saat inkubasi.
Produksi dan Pemekatan Prebiotik
Kecepatan reaksi enzim akan mencapai maksimum jika semua enzim terdapat sebagai kompleks enzim substrat dan konsentrasi enzim bebas lebih kecil. Keadaan ini akan tercapai pada konsentrasi substrat yang tinggi (Lehninger 1982). Hasil penelitian Purnawan (2013) hasil hidrolisis enzim mananase
Streptomyces cyaenus dengan substrat BIS 0.5% menghasilkan gula pereduksi
sebanyak 0.11-0.22 mg/mL, total gula sebanyak 0.29-1.70 mg/mL, dan derajat polimerasi sebanyak 2.48-12.35. Heterogenitas manan kemungkinan menyebabkan enzim pengurai manan memiliki spesifitas yang berbeda dan memiliki produk hidrolisis yang berbeda pula.
Perbandingan nilai total gula dengan gula pereduksinya menunjukkan panjang oligosakarida (Yang et al. 2009). Derajat polimerasi (DP) digunakan untuk mengetahui komposisi oligosakarida yang terkandung dalam BIS. DP merupakan perbandingan antara nilai total gula dan gula perduksi. Total gula adalah keseluruhan kandungan total gula baik monosakarida maupun oligosakarida. Gula pereduksi adalah golongan gula yang dapat mereduksi senyawa penerima elektron seperti MOS. Gula pereduksi akan meningkat ketika aktivitas enzim meningkat (Lehninger 1982).
Perlakuan pemanasan dapat mempengaruhi kandungan gula secara kuantitatif karena panas dapat mendegradasi gula (Hana 2007). MOS memiliki nilai DP berkisar 2-15, yang dihasilkan oleh Candida albicans (Faille et al. 1991).
10
Uji Pengaruh Prebiotik terhadap Pertumbuhan Bakteri Asam Laktat dan Bakteri Patogen
Penggunaan tiga jenis media (normal, minimal, prebiotik) untuk uji prebiotik bertujuan untuk melihat pengaruh prebiotik secara nyata terhadap pertumbuhan bakteri asam laktat maupun bakteri patogen. Media normal dan minimal dimaksudkan sebagai kontrol pembanding dan starter untuk adaptasi bakteri terhadap penggunaan prebiotik.
Berdasarkan hasil Total Plate Count (TPC), pertumbuhan Pediococcus
pentosaceus pada media MRS yang disubstitusi prebiotik mengalami kenaikan
setelah masa inkubasi 48 jam kedua. Hal ini menunjukkan Pediococcus
pentosaceus mampu memanfaatkan prebiotik sebagai sumber karbon untuk
pertumbuhannya. Salmonella sp. masih bisa tumbuh pada media LB yang disubstitusi prebiotik dimungkinkan karena Salmonella sp. memanfaatkan terlebih dahulu sumber karbon seperti tripton untuk pertumbuhannya. Setelah 24 jam kedua, pertumbuhan Salmonella sp. mengalami penurunan (Tabel 3), hal tersebut menunjukkan Salmonella sp. tidak bisa memanfaatkan prebiotik sebagai sumber karbon untuk pertumbuhannya. Oleh karena itu, secara in vitro penambahan prebiotik secara terus menerus bisa meningkatkan pertumbuhan Pediococcus
pentosaceus dan menekan pertumbuhan Salmonella sp..
Menurut Baurhoo et al. (2007) mekanisme MOS sebagai prebiotik secara selektif tidak menyebabkan peningkatan pertumbuhan bakteri yang menguntungkan, tetapi melalui kemampuannya yang dapat melekat pada lektin spesifik manosa dari bakteri patogen Gram negatif tipe I fimbriae seperti
Salmonella sp. dan E.coli yang kemudian akan dikeluarkan dari saluran
pencernaan. Aksi anti infeksi dari MOS pada fimbriae Salmonella sp. menyebabkan tidak terjadinya kolonisasi Salmonella sp.. Manan membantu perlawanan terhadap kolonisasi bakteri patogen dengan berperan sebagai reseptor analog untuk fimbrae Tipe I dalam E.coli dan Salmonella sp. (Oyofo et al. 1989).
Berdasarkan penelitian Spring et al. (2000) MOS menurunkan konsentrasi
S.typhimurium secara in vitro, namun tidak mempengaruhi konsentrasi Lactobacilli, Enterococci, bakteri anaerob dalam usus, asam laktat, asam lemak
volatil, dan pH usus. Penggunaan manosa efektif untuk mencegah kolonisasi
Salmonella pada ayam pedaging (Oyofo et al. 1989). MOS yang ditambahkan
pada pakan dapat mempengaruhi mikroflora usus ayam (Fernandez et al. 2000).
SIMPULAN DAN SARAN
Simpulan
Isolat W6 memiliki indeks mananolitik sebesar 6.5 dan memiliki aktivitas enzim optimum pada pH 5.5 suhu 30°C dengan waktu inkubasi 96 jam sebesar 0.0265 U/mL. Isolat ini mampu menghasilkan manooligosakarida (MOS) dengan
11
konsentrasi BIS 10%. Hal ini berarti bahwa isolat ini dapat dijadikan kandidat penghasil prebiotik. Prebiotik MOS dapat meningkatkan pertumbuhan
Pediococcus pentosaceus serta mampu menekan pertumbuhan Salmonella sp..
Isolat W6 memiliki kemampuan untuk memproduksi MOS dengan derajat polimerasi sebesar 3.3 – 8.6 yang potensial sebagai prebiotik
Saran
Produk hidrolisis perlu dianalisis kandungan gulanya misalnya dengan metode Kromatografi Lapis Tipis (KLT) sehingga bisa diketahui pengaruh komponen gula terhadap potensi dari prebiotik MOS, selain itu perlu dilanjutkan kembali mengenai pengaruh konsentrasi BIS diatas 10% untuk melihat produk hidrolisis yang dihasilkan.
DAFTAR PUSTAKA
Baurhoo B, Letellier A, Zhao X, Ruiz-Feria CA. 2007. Cecal populations of
Lactobacilli and Bifidobacteria and Escherichia coli populations after in
vivo Escherichia coli challenge in birds fed diets with purified lignin or mannanooligosaccharides. Poultry Scien. 86: 2509-2516.
Daud M. 2005. Performan ayam pedaging yang diberi probiotik dan prebiotik dalam ransum. J Ilmu Tern. 5 (2): 75-79.
Downie B, Hilhorst HWM, Bewley JD. 1994. A new assay for quantifying endo-ß-D-mannanase activity using congo red dye. Phytochemistry 36 (4): 829-835
Dubois M, Giles KA, Hamilton JK. 1956. Colorimetric method for determination of sugars and related substances. Anal Chem. 28: 350-356.
Faille C, Wieruszeski JM, Lepage G, Michalski JC, Poulain D, Strecker G. 1991. 1H-NMR spectroscopy of manno-oligosaccharides of the β-1,2-linked series released from the phosphopeptidomannan of Candida albicans source VW-32 (serotype A). Biochem and Biophys Research Commun. 181(3): 1251-1258.
Fernandez F, Hinton M, Gils VB. 2002. Dietray mannan-oligosaccharides and their effect on chicken caecal microflora in relation to Salmonella enteritidis colonization. Avian Pathology. 31: 49-58.
Gibson GR, Roberfroid MB. 1995. Dietary Modulation of the Human Colonie Microbiota: Introducing the Concept of Prebiotics. J Nutr. 125 (6): 1401-1412.
Hana. 2007. Pengaruh pemanasan terhadap kemampuan ekstrak gula talas (Colocasia esculenta (L) Schott) untuk mendukung pertumbuhan bakteri
12
asam laktat dan evaluasi in vivo potensi prebiotik [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Hilge M, Gloor SM, Rypniewski W, Sauer O, Heightman TD, Zimmermann W, Winterhalter K, Piontek K. 1989. High-resolution native and complex structures of thermostable mannanase from Thermomonospora fusca substrate specificity in glycosil hydrolase family 5. Structure. 6 (11): 1433-1444.
Iskandar S, Sinurat AP, Tiesnamurti B, Bamualim A. 2008. Bungkil inti sawit potensial untuk pakan ternak. Warta Penelitian dan Pengembangan
Pertanian. 30 (1): 16-17.
Lehninger AL.1982. Dasar-Dasar Biokimia. Thenawijaya M, penerjemah. Jakarta (ID): Erlangga. Terjemahan dari: Principle of Biochem. Ed ke-1.
Meryandini A, Ambarawati D, Rachmania N. 2008a. Pencirian mananase
Streptomyces costricanus 451-3. JIPI. 13 (1): 1-6.
Meryandini A, Anggreandari R, Rachmania N. 2008b. Isolasi bakteri mananolitik dan karakterisasi mananasenya. Biota. 13(2): 82-88.
Miller GL. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem. 31: 426-428.
Oyofo BA, Deloach JR, Corrier DE, Norman JO, Ziprin RL, Mollenhauer HH. 1989. Effect of carbohydrates on Salmonella typhimurium colonization in broiler chickens. Avian Diseases: 33: 531-534.
Petkowicz CLO, Reicher F, Chanzy H, Taravel FR, Vuong R. 2001. Linear mannan in the endosperm of Schizolobium amazonicum. Carbohydrate
Polymer. 44:107- 112.
Purnawan A. 2013. Hidrolisis karbohidrat manan (bungkil inti kelapa sawit) dengan enzim mananase dari Streptomyces cyaenus untuk menghasilkan oligosakarida [tesis]. Bogor (ID): Sekolah Pascasarjana, Institut Pertanian Bogor.
Rahayu WP. 2013. Isolasi dan pencirian bakteri mananolitik pendegradasi bungkil inti sawit [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Spring P, Wenk C, Dawson KA, Newman KE. The effect of dietary mannanooligosacharides on cecal parameters and the concentrations of enteric bacteria in the ceca of Salmonella-challenged broiler chicks. Poultry
Scien. 79: 205-211.
Suhartono MT. 1989. Enzim dan Bioteknologi. Bogor (ID): Departemen Pendidikan dan Kebudayaan Direktorat Jendral Pendidikan Tinggi Antar Universitas Bioteknologi Institut Pertanian Bogor.
Sumardi. 2005. Isolasi, karakterisasi, dan produksi β-mananase ekstraseluler dari
Geobacillus stearothermophilus L-07 [tesis]. Bogor (ID): Institut Pertanian
Bogor.
Tafsin M, Sofyan LA, Ramli N, Wiryawan KG, Zarkasie K, Piliang WG. 2007. Polisakarida mengandung mannan dari bungkil inti sawit sebagai antimikroba Salmonella typhimurium pada ayam. Media Petern. 30(2):139-146.
Utami W. 2010. Pembuatan prebiotik dari bungkil inti sawit menggunakan bakteri mananolitik dan pemanfaatannya untuk menghambat pertumbuhan E. coli dan memicu pertumbuhan L. casei [tesis]. Bogor (ID): Sekolah Pascasarjana, Institut Pertanian Bogor.
13
Williams, Wilkins. 1996. Biokimia Kedokteran Dasar: Sebuah Pendekatan Klinis. Pendit BU, penerjemah; Suyono J, Sadikin V, Mandera LI, editor. Jakarta (ID): EGC. Terjemahan dari: Basic Medical Biochemistry: A Clinical Approach. Ed ke-1.
Yang P, Li Y, Wang Y, Meng K, Luo H, Yuan T, Bai Y, Zhan Z, Yao B. 2009. A novel beta-mannanase with high specific activity from Bacillus circulans CGMCC1554: gene cloning, expression and enzymatic characterization.
Appl Biochem Biotechnol. 159(1):85-94.
Yopi, Purnawan A, Thontowi A, Hermansyah H, Wijanarko A. 2006. Preparasi manan dan mananase kasar dari bungkil inti sawit. J Teknol. (4):312-319.
14
Lampiran 1 Komposisi reagen dinitrosalicylic acid (DNS) Komposisi reagen dinitrosalicylic acid (DNS):
NaOH padat = 10 g KNa tartat = 182 g
Na2SO3 = 0.5 g
DNS = 10 g
15
Lampiran 2 Cara perhitungan aktivitas enzim Cara perhitungan aktivitas enzim yaitu:
A= OD sampel – OD blanko B= OD kontrol – OD blanko
Kurva standar gula pereduksi manosa dibuat dengan berbagai konsentrasi manosa, sehingga diperoleh persamaan y= ax + b
Gula pereduksi sampel (mg/mL) = (A + b) / a = X sampel Gula pereduksi kontrol (mg/mL) = (B + b) / a = X kontrol Aktivitas enzim (U/mL) = (X sampel – X kontrol) x fp x 1000
BM manosa x waktu inkubasi (menit) fp = faktor pengenceran y = 2.8061x - 0.0658 R² = 0.9849 -0.2 -0.1 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 Ab so rb an si Konsentrasi manosa (mg/mL)
Kurva Standar Manosa
16
Lampiran 3 Cara perhitungan total gula dan derajat polimerasi Cara perhitungan total gula:
A = OD sampel – OD blanko
Kurva standar total gula manosa dibuat dengan berbagai konsentrasi, sehingga diperoleh persamaan y = ax + b
Total gula sampel (mg/mL) = (A + b) / a
Cara perhitungan derajat polimerasi (DP) = Total gula / Gula perduksi
y = 9.8845x + 0.0005 R² = 0.9989 0 0.2 0.4 0.6 0.8 1 1.2 0 0.01 0.02 0.03 0.04 0.05 0.06 0.07 0.08 0.09 0.1 A b sor b an si Konsentrasi
17
Lampiran 4 Komposisi media MRS dan LB a Komposisi media MRS 100 mL
Bahan MRS normal MRS minimal MRS prebiotik
Pepton 1.8 g 0.18 g 0.18 g Ekstrak khamir 0.4 g 0.04 g 0.04 g K2HPO4 0.2 g 0.2 g 0.2 g Tween 80 0.1 mL 0.1 mL 0.1 mL MgSO4 . 7H2O 0.02 g 0.02 g 0.02 g MnSO4 0.004 g 0.004 g 0.004 g Glukosa/prebiotik 1 g 0.1 g 1 g Diamonium hidrogen sitrat 0.2 g 0.2 g 0.2 g b Komposisi media LB 100 mL
Bahan LB normal LB minimal LB prebiotik
Tripton 1 g 0.1 g 0.1 g
Ekstrak
khamir/prebiotik 0.5 g 0.05 g 0.5 g
18
RIWAYAT HIDUP
Penulis dilahirkan di Tasikmalaya pada tanggal 24 April 1992 dari Ayah Yayat Ruhiyat dan Ibu Nani Tartiyah. Penulis merupakan putri kedua dari tiga bersaudara. Tahun 2010 penulis lulus dari MA Al Amin Tasikmalaya dan pada tahun yang sama diterima di Program Studi Biologi, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor melalui jalur ujian seleksi masuk IPB (USMI).
Penulis pernah melaksanakan studi lapang mengenai Isolasi dan Karakterisasi Bakteri Penghasil Selulase dari Sampel Tanah Asal Taman Nasional Gunung Gede Pangrango (TNGGP) pada tahun 2012 serta praktik lapangan dalam bidang Pemeriksaan Darah dan Urin di Laboratorium Klinik Rumah Sakit Umum Daerah (RSUD) Kota Tasikmalaya pada bulan Juli sampai Agustus 2013.
Selama masa perkuliahan penulis aktif dalam organisasi kemahasiswaan yaitu sebagai anggota divisi Bioworld, Himpunan Mahasiswa Biologi (Himabio) IPB (2012/2013). Selain itu penulis aktif dalam berbagai kepanitiaan event kampus diantaranya yaitu, staf divisi konsumsi Lomba Cepat Tepat Biologi (LCTB) IPB (2011); ketua divisi Leader Operasional (LO) LCTB dan anggota divisi LO Pesta Sains Nasional FMIPA IPB (2012); divisi konsumsi Biologi on Experiment (BOX) (2012); bendahara Himabio Go Field (2012); staf divisi konsumsi BOX (2013). Penulis pernah menjadi asisten praktikum Biologi Dasar periode semester ganjil 2012/2013 serta periode semester ganjil dan genap 2013/2014.