i
KINETIKA BIOTRANSFORMASI SUKSINONITRIL
MENJADI ASAM SUKSINAT OLEH
Pseudomonas
sp.
TUTI UTAMA SIREGAR
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ii
ABSTRAK
TUTI UTAMA SIREGAR. Kinetika Biotransformasi Suksinonitril menjadi Asam
Suksinat oleh Pseudomonmas sp.. Dibimbing oleh GUSTINI SYAHBIRIN dan
BAMBANG SUNARKO.
Asam suksinat (HOOC(CH2)2COOH) merupakan senyawa komersial yang banyak
digunakan dalam bidang pangan, misalnya pada industri kecap, sari buah, anggur, dan produk-produk susu lainnya. Selama ini asam suksinat yang kita gunakan dihasilkan melalui serangkaian reaksi kimia yang sangat berbahaya sehingga diperlukan alternatif baru yang lebih aman dan murah melalui proses biotransformasi dengan memanfaatkan
aktivitas Pseudomonas sp. Untuk mengetahui kemampuan Pseudomonas sp. dalam proses
biotransformasi digunakan sistem fermentasi batch untuk menentukan pola pertumbuhan,
perubahan pH, produksi amonium, laju penguraian substrat, dan pembentukan produk. Analisis terhadap penguraian suksinonitril dan asam suksinat yang terbentuk menggunakan kromatografi cair kinerja tinggi yang menggunakan metanol 80% sebagai
fase gerak, fase diam kolom organik C18 10 , dan detektor ultraviolet yang diukur pada
panjang gelombang 220 nm, sedangkan amonium yang terbentuk diukur dengan menggunakan spektrofotometer pada panjang gelombang 400 nm. Hasil penelitian menunjukkan produksi optimum enzim terjadi pada fasa log. Laju pembentukan asam suksinat sebesar 0.982 mM/mL.jam seiring dengan penurunan konsumsi substrat sebesar 1.235 mM/mL.jam. Perolehan asam suksinat setelah 81 jam fermentasi sebesar 81 %. pH
optimum, temperatur optimum, Km, dan Vmaks adalah 7, 27 °C, 90 mM, dan 0.0002
µM/mL.jam.
ABSTRACT
TUTI UTAMA SIREGAR. Biotransformation Kinetic of Succinonitrile to Succinic
Acid by Pseudomonas sp.. Supervised by GUSTINI SYAHBIRIN and BAMBANG
SUNARKO.
Succinic acid (HOOC(CH2)2COOH) is a commercial compound used in many food
industry, such as soy sauce, juice, wine, and dairy products. So far, succinic acid is produced through a dangerous chemical reaction series, so a safer and cheaper alternative
is needed by means of biotransformation process with application of Pseudomonas sp.
activity. To investigate the ability of Pseudomonas sp. in biotransformation process
batch fermentation system was used in this research to determine the pattern of growth, pH alteration, ammonium production, substrate degradation rate, and product formation. Analysis towards the succinonitrile and succinic acid degradation was carried out using high performance liquid chromatography with 80% methanol as mobile phase, organic
coloumn C18 10µ as stationary phase, and ultraviolet detector at 220 nm whereas the
ammonium formation was measured using spectrophotometer at 400 nm. The result of this research showed that the production of optimum enzyme occurred on log phase. Formation rate of succinic acid was 0.982 mM/mL.hr along with the decrease of substrate consumption of 1.235 mM/mL.hr. The yield of succinic acid after 81 hours of
fermentation was 81%. Optimum pH, optimum temperature, Km, and Vmaks were 7, 27 ºC,
iii
KINETIKA BIOTRANSFORMASI SUKSINONITRIL
MENJADI ASAM SUKSINAT OLEH
Pseudomonas
sp.
TUTI UTAMA SIREGAR
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Kimia
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
iv
Judul : Kinetika Biotransformasi Suksinonitril menjadi Asam Suksinat
oleh Pseudomonas sp.
Nama : Tuti Utama Siregar
NIM : G44201001
Disetujui
Pembimbing I Pembimbing II
Dra. Gustini Syahbirin, M.S Dr. Bambang Sunarko NIP 131 842 414 NIP 320 004 933
Diketahui
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor
Prof. Dr.Ir. Yonny Koesmaryono, M.S NIP 131 473 999
v
PRAKATA
Bismillaahirrahmaanirrahiim.
Alhamdulillah, segala puji dan syukur penulis panjatkan ke hadirat Allah SWT yang telah melimpahkan rahmat dan hidayah-Nya serta shalawat beriring salam kepada Nabi Muhammad SAW sehingga penulis dapat menyelesaikan karya ilmiah ini. Penelitian ini
berjudul Kinetika Biotransformasi Suksinonitril oleh Pseudomonas sp. yang dilaksanakan
sejak bulan Oktober 2005 sampai April 2006 di Laboratorium Bioproses, Pusat Penelitian Bioteknologi, LIPI, Cibinong dan Laboratorium Ekologi dan Fisiologi Mikrob, Bidang Mikrobiologi, Pusat Penelitian Biologi, LIPI, Bogor.
Terima kasih penulis ucapkan kepada berbagai pihak yang telah membantu terutama kepada Bapak Dr. Bambang Sunarko selaku pembimbing dari Puslit Bioteknologi-LIPI, Cibinong dan Ibu Dra. Gustini Syahbirin, M.S selaku pembimbing dari Departemen Kimia-FMIPA, IPB. Penulis juga mengucapkan terima kasih kepada seluruh staf, peneliti, serta teman-teman di laboratorium atas segala bantuan dan diskusinya.
Ucapan terima kasih tak terhingga penulis sampaikan kepada Ayahanda dan Ibunda serta kakak dan adikku atas kasih sayang, dukungan, cinta, dan doanya. Untuk Raja terima kasih atas kebersamaan, cinta, dan doanya serta tak lupa untuk teman-teman Kimia 38 dan anak-anak Ciwaluya 22.
Semoga apa yang penulis lakukan menjadi suatu amal ibadah dan diberikan keridoan-Nya. Mudah-mudahan karya kecil ini bermanfaat bagi kemaslahatan manusia dan perkembangan ilmu pengetahuan.
Bogor, September 2006
vi
RIWAYAT HIDUP
Penulis dilahirkan di Medan, Sumatera Utara pada tanggal 22 Februari 1982 sebagai
putri ketiga dari empat bersaudara, dari ayah Tajuddin Siregar dan ibu Nuraini Tanjung. Tahun 2001 penulis lulus dari SMUN 3 Sipirok dan pada tahun yang sama diterima diDepartemen Kimia, Institut Pertanian Bogor, melalui jalur Undangan Seleksi Masuk IPB (USMI). Tahun 2004, penulis melaksanakan praktik kerja lapangan di Laboratorim Ekologi dan Fisiologi Mikrob, Bidang Mikrobiologi, LIPI, Bogor dan pada tahun 2005, penulis melaksanakan penelitian tugas akhir di Laboratorim Bioproses, Pusat Penelitian Bioteknologi, LIPI, Cibinong dan Laboratorium Ekologi dan Fisiologi Mikrob, Bidang Mikrobiologi, Pusat Penelitian Biologi, LIPI, Bogor.
Selama mengikuti perkuliahan penulis aktif mengikuti berbagai organisasi dan
mengajar di lembaga Bimbingan Belajar Nurul Ilmi dan Private Community padaTahun
vii
DAFTAR ISI
Halaman
DAFTAR GAMBAR... viii
DAFTAR LAMPIRAN... viii
PENDAHULUAN ... 1
TINJAUAN PUSTAKA Suksinonitril dan Asam Suksinat... 1
Biokonversi Suksinonitril... 1
Enzim Pendegradasi Nitril ... 2
Proses Fermentasi dan Kinetika Fermentasi ... 3
NHase dan Amidase... 4
Karakterisasi Enzim ... 4
BAHAN DAN METODE Bahan dan Alat ... 5
Metode ... 5
HASIL DAN PEMBAHASAN Fermentasi Pseudomonas sp. ... 6
Aktivitas Enzim Pengkonversi Suksinonitril Selama Fermentasi ... 8
Pengaruh pH terhadap Aktivitas Enzim NHase ... 9
Pengaruh suhu terhadap Aktivitas Enzim NHase ... 9
Kinetika Biotransformasi Suksinonitril... 9
Biotransformasi Suksinonitril ... 10
SIMPULAN DAN SARAN... 11
DAFTAR PUSTAKA ... 11
viii
DAFTAR GAMBAR
Halaman
1 Biokonversi suksinonitril ... 2
2 Biokonversi senyawa nitril oleh NHase,amidase dan nitrilase... 2
3 Proses fermentasi ... 3
4 Struktur NHase dan Amidase... 4
5 Pola dan pH pertumbuhan Pseudomonas sp. dalam suksinonitril ... 7
6 Penurunan konsentrasi suksinonitril dan pembentukan asam suksinat ... 7
7 Kromatogram standar suksinonitril... 8
8 Kromatogram standar asam suksinat... 8
9 Kromatogram sampel pada jam ke-0 ... 8
10 Kromatogram sampel pada jam ke-81.. ... 8
11 Aktivitas total dan spesifik NHase selama fermentasi ... 9
12 Pengaruh pH terhadap aktivitas NHase... 9
13 Pengaruh suhu terhadap aktivitas NHase... 10
14 Pengaruh konsentrasi substrat terhadap kecepatan awal reaksi NHase ... 10
15 Konsentrasi NH4 + yang dihasilkan pada kec.pembentukan NH4 + ... 10
16 Konsentrasi NH4+ yang dihasilkan pada kec. relatif pembentukan NH4+... 11
17 Konsentrasi NH4+ yang dihasilkan pada proses biokonversi suksinonitril ... 11
DAFTAR LAMPIRAN
Halaman 1 Bagan alir penelitian ... 14
2 Analisis NH4+dengan metode Nessler ... 15
3 Pengukuran kadar protein dengan metode Biuret ... 16
4 Kurva standar protein dan sampel protein selama fermentasi... 17
5 Kromatogram standar dan sampel berdasarkan analisis KCKT ... 18
6 Konsentrasi suksinonitril dan asam suksinat selama fermentasi ... 21
7 Pola pertumbuhan Pseudomonas sp.pada suksinonitril ... 21
8 Aktivitas enzim selama proses fermentasi... 22
9 Pengaruh substrat terhadap kec. reaksi enzimatik pada penentuan Km dan Vmaks 23
1
PENDAHULUAN
Suksinonitril (CNCH2CH2CN) disebut
juga butanadinitril dan termasuk dalam kelompok senyawa dinitril alifatik. Suksinonitril pada konsentrasi tinggi dapat menyebabkan iritasi pada kulit dan gangguan pada saluran pernafasan (Benesovsky 1981).
Suksinonitril ádalah senyawa yang dapat digunakan sebagai substrat dan diubah menjadi asam suksinat melalui proses biokonversi dengan bantuan mikrob yang mampu memproduksi enzim pendegradasi suksinonitril (Komeda et al. 1996). Asam suksinat (HOOC)2(CH2)2 merupakan senyawa
komersial yang banyak digunakan dalam industri. Asam suksinat banyak digunakan dalam bidang pangan, misalnya pada industri kecap, sari buah, anggur, dan produk-produk susu lainnya. Selain itu, asam suksinat juga digunakan pada industri tekstil pada proses pewarnaan, industri obat, pernis, cat, dan lempeng fotografi.
Penggunaan asam suksinat yang cukup luas menyebabkan kebutuhannya semakin meningkat. Umumnya, asam suksinat yang dihasilkan selama ini dibuat secara industri melalui serangkaian reaksi kimia yang menghasilkan produk samping yang berbahaya yaitu, asam sianida.
Perkembangan bioteknologi atau bioproses memberikan alternatif yang lebih aman dan murah dalam sintesis asam suksinat, yaitu melalui proses biotransformasi dengan memanfaatkan substrat yang berlimpah dan murah, terutama substrat yang berasal dari limbah industri.
Pembuatan asam suksinat secara mikrobial menggunakan mikrob yang dapat melakukan proses degradasi atau dekomposisi substrat yang melibatkan reaksi enzimatik. Proses ini memiliki banyak keuntungan dibandingkan dengan reaksi kimia biasa, sebab dapat menghasilkan produk dengan tingkat kemurnian tinggi, hemat energi, biaya produksi rendah, dan aman terhadap lingkungan.
Sel bakteri Pseudomonas sp. yang diisolasi dari limbah PT Pusri Palembang, merupakan koleksi bakteri Eko-Fisiologi Mikrob, LIPI. Pseudomonas sp. tersebar luas di alam dan galurnya memiliki kemampuan beradaptasi secara genetis terhadap lingkungannya dan mampu mendegradasi sejumlah besar senyawa. (Holloway et al.
1998). Bakteri ini dapat hidup dengan baik dan memiliki rapat optis yang tinggi ketika
ditumbuhkan dalam medium yang mengandung suksinonitril. Bakteri ini berpotensi menghasilkan asam suksinat melalui proses biokonversi. Oleh karena itu, perlu dipelajari kemampuan Pseudomonas sp. dalam memproduksi asam suksinat dan enzim yang terlibat di dalam biokonversi suksinonitril oleh sel tersebut.
Penelitian ini bertujuan untuk mempelajari kinetika biotransformasi suksinonitril menjadi asam suksinat oleh
Pseudomonas sp. serta mempelajari karakteristik enzim yang terlibat di dalamnya.
Hipotesis yang mendukung penelitian ini adalah, enzim NHase yang diproduksi oleh
Pseudomonas sp. mampu mengkonversi suksinonitril menjasi asam suksinat. Hasil penelitian ini diharapkan dapat bermanfaat bagi pengembangan industri yang menggunakan asam suksinat dengan harga yang lebih murah dan mengurangi kerusakan lingkungan akibat akumulasi senyawa nitril.
TINJAUAN PUSTAKA
Suksinonitril dan Asam Suksinat
Suksinonitril (CNCH2CH2CN) disebut
juga butanadinitril, adalah substansi stabil barupa padatan yang larut dalam air dengan titik didih 265 ºC, titik leleh 54 ºC, dan titik nyala 132 ºC. Senyawa ini memiliki kelarutan sebesar 1.02 g/cm3 dalam air dan merupakan senyawa toksik dengan toksisitas akut oral LD50 pada tikus sebesar 450 mg/kg (Pollak
1991). Suksinonitril pada konsentrasi tinggi dapat menyebabkan iritasi pada kulit dan gangguan pada saluran pernapasan (Benesovsky 1981).
Asam suksinat (COOH)2 (CH2)2, disebut
juga asam butanadioat adalah substansi stabil berupa padatan yang larut dalam air dengan bobot molekul 118 g/mol dengan titik didih 235 ºC, titik leleh 185 ºC, dan titik nyala 206 ºC. Senyawa ini memiliki kelarutan 1.56 g/cm3 dalam air. Asam suksinat pada konsentrasi tinggi dapat menyebabkan luka bakar, iritasi pada kulit, dan kerusakan pada mata. Asam suksinat banyak digunakan sebagai perasa pada makanan dan minuman, obat penghilang rasa sakit, sebagai antioksidan, dan antiflek pada kosmetik.
Biokonversi Suksinonitril
2
mikroorganisme yang melibatkan reaksi enzimatik. Suatu mikrob dapat melakukan reaksi biokonversi karena mikrob memiliki enzim yang dapat mengkonversi substratnya menjadi produk. Umumnya proses ini tanpa pembentukan hasil samping yang berbahaya.
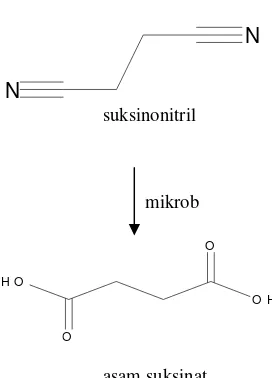
Suksinonitril yang merupakan produk limbah dapat diubah menjadi asam suksinat oleh mikrob yang memiliki aktivitas enzim pendegradasi nitril dengan mengubah dua gugus sianida menjadi gugus karboksilat yang relatif tidak toksik. Berikut adalah biokonversi suksinonitril dengan bantuan mikrob.
N
N
suksinonitril
mikrob
O
O H
O H O
asam suksinat
Gambar 1 Biokonversi Suksinonitril.
Menurut Crueger & Crueger (1984), reaksi biokonversi memiliki beberapa kelebihan dibandingkan dengan reaksi kimia biasa antara lain, spesifik terhadap substrat, spesifik terhadap situs tertentu (regioselektif), memiliki kespesifikan stereo dan enansio (stereoselektif dan enensioselektif), dan reaksi biokonversi tidak membutuhkan kondisi reaksi yang ekstrim.
Aplikasi reaksi biokonversi antara lain untuk sintesis berbagai senyawa organik seperti akrilamida dan nikotin amida (Kobayashi 1993), sintesis asam askorbat oleh
Acetobacter suboxydans, dan sintesis berbagai jenis antibiotik oleh mikrob penghasilnya, misalnya sintesis penisilin dari Penicillium crysogenum. Dalam bidang pertanian, reaksi biokonversi digunakan untuk mendegradasi pestisida dengan cara menggunakan pestisida sebagai substrat untuk pertumbuhan mikrob dan produksi energi (Crueger & Crueger 1984).
Penggunaan mikrob dalam skala industri untuk memproduksi akrilamida, nikotin amida, dan senyawa amida lainnya melalui proses biokonversi menggunakan enzim NHase yang telah terimobilisasi dalam bentuk sel utuh dari Pseudomonas chloraphis B32,
Rhodococcus sp. R312, dan Rhodcoccus rhodochorous J1 (Liese 2000).
Enzim Pendegradasi Nitril
Senyawa nitril digunakan secara luas pada industri manufaktur dan industri kimia. Senyawa tersebut sangat toksik dan pada umumnya merupakan senyawa yang sulit terdegradasi, tetapi beberapa mikroorganisme mampu mendegradasi senyawa nitril dengan cara menggunakan nitril sebagai sumber karbon dan nitrogen.
Enzim yang terlibat dalam hidrolisis senyawa nitril adalah NHase, amidase, dan nitrilase. Secara umum terdapat dua tipe reaksi utama metabolisme pada proses hidrolisis senyawa nitril dengan menggunakan aktivitas mikrob, masing-masing untuk senyawa nitril alifatik dan senyawa nitril aromatik.
1RCN NHase RCONH2 amidase RCOOH
+ + + H2O H2O NH4
2 R’CN + H2O nitrilase RCOOH + NH4
Gambar 2 Biokonversi senyawa nitril oleh NHase ,amidase dan nitrilase.
Pada reaksi tahap pertama, NHase mengubah senyawa nitril menjadi senyawa amida dan dilanjutkan menjadi amidase untuk mengubah senyawa amida menjadi amonium (NH4+) dan asam karboksilat, sedangkan pada
reaksi kedua, nitrilase langsung mengubah senyawa nitril menjadi asam karboksilat dan amonium (NH4
+
) (Ciskanik et al. 1995). Kedua tahap reaksi dapat bersama-sama dalam suatu mikrob yang memiliki ketiga enzim tersebut. Umumnya, nitrilase aktif terhadap senyawa nitril aromatik (Tauber 2000). Beberapa mikrob yang dilaporkan mampu memproduksi enzim NHase dan amidase antara lain Corynebacterium D5,
3
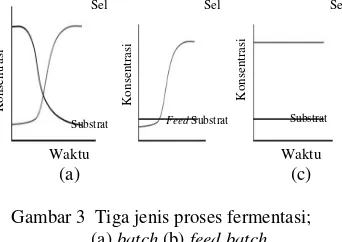
Proses Fermentasi dan Kinetika Fermentasi
Proses fermentasi adalah suatu proses mendayagunakan aktivitas metabolisme suatu mikrob tertentu atau campuran dari beberapa spesies mikrob untuk menghasilkan senyawa tertentu (Rahman 1992). Terdapat tiga macam proses fermentasi cair yaitu, fermentasi
batch, kontinyu, dan feed batch.
Gambar 3 Tiga jenis proses fermentasi; (a) batch (b) feed batch
(c) kontinyu.
Pada fermentasi batch, setelah inokulasi tidak dilakukan penambahan substrat ke dalam media. Pada fermentasi ini umumnya dijumpai empat fase pertumbuhan mikrob yaitu, fase lag, log, stasioner, dan fase kematian. Kondisi fisiologi dan metabolisme sel pada keempat fase tersebut berbeda.
Fase lag yang merupakan awal pertumbuhan mikrob, dimulai sejak inokulasi sampel ke media. Pada fase ini sel butuh penyesuaian terhadap kondisi lingkungan yang baru, sel mengalami perubahan komposisi kimiawi, bertambah ukuran, dan substansi seluler sehingga pertumbuhannya sangat sedikit.
Pada fase eksponensial terjadi pembelahan dan pertumbuhan sel yang cepat. Pada fase ini, populasi meningkat secara signifikan dan mengikuti persamaan eksponensial yang kurva pertumbuhannya berupa kurva linear. Sel yang berada pada fase ini sering digunakan untuk mempelajari enzim dan komponen lainnya. Fase ini sangat dipengaruhi oleh temperatur dan komposisi media kultur.
Fase stasioner merupakan fase yang tidak terjadi peningkatan atau penurunan jumlah sel. Pertambahan populasi dan kematian mikrob berlangsung dengan seimbang. Pada fase ini terjadi penumpukan produk beracun atau kehabisan nutrien, sehingga sel mati dan sebagian tumbuh membelah. Pada beberapa organisme pertumbuhan yang lambat mungkin terjadi pada masa ini. Setelah mencapai fase
stasioner, populasi mikrob akan mengalami fase kematian. Pada fase ini sebagian besar mikrob mati, karena substrat atau media untuk pertumbuhan sudah tidak memadai untuk pertumbuhan mikrob.
Pada fermentasi kontinyu, ada penambahan dan pengurangan kultur selama proses fermentasi, sehingga volume tetap dan kondisi fisiologi sel cenderung konstan, sedangkan pada fermentasi feedbatch, ada penambahan substrat pada fase pertumbuhan tertentu, yang bertujuan untuk memperpanjang fase pertumbuhan yang diinginkan.
Mikroorganisme tumbuh dalam suatu spektrum lingkungan fisik dan kimiawi yang sangat luas, pertumbuhan dan kegiatan-kegiatan fisiologik lainnya merupakan suatu respon terhadap lingkungan fisika-kimiawinya. Kinetika fermentasi menggambarkan pertumbuhan dan pembentukan produk oleh mikroorganisme, bukan hanya pertumbuhan sel aktif tetapi juga kegiatan sel istirahat dan sel mati (Said 1987).
Pertumbuhan sel didefenisikan sebagai kenaikan biomassa sel persatuan waktu (White 1995). Kecepatan pertumbuhan merupakan perubahan dalam jumlah sel persatuan waktu. Waktu penggandaan (td) atau
Generation Time adalah waktu yang dibutuhkan untuk menggandakan sel pada fase eksponensial.
Nt = No x 2n
td =
t
n
Keterangan
Nt = jumlah sel akhir
No = jumlah sel awal
n = jumlah generasi
µ = konstanta laju pertumbuhan
µ digunakan untuk mengukur jumlah/banyaknya generasi per unit waktu dalam pertumbuhan eksponensial.
Produktivitas (P) =
t
P
P
t−
oPerolehan/Yield (Y) =
o t o t
S
S
P
P
−
−
KeteranganPt = konsentrasi akhir produk (mM)
Po = konsentrasi awal produk (mM)
St = konsentrasi akhir substrat (mM)
So = konsentrasi awal substrat (mM)
t = waktu fermentasi (jam) Waktu (a) Waktu (c) K o n se n tr as i K o n se n tr a si
Sel Sel Sel
Feed Substrat Substrat
4
NHase dan Amidase
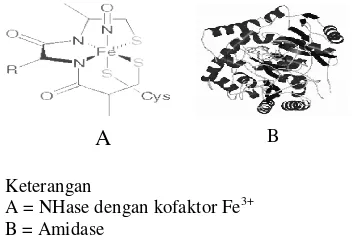
Tauber et al. (2000) menyatakan bahwa NHase dari sel Rhodococcus rhodochrous
merupakan enzim yang memiliki dua sub unit, yaitu sub unit α yang memiliki 206 residu asam amino dengan berat molekul (BM) sebesar 22,786, g/mol dan sub unit β yang memiliki 418 residu asam amino dengan BM sebesar 23,428 g/mol. Enzim ini memiliki kofaktor berupa ion kobal (Co) atau besi untuk aktivitas katalitiknya.
Amidase dari sel Pseudomonas chlororaphis B23 merupakan enzim dengan BM 105 g/mol yang memiliki dua sub unit berupa homodimer dengan BM 54 g/mol (Ciskanik 1995). Residu sisi aktif dari enzim amidase pada sel Rhodococcus rhodochrous
Jl adalah Asp 191 dan Ser 195.
Asp 191 akan mengambil proton dari residu Ser 195 sehingga Ser 195 bertindak sebagai nukleofil terhadap gugus karbonil dari ikatan amida pada substrat (Kobayashi 1997). Amidase tidak membutuhkan kofaktor untuk aktivitas katalitiknya. Umumnya, enzim pentransformasi nitril terakumulasi pada fasa log pertumbuhan sel.
Keterangan
A = NHase dengan kofaktor Fe3+ B = Amidase
Gambar 4 Struktur NHase dan Amidase.
Karakterisasi Enzim
Bakteri mengandung enzim konstitutif dan induktif. Enzim konstitutif merupakan enzim yang terdapat dalam sel bakteri dalam jumlah tetap dan tidak bergantung pada keadaan metabolisme organisme tersebut seperti enzim yang terlibat dalam lintasan glikolisis. Enzim induktif dalam sel bakteri dalam sel bakteri terdapat dalam berbagai konsentrasi. Dalam keadaan normal terdapat dalam jumlah kecil, tetapi konsentrasinya akan meningkat dengan cepat bila substratnya terdapat dalam medium terutama jika substrat tersebut merupakan sumber karbon satu-satunya bagi sel (Lehninger 1994).
Rhodococcus erythropolis A10 bila ditempatkan di media kultur yang berisi asetonitril sebagai satu-satunya sumber karbon, nitrogen, dan energi dalam waktu yang relatif cepat akan mensintesis nitril hidratase dan amidase dalam jumlah besar (Acharya & Desai 1997). Bila bakteri yang terinduksi asetonitril dipindahkan ke medium yang mengandung glukosa dan garam amonium, maka sintesis enzim NHase dan amidase akan terhenti. Dengan kata lain, induksi enzim merupakan proses yang ekonomis, sebab enzim induktif sanya dihasilkan bila diperlukan oleh sel. Senyawa yang dapat menginduksi sintesis enzim disebut penginduksi atau induktor (Lehninger 1994).
Beberapa enzim mempunyai spesifitas yang tinggi terhadap substrat tertentu dan tidak akan bekerja bahkan terhadap senyawa yang mirip secara struktural (Lehninger. 1994). Kobayashi et al. (1997) melaporkan bahwa enzim nitril hidratase dari
Rhodococcus rhodochrous K22 spesifik pada senyawa nitril alifatik, tetapi ada pula enzim yang mempunyai spesifitas yang rendah, artinya dapat bekerja pada berbagai senyawa dengan ciri struktur yang sama seperti yang dilaporkan oleh Acharya & Desai (1997), misalnya enzim nitril hidratase oleh
Rhodococcus erythropolis A10 mempunyai spesifitas terhadap substrat nitril alifatik dan aromatik. Aktivitas enzim terhadap substrat dipengaruhi oleh beberapa faktor yaitu, pH, suhu, konsentrasi substrat, dan aktivator (koenzim dan kofaktor) dan inhibitor.
pH
Efek pH pada enzim berkaitan dengan keadaan ionisasi dari sistem yang dikatalisis, termasuk substrat dan enzim itu sendiri. Perubahan pH dapat mempengaruhi keadaaan ionisasi dari asam-asam amino pada sisi aktif enzim sehingga akan mempengaruhi interaksinya dengan molekul substrat. Kadar pH yang terlalu tinggi atau terlalu rendah akan menyebabkan ketidakstabilan pada konformasi enzim sehingga menyebabkan struktur pada enzim rusak. Enzim mempunyai pH optimum yang khas yang akan menyebabkan aktivitas maksimal. Keadaan optimum ini dihubungkan dengan saat gugus pemberi proton atau penerima proton yang aktif pada sisi enzim berada pada kondisi ionisasi yang tepat. Keadaan optimum tidak harus sama dengan pH lingkungannya. Aktivitas enzim dalam sel sebagaian diatur
5
oleh pH media kulturnya (Lehninger 1994). Enzim NHase memiliki pH optimum pada pH 7, sedangkan amidase pada pH 6 (Purnomo 2000; Adityarini 1999).
Suhu
Suhu mempunyai dua pengaruh yang saling bertentangan. Suhu dapat meningkatkan aktivitas enzim, tetapi dapat pula merusak stuktur enzim. Suhu optimum merupakan batas keduanya (Dixon & Webb 1978). Peningkatan suhu sebelum tercapai suhu optimum akan meningkatkan kecepatan reaksi katalitik enzim karena energi kinetik molekul-molekul yang bereaksi, yaitu pada saat kompleks enzim-substrat melampaui energi aktivasi terlalu besar, sehingga memecah ikatan sekunder pada konformasi enzim dan sisi aktifnya. Hal ini mengakibatkan enzim terdenaturasi dan kehilangan sifat katalitiknya (Martin 1981). NHase mempunyai suhu optimum pada 25 °C, sedangkan amidase pada suhu 50 °C (Adityarini 1999).
Konsentrasi Substrat
Konsentrasi substrat sangat mempengaruhi kecepatan reaksi enzimatik, kecepatan maksimum (Vmaks) suatu enzim
dicapai ketika enzim telah jenuh oleh substratnya sehingga tidak dapat berfungsi lebih cepat. Konsentrasi substrat tertentu pada saat enzim mencapai setengah kecepatan maksimum disebut Km atau konstanta
Michaelis-Menten. Nilai Km bersifat khas
pada setiap enzim tertentu. Hubungan antara Km dan Vmaks dinyatakan dalam persamaan
Michaelis-Menten (Lehninger 1982).
S
+ =
m makss o
K [S] V
V
Persamaan Michaelis–Menten merupakan dasar bagi aspek kinetika kerja enzim. Apabila nilai Km dan Vmaks diketahui, maka
kecepatan reaksi suatu enzim pada setiap konsentrasi substrat dapat dihitung. Nilai Km
dan Vmaks yang lebih tepat dapat diperoleh
dengan memetakan data yang ada ke dalam persamaan Lineweaver-Burk sebagai berikut.
makss makss
m
V
1
[S]
1
V
K
Vo
1
+
=
Aktivator (kofaktor dan koenzim) dan Inhibitor
Lehninger (1994), menyatakan beberapa enzim membutuhkan komponen tambahan bagi aktivitasnya. Bila komponen tambahan tersebut berupa senyawa anorganik disebut kofaktor, sedangkan bila senyawa organik disebut koenzim. Pada beberapa enzim, kofaktor dan koenzim terlibat langsung pada proses katalitik, tapi ada juga yang berfungsi sebagai pembawa gugus fungsional tertentu. Hampir semua enzim dapat dihambat oleh senyawa kimia tertentu misalnya ion logam, senyawa pengkelat, senyawa organik, bahkan substrat enzim itu sendiri.
METODE PENELITIAN
Alat dan Bahan
Alat-alat yang digunakan adalah spektrofotometer UV/VIS, neraca analitik, pH meter, sentrifuse, kromatografi cair kinerja tinggi (KCKT), dan alat-alat gelas.
Bahan-bahan yang digunakan adalah suksinonitril, asam suksinat, larutan standar amonium, bufer fosfat, pereaksi Nessler, NaOH, bovine serum albumin (BSA), dan bakteri Pseudomonas sp. (koleksi kelompok Eko-Fisiologi Mikrob, Bidang Mikrobiologi, LIPI) yang diisolasi dari limbah PT Pusri Palembang.
Media Tumbuh Pseudomonas sp.
Media yang digunakan untuk menumbuhkan Pseudomonas sp. adalah media mineral dengan komposisi sebagai berikut: Na2HPO4.2H2O 0.4475 g; KH2PO4 0.1 g;
MgSO4.7H2O 0.1 g; CaCl2.2H2O 0.01 g;
FeSO4.7H2O 0.001 g; yeast exstract 0.001 g;
mikroelemen 1 mL; dan ditambahkan akuades sampai 1000 mL. Adapun komposisi mikroelemen adalah sebagai berikut; ZnSO4.7H2O 0.1 g; MnCl2.4H2O 0.003 g;
H3BO3 0.3 g; CoCl2.6H2O 0.2 g; CoCl2.2H2O
0.01 g; NiCl2.2H2O 0.02g; Na2MO4.2H2O 0.9
g; Na2S2O3 0.02 g; dan ditambahkan akuades
samapai dengan 1000 mL.
Metode Penelitian
Fermentasi Pseudomonas sp.
Fermentasi diawali dengan pembuatan inokulan dengan cara menumbuhkan
6
mL) yang berisi 50 mL media mineral yang mengandung 90 mM suksinonitril. Kultur diinkubasi di atas shaker pada suhu kamar selama 45 jam.
Pola pertumbuhan Pseudomonas sp. dilakukan dengan cara menginokulasikan sebanyak 45 mL inokulan ke dalam Erlemeyer (2000 mL) yang berisi 800 mL media pertumbuhan yang mengandung 90 mM suksinonitril dan diinkubasi pada suhu kamar. Setiap 3 jam, sampel diambil secara periodik untuk diamati pertumbuhannya pada OD 600 nm, perubahan pH, aktivitas enzim, kadar protein, konsentrasi NH4
+
dan asam suksinat yang terbentuk.
Produksi Biomassa Sel
Produksi biomassa sel dilakukan dalam Erlemeyer 3 liter yang mengandung 1.5 liter media pertumbuhan dan diinkubasi pada suhu kamar. Sel dipanen pada waktu fermentasi yang menunjukkan produksi sel optimum (waktu yang menunjukkan nilai OD maksimum selama fermentasi). Pemanenan sel dilakukan dengan mensentrifus larutan dengan kecepatan 10.000 rpm selama 15 menit pada 4 ºC. Pelet yang didapat dicuci dengan bufer fosfat pH 7.2 sebanyak dua kali, supernatan dibuang dan pelet dikumpulkan. Pelet yang didapat adalah Pseudomonas sp. (resting cells) yang akan digunkan untuk penentuan karakteristik enzim dalam sel utuh.
Penentuan Aktivitas Enzim
Campuran reaksi yang telah mengandung 1.5 % (b/v) suksinonitril dalam bufer fosfat, diinkubasi selama 15 menit pada suhu kamar. Aktivitas enzim dihentikan dengan penambahan 0.20 mL HCl 4 N, kemudian disentrifuse dan kadar NH4+ dalam supernatan
diuji dengan metode Nessler untuk menentukan jumlah NH4+.
Biokonversi Suksinonitril Menggunakan Sel Utuh
1.5 % (b/v) sel dan suksinonitril 90 mM ditambahkan ke dalam Erlemeyer berisi 50 bufer fosfat pH 7.2, kemudian diinkubasi diatas shaker dan setiap inteval waktu tertentu sampel diambil dan kandungan NH4+
ditentukan dengan metode Nessler.
Penentuan Suhu dan pH Optimum
Pengaruh suhu ditentukan dengan cara menambahkan sebanyak 1.5 % sel (b/v) ke dalam 10 ml bufer fosfat pH 7.0 kemudian diinkubasi pada kisaran suhu 5-60 ºC dalam Erlemeyer tertutup. Aktivitas enzim ditentukan dengan cara seperti tersebut di atas. Penentuan pengaruh pH dilakukan dengan cara yang sama seperti pada penentuan suhu tetapi diinkubasi pada kisaran pH 4−10.
Penentuan Km dan Vmaks
Sebanyak 1.5 % sel (b/v) ditambahkan ke dalam bufer fosfat 10 mL pH 7.2 pada suhu kamar, setelah itu substrat suksinonitril ditambahkan dengan konsentrasi 0−120 mM. Aktivitas enzim ditentukan dengan cara yang telah disebutkan di atas.
Penentuan Kadar Protein
Kadar protein ditentukan dengan menggunakan metode Biuret. Konsentrasi sampel ditentukan berdasarkan kurva standar
BSA (bovine serum albumin).
Penentuan Konsentrasi Asam Suksinat dan Suksinonitril dengan Menggunakan Kromatografi Cair Kinerja Tinggi (KCKT)
Supernatan hasil fermentasi dilarutkan dalam pelarut metanol. Larutan tersebut diambil sebanyak beberapa µL lalu disuntikkan dalam KCKT. Kromatogram yang didapat dibandingkan dengan standar suksinonitril atau asam suksinat untuk menentukan konsentrasi sampel dengan rumus sebagai berikut (Lindsay 1992).
[Standar] standar
area Luas
sampel area Luas
[Sampel]= ×
Kondisi KCKT adalah sebagai berikut, fasa gerak metanol 80%, fasa diam kolom organik C 18 10 µ, dan detektor UV (λ 220 nm).
HASIL DAN PEMBAHASAN
Fermentasi Pseudomonas sp.
7
5,8 6 6,2 6,4 6,6 6,8 7 7,2 7,4 7,6 7,80 15 30 45 60 75
waktu (jam) p H 0 1 2 3 4 5 6 7 8 p e rt u m b u h a n ( 6 0 0 n m )
Gambar 5 Pola dan pH pertumbuhan
Pseudomonas sp. dalam suksinonitril.
Keterangan :
( ) Pola pertumbuhan Pseudomonas sp. ( ) pH pertumbuhan Pseudomonas sp.
Pseudomonas sp. tumbuh melalui empat fasa pertumbuhan yaitu, fasa lag, log, stasioner, dan fasa kematian. Fasa lag berlangsung ± 18 jam. Pada fasa ini pertumbuhannya masih sangat lamban karena bakteri masih beradaptasi dengan lingkungan mediumnya dan amonia yang terbentuk dari hasil hidrolisis suksinonitril masih sedikit, tetapi selama fasa ini terjadi penurunan pH medium pertumbuhan. Hal ini diduga karena terakumulasinya asam suksinat yang terbentuk sehingga pH turun dari 6.58 menjadi 6.41.
Pseudomonas sp. mengalami fasa log selama ± 45 jam, (jam ke 18−63). Selama fasa ini terjadi peningkatan konsentrasi amonium secara cepat dan terjadi peningkatan pH dari 6 menjadi 7. Hal ini dapat disebabkan karena mikrob belum memanfaatkan seluruh amonium yang terbentuk untuk metabolismenya sehingga amonium terakumulasi dalam medium.
Pertumbuhan Pseudomonas sp. mencapai maksimum pada jam ke-66 dengan pH 7.46 dan akhirnya masuk ke dalam fasa stasioner yang singkat. Pada fasa ini penambahan jumlah sel setara dengan kematian sel. Hal ini mungkin disebabkan oleh semakin berkurangnya persediaan nutrien dan terjadinya produk samping yang menghambat pertumbuhan. Jumlah sel aktif tetap, tetapi produksi sel-sel baru sebanding dengan kematian sel yang lain sehingga rata-rata pertumbuhan nol. Fasa kematian diduga terjadi setelah 90 jam pertumbuhan.
Waktu penggandaan (td) dan laju
pertumbuhan spesifik ( ) untuk Pseudomonas
sp. masing-masing ditentukan sebesar 3 jam dan 0.346 jam-1, merupakan nilai yang spesifik dan sangat ditentukan oleh jenis mikroorganisme serta kondisi fermentasi.
pH medium selama pertumbuhan berkisar antara 6.2−8.1 dan nilainya cenderung naik seiring dengan meningkatnya waktu fermentasi. Kecenderungan medium bersifat basa karena dihasilkan NH4+. Hasil yang sama
dilaporkan Fitri (2005), bahwa pH medium fermentasi menggunakan isolat bakteri
Pseudomonas sp. dalam substrat adiponitril cenderung bersifat basa.
Selama proses pertumbuhan sel produk biotransformasi (NH4
+
dan asam suksinat) terbentuk seiring dengan konversi substrat. Pembentukan NH4+ mengikuti pola
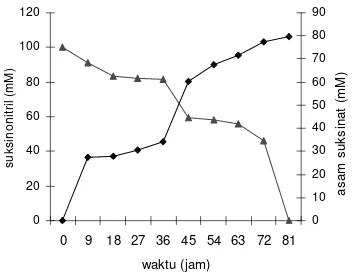
pertumbuhan sel (optimum terbentuk pada akhir fasa log), sedangkan pembentukan asam suksinat mengalami kenaikan seiring dengan bertambahnya waktu fermentasi. Berdasarkan hasil pengukuran dengan menggunakan KCKT dan kromatogram yang terbentuk, kecepatan penguraian suksinonitril lebih cepat dibandingkan pembentukan asam suksinat dengan laju penguraian suksinonitril sebesar 1.235 mM/mL jam yang sudah terurai pada awal pengukuran, sedangkan pembentukan asam suksinat atau produktivitas sel dalam memproduksi asam suksinat sebesar 0.982 mM/mL jam yang terukur maksimum pada akhir pengukuran (Gambar 6).
0 20 40 60 80 100 120
0 9 18 27 36 45 54 63 72 81
waktu (jam) s u k s in o n it ri l (m M ) 0 10 20 30 40 50 60 70 80 90 a s a m s u k s in a t (m M )
Gambar 6 Penurunan konsentrasi suksinonitril dan pembentukan asam suksinat. Keterangan :
( ) Penurunan konsentrasi suksinonitril ( ) Pembentukan asam suksinat
8
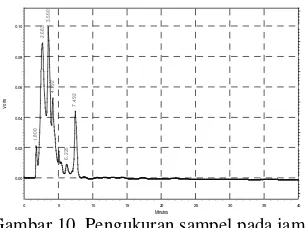
serta asam suksinat dengan luas area 2891620 dan waktu retensi 2.700 yang dijadikan pembanding terhadap sampel yang akan diukur. Pada Gambar 9, terlihat bahwa puncak yang muncul adalah suksinonitril dengan luas area 5939034 dan waktu retensi 4308 dengan konsentrasi optimum 100.0021 mM dibanding pengukuran sampel pada waktu yang lain. Konsentrasi optimum suksinonitril pada waktu awal pengukuran membuktikan bahwa laju penguraian suksinonitril lebih cepat dibandingkan laju optimum pembentukan asam suksinat dengan konsentrasi 86.8488 mM pada waktu akhir pengukuran dengan luas area 2511340 dan waktu retensi 2683 (Gambar 10).
Minutes
0 1 2 3 4 5 6 7 8 9 10
V o lt s 0 .0 0 .1 0 .2 0 .3 0 .4 0 .5 V o lt s 0.0 0.1 0.2 0.3 0.4 0.5 2 .7 5 0 4 .4 2 5
Gambar 7 Standar Suksinonitril.
Minutes
0.00 0.25 0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00 3.25 3.50
V o lt s 0 .0 0 0 0 .0 0 2 0 .0 0 4 0 .0 0 6 0 .0 0 8 0 .0 1 0 V o lt s 0.000 0.002 0.004 0.006 0.008 0.010 2 .1 3 3 2 .7 0 0
Gambar 8 Strandar Asam Suksinat.
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 2 .5 2 5 3 .5 0 0 3 .9 3 3 5 .1 3 3
Gambar 9 Pengukuran sampel pada jam ke-0.
Minutes
0 5 10 15 20 25 30 35 40
V o lt s 0.00 0.02 0.04 0.06 0.08 0.10 V o lt s 0.00 0.02 0.04 0.06 0.08 0.10 1 .8 0 0 2 .6 8 3 3 .5 5 0 4 .1 9 2 6 .2 2 5 7 .4 9 2
Gambar 10 Pengukuran sampel pada jam ke-81.
Perbedaan kecepatan tersebut mengindikasikan bahwa sebagian asam suksinat yang terbentuk digunakan untuk pertumbuhan Pseudomonas sp. Perolehan (Yield) asam suksinat yang didapat sebesar 80% setelah 81 jam fermentasi. Pada sistem fermentasi batch menggunakan Pseudomonas
sp. dalam mengkonversi akrilamida 62.5 mM membutuhkan waktu selama 24 jam, sedangkan apabila sel Pseudomonas sp. diimobilisasi menggunakan kalsium alginat konversi akrilamida hanya membutuhkan waktu kurang dari 6 jam untuk membentuk produknya (Nawaz et al. 1993).
Aktivitas Enzim Pengkonversi Suksinonitril Selama Fermentasi
9
0 0,2 0,4 0,6 0,8 1 1,2
0 9 18 27 36 45 54 63 72
waktu fermentasi (jam)
a
k
ti
v
it
a
s
t
o
ta
l
(U
)
0 20 40 60 80 100 120
a
k
ti
v
it
a
s
r
e
la
ti
f
(U
/m
g
)
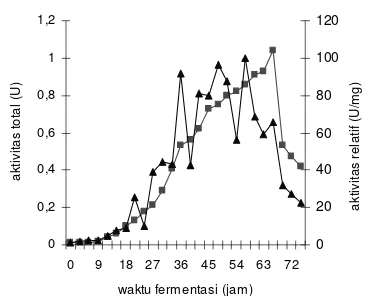
Gambar 11 Aktivitas total dan spesifik NHase selama fermentasi.
Keterangan :
( ) Aktivitas total NHase selama fermentasi ( )Aktivitas spesifik NHase selama
fermentasi
Walaupun tidak dilakukan pengukuran secara kuantitatif, diduga aktivitas total enzim amidase pada suksinonitril mirip dengan aktivitas total amidase pada adiponitril yang juga merupakan dinitril alifatik jenuh. Fitri (2005) melaporkan bahwa dengan substrat adiponitril yang juga merupakan dinitril alifatik jenuh, aktivitas total amidase memiliki nilai 4 kali lebih tinggi dari aktivitas NHase. Tingginya aktivitas amidase dapat diakibatkan karena kondisi medium yang menunjang aktivitas enzim amidase tersebut yaitu suhu, pH, tekanan oksigen, konsentrasi substrat, dan produk-produk yang terdapat dalam media ( Sa’id 1987).
Pengaruh pH Terhadap Aktivitas Enzim NHase
Salah satu faktor yang mempengaruhi aktivitas enzim adalah pH. Pengaruh pH terhadap aktivitas enzim ditunjukkan pada Gambar 12. Aktivitas maksimum NHase dari
Pseudomonas sp. ditunjukkan pada pH 7.
0 20 40 60 80 100 120
0 5 10 15
pH
a
k
ti
v
it
a
s
r
e
la
ti
f
(%
)
Gambar 12 Pengaruh pH terhadap aktivitas relatif NHase dari sel
Pseudomonas sp..
Aktivitas enzim menurun akibat perubahan pH di bawah dan di atas pH optimum, karena perubahan pH medium menyebabkan perubahan pada sifat ionik gugus karboksilat dan gugus amino enzim sehingga daerah katalitik dan konformasi enzim berubah. Enzim merupakan protein yang juga memiliki sifat asam atau basa tergantung dari asam amino penyususnnya. Penurunan pH satu satuan dari pH optimum menurunkan aktivitas relatifnya sebesar 26.13%, sedangkan kenaikan pH satu satuan dari pH optimum menurunkan aktivitas relatifnya sebesar 16.11%. Perubahan pH lingkungan akan mempengaruhi lipatan struktur tersier dan juga struktur kwartenernya. Oleh karena itu, perlakuan pH yang terlalu ekstrim (terlalu asam atau terlalu basa) dapat menyebabkan enzim mengalami denaturasi.
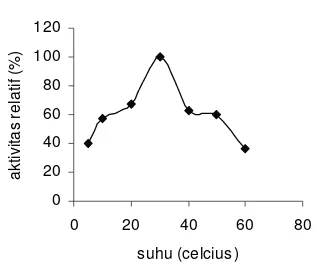
Pengaruh Suhu TerhadapAktivitas NHase
10
0 20 40 60 80 100 120
0 20 40 60 80
suhu (celcius )
a
k
ti
v
it
a
s
r
e
la
ti
f
(%
)
Gambar 13 Pengaruh suhu terhadap aktivitas relatif enzim NHase dari sel Pseudomonas sp..
Penurunan suhu satu satuan akan menurunkan aktivitas relatifnya sebesar 32.72%, sedangkan kenaikan suhu satu satuan akan menurunkan aktivitas relatifnya sebesar 37.13%. Perbedaan nilai ini diakibatkan karena penurunan suhu satu satuan lebih dekat pada kisaran suhu optimum sehingga aktivitas relatif yang diturunkan lebih kecil dibandingkan kenaikan suhu satu satuan. Suhu di atas 40 °C mengakibatkan aktivitas enzim turun dengan tajam. Hal ini disebabkan enzim sebagai protein akan mengalami denaturasi jika suhunya dinaikkan. dan menurut Lee (1992), protein akan terdenaturasi pada suhu sekitar 40−50 °C.
Kinetika Biotransformasi Suksinonitril
Km dan Vmax NHase
Penentuan nilai Km dan Vmax berguna
untuk mengetahui kinetika enzim sehingga dapat diketahui karakteristik dan kecepatan reaksi enzim dalam menguraikan substrat, selain itu Km dan Vmax dapat digunakan untuk
analisis lebih lanjut tentang penghambatan akrivitas enzim dan untuk mengetahui efisiensi aktivitas katalitik enzim. Pengaruh substrat terhadap kecepatan reaksi enzimatik NHase dapat dilihat pada Lampiran 9.
Gambar 14 menunjukkan bahwa enzim NHase memiliki kecepatan reaksi (Vo) akan
mengikuti pola Michaelis-Menten. Fitri (2005) melaporkan bahwa amidase dengan substrat adiponitril juga mengikuti pola Michaelis-Menten sehingga dapat disimpulkan bahwa kedua enzim merupakan enzim alosterik.
0 0,1 0,2 0,3 0,4 0,5 0,6
0 100 200 300
suksinonitril (mM)
V
o
(
u
M
/m
L
.m
in
)
Gambar 14 Pengaruh konsentrasi substrat terhadap kecepatan awal reaksi enzimatik NHase.
Nilai Km dan Vmaks enzim didapat dengan
memplot kurva pada Gambar 14 ke dalam persamaan Line-wever Burk. Nilai Km NHase
sebesar 90 mM dan Vmaks 0.0002 µ M/mL.min.
Nilai Km yang tinggi dapat diartikan bahwa
enzim dapat dengan mudah membentuk kompleks enzim substrat (E-S). sedangkan nilai Km yang kecil menunjukkann bahwa
energi yang diperlukan untuk memulai terjadinya reaksi enzimatik lebih sedikit sehingga reaksi lebih mudah terjadi.
Biotransformasi Suksinonitril
Pola transformasi suksinonitril dan suksinamida oleh Pseudomonas sp. dapat dilihat pada Gambar 15, 16, dan17.
0 10 20 30 40 50 60 70
0 50 100 150 200
waktu menit)
N
H
4
+
(
m
M
)
Gambar 15 Konsentrasi NH4 +
11
0 2 4 6 8 10 120 50 100 150 200 waktu (menit) k e c e p a ta n p e m b e n tu k a n N H 4 + ( m M /m L .m in )
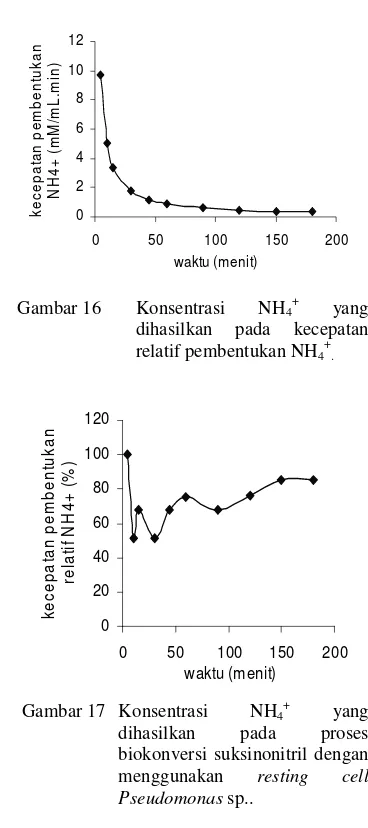
Gambar 16 Konsentrasi NH4 +
yang dihasilkan pada kecepatan relatif pembentukan NH4+.
0 20 40 60 80 100 120
0 50 100 150 200 waktu (menit) k e c e p a ta n p e m b e n tu k a n re la ti f N H 4 + ( % )
Gambar 17 Konsentrasi NH4 +
yang dihasilkan pada proses biokonversi suksinonitril dengan menggunakan resting cell Pseudomonas sp..
Secara stoikiometri, apabila aktivitas amidase tinggi maka jumlah asam suksinat dan amonium yang terbentuk menunjukkan aktivitas NHase yang tinggi juga, karena asam suksinat dan amonium yang dihasilkan oleh aktivitas amidase yang berasal dari suksinamida yang merupakan produk hidrolisis suksinonitril oleh NHase.
Pada Gambar 15, dapat dilihat bahwa konversi suksinonitril selama 180 menit-1 menghasilkan NH4+ sebanyak 59.68 mM
dengan produktivitas sel sebesar 0.33 mM/mL min. Pada biokonversi suksinonitril kecepatan pembentukan relatifnya berkisar antara 51.51%−100%. Biokonversi dengan isolat bakteri terpilih BA 12 (Pamuji 2003) sebesar 4.44 mM pada menit ke-180. Dengan demikian biokonversi Pseudomonas sp. lebih tinggi dibandingkan dengan isolat BA 12.
Pada menit pertama. NH4+ yang dihasilkan
dari biokonversi senyawa amida lebih banyak terbentuk dibandingkan senyawa dinitril, tetapi pada menit-menit berikutnya pembentukan NH4+ dari biokonversi dinitril
lebih banyak dibandingkan amidase dan terus menurun secara signifikan pada menit-menit berikutnya (Fitri 2005).
SIMPULAN DAN SARAN
Simpulan
Sel Pseudomonas sp. yang ditumbuhkan dalam suksinonitril memiliki optical dencity
yang tinggi. Laju pembentukan asam suksinat sebesar 0.982 mM/mL.jam sedangkan laju penguraian suksinonitril adalah 1.235 mM mL.jam.
pH optimum enzim NHase adalah 7 sedangkan suhu optimumnya 27 °C. Km dan
Vmaks enzim NHase adalah 90 mM dan 0.0002
µM/mL/min.
Saran
Sebaiknya perlu dilakukan penelitian lebih lanjut mengenai optimasi media tumbuh dan imobilisasi sel untuk lebih meningkatkan efisiensi pembentukan produk
DAFTAR PUSTAKA
Acharya A, Desai AJ. 1997. Studies on utilization of acetonitrile by Rhodococcus erythropolis A 10. J Microbiol Biotechnol
13:175−178.
Adityarini. 1999. Isolasi dan seleksi mikrob pendegradasi asetonitril serta karakterisasi enzim yang berperan di dalamnya [tesis]. Depok: Program Pasca Sarjana. Universitas Indonesia.
Bandyopadhyay AK et al. 1986. Purification and characterization of benzonitrilases from Arthrobacter sp. strain J-1. Appl Environ Microbiol 51:302−306.
Benesovsky F. 1981. Encyclopedia of Chemical Technology. New York: J Wiley.
12
Pseudomonas chlororaphis B23.
Appl .Environ Microbiol 61:998−1003.
Crueger W, Crueger A. 1984. Biotechnology:
A Text Book of Industrial Microbiology. Madison: Science Tech.
Dixon M & Webb EC. 1978. Enzymes. New York: Academic Press.
Fitri EY. 2005. Kinetika fermentasi dan karakterisasi Enzim yamg terlibat dalam proses produksi asam adipat oleh
Pseudomonas sp. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Holloway et al. 1998. Ability of microbe in strain from Rhodococcus rhodochrous. Am Society for Biochem and Molecul Biol
291:15546−15692.
Kobayashi M, Hidenobu K, Toru N, Hideaki Y, Sakayu S. 1993. Occurrence of amidases in the industrial microbe
Rhodococcus rhodochrous J1.
Biosci Biotech Biochem 57:1949−1950.
Komeda H, Kobayashi M, Sakayu S. 1996. A novel gene cluster including the
Rhodococcus rhodochrous J1 nhIBA
genes encoding a low molecular mass nitrile hidratase (L-NHase) induced by its reaction product. Am Society for Biochem and Molecul Biol 271:15796−15802.
Kobayashi M, Yoshie F, Masahiko G, Hidenobu K, Sakayu S. 1997. Identification of active sites in amidase: evolutionary relationship betwen amidae bond and peptide bond-cleaving enzymes.
Proc Natl Acad Sci 94:11986−11991.
Lee. 1992. Analysis Protein Method. New York: Academic Press.
Liese A, Seelbach K, Wandrey C. 2000.
Industrial Biotransformations. Weinheim: Wiley,VCH.
Lehninger AL. 1994. Dasar-Dasar Biokimia. Jakarta: Erlangga.
Linsay S. 1992. High Performance Liquid Chromatography.. London: J Willey.
Martin DW. 1981. Harper’s Review of Biochemistry. Calipornia: Medical Publication.
Nawaz MS, Franklin W, Cerniglia CE. 1997. Degradation of acrylamide by immobilized cells of Pseudomonas sp. And Xanthomonas maltophilia. Can J Microbial.73:465−576.
Pamuji EW. 2003. Biokonversi Adiponitril menjadi Asam Adipat oleh Isolat Bakteri Terpilih [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Pollak P, Romeder G, Hagedorn F, Heinz PG. 1991. Nitriles. Biotechnol 17:349-361.
Purnomo D.1999. Biokonversi Akrilonitril menjadi Akrilamida dan Asam Akrilat oleh Sel Corynebacterium D5 [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Rahman A. 1992. Teknologi Fermentasi. Jakarta: Arcan.
Sa’id EG. 1987. Bioindustri: Penerapan Teknologi Fermentasi. Jakarta: Mediatama Sarana Perkasa.
Sunarko B. 1996. Kemampuan Isolat Bakteri dalam Mendegradasi Asetonitril J.
Mikrob Trop. 1: 13−19
Tauber MM, Cavaco-Paulo A, Robra KH, Gubitz GM. 2000. Nitrile hidratase and amidase from Rhodococcus rhodochrous
hydrolyze acrilic fibers and granular polyacrylonitriles. Appl Environ Microbial 66:1634−1638.
White D. 1995. The Physiology and Biochemistry of Prokaryotes. New York: Oxford University Press.
Wu ZL & Li ZY. 2002. Enhancement of enzyme activity and enentioselectivity in nitrile via cultivation metabolism by
15
Lampiran 2 Analisis NH
4+dengan metode Nessler (Gerhardt & Krieg 1994) pada
panjang gelombang 400 nm dan kurva standar NH
4+Blanko Standar Sampel
Aquades (ml) 0.1 - -
Standar (ml) - 0.1 -
Sampel (ml) - - 0.1
NaOH 0.1 N (ml) 9.9 9.9 9.9
Pereaksi Nessler (ml) 0.2 0.2 0.2
Konsentrasi amonium Absorbans
(ppm) I II rerata
1 0.0091 0.0091 0.0091
5 0.0210 0.0212 0.0211 10 0.0230 0.0238 0.0234 30 0.0892 0.0892 0.0892 50 0.1183 0.1183 0.1183 70 0.1578 0.1584 0.1581 100 0.1940 0.1942 0.1941 120 0.2372 0.2372 0.2372 150 0.2825 0.2825 0.2825 180 0.3420 0.3422 0.3421 200 o.3992 0.3992 0.3992
y = 0.0019x + 0.0157 R2 = 0.9934
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45
0 50 100 150 200 250
konsentrasi (ppm)
ab
so
rb
an
s
16
Lampiran 3 Pengukuran Kadar Protein dengan Metode Biuret (Wood & Crieg
1994).
Suspensi sel (1 ml) + 0.2 ml NaOH 2 N + 0.4 ml H2O
Rebus 10 menit (100°C)
Dinginkan dengan es
+ 0.2 ml 4 N TCA
Supernatan
Sentrifuse 10000 g, 10 menit Diamkan 20 menit Pellet + Reagen Biuret 5 ml Sentrifuse 10000 g, 10 menit
17
Lampiran 4 Kurva standar protein dan sampel protein selama fermentasi
Kurva Standar
BSA Absorbans
(mg) I II rerata
0.1000 0.0021 0.0021 0.0021
0.0200 0.0041 0.0045 0.0043
0.0400 0.0096 0.0096 0.0096
0.1600 0.0259 0.0259 0.0259
0.2000 0.0340 0.0346 0.0343
0.2400 0.0368 0.0372 0.0370
0.2800 0.0481 0.0483 0.0484
0.3200 0.0541 0.0541 0.0541
y = 0.174x - 0.0026
R2 = 0.9224
0 0.01 0.02 0.03 0.04 0.05 0.06
0 0.1 0.2 0.3 0.4
BSA (mg)
ab
so
rb
a
n
s
Kurva Standar BSA
0 0.005 0.01 0.015 0.02 0.025 0.03 0.035 0.04 0.045 0.05
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48 51 54 57 60 63 66 69 72 75
waktu (jam)
k
a
d
a
r
p
ro
te
in
(
m
g
/m
L
)
18
Lampiran 5 Kromatogram standard dan sample berdasarkan analisis dngan
menggunakan KCKT
No Standar/ Sampel jam ke- Kromatogaram
1 Standar Suksinonitril
Minutes
0 1 2 3 4 5 6 7 8 9 10
V olt s 0 .0 0 .1 0 .2 0 .3 0 .4 0 .5 V olt s 0.0 0.1 0.2 0.3 0.4 0.5 2 .7 5 0 4 .4 2 5
2 Standar Asam Suksinat
Minutes
0.00 0.25 0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00 3.25 3.50
V o lt s 0 .0 0 0 0 .0 0 2 0.0 0 4 0 .0 06 0 .0 0 8 0 .0 1 0 V o lt s 0.000 0.002 0.004 0.006 0.008 0.010 2 .1 3 3 2 .7 0 0
3 0
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 2 .9 1 7 4 .0 0 8 4 .3 0 8
4 9
Minutes
0 2 4 6 8 10 12 14 16 18 20
19
lanjutan Lampiran 5
5 18
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 0.30 V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 0.30 2 .5 9 2 3 .4 0 8 5 .5 3 3
6 27
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lts 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 V o lts 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 2 .5 0 0 3 .3 9 23 .5 3 3 5 .8 8 3 36
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 V o lt s 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 2 .5 0 0 3 .3 9 23 .5 3 3 5 .8 8 3
8 45
Minutes
0 2 4 6 8 10 12 14 16 18 20
20
lanjutan Lampiran 5
9 54
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.00 0.02 0.04 0.06 0.08 0.10 V o lt s 0.00 0.02 0.04 0.06 0.08 0.10 1 .7 0 8 2 .5 6 7 3 .2 5 0 3 .5 3 3 3 .8 0 8 4 .9 0 8 5 .4 0 8
10 63
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.000 0.025 0.050 0.075 0.100 0.125 0.150 V o lt s 0.000 0.025 0.050 0.075 0.100 0.125 0.150 1 .6 5 8 2 .7 5 8 3 .1 1 7 3 .5 0 8 3 .8 0 8 4 .9 2 5 5 .4 6 7
11 72
Minutes
0 2 4 6 8 10 12 14 16 18 20
V o lt s 0.00 0.02 0.04 0.06 0.08 0.10 0.12 V o lt s 0.00 0.02 0.04 0.06 0.08 0.10 0.12 1 .7 5 8 2 .6 2 5 3 .1 9 2 3 .4 3
3 3.90
0
5
.7
0
8
12 81
Minutes
0 5 10 15 20 25 30 35 40
21
Lampiran 6 Konsentrasi Suksinonitril dan asam suksinat selama fermentasi
berdasarkan KCKT
Waktu Suksinonitril Asam Suksinat
(Jam) Rt L.area (mM) Rt L.area (mM)
Standar 4.425 5938905 100 2.700 2891620 100
Standar
0 4.308 5939034 100.0021 0 0 0
9 3.500 5763214 97.0417 2.525 671458 23.2208
18 3.408 5419637 91.2565 2.592 913675 31.5973
27 3.392 5134162 86.4496 2.500 115397 38.9192
36 3.492 4861294 818550 2.525 1461155 50.5306
45 3.900 4541315 76.4672 2.550 1866938 63.8720
54 3.533 4006315 67.4588 2.567 1951347 67.4828
63 3.508 3847462 64.7840 2.758 2017936 69.7856
72 3.900 2001705 337049 2.625 2341625 80.9796
81 0 0 0 2.683 2511340 86.8488
Lampiran 7 Pola pertumbuhan
Pseudomonas
sp. pada suksinonitril
Waktu OD 600 NH4+ pH
(jam) (nm) (mM)
0 0.133 2.212 6.491
3 0.199 2.625 6.524
6 0.231 3.860 6.583
9 0.297 4.210 6.405
12 0.365 4.998 6.66
15 0.451 5.620 6.680
18 0.671 6.214 6.820
21 0.820 7.120 6.838
24 1.010 8.440 6.844
27 1.170 13.230 6.945
30 1.290 14.560 7.061
33 2.010 15.670 7.078
36 2.561 16.680 7.192
39 3.230 17.520 7.116
42 3.870 18.290 7.226
45 4.253 19.200 7.279
48 4.781 19.909 7.280
51 4.790 20.010 7.206
54 5.120 21.620 7.335
57 5.691 22.221 7.524
60 6.020 23.320 7.326
63 6.543 24.620 7.362
66 6.875 28.320 7.445
69 6.225 26.690 7.461
72 5.875 15.311 7.479
22
Lampiran 8 Aktivitas enzim selama proses fermentasi
Absorbans NH4+ Kadar Aktivitas Aktivitas Aktivitas
Waktu NHase rerata (mM) Protein enzim spesifik
Relatif (%)
(jam) I II NHase NHase (mg/mL) NHase (U/mg) NHase
0 0.0101 0.0101 0.0101 0.1013 0.0115 0.01 0.59 1.44
3 0.0101 0.0105 0.0103 0.1323 0.0115 0.01 0.77 1.89
6 0.0135 0.0141 0.0138 0.1923 0.0148 0.01 0.87 2.13
9 0.0274 0.0273 0.0274 0.2769 0.0207 0.02 0.89 2.19
12 0.0312 0.0318 0.0315 0.6737 0.0232 0.04 1.94 4.76
15 0.0281 0.0281 0.0281 0.8314 0.0177 0.06 3.13 7.71
18 0.0286 0.028 0.0283 1.4311 0.0269 0.10 3.54 8.70
21 0.0561 0.0562 0.0562 1.9269 0.0125 0.13 10.32 25.36
24 0.0531 0.0513 0.0522 2.6301 0.0231 0.18 4.14 10.17
27 0.0274 0.0274 0.0274 3.2101 0.0136 0.21 15.76 38.73
30 0.0135 0.0135 0.0135 4.3613 0.0161 0.29 18.04 44.33
33 0.0281 0.0273 0.0277 6.2175 0.0237 0.41 17.51 43.04
36 0.0264 0.0261 0.0263 7.9815 0.0143 0.53 37.34 91.78
39 0.0312 0.0332 0.0322 8.3921 0.0323 0.56 17.32 42.57
42 0.0511 0.0516 0.0514 9.2314 0.0187 0.62 32.91 80.89
45 0.0274 0.0274 0.0274 10.9915 0.0225 0.73 32.51 79.91
48 0.0427 0.0402 0.0415 11.2316 0.0191 0.75 39.14 96.21
51 0.0283 0.0283 0.0283 11.9891 0.0224 0.80 35.68 87.70
54 0.0312 0.0312 0.0312 12.3124 0.0361 0.82 22.74 55.89
57 0.0281 0.0279 0.0280 12.8767 0.0211 0.86 40.68 100.00
60 0.0511 0.0499 0.0505 13.6675 0.0326 0.91 27.95 68.70
63 0.0421 0.0417 0.0419 13.9888 0.0389 0.93 23.97 58.93
66 0.0283 0.0281 0.0282 15.6676 0.0392 1.04 26.65 65.49
69 0.0274 0.0274 0.0274 7.9813 0.0412 0.53 12.91 31.74
72 0.0271 0.0269 0.0270 7.0128 0.0426 0.47 10.97 26.97
75 0.0215 0.021 0.0213 6.3154 0.0458 0.42 9.19 22.60
Ket. Satu unit aktivitas (U) didefenisikan sebagai banyaknya enzim yang memproduksi
23
Lampiran 9 Pengaruh substrat terhadap kecepatan reaksi enzimatik pada
penentuan K
mdan V
makssuksinonitril Absorbans NH4
+
Kec.Reaksi (Vo) 1/S 1/Vo
(mM) (nm) (mM) ( M/ml.min) (1/mM) (1/ M/ml.min)
20 0.3936 25.6621 0.1700 0.0500 5.8452
40 0.4771 31.0594 0.2100 0.0250 4.8294
60 0.5106 33.2235 0.2200 0.0167 4.5148
100 0.3680 72.0349 0.4800 0.0100 2.0823
120 0.3451 67.5969 0.4500 0.0083 2.2190
140 0.3451 67.5969 0.4500 0.0071 2.2190
160 0.3541 67.5969 0.4500 0.0063 2.2190
180 0.3244 63.5853 0.4200 0.0056 2.3590
200 0.3222 63.1589 0.4000 0.0050 2.3749
Lampiran 10 Analisis produk (NH
4+) pada biotransformasi suksinonoitril dengan
menggunakan sel
utuhwaktu Absorbans [NH4
+]
Kec. Pembentukan Aktivitas Relatif
(menit) (nm) (mM) (mM/ ml.min) (%)
5 0.151 48.57 9.71 100.00
10 0.156 50.03 5.00 51.51
15 0.158 50.61 3.37 67.45
30 0.163 52.08 1.74 51.44
45 0.167 53.25 1.18 68.16
60 0.168 53.54 0.89 75.41
90 0.171 54.42 0.60 67.76
120 0.173 55.00 0.46 75.81
150 0.185 58.51 0.39 85.10
180 0.189 59.68 0.33 85.00