PENGGUNAAN
RHIZOBIUM
DAN
BAKTERI PELARUT
FOSFAT PADA
TANAHMINERAL MASAM
UNTUK
MEMPERBAlKl PERTUMBUHAN
BIBIT SENGON
(Paraserianthes
falcafuria
(L.)
Nielsen)
OLEH:
DEN1 ELFIATI
SEKOLAH PASCASARJANA
INSTITUT
PERTANIAN
BOGOR
DENI ELFIATT. Penggunaan Rhizobium dan bakteri pelarut fosfat pa& tanah
mineral masam untuk memperbaiki pertumbuhan bibit sengon (Pmaserianthes
falcutaria (L.) Nielsen). Dibimbing oleh IS WAND1 AN AS, SARWONO HARDJOWIGENO, GLJNAWAN DJAJAKlRANA, YADI SETIADI
dan
LUKMAN GUNARTO,Permasatahan yang utarna dari mah masam adalah cadangan unsur hara dm
kesuburan tanah yang rendah, kandungan Al dapat ditukar tin& dan mempunyai kapasitas retensi P yang tinggi. Kernasaman tanah pada tanaman legurninosa a h
rnenyebabkan terganggunya pertumbuhan tanaman, j urnlah rbizobia dan pembentukan bintil akar berkurang serta terhambatnya fungsi dan perkembangan bintil. Sehubungan dengan ha1 ini penelitian dilakukan untuk mendapatkan isolat Rhljobium asal sengon yang toleran terhadap kemasaman dan Al sekaligus efektif
dalam menarnbat nitrogen serta mendapatkan bakteri pelarut P yang efektif dalarn meningkatkan ketersediam P sehimgga mampu untuk menunjang pertumbuhan
tanaman sengon.
Untuk tujuan tersebut dilakukan penelitian yang terdiri atas: (I) penelitian Rhizobium yang meliputi (a) koleksi isolat Rkizobium dari bintil akar tanaman
sengon, (b) seleksi isolat Rhizobim yang toleran kemasaman dan Al, (c) seleksi
isolat Rhizobium berdasarkan kernampuan menambat nitrogen, (d) penandaan
isolat yang terpilih dengan antibiotik, dan seleksi mutan yang terbentuk, (e)
pengujian efekt ivitas isolat yang terpilih pada tanah masam di rumah kaca; (2) penelitian bakteri pelarut P yang meliputi (a) isolasi bakteri pelarut P, (b) seleksi
isolat bakteri pelarut P dalam melarudcan P dari batuan fosfat dan AlPOs pa& media cair dm
tanah masam,
(c) i d e n t i f h i isolat yang terpilih, (d) penetapan asarn organik isolat yang terpilih, (e) pengujian efektivitas bakteri pelamt P dirumah kaca, (3) koinokulasi antara Rhizobium dan bakteri pelarut P di rumah kaca
dan evaluasi di lapang.
Hasil penelitian menunjukkan bahwa isolat Rhizobium GR2-7 dan GR3-4 toleran terhadap kemasaman dan Al yang tinggi di media cair, sekaligus efektif
&lam menambat nitrogen. Kedua isolat ini mampu meningkatkan bobot kering
tanaman berturut-tumt sebesar 167
dan 132%
dibanding kontml pada Ultisol dan294 dan 265% pa& Inceptisol. Inokulasi dengan isolat Rhizobium GR2-7 dan GR3-4 meningkatkan serapan nitrogen 353 dan 258% pada Ultisol clan 575 dan
642% pada lnceptisol bila dibandin* dengan konlml. Bakteri pelarut
P,
Ertferobacter gergoviae-CKP3-3 dan BaciIw subfilis-GP3-2 efektif dalam
meningkatkan ketersediaan P pada tan& masam yaitu 222 dan 209% dibanding kontrol. Inokulasi kedua bakteri ini dapat meningkatkan bobot kering tanaman sengon beriurut-brut sebesar 93 dm 98% pada Ultisol dan pningkatan yang sama
yaitu sebesar 35% pada Inceptisol, sedangkan serapan
P
meningkat 5 12 dan 496% pada Ultisol dan 307 dan 327% pada Inceptisol dibanding dengan kontrol. Koinokulasi antara Rhizobium dan bakteri pelarut P dapat meningkatkan hampirsemua parameter yang diamati pada Ultisol pada percobaan m a h kaca dan
ABSTRACT
DEN1 ELFLATI. The use of Rhizobium and phosphate solubilizing bacteria in acid mineral soil to enhance seedling growth of sengon (Parnserianthes Sicataria (L.)
Nielsen). Supervised by IS WAND1 AN AS, SARWONO HARDJOWIGENO,
GUNA WAN DJ AJ AKIRAN A, YADI SETIADI AND LUKMAN GUNARTO.
The main problem of acid soil is the availability of nutrients and high exchangeable A1 content. Soil acidity will reduce many legum plant growth, decrease number of rhizobia and nodule initiation and inhibit the function and growth of
ndule. Based on these problems, the experiment was cmducted to obtain the acid
and A1 tolerant strain of Rhizobium isolated from sengon nodules, and to obtain the effective strain of Rhizobium in fixing atmospheric nitrogen, and phosphate solubilizing bacteria in increasing P availability, thus support plant growth.
The study was focused on three aspects: ( I ) Research on Rhizobim consist of (a) collection of Ahitobiurn strains from sengon nodules, (b) screening Rhizobium
toward acidity and high concentration of Al, (c) screening Rhizobium based on their
ability to fix nitrogen, (d) marking the selected Rhizobim with antibiotic, and
screening the resistant mutants, (e) assesing the effectiveness of the selected
Rhizobim on acid soil in greenhouse; (2) Research on phosphate solubilizing bacteria, consist of (a) isolation of phosphate solubilizing bacteria, (b) selection of phosphate
solubilizing bacteria in dissolving P from rock phosphate and Alms in liquid medium and acid soil, (c) identification of selected phosphate solubilizing bacteria, (d) determination of organic acid produced by selective phosphate solubilizing bacteria,
(e) assessing the effectiveness of selected phosphate solubilizing bacteria in
greenhouse; (3) winmulation between Rhizobim and phosphate solubilizing bactetia
in greenhouse and field experiments.
The results showed that Rhizobiurn strains GR2-7 and GR34 were considered
as acid and Al tolerant strains in liquid medium even at pH 4 with A1 concentration in solution until 3.5 ppm. As corn& to the control, strains of GR2-7 and GR34
increased sengon plant dry weight grown in Ultisol by 167 and 132% respectively, and
increased N- uptake by 353 and 258% respectively. Meanwhile, inoculation of GR2-7
and GR34 on sengon grown in Inceptisol, increased plant dry weight by 294 and
269% respectively, N-uptake by 575 and 642%, respectively as compared to the control. Phosphate solubilizing bacteria Enterobmter gergoviaeCKP3-3 and Bmilus subtilis-GP3-2 were selected based on their ability to dissolve unsoluble phosphate.
Isolates CKP3-3 and GP3-2 could increase P availability in acid soil by 222 and 209%
as compared to the control, respectively. Inoculation of phosphate solubilizing bacteria
CKP3-3 and GP3-2 on sengon grown in Ultisol could increase plant dry weight by 93 and 98% respectively as compared to the control, increased plant dry weight by 35% than the control. The similar result were also found in their effect on P-uptake. Sengan grown in Ultisol provided the higher (512 and 496%) P-uptake as compared to the control if it was inoculated by CKP3-3 and GP3-2, respectively, while in Znceptisol these bacteria could increase P-uptake by 307 and 327% respectively as compared to the control. Co-inoculation between Rhizobium (strain GR2-7) and phosphate solubilizing bacteria (CKP3-3 and GP3-2); and between Rhizobim (strain GR3-4) and
phosphate solubilizing bacteria (CKP3-3 and (23-2) provided beneficial effects on
sengon plant when it was grown in Ultisol either under greenhouse study or field
Dengan ini saya menyatakan bahwa Disertasi yang berjudul:
Penggunam Rhizobium dan Brlkteri Pelarut Fosfat pada Tanah Mineral Masam untuk Memperbaiki Pertumbuhan Bibit Sengon (Paraserianthes falcatmia (L.) Nielsen)
adalah benar merupakan hasil karya saya sendiri d m belum pernah dipublikasikan.
Semua sumber data dan infonnasi yang digunakan tehh dinyatakan secara jelas
dan dapat diperiksa kebenarannya
Bogor, Juni 2004
DEN1 ELFZATI
~ ~ E ~ AYKK L 1 UMJ~U-J K l
BIBIT SENGON ( P a m e r i a ~ t h ~ fdcafaria
(L.)
Nielsen)OLEH
DENI ELFIATI
Dhertmi
Sebagi salab satu syarat untuk memperoleb gelar
Doktor pada
Program Studi Ilmu Tanah
SEKOLAH PASCASARJANA
INSTITUT P E R T A M BOGOR
Judul Disertasi : PENGGUNAAN RHIZOBIVM DAN BAKTERI PELARUT FOSFAT PADA TANAH MINERAL MASAM UNTUK
MEMPERBAIH PERTUMBUH AN BIBIT SENGON
(Puraserianthes falcutariu (L.) Nielsen)
Nama Mahasiswa : DEN1 ELFIATI
Nomor Pokok : -985023
Program studi : ILMU TANAH
Menyetuj ui:
Komisi Pembimbing
Dr Ir Iswandi Anas, M.Sc
Ketua
Dr
Ir Y adi Setiadi M.ScAnggota Anggo ta
Ketua Program Studi Ilmu Tanah
Dr Ir Komaruddin Idris MS
Penulis dilahitkan di Batusangkar, Sumatera Barat, pada tanggal 14
Desember 1968. Ayah bemama Jainir dan Ibu bernama Fatirnah. Penulis
mmpakan an& ketiga dari empat bersaudara.
Penulis menyelesaikan Sekolah Dasu (SD) pada tahun 1982 di SD Negeri
7 Batusangkar, m a t SMP Negeri I Batusangkar tahun 1985, lulus SMA Negeri
Batusangkar tahun 1988. Pada tahun 1988 diterima pada Jurusan Tanah Fakultas
Pertanian Universitas Andalas di Padang dan lulus pada tahun 1993. Selanjutnya
pada tahun 1995 penulis mengikuti pendidikan S2 pa& Program Pascasarjana
Universitas Andalas dan lulus tahun 1998. Kemudian pada tahun 1998 penulis
mengikuti pendidikan Doktor (S3) pada Sekolah Pascasarjana Institut Pertanian
Bogor, pada program studi Ilmu Tanah. Selama menempuh pendidikan S3 penulis
mendapat kesempatan menjadi asisten praktikurn pada mata kuliah Biologi Tanah
pada tahun ajaran 1999/2000,2000/200 1,200 1 /ZOO2 dm 2002/2003.
Pada tahun 1998 penuli diterima s e w 1 staf pengajar di Program Ilmu
PRAKATA
Syukur ke hadirat Allah Swt penulis panjatkan karena berkat rahmat dan
karunia-Nya, penulis dapat menyelesaikan rangkaian penelitian dm penulisan
disertasi yang berjudul: Penggunaan Rhizobium dan Bakteri Pelarut Fosfat pada
Tanah Mineral M a m untuk Mem- Pertumbuhan Bibit Sengon
(Parme r i m t h s farcataria
(L.)
Nielsen).Penulis sangat berterima kasih kepada Bapak Dr Ir lswandi h a s , MSG.,
Bapak Prof. Dr Ir Sarwono Hardjowigeno, M.Sc., Bapak Dr Ir Gunawan
Djajakirana, M.Sc., Bapak Dr Ir Yadi Setiadi, M.Sc., dm Bapak Dr Tr Lukman
Gunarto, MS selaku Komisi Pernbim bing y ang telah memberikan saran, arahan
serta petunjuk sejak menyusun rencana penelitian sarnpai penyusunan tulisan hi.
Ucapan terima kasih juga disampaikan kepada:
1. Rektor dan Dekan Sekolah Pascasarjana Tnstitut Pertanian Bogor atas
kesempatan y ang di berikan kepada penulis untuk mengikuti pendidikan
pada P r o m Doktor (S3) pada Program llmu Tanah
2. Proyek Research for Graduate Education (URGE) atas beasiswa yang telah diberikan
.
3. Rektor Universitas Sumatera Utara, dm Ketua Pengelola Program Ilmu
Kehutanan Universitas Sumatera Utara atas segala bantuan y ang telah
diberikan.
4. Teman-ternan HMPIT dan kepada semua pihak yang telah membantu
5. Ayahanda Jainir, Ibunda Fatimah, kakak dan adik atas segala dorongan
sernangat, banhlan yang tulus ikhlas dan doa yang tak pernah henti selarna
menempuh pendidikan sehingga penyelesaian disertasi ini.
Semoga disertasi ini bermanfaat bagi kita semua. Amin.
Bogor, Juni 2004
DAFTAR IS1
...
DAFTAR TABEL...
DAFTAR GAMB AR
...
PEND A mUAN...
Latar Belakang
...
Tujuan
...
Hipotesis
...
TlNJAU AN PUSTAKA...
Tanah Masam dm Permasalahannya
...
Tanarnan Sengon
Rkizobiurn
...
...
Simbiosis Tanaman Sengon dengan Rhizobium....
Hasil-hasil Penelitian Rhizobium pada Tanaman Sengon
...
Pengaruh Faktor-faktor Lingkungan...
Bakteri Pelanit Fosfat
...
Peranan P &lam Tanaman...
Interaksi ~ h i z o bium. Bakteri Palarut Fosfat dan Tanaman
...
BAHANDAN
METODATempat dan Waktu
...
...
Metoda...
Rhizobium...
.
...
2.
Penamaan isolat...
.
3 Uji autentikmi bakteri bintil akar sengon
4
.
Seleksi isolat Rhizobium yang toleran rnasarn dan aluminium5
.
Seleksi isolat Rhuobium berdasarkan kemampuan menambat...
nitrogen6.Penandaan isolat
.
Rhizobium yang terpilih dengan *...
menggunakan ant~brotik7
.
Pengujian efektivitas isolat Rhuobium pada tanah di rumah...
kaca dm uji penguasaan bakteri terhadap bintil
...
Bakteri Pelarut Fosfat.
...
1 Pengambilan contoh tanah...
2.
Isolasi bakteri pelarut fosfat...
...
.
3 Penamaan isolat
..
...
4. Seleksi bakteri pelarut fosfat...
5
.
Identifikasi isolat...
6
.
Penetapan asam organik7
.
Pengujian efektivitas bakteri pelarut fosfat di rumah kaca....
...
Koinokulasi antara Rhizobium dan Bakteri Pelantt Fosfat
...
1.
Pembaan Rumah kaca...
2
.
Permbaaan lapang...
HASIL DAN PEMBAHASAN...
Rkizobiurn
...
1.
Isolasi Rhizobium dari Bintil Akar Tanaman Sengon...
2.
Uj i Autentikasi Rhizobiumxii
4
.
Seleksi Isolat Rhizobium Berdasarkan Kemarnpuan Menambat...
N itmgen
4.1. Bobot Kering Tanaman
...
...
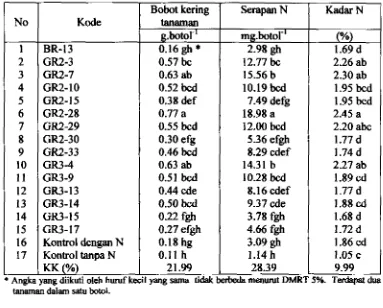
4.2. Kadar dm Serapan N Tanaman
...
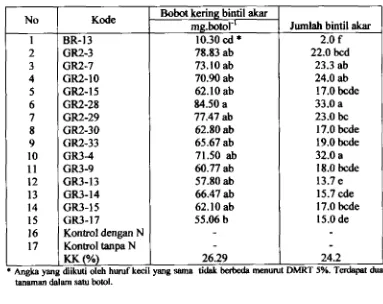
4.3. Jumlah dan Bobot Kering Bintil Akar5
.
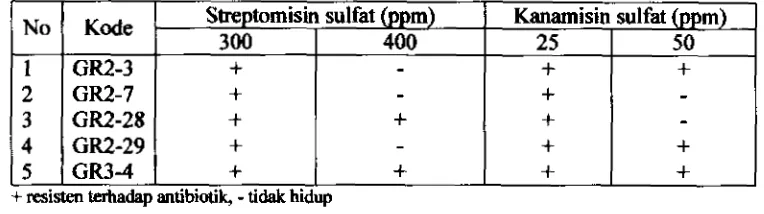
Penandaan Bakteri BintiI Akar Terpilih dengan Antibiotik...
...
5.1. Penandaan Rhizobium dengan Antibiotik
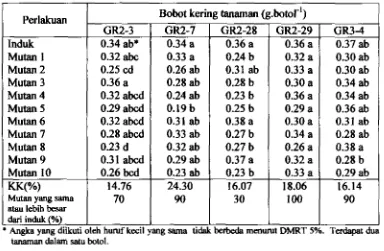
5.2. Bobot Kering Tanaman yang Diinokulasi dengan Mutan
...
5.3. Serapan N Tanarnan yang Di inokulasi dengan Mutan
...
5.4. Kemarnpuan Mutan Membentuk Bintil Akar
...
6
.
Pengujian Efektivitas Isolat Rhizobium pada Tanah di Rumah Kaca dan Uji Penguasaan Bakteri terhadap Bintil Akar...
...
6.1. Kemarnpuan Pernbentukan Bintil Akar
6.2. Kemampuan Okupansi Bintil Akar oleh lsolat Resisten
...
...
6.3. Perturnbuhan Tanaman Sengon...
6.3.2. Diameter Batang...
6.3.3. Bobot Kering Tanaman...
6.4. Kadar dan Serapan N...
Bakteri Pelarut Fosfat...
1.
Isolasi Bakteri Pelarut Fosfat...
2.
Seleksi Bakteri Pelarut Fosfat2.1. Seleksi Kemampuan Bakteri &lam Melarutkan Batuan
...
Fosfat dan A N 42.2. Sejeksi Kemampuan Bakteri dalarn Meningkatkm
...
Ketersediaan P pa& Ultisol dan Inceptis01...
.
...
4
.
Penetapan Asam Organik5
.
Penguj ian Efektivitas Bakteri Pelanrt Fosfat di Rumah Kaca...
...
5.1. P-tersedia tanah...
5 .2
.
Pertumbuhan Tanaman Sengon.
.
...
5.2.1
.
T ~ n g g ~ Tanaman...
5.2.2. Diameter Batang...
5.2.3. Bobot Kering Tanaman
...
5 .3. Kadar P dan Serapan P Tanaman...
Koinokulasi antara Rhizobium dm Bakteri Pelarut Fosfat
...
.
1 Percobam Rumah kaca
...
.
1
.
1 P-tersed ia Tanah...
I.
2. Pembentukan Bintil Akar...
1.3. Pertumbuhan Tanaman.
.
...
1.3.1
.
Tmgg~ Tanaman...
1.3.2. Diameter Batang...
1 -3.3. Bobot Kering Tanaman
...
1 .4. KadarN
dan Serapan N...
1.5. Kadar P dan SerapanP
tanaman...
2
.
Percobaan lapang...
2.1. Tinggi Tanaman....
2.2. Diameter Batang...
Pembahasan Umum...
KESIMPULAN DAN SARANxiv
...
Saran....
DAFTAR PUSTAKA..DAFTAR TABEL
No. Teks Halam an
Karakteristik lahan yang sesuai untuk tanaman sengon
. . .
. . .
.
9Lokasi tempat pengambilan bitil akar sengon
.... . .
. .
,.. . . ...
24Penamaan isolat RhizObium yang diisolasi..
.. . .
. . . .
. . .
.
. . ..
25Kandungan hara Ultisol dan hceptisol yang dipergunakan
. . .
...
32Hasil isolasi Rhirabium dari berbagai lokasi pengambilan bintil akar tanaman sengon
. .
...
... . ... .
...
.
.
.
.
.
.
. ... . ...
. ...
...
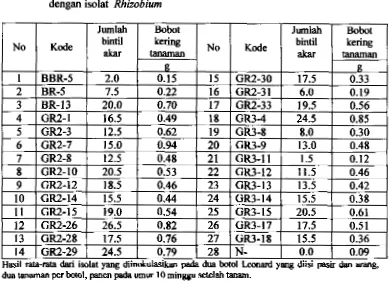
44Jumlah bintil akar dm bobot kering tanaman sengon akibat inokulasi dengan isolat Rhizobium.
. . .
.
.
.
. . . .. . . .
.
. . .
.
. . .
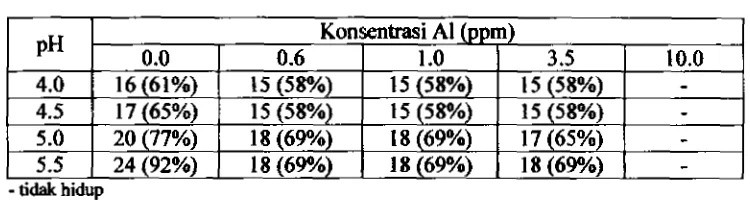
45Jumlah isolat yang berhasil hidup pada pH 4.0 sampai 5.5 dan konsentrasi AlO.O sarnpai 10.0 ppm
. . .
.
.
.
. . .
. . . .
. . .
. . .
. ..
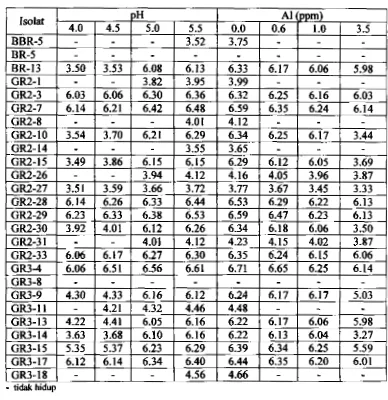
47Populasi (log 10) isolat yang mampu bertahan hidup pada pH 4.0 sampai 5.5 clan konsentrasi A1 0.0 sarnpai 3.5 ppm (7 hari setelah inkubasi).
. ... .
....
. .
... . ...
...,
... . ... . ... . . . ... . ..
48Pengaruh inokulasi Rhizobium terhadap bobot kering tanaman, kadar N dan serapan N tanaman sengon..
.
.
. . .
.
.
.
. . . ....
52Pengad inokulasi Rhizobiwn terhadap bobot kering dan jumlah bintil akar tanaman sengon.
... . .. ... .
..*.. . ...
...
.
...
. ... ..
54Resistensi spontan lima isolat terpilih terhadap antibiotik..
. . . ..
57Bobot kering tanaman yang diinokulasi dengan mutan dm induknya dari kelima isolat yang diuji..
..
.
. . .
...
. .
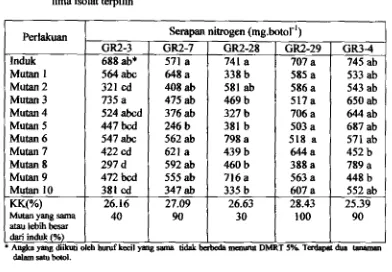
5 8 Serapan nitrogen tanaman sengon y ang diinokulasi dengan mutan dari r i a isolat terpilih... .
.
. . .
.
. .
.
...
. . .
....
6030. Pengaruh koinokulasi antara Rhizobium dan bakteri pelarut P terhadap bobot basah bmtil akar sengon (data ditransforrnasi
It2
dengan (XM.5) ).
. .
. . .
.
. . . .
. . .
. . .
. . .
.
. . .
97 31. Pengaruh koinokulasi antara Rkuobium dan bakteri pelanrt Pterhadap tinggi dan diameter batang tanaman sengon (I0
MST).
. . .
.
. . .
. . . .
. . .
. . .
.
. .
.
. . .
. . .
9932. Pengaruh koinokulasi antara Rkizobium dan bakteri pelarut
P
terhadap bobot kering tanaman sengon (10 MST).
. . .
. . .
.
., 10 133. Pengaruh koinokulasi antara Rhizobium dan bakteri pelarut P
t e r h h p kadar
N
dan serapanN
tanaman sengon (1 0 MST).. ...
10434. Pengaruh koinokulasi antara Rhizobium dan bakteri pelarut P terhadap kadar P dan serapan P tanaman sengon (10
M
SQ.. . .
.
.
. . .
. . .
.
.
.
.
.
. . .
.
. . .
. .
. . .
. . . .
.
. . .
. . .
.
. . .
. .
10635. Pengaruh koinokulasi antara Rhizobium dm bakteri pelarut P terhadap tinggi dm diameter batang tanaman sengon di lapang
(16 MST)
...
.
.
...
109No. Tekr Halaman
1. Kamposisi media yang digunakan dalam penelitian
.... . ... .
...
13 12. Komposisi larutan hara bebas N (Speidel dan Wollum, 1980)..
.
,.
1324. Hasil Analisis Profil Tanah..
. . .
.
. . .
. .
. . .
....
. .. . . ...
1355 , Hasil analisis sifat-sifat kirnia, fisik dan biologi Ultisol
dan
Inceptisol yang digunakan
. . .
. . .
. . .
.
. . .
..
. . .
.
. . .
. .
1366. Hasil analisis sifat-sifat kirnia kompos (bahan pembawa).
. .
. . .
..
1377. Hasil analisis beberapa sifat kimia batuan fosfat Ciamis..
. . .
..
.
1378. Hasil isolasi Rhizobium dari biitil aka- yang diambil dari enrun
DAFTAK
GAMBAR
No.
1.
2.
No.
1.
2.
Penyiapan botol Leonard yang dimodifikasi (Sagiman, 200 1).
....
26Pengaruh inokulasi Rhizubium terhadap bobot kering tanaman
mgon (1 0 MST).
...
70Perbedaan pertumbuhan tanaman sengon pada Ultisol dan
Inceptisol akibat inokulasi dengan Rhizobium..
...
77Pengaruh inokutasi bakteri pelarut P terhadap P-tersedia tanah..
.
83P e r k b a n pertumbuhan tanaman sengon pada Ultisol dan
Inceptisol akibat inokulasi dengan bakteri pelarut P
...
92Lam piran
Uji autentikasi
bakteri
bintil akar Rhizobium.....
154 Uji efektivitas penambatanN
oleh RhRkbium pada tanamansengon di rurnah kaca pada Ultisol (foto atas) dm Inceptisot
...
(foto bawah). 155
Penampakan maman sengon di rurnah kaca pada Ultisol akibat
koinokulasi antara Rhizubiuw dan bakteri p e h t P (10 MST).
...
156Penampakan tanaman sengon di rumah kaca pada Inceptisol
akibat koinokulasi mtara 1Phkubium dan bakteri pelarut P (10
23. Sidik ragam pengaruh koinokulasi antma Rhizobium dan bakteri
p e h t P terhadap tinggi dan diameter batang tanaman sengon di
Latar Belakang
M a n kering di Indonesia yang didominasi oleh tanah-- masam
mempunyai areal yang cukup luas. Tanah masam tasebut di antaranya adaiah
Ultisol dm sebagian Inceptisol. Pusat Penelitian Tanah dan Agroklimat (2000)
melaporkan bahwa dari 188.2 juta ha total luas tanah di Indonesia sebaran Ultisol
menapai 45.8 juta ha (24.3%), dangkan Inceptisol mencapai 70.5 juta ha
(37.5%) yang 5.2 juta ha (7.4%) di antaranya bersifat masam.
Pada kondisi tsopika basah dengan curah hujan yang tinggi, tanah-ternah ini
mengalami pencucian tinggi. Kondisi ini menyebabkan tanah mempunyai reaksi
masarn, kandungan liat tinggi yang didominasi oleh tip liat 1 : 1, cadangan unsur
hara dan kesuburan tanah rendah, kandungan A1 dapat ditukar (Al-dd) tinggi, dan
kapasitas retensi P juga tinggi (Sanchez, 1992; Hardjowigeno, 1993). R&i tanah
yang masam pada sebagian besar tanaman leguminosa menyebabkan terganggunya
pertumbuhm tanaman, jurnlah rhizobia dan pembentukan biitil akar berkurang
serra terhambatnya fungsi dan perkembangan bintil (Reddell, 1993). Untuk
mengatmi keadaan tersebut clapat dilahkm dengan beberapa cara di antaranya
yaitu penggunaan tanaman dan mikroorganisme tanah yang toleran dengan kondisi
tanah yang masam.
Tanaman sengon (Parmeriantks faIcafaria (L.) Nielsen) merupakan salah
satu jenis tanaman yang dapat digumkm, kmm tamman ini dapat tumbuh pada
kondisi tanah yang sangat masam dengan kejenuhan Al mencapai 60% (Binkley
dan Giardina, 1997). Sengon merupakan p h o n leguminosa yang tumbuh
d i m a n f a a h untuk kayu olahan dan bahan baku industri bubur kertas. Daun
sengon berpotensi untuk pupuk hijau dan pakan ternak. Pohon ini dapat
bersimbiosis dengan Rhizobiurn dalam menambat
Nz
(Suharti et dl., 1991). DiIndonesia tanaman sengon umumnya banyak tumbuh pada jenis tanah Ultitisol dm Inceptisol, sehingga pada penelitian hi jenis tanah yang digunakan adalah Ultisol
dan inceptisol.
Untuk menunjang simbiosis yang efektif antara Rhizobium clan tanaman
sengon, salah satu strategi yang dapat dilakukan &ah dengan rnelakukan se1eksi
untuk mendapatkan Rhizobiarns yang toleran t e h d a p tanah masam dan kandungan
Aldd tinggi. Penelitian mengenai simbiosis Rhizobium dm tanaman sengon sudah banyak dilakukan di Indonesia (Danimihardja et af., 1988; Agustiyani, 1 99 1 ; K m n o dan Prana, 1994; Wasis, 1996; Kmono, 2 0 ) , namun belurn banyak
dilaporkan penelitian yang memfokuskan mengenai penggunaan Rhizubium yang
toleran terhadq tanah masam dan dapat bersimbiosis dengan baik dengan tanaman
sengon.
Fosfor
(P)
merupakan salah satu unsur hara makro esensiril untuk pertumbuhan tanaman dm merupakan salah satu faktor pembatas dalarnpertumbuhan maman. Pada tanaman legum, unsur P diperlukan untuk
merangsang penambatan
Nz
melalui peningkatan jumlah bintil pada perakaransehingga dapat meningkatkan kandungan nitrogen dan pertumbuhan tanaman
(Subba Rao, 1994). Pada tanah masam P bemyawa membentuk AI-P, Fe-P dm
occlude&P yang sukar larut. Hal ini menyebabkan pupuk P yang dl'berikan menjadi kurang efisien, sehingga perlu diberikan dalam takaran yang tinggi.
Menurut Jones (1 982) tanman mernanfaatkan P h y a sebesar 10-30 persen dari
Kekurange fisienan penggunaan pupuk P ini &pat diatasi dengan berbagai
cam Salah satu di antamya dengm memanfaatkan balcteri pelarut
P.
Manfaatbakteri pelarut P terhadap tanman telah lama dilaprkan (Gemsen, 1948).
Berbgai penelitian yang melibatkan bakteri pelarut P menunjukkan bahwa bakteri
pelarut P tertentu mempunyai kemarnpuan mendorong proses pelamtan P &lam
senyawa yang relatif sukar larut dengan cara rnengeluarkan asam organik dari
&lam selnya. Asarn organik ini akan mengkhelat Fe dan A1 yang biasanya
mengikat
P
sehingga kelanrtan P meningkat (Illmer et al., 1995). Di Indonesiapenelitian mengenai bakteri pelarut P sudah dilakukan untuk tanaman pertanian
seperti jagung (Buntan, 1W; Prihatini dan Komarhh, 1991), kacang tanah
(Gunarto, 2000), tebu (Premono, 1994), dan kmbakau (Setiawati, 1998). Belum
banyak laporan yang memfokuskan penelitian mengenai penggmaaa bakteri
pelarut P untuk kehutanan ternw.uk tanaman sengon.
Untuk mendapatkan bibit sengon yang berkualitas tinggi dapat dilakukan
dengan menginokuiasi Rhhubium dan bakteri pelarut P. Hasil penelitian
menunjukkan bahwa berkteri Rhizobium mampu meningkatkan m b u h a n tanaman sengon (Garcia er a!., 1988; Daniiihardja et al., 1988; Ksrsono, 2000). Bakteri pelarut P dapat mehutkan senyawa P yang sangat diperlukan untuk
pembentukan bintil akar, Dengan demikian perlu dihkukan suatu penelitian untuk
mendapadran Rhizobium yang tahan terhadap tanah masam dan Aldd yang tinggi
sekaligus efektif dalam menambat nitrogen serta bakteri pelarut P yang
mempunytii kemarnpuan yang tinggi &lam melepaskan senyawa P yang k&at
Tuj uan
1. Mendapatkan isolat Rhizobium yang toleran rnasam, toleran terhadap Aldd yang tinggi, dan memiliki efektivitas yang tinggi &lam menambat nitrogen
pada tanah mineral masam.
2. Mendapatkan isolat bakteri p e l a t
P
yang efektif dalam meningkatkanketmdiaan P dan serapan P tanaman sengon pada tanah mineral masam.
3. Mernpelajari pengaruh inokulasi isolat Rhizobium dan bakteri pelarut
P
secaratun& serta koinokulasi antara M m y a terhadap efektivitas penambatan
Nz,
serapsrn P oleh tanaman, dm perturnbuhan bibit sengon pada tanah mineral masam baik di rumah h a maupun di Irrpang.
Hipotah
Inokulasi isolat RhiZobim dan bakteri p e k t P secara tun& serta koinokulasi antara keduanya pada tanah mineral rnasam &pat meningkatkan
penambatan
Nz,
serapan P oleh maman, dan %at mem- perhunbuhanTINJAUAN PUSTAKA
Tanab Masam dan Permasahbaanya
Kemasaman tanah yang tinggi m e r u m suatu rnasalah utama yang sering
diternui pada tanah-tanah di wilayah M l i m tmpika basah. Tanah ymg
digolongkan ke dalam tanah masam adalah tanah yang nilai pHnya k m g dari 5.5
(Nyakpa et al., 1988; Hardjowigeno, 1992; Tan, 1998). Menurut Pusat Penelitian
Tanah dan Agroklimat (2000) yang tergolong pada h a h masam tersebut d q a t
termasuk ordo Inceptisol, Ultisol, Entisol, Oxisol dan SNosol.
Inceptisol merupakan tanah ymg belum matang dengan perkernbangan
profil yang lebih lemah dibanding dengan tanah matang dm masih banyak
menyerupai sifat bahan induknya (Hardjowigeno, 1993). Menurut S u b o et a!.
(2000), Inceptisol mempunyai &si tanah yang masam sampai netral. Kandungan
bahan organik, kandungan P-potensial dan K-potensid bervariasi dari rendah
sampai sedang. Jumlah basa-basa dapat tukar, kejenuhan basa dan kapasitas tukar
kation bervsriasi dmi rendah sampai tinggi. Potensi kesuburan alami lnceptisol
krkisar dari rend& sampai tinggi.
Ultisol b l a h tanah dengan horison argilik, bersifat masam dengan
kejenuhan basa (jumlah kation), pada kedalaman 180 cm, kurang dari 35% dan
umumnya berkembang dmi bahan induk tua. Di Indonesia banyak ditemukan di
d a e d dengan bahan induk batuan liat (Hardjowigeno, 1993). Ultisol mempunyai
reaksi tanah sangat masam sampai rnasam. Kandungan P-potmsial dm
K-
poknsial serta kandungan bahan organik rendah. Jumlah basa-basa dapt tukar,
kejenuhm basa dan kapasitas tukar kation rendah. Potensi k e s u b m alami Ultisol
Entisol merupakan tanah mineral yang masih b u r muda mu baru
b e r k a b & tanah baru dien@kan atau masih & i t mengalmi p d a p b (Hrlrdjowigmo, 1993). Menurut Subagyo et al. (2000) reaksi tanah pada Entisol bervariasi dari masam sampai agak masam, jumlah basa basa dapat idm, w i t a s
tukar kation dan kejenuhan basa sebqian tergolong rendah dm sebagian d a n g
sampai tinggi. Potensi kesuburan alami Entisol bervariasi dari rendah sampai
tinggi.
Oxisol adalah tanah-tanah yang sudah mengalami pelapukan yang sangat
lanjut, mempunyai kandungan oksida-oksida Fe dan A1 relatif tinggi, kandungan
mineral mudah lapuk rendah dan k m d u n p liat tinggi tetapi tidak aktif sehingga
kapasitas tukar kation rendah (kumng dari 16 cmol(+).kg-' liat) (Hardjow igeno,
1993). Oxisol mernpunyai reaksi tanah yang sangat masam sampai masam, jurnlah
basa-&a dapat tukar, kandungan P dan K-potensial kejenuhan basa, dan kapasitas
hrkar k d o n sangat rendah. Potensi k e s u b m almi Oxisol sangat rendah sampai
rendah (Subagyo et al., 2000).
Spodosol rnerupdm tanah-tanah di mana di horizon bawah terjadi
penimbunan Fe dm Al-oksida dan humus (horizon spodik) sedangkan di lapisan
atas tenlapat horizon eluviasi yang berwarna pucat (Hardjowigeno, 1993).
Spodosol mempunyai reaksi tanah yang sangat masam, kmdungan organik, kmdungan P dan K-potensial sangat rendah, Jumlah basa-basa dapat
tukar,
kejenuhan dan kapasim tukar kation sangat rendah. Potensi kesuburan ahmi
Spodosol sangat rend& (Subsrgyo et al., 2000).
Menurut Sanchez (1992) dan Tan (1998) masalah y a w dhada~i
pads tanah masam adalah tingginya kandungan Al drln Mn yang Itrut d W t
ini akan rnenyebabkan P rnenjadi terikat dan kersediaannya rnenjadi berkurang
untuk tamman. Di samping itu juga terjadi kahat unsur ham N,
K,
Ca,Mg
dan Mo.Unsur
M o
sangat diperlukan oieh tanaman legum untuk pemkntukan bintil akar,sehingga bila ketediaannya rendah akan mengharnbt terjadinya proses
penambatan nitrogen. Menurut Bohn et al. (1 985) p d tanah masam dengan pH kurang dari 5.0 bentuk Al yang dominan addah A?+.
Flis et nl. (1993) menyatakan bahwa pertumbuhan dan pembintilan akar
dari kberapa tanaman legum dipengaruhi oleh A1 yang tenlapat @a tanah.
Secara langsung Al akan mempengamhi pertumbuhan dan survival serta jumlah
rhizobia di dalam tanah, ymg pada akhimya akan mempengtlruhi pembentukan
b i i l .
Menurut Anas (1989) kisaran pH optimal untuk bakteri bintil akar sedikit di
bawah netral hingga agak basa. Pada pH 5.0 beberapa strain bintil akar mas&
&pat hidup. Pada pH yang rendah masalah-masalah yang Mubungan dengan
pernbentukan bintil akar merupakan masalah yang rurnit, karena terdapat tiga
madah yang saling berkaitan dan perlu dipedmhn yaitu: (1) faktot-f&or di
dalam tanah yang dipengaruhi oleh pH yang langsung berpengaruh pada strain
bakteri bintil akar sebagai mikrosimbion, (2) kkbr-f8ktor di dalam tanah ymg
dipengaruhi pH yang langsung berpengaruh pada tanaman dan (3) hktor-faktor
&lam tanah ymg dipenganrhi pH yang mempengaruhi proses mfeksi hktmi bintil
akar pada akar tanaman dan interaksi antma bakkri bintil akar dengan tanaman
Taaaman Sengon
Sengon (Paraserionfks fakataria (L) Nielsen) yang &ulu dikenal
dengan nama Albizia fakataria
(L.)
Fosberg menlpakan yang termasukke dalarn suku Mimosaf.~ole. Tanaman ini merupakan tanaman asli Indonesia yang
berasaI dari kepulauan Maluku dm Irian Jaya, clan pertama kali dibawa ke kebun
Raya Bogor pada bhun 1871, kemudian disebarluaskan ke seluruh nusantara
(Suharti ei al., 199 1).
Pohon hi pernah dmyatakan sebagai pohon ajaib (miracle tree) karma sebagai pohon-pohonm dapat turnbuh dengan cepat. Bila ditanm pada tanah yang
subur dan iklim yang sesuai tingginya dapt mencapai 7 m pada umur 1 tahun, 18
rn pada umur 3 tahun dan 30 m pada umur 9-10 tahun. Tinggi maksimurn sekitar
45 m. Dsllam kondisi optimum pertambahan diameter lingkaran batang setmar 5-7
cm per tahun (Satjapraja dan Las, 1989).
Tanaman sengon dapat turnbuh baik daemh yang W i m lembab dan
panas sehingga sangat sesmi untuk daerah tropis basah yang subur. Pada kondisi
agroklimat yang demikian, tangunan ini dapat tumbuh dengan baik dan subur
sehingga pengembangannya sebagai salah satu tanaman hutan tanaman industri
(HTI) akan menguntungkm (Deparkmm Kehutanan, 1990). Pada daerah yang
kurang subur dm pada daerah dengan ikiim yang agak kering, tanaman sengon
juga &pat tumbuh, akan tetapi m b u h a n n y a ti& s e w dan sesubur seperti yang diharapkan. Dalam keadaan demikian diperlukan pemupukan dan peny iramm
untuk mendapatkan pertumbuhan yang baik (Fmjadinata dan Masano, 199 1).
Sengon dapat tumbuh pada sebagii besar jenis tanah kecuali Vertisol,
untuk membuat peti, clapat digunakan untuk bhan bangunan rumah, serta sebagai
Wan baku kertas,
anak
korek api dan tusuk gigi; (3) dam sengon dapat digunakansehgai makanm temak dan sebagai pupuk hijau.
Menurut Departemen Kehutman (1990) di propinsi Jawa Barat, kabupaten
Sukabumi dan kabupaten Bogor merupakan tempat pertanaman sengon yang terluas. Di propinsi Jawa Tengah, kslbupaten Wonosobo merupakan tempat
pertanaman sengon yang terluas, sedangkan di propinsi Jawa Timur, tempat
pertanaman sengon yang terluas addah kabupaten Kediri.
Di pulau Jawa sengon terkenal se@i tanaman hutan rakyat. Banyaknya
masyarakat menanam mgon berkaitan dengan meningkatnya prmmtaan dari
sektor industri dan juga dimotivasi oleh besarnya keuntungan yang
aksn
diproleh.Seeam ekonomis kayu sengon rnemang memberikan harapan yang besar. Dengan
umur tebang yang berkisar antm 5-6 tahun, tanaman sengon telah berdiameter
antara 20-30 cm atau k o l u m e 0.5 rn3 (Ahosuseno, 1994). Menurut Suryawan
et al. (2000) hslrga kayu sengon di pasaran bebas dapat mencapai Rp 120 000
sampai Rp 130 000 per m3 untuk diameter kayu yang lebh hsar dari 10 cm,
sedangkan hsrrga kayu dengan diameter lebih kecil dari 10 cm harganya seksar Rp
80 000. Dengan perhitungan tersebut apabila dalarn 1 ha terdapat 1485 b m g
mgon dengan jarak tanam 3x2
m
maka petani sengon mampu meraih keunmganseksar Rp 4.2-5.5 juta per ha.
Rkitobium
Famili Rhizobiaceae memiliki 5 genus yaitu Rhizobium, Bra&rhizobium,
Mesorhizobium, Simrhizobium dan Azorhizobium yang diklasifrkasikan
mempakan bakteri yang mempunyai pertumbuhan cepat Vast growing) yang
menghasilkan reaksi asam dalam media agar manitol eksfmk khamir (YEMA)
yang mengandung biru bromtimol (BTB). Koloni Rhizobium pada media agar manitoi ekstrak khamir behntuk bundar dan cembung, tepian licin, konsistensi
lengket dan berlendir setengah ternbus cahaya, &pat mencapai diameter koloni 2 4
mm dengan masa inkubasi 3-5 hari. Rkizobium mempunyai morfologi sel berbentuk batang berukuran 0.5-0.9 x 1.2-3.0 pm, penataan sel dapat tmggal atau
berpasangan, mampu bergerak den@ flagela polar atau sub polar, bersifat gram
negatif, tidak berspora. Sel bakteri tersebut mengandung butiran poli bhidroksi
butirat yang berfungsi sebagai cadzingan makanan untuk sel. Bakteri ini hidup
secara aerobik dan heterotropik dengan memmfaatkan bebempa mawn karbohidrat seperti manitol, glukosa, d m fiuktosa sebagai sumber karbon.
(Alexander, 1978; Subba Rao, 1994; Somasegaran clan Hoben, 1994).
Simbiosi Tanaman Sengon dengan RRbbiwn
Rhizobium dam membentuk suatu sirnbiosis yang saling menguntungkan
dengan tanaman legum. Kerjasama ini diwujudkan dengan pemhtukan bintil
akar pada tanaman hang. Rhizobiurn dapat menembus bulu-bulu akar yang
terdapat pada lapisan tanah permukaan dengan aerasi yang baik.
Di
d a r n bintilakar, Rhizobium dapat menambat nitrogen dari udara tanah dan mengubahnya rnenjadi suatu senyawa yang dibuhrhkan oleh tanman, sdangkan untuk kegiatan
bakteri senyawa karbon diperoleh dari tanaman inang (Paul dan Clark, 1989;
Subba Rao, 1994). Menurut Jensen et d. (1995) pembentukan binti1 akar ini
di antaranya yaitu asam indolasetat
(IAA)
yang berperan penting dalamperhrmbuhan dan perkernbangan b i i l .
Menurut Turk dan Keyser (1W2) tanaman sengon dapat bersimbiosis
dengan genus Ririzobium maupun Bm&rhizobium. Hasil penelitian Mansur (2000) meny impulkan bahwa genus Bradyrhizobiurn yang bersimbiosis dengan
tanaman sengon d i k l a s i f b s i i ke &lam jenis Bmdyrhizobium japnicum dan B.
elkunii. Menunrt Binkley dan Giardina (1 997) genus Rhizobium yang bersimbiosis dengan tmamrm sengon termasuk ke dalam jenis Rhizobium sp.
Proses pembentukan bintil akar menurut
Moat
dm Foster (1995); Graham(1998)
dan
Madigan el d (2000) d a p t melalui tahapan-tahapan sebagai berikut: (1) pemilihan pasangan tanaman yang tepat oleh bakteri bintil akar yangdilanjutkan dengan pelekatan Mkfi pada bulu-bulu akar, (2) penyerbuan M
biintil akar ke &lam bulu-bulu akar melalui benang-benang infeksi, (3) pe jalanan
bakteri bintil menuju akar utama, (4) pembentukan strulaur baktmid dan
dimulainya penambatan nilrogen, (5) pembelahan sel tanaman terus menerus
sehingga terbentuk bintil akar.
Van Rhijn dan Vanderleyden (1 995) menyatakan bahwa pelekatan
Rhizobim pada bulu-bulu akar bgsmtung kepada k e t e p h senyawa
makromolekul yang dikeluarkrln oleh tanaman dengan plisakarida yang terdapat
pada permukaan sel Rhhobiarm. Ssllah satu senyawa p g diiiuarkan oleh
tanaman legum adalah lektin. Senyawa ini dapat meningkatkan pertumbuhan
bslktai di daerah perakamn tanaman.
Menurut Madigan et al. (2000) gen yang berperan dalam pembtukan
menginduksi terjadinya pembengkokan akar rambut dan pembelahan sel tanaman
adalah gen md ABC yang disebut juga sebagai f h r Nods.
Di dalam akar bakteri akan m e m h t u k suatu s t m h r ymg menggembung serta clapat mengikat nitrogen dari u hyang dikenal dengan nma bakkroid.
D
i
dalam bintil yang aktif menambat nitrogen, kehadiran W r o i d ini ditunjukkan
oleh warna kemerah-merahan karma adanya leghemoglobin yang tdcumulasi
pada sel-sel bitil akar (Alexander, 1978; Subba Rm, 1 994; Graham, 1998).
Di dalarn skuktur b d d m i d &&pat enzim nitrogenase y m g merupakan
kunci utama dalarn reaksi penambatan nitrogen. Enzirn tersebut dikenal juga
dengan nama kompleks nitrogen= yaitu surltu enzim yang terdiri dari mtai
polipeptida dan tersusun dari dua kornponen utama: (1) komponen dinitmgenase yang mempunyai rantai Fe-Mo-polipptida dengan berat molekul sebesm 200.000
dan (2) komponen dinitrogenase reduktase yang mempunyai rantai Fe-polipeptida
dengan h tmolekul sebesar 60.000 (Salisbury dan Ross, 1995; Subba Rao, 1994;
Zuberer, 1998; Madigan et al., 2000). Menurut McCardell et d. (1993) dan
Postgate (1998) gen yang terliht dalm proses penambatan nitrogen disebut
dengan gen n$ Gen nif A dan nif
L
mengontrol semm gen nif yang lainnya.Protein F a M o m e r u m kumpulm dari gen nif
K
dm nif D, sedangkan kofaktorFe-Mo yang dibutuhkan untuk mengaktivasi protein Fe-Mo mmpakan kumpulan
dari gen nif$ V,
N
dan E. Protein Fe dibentuk oleh gen nrfH.Pada reduksi
N2
oleh enzim nitrogenase diperlukan ikatan berenergi tinggi(ATP) yang dihasilkan melalui fosforilasi oksidatif. Energi ymg diperlukan untuk
mereduksi N2 menjadi NH3 setam dengan 12 ATP untuk setiap molekul
N2.
Agarkebutuhan energi yang tinggi dapat dipenuhi maka bakteroid meiakukan respirasi
bebas. Sehubungan dengan itu diperlub protein yang mengatur kadar oksigen
sehingga ti& rnernbahayakan nitmgenase, tugas ini dilaksanakan oleh
leghemoglobin. Hasil reduksi
N2
adalahN&
yang kemudian akan tergabungm e m h t u k asam glutamat, glutamin, asam asptat dan dain
(Subba
Rm, 11994;Postgate, 1998). Hasil simbiosis asam amino hi kemudian ditransportasikan ke
bagian tamman melalui jaringan xilem sehingga dapt dipergunakm oleh tamman
inang (Killham dan Foster, 1995).
Kemampuan bakteri bintil akar menyusup ke dalarn akar rambut disebut
infektivitas. Infektivitsls bakteri ditentukan oleh kemampuan bersaing hizobia
dengan bakteri bintil akar alami. Kemmpuan Wompetisi dari Rhizobim
terhdap bakteri bintil akar alami sangat penting dalam memperebutkan tempat
pada rizosfer dan akar rambut. Semakm tinggi kemampuan kompetisi akan makin
besar proporsi bintil akar yang dibentuknya (Danso et ul., 1992). Menurut
Palaniapp et
d.
( 1997) untuk rnendapkm penguasaan (okupansi) wilayah bintilakar oleh rhizobia yang diinokulasii lebih dari 50% diperlukan aplikasi rhiibia
minimal 1 OOO kali lebih banyak dari populmi fhizobia alarni.
Kemampuan bakteri bintit akar d a b menambat
N2
dalam simbiosisdengan tanaman inang disebut efektivitas. Strain Rhizobiwn yang memiliki
efektivitas tinggi &pat menyumbang
N
pada tanaman inangnya &lam jumlahymg besar. Fkda tanaman sengon &tar 40 sam@ 80% nitrogen yang d b b a t
digunakan untuk memenuhi kebutuhan N tanaman (Binkley dan Gidina, 1997).
Hasibbasil Peaelitian RhbbJum pada Tanaman Sengon
Hasil penelitim Sukirnan et al. (1998) pada media pasir steril selama 4
meningkatkan bobot kering bintil
stkar
dan bobot kering tanaman d i b d i g k a ndengan perlakuan kontrol tanpa inokulasi dm tanpa pupuk
N.
Bobot kering tanaman meningkat dari 0.13 pada perlakuan kontrol menjadi 0.44 g padaperhkmn inokulasi dengan isu1at BF 3.1, sedangkm bobot kering bintil
meningkat menjadi 0.32 g pada inokulasi dengan isolat
JF
2 -2.Agustiyani (1991) melaporkan bahwa inokulasi Rhizobium pada tanah
masam dapat meningkatkan tinggi tanaman dan b o b t kering tanaman sengon.
Terjadi peningkatan tin@ dan bobot kering tanarnan dibanding plakuan konml
berturut-brturut sebesar 51 dan 137% akibat inokulasi dengan biak Bio205R.
Selanjulnya hasil penelitian Karsono (2000) menunjukkan bahwa inokulasi
Rhizobium pa& tanaman sengon dapat meningkatkan jumlah clan bobot kering bintil akar sda bobot kering tamman. Bobot kering tamnm meningkat dari 2.06
g pada perlakuan kontrol menjadi 3.3 1 g.pot-' p d a inokulasi dengan Biak No.2-
PF, sedangkan pda koniml dengan pernberian N bobot kering yang dihasilkan
2.71 g.pf'.
Garcia et al. (1988) melaporkan bahwa inokulasi dengan Rhizobium dapat rneningkatkan b o b t kering bintil akar serta meningkatkan pertumbuhm tanaman
smgon. Terjadi peningkatan bobot kering tanarnan s e b 3-12 1% bila d i h d i n g
dengan perlakuan kontrol tanpa N tanpa inokulasi. Selanjutnya dilaporkan bahwa
penggunaan pup& N yang dikombinasikan dengan inokulasi Rhizobium
menyebabkan tertekannya pembentukan bitil akar. Hasil penelitian Wasis (1 996)
menunjukkan bahwa inokulasi dengan Rhizobiran &pat menin- jumlah dan
b o b t bintil akar serta meningkatkan perhrmbuhan tanaman sengon. Terjadi
peningkatan bobot kering tanaman dan indeks mutu bibit bertumt-tumt sebesar 26
(1 997) melaporkan bahwa terjadi penhgkatan bobot kering tanaman sengon dari
0.18 g pala perhkuan kontrol menjadi 0.56 g pada perlakuan inokuiasi dengan
Rhizobiurn.
Dela Cruz et al. (1988) m e l a p o h bahwa inokulasi dengan Rhi~obim
dapat meningkatkan tinggi dm diameter batang tanaman sengon, di rnana te jadi
peningkatm bertunrt-turut sebesar 50 dan 62% dibanding perlakuan kontrol tanpa
inokulasi. Narnun peningkatan tinggi dan diameter batang berturut-turut m e n q a i
600 dan 323% bila tanaman sengon diinokulasi dengan Rhizobium dan mikorh. Hasil yang m a didapatkan oleh Suhardi et al. (1997), di rnana koinokulasi antara
Rhizobium dan mikon'za menghasilkan tin& dm bobot kering tanaman sengon
yang lebih b w dibanding bila diinokulasi dengan Rhizubium saja.
Hasil penelitian Danimihardja et al. (1988) pada tanman sengon di lapang
setelah diinokulasi pada persernaian menunjukkan bahwa pada umur 13 bulan
setelah tanam dapat meningkatkan tinggi dan diameter batang tanaman dl'banding
perlakm konml. Terjdi peningkatan tinggi tanaman sebesar 22% dan diameter
bamg s&sar 53%.
Pengamb Faktor-faktor Lingknngan
Sifat-sifat tanah seperti suhu, kelembaban, kernasaman tanah, unsur-unsur
dan senyawa k i i a tertentu di dalm tanah pada urnumnya mempengaruhi
Rhizobium. Selain itu rltizobia mempunyai musuh dmi yaitu rhizobiofag dan
.
bakteri yang mempakan parasit terhadap Rkuobium (Giller dm Wilson, 1991).
Menurut Subba R.ao (1994) kisaran suhu yang optimal untuk pembentukm
jaringan bakteroid di &lam bintil akar adalah 20-30 "C, di bawah suhu 10 "C dan
Kisaran pH tanah yang sesuai &ah d k i t netral sampai agak alkali, pada
pH tanah 5.0 hanya sebagian rhizobia ymg hidup. Kernasaman tanah kqmgaruh
tehadap perkembangan bakteri bintil a h , tanaman inang dan pembentukan bitil
akar (Graham, 1 998). Kernmaman
tanah
menyebabkan meningkatnya konsentrasiion H
,
Al,Mn
dan Fe, k u m g krsedianya unsur Ca, Mo dm P serh menjdi fhklor p e m h pa& pertumbuhan, kolonimi dm survival b&mi bid1 akar (Giller danWilson, 1991). Jika konsentrasi ~ 1 ~ ' tinggi maka sei bakteri rhizobia dapat diikat
oleh A1 sehingga mengakibatkan pergemkm rhizobia h h m b a t yang memperkecil
kemampuan infeksi dm pembentukan bintil akar (Flis et al., 1993).
Gunarto (1991) menyatakan bahwa banyak rhiwbirr sensitif tehsrlap
kernasaman, namun yang mampu memproduksi ehpolisakarida dapat
menginduksi pembentukan bintil h r pa& tanah masam. ~ r k i z u b i t i m yang
mampu menghasilkan banyak eksopolisakarida dam bertahan hidup pada pH 4.0
dan 4.5. Menurut Cunningham dan Munns (1984) produksi eksoplisakan'da
mempakan suatu strategi untuk menetralisir kondisi lingkungan yang masam dan
efek meracun dari aluminium.
Menurut Reddell (1 993) dan Bmckwell et a/. (1 995) air mempengaruhi gerakan rhizobia di dalm tanah, kecepatan menginfeksi bulu a h , pertumbuhan
dan efektivitas bintil akar. Penambatan
N2
yang optimum tejadi pada kapasitaslapang. K e k u m p air menyebabkan terganggunya pembentukan bintil akar,
s&gkm kelebihan air menyebabkan menurunnya suplai oksigen ke bintil akar.
Bakteri Pelarut Fmfat
Umumnya di dalm tanah ditemukan mikroba p e h t P anorganik sekitm
yang terdiri dari b a k r i dan fungi (Moersidi, 1992). Bakkri yang sering dilaporkan &pat melanttkan P adalah smggota dari genus Pseruhmonas, Bacillus,
Mycobacterium, Micrococcars, Flavobmteriwn
,
Bacterium, Citrobacter danEntembacter (Alexander, 1978; Buntan, 1992; Subba Rao, 1994), sedangkm dari
jenis fungi dilaporkan beberapa AspergiIItu sp, PeniciZlium sp, Sclerofim dm
Fusarium dapat melmtkan Al-P dm Fe-P (Alexander, 1978; Premono, 1994;
Illmer d al., 1 995).
Unsur
P
relatif ti& mudah tercuci seperti unsur N, -pi karena pengaruhlingkungan maka statusnya dapat berubah dari P yang tersedia bagi tanaman
menjadi kurang tersedia yaitu d a m bentuk Ca-P, Al-P, Fe-P, atau occlded-P
(Nyakpa er al., 1988). Sebagai jasad hidup rnikroba melakukan fungsi metabolisme. Dalam proses metabolisme ini stkan dihasilkan senyawa metablit
sekunder berupa asam-asam organik seperti sitrat, mait, oksalat, suksinat,
glukonat dan glikolat (Alexander, 1978; Subba Rao, 1994; Illmer et d , 1995;
Beauchamp dan
Hume,
1997). Asam- asam organik ini dapt melepaskan ikatan Pdari kompleks jempn sehingga &pat dipergunslkan maman. K m m p u m
mikroba pelarut
P
sangat kragam tergantung dari jenis dm daya adaptasi terh-lingkungan h n y a .
Asam organik mampu meningkatkan ketersediaan
P
di dalam tanah melaluibebrapa mekanisme, di antmtnya sdalah: (1) anion organ& bersaing dengan
ortofosfat pada prmukaan tapak jeraprul koloid yang bermuatan positif (Nagmjah
et al., 1970 dalum Premono, 1994); (2) pelepasan ortofosfat dari &&tan logam-P melalui pembentukan kompleks logam organik (Beauchamp dan Hume, 1 997); (3)
1 999). Secara sederhana mekanisme pelarutan P tembut dapat digambarkan
/OH / OH
Al
-
OH + Asam organik ---> Al-
OH+
HzPOi\ I-hPO4 \ Asam organik
Penelitian Buntan (1 992) memperlihatkan bahwa bakteri pelarut P
(Pseudomo~s pufida dan Enterobacfer gergoviae) mampu meningkatkan
kelamtan P, serapan P d m pertumbuhan tanaman jagung pada Ultisol. Inokulasi
dengan
P.
putida dan E. gergoviue meningkatkan bobot kering tanaman jagungberturut-tumt sebesar 65% dan 293'0. Pada tanaman tebu penggunaan bakteri
pelarut
P
(P. Jwrecens, P. pu~ida) dapat meningkatkan bobot kering tanamansebesar 5-40"! dan meningkatkan efisiensi pggunaan pupuk P asal TSP
sebanyak 60- 1 3 5% (Premono, 1 994). Penelitian Setiawati (1 998) pada tanaman
tembakau, dengan menginokulasikan bakteri pelarut P dapat meningkatkan serapan
P clan baht kering tanaman.
Pal (1998) melaporkan bahwa bakteri pelarut P (BaciZlus sp.) pada tanah
yang dipupuk dengan batuan fosfat dapat meningkatkan ketersediaan P, jumlah dm
b b o t kering b i l akar serta hasil biji tanman pada bekrapa tanaman yang
toleran masam Cjagung, bayam dan kacang panjang). Menurut Dubey (1997)
inokulasi dengan
P.
srriata dengan periambahan superfosfat maupun batuan fosfatdapat meningkadran pemkntukan bintil dan serapan N pada tanaman kedelai dan
bakteri ini dapat dikokulturkan dengan Bradyrkizobium japonicum tanpa efek yang
merugikan.
Beberap peneliti mengemukakan bahwa efekti fnya bakteri pelarut
P
tidakdisebabkan karena kemampuamya dalam menghasilkan zat pengatur tumbuh
seperti
IAA
( M a d dan Frankenberger, 1993; Srinivslssln et ai., 1996; Patten danClick, 1996).
Pernoan
P
dalnm TanimaoFosfor disemp tanaman dari larutan tanah terutama dalam bentuk ion
ortofosfat
(&mi
dan ~ ~ 0 4 ~ ~ ) dimma jumlah dari masing-masing bentuktergantung dari pH larutan tanah. Pada pH 7.2 jumlah antara H2P04 dan HPOZ-
relatif sama, di bawah pH tersebut HZP& merupakan bentuk yang dominan,
sedangkan di atas pH 7.2 h t u k ~ ~ 0 4 ' - yang dominan, HPO:~ lebih larnbat
diserap tanaman dibanding HzPOi Wvlin ei al., 1 999).
Salisbury dan Ross (1995) mengemukakan bahwa
P
sangat berpengamhterhsadap perkembangan dan pertumbuhan tanman, karma P merupakan bagian
esensial dari banyak gula fosfat pada nukleotida, seperti DNA dan RNA, serta
hagian dari fosfolipid pada membran.
Fosfor berperan penting &lam penimbunan dan pelepasm energi. Energi
yang didapatkan dari fotosinmis dan metablisme karbohidrat ditumpuk dalm
senyawa fosfat untuk digunakan dalam pertumbuhan dan proses reproduksi.
Fosfor diinkorporasikm &lam hntuk ATP yang rnmpdm p&et energi umum
bagi semua sel hidup (Willet et al., t 998; Havlm el al., 19W).
Dari sejum1ah P total yang terdapat dalam tanaman sebagian besar tmdapat
dalm bentuk persenyawam organik yaitu sewfitin, fosfolipid, nukleoprotein
dan sebagai koenzim tertentu ymg menjsrdi pmmtara di dalam proses metabolisme
(Jones, 1982; Havlin et al., 19W). Hidrolisis fitin secara enzimatik s e m
sedang tumbuh. Fosfolipid yang merupakan cadangan dalarn biji diperlukan untuk
pertumbuhan selanjutnya (Willet et al., 1998).
Soepardi (1983) mengemukakan peranan
P
antara lain penting untukpertumbuhan sel, pemkntukan akar halus dan m b u t akar, memperkuat jerarni
agar tanaman tidak mudah rebah, mernprbaiki kualitas tanaman, pembentukan
bunga, buah dan biji serta rnemperkwt daya tahan tanaman t e d d q penyakit
Interaksi Rhbbium, Bakteri Pelarut Fosfat dan Tanaman
Keberhasi lm inokulasi Rhkob ium pstda tanaman iegurn sangat bergantung
pada kondisi hrva dari lingkungmya terutama fosfor. Fosfor sangat diperlukan
untuk perkembangan tanaman dan pembentukztn bintil akar. Untuk meningkatkan
keberhsilan inokulasi seringkali ditambahkan pupuk P dan atau menginokulasi
dengan mikroba yrulg menguntungbn seperti jamur mikorila vesikuhr arbuskular
dan pelmt fosfat (Danso et al., 1992).
Proses penambatan Nz oleh tanaman legurn yang r n e l i m Rkizobium
dan bakteri p e l a t P dilaporkan berinteraksi seam sinergis. Inokulasi
Bredyrhizobium dan
P.
JIuorescem pads tanman kedelai mmpu meningkatkmpembentukan binti1 akar (Nishijima et at., 1988). Hasil penelitian Dubey ( 2 997) menunjukkan koinokulasi antara P. sfriara dengm Brcsdyrhizobitun japonicum
tanpa ataupun dengan penambahan superfosfat dan baW fosfat meningkatkm
pembentukan bintil akar, kandungan
N
dan P, b o b t kering tanaman sda hasil bijipada tanaman kedelai. Chebotar et al. (2001) melaporkan bahwa koinokulasi
anbra B. japonieum dan
P.
fluoresdem rneningkatkan pemhtukan bintil akar danBAHAN
DAN METODATempt dan Waktu
Penelitian dilaksanakan di Laboratorium Biologi Tanah, k p r k r n e n Tanah
Fakultas Pertanian IPB Bogor, Laboratorium Mikrobiologi Tanah Balai Penelitian
Bioteknologi dan Surnberdsya M e t & Pertanian Bogor, rumah k&ca Fetkultas
Pertanian IPB dan dilanjutkan dengan evaluasi lapangan di Kebun Percobaan
h p u s IPB Damaga Bogor. Penelitian dihkukan prtda November 2000 sampai
April 2003.
1. h b i RhizoBiwu dari biatil akar tanamaa sengon
Percobatin ini bertujuan untuk mengisolrlsi Rhizobimctn sekaligus mengkoleksinya dan menguji keautentihnya sebagai bakteri bhtil dm.
Binti1 akar diambil dari b e h p a pohon sengon yang tumbuh pada dua jenis tanah yaitu Uttisol dan Inceptisol. Pada Take1 2 @at dilihat lokasi tempat
pengambilan bintil a h . Pengumpulan isolat, pemurnian dan wtentikasi bakteri
bintil akar menggunaksln metode yang dijelasksm oleh Somasegatan dan Hoben
(1 994).
Isolasi bkteri bintil akar dilakukan secara langsung dari biti1 akar yring
telah dikurnpulkan. Permukaan bintil disterilkan dengan mencelupkan bintil akar
ke dalam larutan etanol 95% selama 10 detik, dan kemudian direndm &lam
larutan Hz% 5% selarna tiga menit. Setelah itu bintil dibilas dengan air steril