IDENTIFIKASI MOLEKULAR KANDIDAT MUTAN PROTEIN
HETEROTRIMERIK G SUBUNIT α (Gα) DARI KEDELAI
(Glycine max
(L.) Merrill) TOLERAN ALUMINIUM KULTIVAR SLAMET
LURIA MARLINA LIMBONG
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
LURIA MARLINA LIMBONG. Identifikasi Molekular Kandidat Mutan Protein Heterotrimerik
G Subunit α (Gα) dari Kedelai (Glycine max (L.) Merrill) Toleran Aluminium Kultivar Slamet. Dibimbing oleh UTUT WIDYASTUTI dan MUHAMMAD JUSUF.
Protein heterotrimerik G Subunit α (Gα) diduga berperan dalam sistem pertahanan kedelai
terhadap cekaman aluminium dan pH masam. Tanaman kedelai yang diberi cekaman aluminium mengalami penghambatan pertumbuhan serta kerusakan pada akar dan pada kondisi ini ekspresi gen Gα juga meningkat. Penelitian ini bertujuan untuk mengidentifikasi enam nomor tanaman
kandidat mutan Gα secara molekular. Keenam tanaman tersebut ditanam dan diseleksi berdasarkan dua ciri mutan Gα, yaitu tanaman kerdil dan stomata menutup pada siang hari. Diantara nomor-nomor tanaman hasil seleksi dipilih tiga nomor-nomor kandidat mutan untuk identifikasi gen Gα menggunakan DNA genom, yaitu 338/3S1.174/12/1/4, 103/3S3.103/8/5/1/4, dan 344/3S1.196/14/1/8, serta Slamet tipe liar (WT) sebagai kontrol. Sebelum identifikasi tanaman, dilakukan pemilihan kombinasi primer yang didesain dari cDNA gen Gα kedelai dan DNA genom
Gα padi dengan metode PCR. Kombinasi primer yang berhasil mengamplifikasi cDNA gen Gα digunakan untuk mengamplifikasi gen Gα pada DNA genom tanaman kandidat mutan dan Slamet WT. Hasil PCR tiga kombinasi primer (U9-L2, L3-L4, dan U3-L4) berhasil pada DNA genom Slamet WT dan kandidat mutan nomor 344/3S1.196/14/1/8, sedangkan pada DNA genom kandidat mutan 338/3S1.174/12/1/4 dan 103/3S3.103/8/5/1/4 hanya dua kombinasi primer yang berhasil, yaitu U9-L2 dan L3-L4, maka terdapat satu kombinasi primer yang tidak berhasil, yaitu primer U3-L4. Hal ini diduga karena kedua kandidat mutan tersebut telah mengalami mutasi gen Gα pada daerah penyandi primer U3 dan primer L4.
ABSTRACT
LURIA MARLINA LIMBONG. Molecular Identification of Heterotrimeric G Protein α Subunit
(Gα) Mutant Candidate from Aluminum Tolerant Soybean (Glycine max (L.) Merrill) cv. Slamet. Supervised by UTUT WIDYASTUTI and MUHAMMAD JUSUF.
Heterotrimeric G protein α Subunit (Gα) is predicted to play an important role in soybean tolerance mechanism to aluminum stress and low pH. Aluminum stress inhibits soybean growth and induces root damage. The expression of Gα gene is induced under Al stress condition. The
objective of this research was to identify six lines of Gα mutant candidates using molecular
technique. Six soybean mutant lines were planted and selected based on representative characters of Gα mutant, which were dwarf phenotypes and stomatal closure during daylight. Identification of Gα gene on mutant candidates was carried out through PCR method by using genomic DNA as a template from mutant and wild type cv. Slamet as the control plant. Three mutant candidates (338/3S1.174/12/1/4, 103/3S3.103/8/5/1/4, and 344/3S1.196/14/1/8) were chosen from all the selected lines to identify Gα gene. Before gene identification, a proper primer choosed by combined a designed primer from cDNA of Gα gene from cv. Slamet and Gα genomic of rice. A
numbers of primers were generated from rice Gα genomic and soybean Gα cDNA sequences.
Three primers were selected based on the ability of the primer to amplify soybean Gα cDNA cv.
IDENTIFIKASI MOLEKULAR KANDIDAT MUTAN PROTEIN
HETEROTRIMERIK G SUBUNIT α (Gα) DARI KEDELAI
(Glycine max
(L.) Merrill) TOLERAN ALUMINIUM KULTIVAR SLAMET
LURIA MARLINA LIMBONG
Skripsi
sebagai salah satu syarat memperoleh gelar
Sarjana Sains pada
Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul : Identifikasi Molekular Kandidat Mutan Protein Heterotrimerik G Subunit
α (Gα) dari Kedelai (
Glycine max
(L.) Merrill) Toleran Aluminium
Kultivar Slamet
Nama : Luria Marlina Limbong
NIM : G34051816
Menyetujui:
Pembimbing I,
Pembimbing II,
(Dr. Ir. Utut Widyastuti, M.Si.)
(Dr. Ir. Muhammad Jusuf)
NIP: 19640517 198903 2 001
NIP: 19500301 197603 1 001
Mengetahui
Ketua Departemen,
(Dr. Ir. Ence Darmo Jaya Supena, M.Si.)
NIP: 19641002 198901 1 002
PRAKATA
Syukur kepada Allah pencipta langit dan bumi atas setiap penyertaan dan berkat melimpah yang diberikanNya kepada penulis sehingga penulis dapat menyelesaikan karya ilmiah ini dengan baik.
Terima kasih penulis sampaikan kepada Dr. Ir. Utut Widyastuti, M.Si dan Dr. Ir. Muhammad Jusuf selaku pembimbing atas segala bimbingan, nasihat, waktu, dan sarana yang telah diberikan kepada penulis. Terima kasih kepada Kepala Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB Darmaga beserta seluruh staf dan karyawan atas sarana, prasarana, dan pertolongannya selama penulis melakukan penelitian di Laboratorium BIORIN (Biotechnology Research Indonesia-The Netherland). Penulis mengucapkan terima kasih kepada proyek KKP3T dengan No. Kontrak 635/LB.620/1.1/2/2009 atas nama Dr. Ir. Utut Widyastuti, M.Si. atas biaya penelitian yang diberikan. Penulis juga mengucapkan terima kasih kepada Dr. Ir. Miftahudin, M.Si. atas saran dan kritik yang diberikan saat ujian sehingga menambah informasi untuk perbaikan karya ilmiah ini.
Terima kasih kepada Mbak Pepi, Bapak Mulya, Bapak Adi, Bapak Drs. Y. Ulung Anggraito, M.Si., Bapak Muzuni, S.Si., M.Si., Ibu Saleha Hannum, S.Si., M.Si., Ibu Dra. Yohana Caecilia, M.Si., Ibu Dra. Sri Listyowati, M.Si., Ibu Dra. Ratna Yuniati, M.Si., Mbak Niken Trisnaningrum, S.Si., M.Si., Mbak Ulfa Mushofa, S.P., Mbak Ika Atifah Zahro, S.Si., Kak Syahnada Jaya, S.Si., Kak Rizky Darojat, S.Si., Goto Giok, S.Si., dan Fajri atas diskusi, saran, bantuan, dan persahabatan hangat yang diberikan. Terima kasih juga kepada sahabat terkasih Frahel, Ester, Tiur, Robert, dan Dmitry serta teman-teman Biologi 42 yang tidak dapat disebutkan satu-persatu. Secara khusus penulis sampaikan terima kasih kepada orang tua, abang, dan semua saudara atas doa, perhatian, dan kasih yang membuat penulis selalu bersemangat.
Kiranya karya ilmiah ini bermanfaat.
Bogor, Juni 2010
RIWAYAT HIDUP
Penulis dilahirkan di Samosir pada tanggal 16 April 1987 dari ayah bernama J. Rolimto Limbong dan ibu bernama Lemsia Nainggolan. Penulis merupakan anak kedua dari dua bersaudara.
Penulis lulus dari SMA Negeri 1 Subang pada tahun 2005 dan diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI).
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 2
Hipotesis Penelitian ... 2
BAHAN DAN METODE Waktu dan Tempat Penelitian ... 2
Bahan dan Alat ... 2
Metode Penelitian ... 2
Penanaman Biji ... 2
Identifikasi Kandidat Mutan dengan PCR ... 2
HASIL Seleksi Tanaman Kedelai Berdasarkan Persentase Stomata Menutup dan Tinggi Tanaman ... 3
Identifikasi Kandidat Mutan dengan PCR ... 3
PEMBAHASAN ... 5
SIMPULAN ... 6
SARAN ... 7
DAFTAR PUSTAKA ... 7
DAFTAR TABEL
Halaman 1 Hasil seleksi kandidat mutan generasi kedua (F2) kedelai toleran asam kultivar Slamet
(Hartini 2008) ... 2
2 Sekuen basa nitrogen pada primer-primer yang digunakan ... 3
3 Kestabilan genetik persentase stomata menutup tanaman kandidat mutan terpilih dan Slamet tipe liar (WT) ... 4
4 Kestabilan genetik tinggi tanaman kandidat mutan terpilih dan Slamet tipe liar (WT) ... 4
DAFTAR GAMBAR
Halaman 1 Struktur subunit Gα pada padi (Accession: A2Y3B5) ... 1
2 (a) Kondisi stomata membuka pada Slamet tipe liar (WT), (b, c, d) Kondisi stomata menutup pada 103/3S3.103/8, 344/3S1.196/14/1/8, 338/3S1.174/12/1/4 (tanpa panah) ... 4
3 Perbandingan tinggi tanaman Slamet WT dan kandidat mutan 338/3S1.174/12/1/4 pada umur dua bulan ... 4
4 Panjang basa nitrogen antar primer pada cDNA heterotrimerik Gα kedelai ... 4
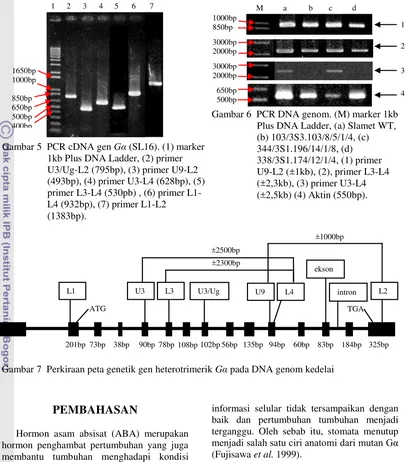
5 PCR cDNA gen Gα (SL16). (1) marker 1kb Plus DNA Ladder, (2) primer U3/Ug-L2 (795bp), (3) primer U9-L2 (493bp), (4) primer U3-L4 (628bp), (5) primer L3-L4 (530pb), (6) primer L1-L4 (932bp), (7) primer L1-L2 (1383bp) ... 5
6 PCR DNA genom. (M) marker 1kb Plus DNA Ladder, (a) Slamet WT, (b)
103/3S3.103/8/5/1/4, (c) 344/3S1.196/14/1/8, (d) 338/3S1.174/12/1/4, (1) primer U9-L2 (±1kb), (2), primer L3-L4 (±2,3kb), (3) primer U3-L4 (±2,5kb), (4) Aktin (550bp) ... 5
7 Perkiraan peta genetik gen heterotrimerik Gα pada DNA genom kedelai ... 5
DAFTAR LAMPIRAN
Halaman 1 Hasil seleksi tanaman kandidat mutan berdasarkan persentase stomata menutup dan
tinggi tanaman generasi ketiga sampai generasi kelima ... 9
2 Kondisi lingkungan saat pengambilan stomata pada F5 ... 10
PENDAHULUAN
Latar Belakang
GTP-binding regulatory protein atau protein G merupakan bagian dari famili protein pengikat guanosin trifosfat yang berperan dalam transduksi sinyal. Protein G dibagi menjadi protein G kecil (small G protein) yang terletak di membran plasma dan protein heterotrimerik G yang berperan meneruskan informasi dari reseptor membran ke efektor intraselular. Protein heterotrimerik
G terdiri atas γ subunit, yaitu subunit α (Gα), subunit (G ), dan subunit (G ) (Ma 1λλ4).
Beragam transduksi sinyal pada mamalia dan tumbuhan telah diketahui melibatkan protein heterotrimerik G khususnya protein hetero-trimerik Gα (Ma 2001).
Pada keadaan tidak aktif protein
hetero-trimerik Gα berikatan dengan dimer G dan
GDP sedangkan pada keadaan aktif protein
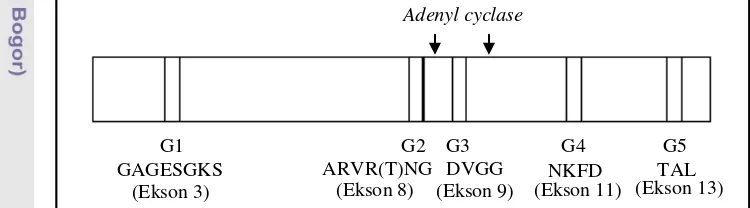
Gα berikatan dengan GTP dan terpisah dengan dimer G (Perfus-Barbeoch et al. 2004). Protein Gα pada umumnya memiliki 5 situs pengikatan GTP, yaitu G1-G5 (Sprang 1997). Penyandi kelima situs tersebut terdapat pada daerah ekson yang berbeda (Gambar 1).
Protein Gα berperan dalam berbagai respon selular, yaitu mengaktifkan enzim adenyl cyclase sehingga dapat mengubah ATP menjadi cAMP untuk menginduksi berbagai respon selular. Protein Gα juga berperan dalam pengaturan kanal ion, salah satunya pengaturan kanal ion K+ pada proses pembukaan stomata (Assmann 1996). Selain itu, protein Gα juga berperan meregulasi 2 jenis second messengers hidrofobik, yaitu inositol 1,4,5-triphosphate (IP3) dan diacylglycerol (DAG). IP3 berperan dalam regulasi produksi H2O2 (Becker et al. 2000).
Tanaman kedelai (Glycine max (L.) Merrill) tidak dapat tumbuh dengan baik pada lahan masam karena lahan tersebut memiliki pH rendah (pH<4) dan tingkat kelarutan
aluminium (Al) yang sangat tinggi (Matsumoto 2000). Aluminium reaktif (Al3+) dapat meracuni akar tanaman dengan menghambat kanal ion K+ pada membran sel penjaga stomata (Perfus-Barbeoch et al. 2004). Pertumbuhan yang terhambat tersebut ditemukan pada kedelai kultivar Slamet pada kondisi cekaman Al dan pH 4 yang ditunjukkan oleh reduksi perpanjangan akar (Mashuda 2006).
Belafiff (2007) membuktikan bahwa transduksi sinyal cekaman Al3+ pada kedelai kultivar Slamet dapat meningkatkan aktivitas
protein Gα melalui peningkatan produksi H2O2. Ekspresi gen Gα juga meningkat pada kondisi cekaman Al dan pH 4 yaitu pada jam ke-24 dan jam ke-48 (Mashuda 2006). Gen Gα diduga berperan dalam sistem toleransi kedelai kultivar Slamet terhadap cekaman Al (Suharsono & Suharsono 2006).
Kedelai kulivar Slamet hanya memiliki satu kopi gen Gα, sehingga keterlibatan gen Gα secara langsung dalam sistem pertahanan kedelai terhadap cekaman Al dapat dibuktikan dengan men-knockout gen tersebut (Suharsono 2004). Hartini (2008) melakukan pendekatan reverse genetic, yaitu dengan
menonaktifkan gen Gα pada kedelai kultivar Slamet dan Lumut melalui irradiasi sinar gamma. Kandidat mutan gen Gα diseleksi berdasarkan ciri-ciri morfologi mutan Gα, yaitu tanaman kerdil (Ma 2001; Fujisawa et al. 1999) dan dengan stomata menutup pada saat siang hari (Perfus-Barbeoch et al. 2004; Nilson & Assmann 2010). Hasil percobaan Hartini (2008), menunjukkan bahwa stomata kedelai kultivar Slamet tipe liar (wild type) membuka maksimal pada pukul 12.00 WIB. Hasil seleksi diperoleh 10 kandidat mutan Gα pada generasi kedua, yaitu 9 tanaman berasal dari kultivar Slamet dan satu tanaman berasal dari kultivar Lumut.
G1 G2 G3 G4 G5
GAGESGKS ARVR(T)NG DVGG NKFD TAL Adenyl cyclase
(Ekson 9)
(Ekson 3) (Ekson 8) (Ekson 11) (Ekson 13)
2
Giok (2010) melakukan pengamatan pada dua tanaman kandidat mutan kultivar Slamet generasi keempat, yaitu 103/3S3.103/8/5/1 dan 338/3S1.174/12/1/4. Pada kedua tanaman tersebut ditemukan adanya kerusakan sel akar di bagian epidermis sampai korteks serta adanya peningkatan reduksi panjang akar yang mencapai 82-96% pada kondisi cekaman 1,6 mM AlCl3 dan pH 4. Hal ini menimbulkan dugaan bahwa kedua tanaman tersebut telah mengalami mutasi pada gen Gα.
Tujuan Penelitian
Penelitian ini bertujuan mengidentifikasi kandidat mutan protein heterotrimerik G
subunit α (Gα) dari kedelai toleran Al kultivar
Slamet secara molekular.
Hipotesis Penelitan
Hipotesis dalam penelitian ini bahwa enam tanaman kedelai kultivar Slamet hasil irradiasi sinar gamma telah mengalami mutasi pada gen Gα.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan mulai bulan Februari 2009 sampai dengan Mei 2010 di Laboratorium BIORIN, Pusat Penelitian Sumber daya Hayati dan Bioteknologi (PPSHB), Institut Pertanian Bogor.
Bahan dan Alat
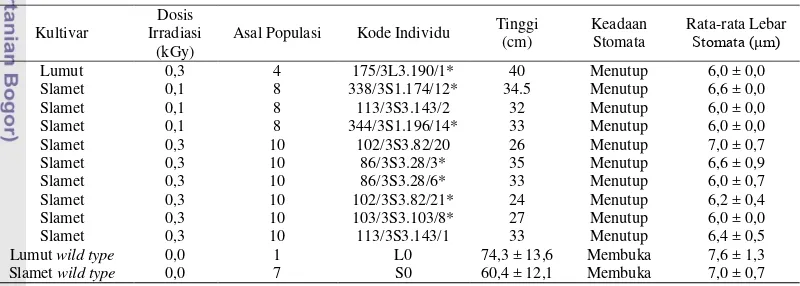
Bahan yang digunakan dalam penelitian ini adalah biji kedelai kultivar Slamet generasi kedua (F2) yang merupakan kandidat mutan protein heterotrimerik Gα dari hasil penelitian Hartini (2008). Kode tanaman kedelai tersebut adalah 338/3S1.174/12, 86/3S3.28/6, 86/3S3.28/3, 344/3S1.196/14, 102/3S3.82/21,
103/3S3.103/8, serta Slamet wild type (Tabel 1).
Alat yang digunakan adalah penggaris skala 50 cm, 4 in 1 environment tester, mikroskop cahaya, kamera digital, autoklaf, mortar, pestel, sudip, tube 1.5 ml, tip, pipet mikro, mesin sentrifus, mesin elektroforesis, UV transiluminator, mesin spektrofotometer, dan mesin PCR.
Metode Penelitian Penanaman Biji
Biji kedelai ditanam di lahan seluas 6,5x4 meter dengan jarak tanam 40x20 cm. Pemupukan dilakukan dua minggu setelah tanam dengan pupuk Urea/TSP/KCL dosis 50/100/50 kg/ha. Penyiangan dilakukan setiap dua minggu. Hama diatasi dengan Acodan dosis 0,5 ml/l dengan dosis penyemprotan 500 l/ha. Setelah tanaman memiliki cukup daun, dilakukan pengamatan terhadap kondisi stomata saat pukul 12.00 WIB dengan metode nail polish swath. Stomata kedelai membuka
maksimal pada pukul 11.00 sampai 12.00 WIB (Hartini 2008). Pengamatan tinggi tanaman dilakukan pada saat muncul bunga dan pada saat panen. Tanaman diseleksi berdasarkan kondisi stomata dan tinggi tanaman. Tanaman hasil seleksi ditanam kembali pada penanaman berikutnya, yaitu sampai generasi kelima (F5). Penanaman dilakukan tiga musim tanam untuk memperoleh kandidat mutan yang cukup stabil berdasarkan kondisi stomata dan tinggi tanaman.
Identifikasi Kandidat Mutan dengan PCR DNA genom kedelai diisolasi menggunakan Qiagen Dneasy Plant Mini Kit. Primer-primer yang digunakan untuk PCR Kultivar
Dosis Irradiasi
(kGy)
Asal Populasi Kode Individu Tinggi (cm)
Keadaan Stomata
Rata-rata Lebar
Stomata ( m)
Lumut 0,3 4 175/3L3.190/1* 40 Menutup 6,0 ± 0,0 Slamet 0,1 8 338/3S1.174/12* 34.5 Menutup 6,6 ± 0,0
Slamet 0,1 8 113/3S3.143/2 32 Menutup 6,0 ± 0,0
Slamet 0,1 8 344/3S1.196/14* 33 Menutup 6,0 ± 0,0
Slamet 0,3 10 102/3S3.82/20 26 Menutup 7,0 ± 0,7
Slamet 0,3 10 86/3S3.28/3* 35 Menutup 6,6 ± 0,9
Slamet 0,3 10 86/3S3.28/6* 33 Menutup 6,0 ± 0,7
Slamet 0,3 10 102/3S3.82/21* 24 Menutup 6,2 ± 0,4 Slamet 0,3 10 103/3S3.103/8* 27 Menutup 6,0 ± 0,0 Slamet 0,3 10 113/3S3.143/1 33 Menutup 6,4 ± 0,5 Lumut wild type 0,0 1 L0 74,3 ± 13,6 Membuka 7,6 ± 1,3 Slamet wild type 0,0 7 S0 60,4 ± 12,1 Membuka 7,0 ± 0,7 Keterangan: (*) Nomor tanaman yang ditanam untuk generasi ketiga
3
(Polymerase Chain Reaction) didesain dari cDNA heterotrimerik Gα kedelai kultivar Slamet (Accession: L27418) dan dari DNA
genom heterotrimerik Gα padi (Accession: L35844) (Tabel 2). DNA cetakan yang digunakan sebagai kontrol dan penentuan primer yang cocok adalah cDNA
hetero-trimerik Gα (SL16). Terdapat 17 kombinasi
primer yang digunakan, yaitu L1-L2, L1-L4, L1-D1, L1-D2, L3-L2, L3-L4, U1-D1, U1-D2, U3-L2, U3-L4, L2, U3/Ug-L4, U9-L2, U9-U3/Ug-L4, U9-D1, L2, dan U10-D1 (Tabel 2). PCR dilakukan dengan
mencampur 100 ng DNA SL16, 1 l buffer Taq 10x, 0,08 dNTP mix 25 mM, 0,4 l DMSO, 1 unit enzim Taq Polymerase, 10 pmol primer forward, 10 pmol primer reverse, dan ddH2O sampai volume akhir mencapai 10
l. Kondisi PCR yang digunakan untuk semua
kombinasi primer adalah denaturasi pra-PCR 94 °C 5 menit, denaturasi pada 94 °C 1 menit, penempelan primer 52 °C 30 detik, pemanjangan DNA 72 °C 1 menit 30 detik dengan pengulangan siklus sebanyak 30 kali, pemanjangan pasca PCR pada 72 °C 5 menit, dan proses pendinginan pada 15 °C selama 10 menit.
Kombinasi primer yang berhasil ditemukan pada SL16 kemudian digunakan untuk mengamplifikasi gen Gα pada DNA genom tanaman kedelai tipe liar (wild type) dan kandidat mutan dengan campuran bahan dan kondisi PCR yang sama seperti pada SL16 kecuali pada pemanjangan DNA, yaitu 72 °C selama 3 menit. Kontrol positif percobaan identifikasi DNA genom kedelai adalah primer aktin yang didesain dari kedelai (Accession: V00450) dengan primer forward
(5’ atggcagatgccgaggatat ‘γ) dan primer
reverse (5’ cagttgtgcgaccacttgca ‘γ).
HASIL
Seleksi Tanaman Kedelai Berdasarkan Persentase Stomata Menutup dan Tinggi Tanaman
Kedelai F3 diseleksi berdasarkan tinggi tanaman, yaitu di bawah tinggi rata-rata Slamet wild type yang ditanam bersamaan
dengan tanaman kandidat mutan dan persentase stomata menutup pada pukul 12.00 WIB, yaitu ≥50%. Hasil panen F3 yang telah diseleksi menghasilkan 76 nomor tanaman. Kedelai F4 yang telah diseleksi berdasarkan tinggi tanaman dan kondisi stomata menghasilkan 23 nomor tanaman. Hasil seleksi panen ketiga (F5) menghasilkan 26 nomor tanaman kandidat mutan (Lampiran 1). Suhu rata-rata saat pengambilan stomata pada tanaman generasi F5 adalah 32,09 °C dengan kelembaban rata-rata 65,15% (Lampiran 2).
Tanaman yang dipilih untuk identifikasi gen Gα adalah Slamet wild type (WT), 338/3S1.174/12/1/4 (F4), 103/3S3.103/8/5/1/4 (F5) (Giok 2010), serta satu tanaman mutan lainnya yang dipilih secara acak yaitu 344/3S1.196/14/1/8 (F4). Hasil pengamatan stomata pada ketiga tanaman tersebut adalah menutup ≥50% (Gambar β, Tabel 3, dan Lampiran 1). Tanaman kandidat mutan relatif lebih pendek dari tanaman Slamet WT (Gambar 3). Ketiga tanaman tersebut memiliki tinggi tanaman yang relatif stabil mulai dari F1 sampai F5 (Tabel 4 dan Lampiran 1).
Identifikasi Kandidat Mutan dengan PCR Hasil kuantifikasi DNA genom keempat tanaman yang terpilih untuk diidentifikasi sudah cukup murni dengan konsentrasi yang baik (Lampiran 3). Sebanyak 6 kombinasi primer berhasil mengamplifikasi bagian dari gen Gα pada cDNA SL16, yaitu L2, L1-L4, U3/UG-L2, U9-L2, U3-L1-L4, dan L3-L4 Asal Nama Primer Sekuen Basa Nitrogen (5’ - ‘γ) Posisi Nukleotida cDNA Gen Gα
Kedelai (Accession: L27418)
L27418-SOYGALP1 (L1) gcttcacacttcacacttaacact (F) 111 nukleotida sebelum ATG L27418-SOYGALP2 (L2) atattgttgtatacctgacctc (R) 114 nukleotida sesudah TGA L27418-SOYGALP3 (L3) gcgcagaatgactttgattctt (F) 309 nukleotida sesudah ATG L27418-SOYGALP4 (L4) cttaggacccactcaaaaagttcc (R) 358 nukleotida sebelum TGA
DNA Genom Gen
Gα Padi (Accession: L35844)
RGA1-D (D1) aaagatcaatcaatggtccacg (R) 138 nukleotida sesudah TGA RGA2-D (D2) ccacttgtgagagttctt (R) 1026 nukleotida sesudah ATG RGA-U1 (U1) gacgtcaacgtgcttcctgg (F) 38 nukleotida sebelum ATG RGA-U3 (U3) ctggctttgatgaggcagaac (F) 775 nukleotida sesudah ATG RGA-U3/Ug (U3/Ug) gcaacttcctgattgtgcaca (F) 1759 nukleotida sesudah ATG
RGA-U9 (U9) cagaatgatggagaccaagg (F) 689 nukleotida sebelum TGA RGA-U10 (U10) gctgcatggtgcacttat (F) 103 nukleotida sebelum TGA Keterangan: ATG = Kodon awal TGA = Kodon akhir
4
338/3S1.174/12/1/4
Slamet Wild Type
(Gambar 4 dan 5). Kombinasi primer yang berhasil mengamplifikasi gen Gα pada DNA genom Slamet WT dan DNA genom kandidat mutan hanya tiga, yaitu U9-L2, U3-L4, dan L3-L4 dengan aktin sebagai kontrol positif
(Gambar 6). Berdasarkan kemiripan gen Gα pada cDNA kedelai dengan cDNA padi, dihasilkan perkiraan peta genetik gen Gα pada DNA genom kedelai (Gambar 7).
Gambar 2 (a) Kondisi stomata membuka (tanda panah) pada Slamet tipe liar WT, (b, c, d) Kondisi stomata menutup (tanda panah) pada 103/3S3.103/8, 344/3S1.196/14/1/8,
338/3S1.174/12/1/4.
Tabel 3 Kestabilan genetik persentase stomata menutup tanaman kandidat mutan terpilih dan Slamet WT
Nomor Tanaman F3 (%) F4 (%) F5 (%) 103/3S3.103/8/5/1/4 33,33 25,62±8,30 82,95±10,43
338/3S1.174/12/1/4 75,93±1,61 41,79±6,41 * 344/3S1.196/14/1/8/4 43,62±11,26 64,58±13,32 81,23±7,60
Slamet WT 27,78±9,62 0,42±0,65 4,44±6,17 Keterangan: (*) Merupakan tanaman generasi keempat
Gambar 3 Perbandingan tinggi tanaman Slamet WT dan kandidat mutan 338/3S1.174/12/1/4 pada umur dua bulan
Tabel 4 Kestabilan genetik tinggi tanaman kandidat mutan terpilih dan Slamet tipe liar WT
Nomor Tanaman F1 (cm)* F2 (cm)* F3 (cm) F4 (cm) F5 (cm) 103/3S3.103/8/5/1/4 35,0 27,0 34,0 17,5 37,0
338/3S1.174/12/1/4 36,0 45,7 52,0 34,0 ** 344/3S1.196/14/1/8/4 30,0 52,3 34,0 29,0 64,0
Slamet WT 60,4 68,6 55,13±12,10 57,33±10,72 67,22±12,68 Keterangan: (*) Hartini (2008)
(**) Merupakan tanaman generasi keempat
Gambar 4 Panjang basa nitrogen antar primer pada cDNA heterotrimerik Gα kedelai
10 cm
a
0,05 mm
b
0,05 mm
c
0,05 mm
d
0,05 mm
1383bp
42bp L4
493bp 795bp
628bp 932bp
530bp
5
1 2 3 4 5 6 7
U3/Ug
L1 U3 L3 U9 L4 L2
ATG TGA
±1000bp ±2500bp ±2300bp 78bp 90bp 38bp 73bp
201bp 108bp 102bp 56bp 135bp 94bp 60bp 83bp 184bp 325bp
Gambar 7 Perkiraan peta genetik gen heterotrimerik Gα pada DNA genom kedelai
PEMBAHASAN
Hormon asam absisat (ABA) merupakan hormon penghambat pertumbuhan yang juga membantu tumbuhan menghadapi kondisi lingkungan yang buruk (cekaman). Sebagai contoh, ketika suatu tumbuhan mulai layu, ABA akan terakumulasi di daun dan menyebabkan stomata menutup, mengurangi transpirasi dan mencegah kehilangan air lebih banyak (Campbell et al. 2006). Nilson & Assmann (2010) menyatakan bahwa protein heterotrimerik G subunit α (GPA1) merupakan protein yang berperan dalam pengaturan efisiensi transpirasi pada Arabidopsis. Mutan Gα pada Arabidopsis hasil penelitian Perfus-Barbeoch et al. (2004) memperlihatkan peningkatan sensitivitas tumbuhan terhadap ABA. Pada kondisi mutan
Gα, ABA menginduksi stomata untuk menutup dan mencegah masuknya ion K+ pada sel penjaga stomata. Transduksi sinyal yang terganggu mengakibatkan
informasi-informasi selular tidak tersampaikan dengan baik dan pertumbuhan tumbuhan menjadi terganggu. Oleh sebab itu, stomata menutup
menjadi salah satu ciri anatomi dari mutan Gα
(Fujisawa et al. 1999).
Kondisi stomata kedelai kultivar Slamet WT telah diamati pada waktu pengamatan yang berbeda, yaitu pukul 10.00 WIB, 11.00 WIB, 12.00 WIB, dan 13.00 WIB (data tidak dipublikasi). Diantara keempat waktu pengamatan tersebut, stomata memiliki persentase membuka paling tinggi pada pukul 12.00 WIB, sehingga waktu yang paling tepat untuk pengamatan kondisi stomata adalah pada pukul 12.00 WIB.
Ciri lainnya dari tanaman mutan Gα pada tumbuhan adalah pertumbuhan fisik yang terhambat, yaitu tinggi tanaman yang kerdil (Ma 2001; Fujisawa et al. 1999). Terhambatnya transduksi sinyal pada sel tumbuhan akibat penerus informasi yang tidak aktif menyebabkan beberapa second messenger tidak tersampaikan ke dalam sel Gambar 5 PCR cDNA gen Gα (SL16). (1) marker
1kb Plus DNA Ladder, (2) primer U3/Ug-L2 (795bp), (3) primer U9-L2 (493bp), (4) primer U3-L4 (628bp), (5) primer L3-L4 (530pb) , (6) primer L1-L4 (932bp), (7) primer L1-L2 (1383bp). 1650bp 1000bp 850bp 650bp 500bp 400bp
Gambar 6 PCR DNA genom. (M) marker 1kb Plus DNA Ladder, (a) Slamet WT, (b) 103/3S3.103/8/5/1/4, (c) 344/3S1.196/14/1/8, (d) 338/3S1.174/12/1/4, (1) primer U9-L2 (±1kb), (2), primer L3-L4 (±2,3kb), (3) primer U3-L4 (±2,5kb) (4) Aktin (550bp).
4
ekson
intron M a b c d
6
sehingga aktivitas respon selular dan pembelahan sel menjadi terganggu yang mengakibatkan tanaman menjadi kerdil (Fujisawa et al. 1999). Kedua ciri mutan tersebut, yaitu stomata menutup dan ukuran tanaman yang kerdil menjadi parameter untuk
menyeleksi kandidat mutan Gα pada
penelitian ini.
Penanaman dilakukan hingga tiga kali untuk memperoleh tanaman kandidat mutan yang stabil secara genetik. Hal tersebut ditunjukkan dengan Standar Deviasi (SD) persentase stomata menutup pada generasi kelima yang lebih kecil daripada SD pada generasi keempat (Tabel 3 dan Lampiran 1), demikian halnya dengan tinggi tanaman pada ketiga tanaman kandidat mutan 103/3S3.103/8/5/1/4, 338/3S1.174/12/1/4, dan 344/3S1.196/14/1/8 yang juga sudah relatif stabil. Diantara ketiga tanaman tersebut tidak ada yang lebih tinggi dari tanaman Slamet WT yang menjadi kontrol pada penanaman yang sama (Tabel 4 dan Lampiran 1).
Sekuen DNA genom gen Gα kedelai belum diketahui sehingga primer spesifik untuk mengamplifikasi gen tersebut juga belum diketahui dengan jelas. Oleh karena itu perlu dilakukan pengujian untuk mendapatkan kombinasi primer yang tepat dengan menggunakan cDNA gen Gα SL16 sebagai DNA cetakan. Sebanyak 17 kombinasi primer yang didesain dari kedelai dan padi digunakan, namun hanya 6 kombinasi primer yang berhasil diamplifikasi melalui PCR (Gambar 5). Terdapat tujuh primer yang tepat berada pada ekson gen Gα sehingga ketujuh primer tersebut dapat digunakan untuk mengamplifikasi gen Gα dari DNA genom tanaman kedelai (Gambar 4).
Enam kombinasi primer yang digunakan untuk mengamplifikasi bagian DNA genom
Gα, tiga diantaranya gagal, yaitu primer
L1-L2, L1-L4, dan U3/Ug-L2. Gagalnya ketiga kombinasi primer tersebut kemungkinan disebabkan oleh sekuen basa yang diamplifikasi terlalu panjang, yaitu >795 bp dan belum termasuk intron (Gambar 7). Enzim Taq Polymerase dengan nama dagang Fermentas dapat mengamplifikasi segmen DNA sampai 5 kb, namun seringkali terjadi kesalahan penempelan primer dan reaksi yang tidak kompeten yang menyebabkan PCR tidak berhasil. Enzim Taq akan bekerja dengan optimal pada panjang basa di bawah 2 kb (Brown 1991).
Tiga kombinasi primer yang berhasil mengamplifikasi gen Gα adalah primer U9-L2, U3-L4, dan L3-L4. Produk PCR untuk
primer U9-L2 dan L3-L4 muncul pada keempat tanaman yang diamati. Sedangkan primer U3-L4 hanya berhasil pada tanaman Slamet WT dan 344/3S1.196/14/1/8 sedangkan pada kandidat mutan 103/3S3. 103/8/5/1/4 dan 338/3S1.174/12/1/4 tidak ada pita. Hal ini menunjukkan bahwa pada kedua tanaman tersebut kemungkinan telah terjadi mutasi gen Gα, tepatnya pada daerah penyandi primer U3 dan L4.
Daerah penyandi primer U3 terletak pada ekson 4 dan primer L4 terletak pada ekson 10 (Gambar 7). Ekson 4 dan ekson 10 mengapit daerah penyandi situs pengikat GTP, yaitu G2 dan G3 serta daerah penyandi pengikat adenyl cyclase (Gambar 1). Adanya mutasi gen Gα pada daerah yang diapit oleh ekson 4 dan ekson 10 dapat mengganggu fungsi protein
Gα dalam mengikat GTP serta mengikat adenyl cyclase sehingga pertumbuhan tanaman mutan, yaitu 103/3S3.103/8/5/1/4 dan 338/3S1.174/12/1/4 menjadi terganggu yang ditunjukkan dengan kondisi stomata yang menutup pada siang hari dan tinggi tanaman yang kerdil.
Pita yang dihasilkan dengan primer U3-L4 tidak setebal pita dengan primer yang lain. Ukuran basa nitrogen yang disandikan oleh kedua primer tersebut terlalu besar, yaitu ± 2,5 kb. Selain itu, kecocokan sekuen primer U3 yang didesain dari padi dengan sekuen cDNA gen Gα kedelai hanya 85% sedangkan primer U9 mencapai 95% dan primer L2 dan L4 mencapai 100%. Kedua faktor tersebut bersamaan menjadikan PCR dengan primer U3-L4 kurang efektif sehingga menghasilkan pita yang tipis.
Tiga dari enam kandidat mutan yang diamati berhasil diidentifikasi secara molekular, yaitu 103/3S3.103/8/5/1/4, 338/3S1.174/12/1/4, dan 344/3S1.196/14/1/8. Sedangkan tiga kandidat mutan lainnya, yaitu 86/3S3.28/6, 86/3S3.28/3, dan 102/3S3.82/21 belum dapat dipastikan letak mutasinya secara molekular.
SIMPULAN
7
SARAN
Perlu dilakukan sekuensing serta desain primer yang baru untuk memperjelas letak mutasi pada daerah penyandi primer U3 dan L4. Selain itu, perlu dilakukan penelitian lebih lanjut untuk menyeleksi dan mengidentifikasi tanaman kandidat mutan Gα yang lain, serta mengidentifikasi tanaman kandidat mutan melalui mRNA kedelai.
DAFTAR PUSTAKA
Assmann SM. 1996. Guard cell G proteins. Tr Plan Sci 1:73-74.
Becker WM, Kleinsmith LJ, Hardin J. 2000. The World of The Cell. 4th Ed. Madison : The Benjamin/Cummings Publishing Company.
Belafiff MB. 2007. Analisis keterlibatan
protein heterotrimerik G subunit α
terhadap cekaman Al3+ melalui identifikasi produksi H2O2 pada kedelai kultivar Slamet [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Brown TA. 1991. Essential Molecular Biology: A Practical Approach. New York: Oxford. hlm 202.
Campbell NA et al. 2006. Biology Concepts and Connections fifth edition. New York: Benjamin Cummings. hlm 84, 664. Fujisawa Y et al. 1999. Suppression of the
heterotrimeric G protein causes abnormal morphology, including dwarfism, in rice. Proc Natl Acad Sci USA 96:7575-7580. Giok G. 2010. Studi anatomi dan histokimia
pada akar kedelai (Glycine max (L.) Merr)
kandidat mutan protein Gα terhadap
cekaman aluminium [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Hartini S. 2008. Induksi mutasi dengan irradiasi sinar gamma pada kedelai (Glycine max (L.) Merrill) kultivar Slamet dan Lumut [tesis]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor. Ma H. 1994. GTP-binding proteins in plants:
New members of an old family. Plant Mol Biol 26:1611-1636.
Ma H. 2001. Plant G proteins: The different faces of GPA1. Curr Biol 11:R869-R871. Mashuda. 2006. Ekspresi gen Gα dan GST
pada kedelai kultivar Slamet yang mendapat cekaman aluminium [tesis].
Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Matsumoto H. 2000. Cell biology of aluminum toxicity and tolerance in higher plants. Int Mol Rev of Cytol 200:1-17. Nilson SE, Assmann SM. 2010. The α
-Subunit of the Arabidopsis Heterotrimeric G Protein, GPA1, Is a Regulator of Transpiration Efficiency. Plant Physiol 152:2067-2077.
Perfus-Barbeoch L, Jones AM, Assmann SM. 2004. Plant heterotrimeric G protein function: Insights from Arabidopsis and rice mutants. Plant Biol 7:719-731. Suharsono UW. 2004. Analisis gen penyandi
protein heterotrimerik G subunit α yang
terlibat dalam sistem toleransi tanaman kedelai terhadap cekaman aluminium. Laporan Penelitian Hibah Bersaing tahun ke-I. Bogor: Institut Pertanian Bogor. Suharsono UW, Suharsono S. 2006. Analisis
gen penyandi protein heterotrimerik G
subunit α yang terlibat dalam sistem
toleransi tanaman kedelai terhadap cekaman aluminium. Laporan Akhir Penelitian Hibah Bersaing XII. Bogor: Institut Pertanian Bogor.
8
9
Lampiran 1 Hasil seleksi tanaman kandidat mutan berdasarkan persentase stomata menutup dan tinggi tanaman generasi ketiga sampai generasi kelima
Nomor Tanaman Stomata Menutup (%) Tinggi Tanaman (cm) Umur Berbunga (HST) Jumlah Biji
F3 F4 F5 F3 F4 F5 F3 F4 F5 F3 F4 F5
43/1S3.101/13/2/4/7 52,19±13,02 72,50±8,99 70,05±6,12 29 29 45 56 38 39 13 117 76
102/3S3.82/21/20/2/5 59,26±8,49 28,82±9,74 59,07±6,16 28,5 23 34 63 41 47 8 52 15
103/3S3.103/8/5/1/4 - 25,62±8,30 82,95±10,43 34 17,5 55 41 41 39 138 80 454
338/3S1.174/12/1/4/6 75,93±1,61 41,79±6,41 44,30±9,88 52 34 68,5 34 38 38 162 102 267
341/3S1.179/2/1/1/6 59,49±6,22 25,56±0,96 56,87±6,79 25 17,5 51 41 43 37 21 7 0
341/3S1.179/2/1/3/4 59,49±6,22 20,82±15,63 73,71±11,81 25 34 57 41 38 33 21 136 47
341/3S1.179/2/1/4/3 59,49±6,22 21,30±8,37 60,95±11,81 25 34,5 60 41 38 32 21 95 134
341/3S1.179/2/1/4/5 59,49±6,22 21,30±8,37 57,17±8,00 25 34,5 48 41 38 35 21 95 51
341/3S1.179/2/1/4/6 59,49±6,22 21,30±8,37 85,90±5,46 25 34,5 62 41 38 35 21 95 233
341/3S1.179/2/1/4/8 59,49±6,22 21,30±8,37 65,00±16,05 25 34,5 53 41 38 37 21 95 106
341/3S1.179/2/1/7/1 59,49±6,22 67,25±11,13 74,66±21,13 25 28 26,5 41 38 41 21 82 0
341/3S1.179/2/1/7/2 59,49±6,22 67,25±11,13 76,09±14,60 25 28 25 41 38 41 21 82 58
341/3S1.179/2/1/7/4 59,49±6,22 67,25±11,13 69,62±11,67 25 28 49 41 38 36 21 82 120
341/3S1.179/2/1/7/8 59,49±6,22 67,25±11,13 86,85±12,25 25 28 42 41 38 36 21 82 89
341/3S1.179/2/3/4/1 61,60±7,18 20,34±13,57 76,58±11,14 56 27 47 56 38 36 83 108 134
341/3S1.179/2/3/4/3 61,60±7,18 20,34±13,57 68,86±24,32 56 27 44 56 38 39 83 108 55
344/3S1.196/14/1/2/4 43,62±11,26 52,10±32,75 62,60±4,67 34 38 52 34 41 41 153 155 121
344/3S1.196/14/1/2/7 43,62±11,26 52,10±32,75 58,74±10,70 34 38 62 34 41 41 153 155 267
344/3S1.196/14/1/4/3 43,62±11,26 26,40±25,49 51,72±5,77 34 32,5 54 34 41 41 153 126 140
344/3S1.196/14/1/4/5 43,62±11,26 26,40±25,49 57,94±13,59 34 32,5 54 34 41 41 153 126 283
344/3S1.196/14/1/4/6 43,62±11,26 26,40±25,49 50,48±11,19 34 32,5 59 34 41 43 153 126 117
344/3S1.196/14/1/4/7 43,62±11,26 26,40±25,49 72,31±6,96 34 32,5 57 34 41 43 153 126 99
344/3S1.196/14/1/8/4 43,62±11,26 64,58±13,32 81,23±7,60 34 29 64 34 41 41 153 124 77
344/3S1.196/14/1/8/5 43,62±11,26 64,58±13,32 83,07±8,25 34 29 70 34 41 45 153 124 166
344/3S1.196/14/1/8/7 43,62±11,26 64,58±13,32 54,98±3,62 34 29 50 34 41 39 153 124 1
344/3S1.196/14/10/4/4 59,63±7,19 28,53±12,96 74,56±7,21 56 31 32,5 56 43 42 32 54 170
344/3S1.196/14/10/6/5 59,63±7,19 87,48±3,21 63,15±15,65 56 38 60 56 41 41 32 132 78
10
Lampiran 2 Kondisi lingkungan saat pengambilan stomata pada F5
Tanggal Pengamatan Suhu (°C)
Kelembaban (% RH)
Intensitas Cahaya (lux)
29 Oktober 2009 31,3 74,8 1343x10 30 Oktober 2009 31,4 63,8 1623x10 31 Oktober 2009 32,8 63,6 1482x10 02 Nopember 2009 33,0 58,2 1402x10 04 Nopember 2009 35,9 51,2 1520x10 10 Nopember 2009 33,6 54,7 1546x10 12 Nopember 2009 32,4 71,7 1392x10 16 Nopember 2009 29,4 72,6 1302x10 17 Nopember 2009 30,3 71,4 1447x10 18 Nopember 2009 30,8 69,5 1700x10 Rata-rata 32,09 65,15 1475,7x10
Lampiran 3 Hasil kuantifikasi DNA genom