II. TINJAUAN PUSTAKA
2.1.
Karakteristik KHVHerpesvirus adalah virus yang berukuran besar. Herpetos berasal dari bahasa Yunani yang artinya mengerikan. Herpesviridae berbiak dalam inti, membentuk badan inklusi yang disebut “cowdry type A”. Virus-virus ini memperoleh amplopnya sewaktu budding melalui membran inti sel (Malole, 1988)
Herpesvirus memiliki sejumlah besar gen, yang telah dibuktikan bersifat peka terhadap kemoterapi anti virus (Brooks, et al, 1995). Menurut Malole (1988), semua anggota Herpesviridae sensitif terhadap ether dan asam. DNA-nya berserabut ganda dengan berat molekul 50–100 x 106 Dalton. Kapsidnya bersimetri kubus memiliki 162 kapsomer (150 heksagonal dan 12 pentagonal). Virion yang beramplop berukuran antara 150–200 nm, tetapi virion yang tanpa amplop juga sering ditemukan dengan ukuran 100–110 nm. Bentuk famili Herpesviridae dapat dilihat pada Gambar 1.
KHV yang termasuk salah satu anggota famili Herpesviridae, dilaporkan menyerang ikan mas dan koi (Cyprinus carpio) di banyak negara dan telah menimbulkan kerugian ekonomi yang cukup besar (Perelberg et al., 2003).
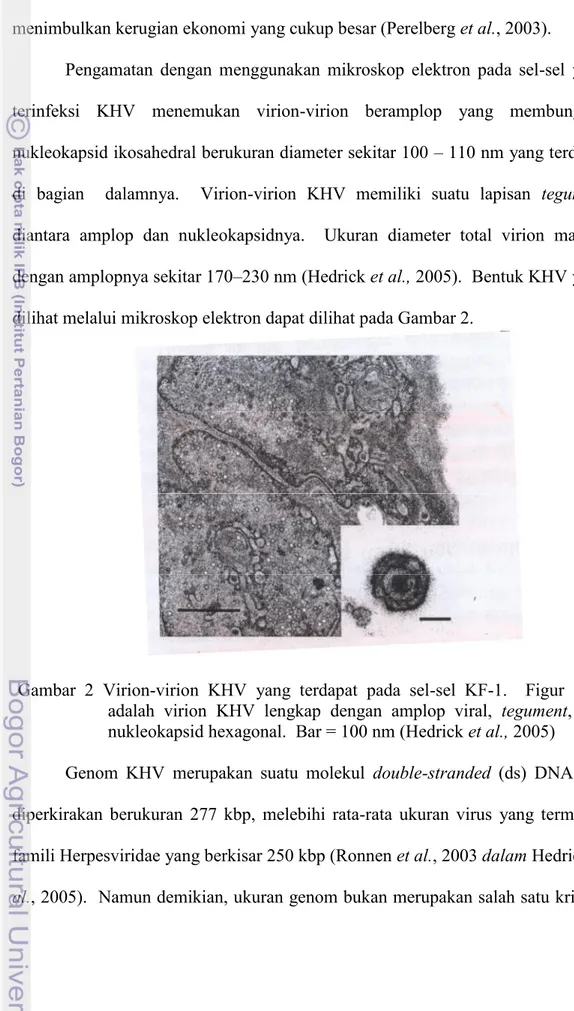
Pengamatan dengan menggunakan mikroskop elektron pada sel-sel yang terinfeksi KHV menemukan virion-virion beramplop yang membungkus nukleokapsid ikosahedral berukuran diameter sekitar 100 – 110 nm yang terdapat di bagian dalamnya. Virion-virion KHV memiliki suatu lapisan tegument diantara amplop dan nukleokapsidnya. Ukuran diameter total virion matang dengan amplopnya sekitar 170–230 nm (Hedrick et al., 2005). Bentuk KHV yang dilihat melalui mikroskop elektron dapat dilihat pada Gambar 2.
Gambar 2 Virion-virion KHV yang terdapat pada sel-sel KF-1. Figur inset adalah virion KHV lengkap dengan amplop viral, tegument, dan nukleokapsid hexagonal. Bar = 100 nm (Hedrick et al., 2005)
Genom KHV merupakan suatu molekul double-stranded (ds) DNA dan diperkirakan berukuran 277 kbp, melebihi rata-rata ukuran virus yang termasuk famili Herpesviridae yang berkisar 250 kbp (Ronnen et al., 2003 dalam Hedrick et al., 2005). Namun demikian, ukuran genom bukan merupakan salah satu kriteria
yang dipertimbangkan dalam mengelompokkan virus ini dalam famili Herpesviridae.
KHV berbeda dari herpesvirus-herpesvirus lainnya yang menyerang ikan-ikan cyprinid seperti Cyprinid herpesvirus 1 yang merupakan agen penyebab carp pox. Hal ini ditunjukkan melalui uji-uji immunofluosescene yang menemukan antibodi anti-CyHV-1 gagal bereaksi dengan KHV (Hedrick et al., 2000). Selanjutnya, perbedaan-perbedaan dalam susunan protein dan sequence genomik virion memberikan bukti tambahan bahwa kedua jenis virus tersebut merupakan agen-agen yang berbeda (Gilad et al., 2002). CyHV-1 dapat menyebabkan mortalitas pada ikan mas dan koi tetapi hanya terjadi pada ikan-ikan yang usianya kurang dari 2 bulan (Sano et al., 1985 dalam Hedrick et al., 2005). Selain itu juga, ikan-ikan yang dapat bertahan hidup dari infeksi CyHV-1 menunjukkan karakteristik pertumbuhan papillomatous-like yang umum diketahui sebagai carp pox (Schubert, 1966 dalam Hedrick et al., 2005). Satu jenis herpesvirus lainnya adalah Cyprinid herpesvirus 2 yang awalnya dinamakan goldfish hematopoietic necrosis virus (GFHNV), telah diobservasi melalui mikroskop elektron dan telah berhasil diisolasi dari ikan koki (Carassius auratus) yang menunjukkan nekrosis yang parah pada sel-sel hematopoietic. Tidak seperti CyHV-1, KHV sangat virulen dan dapat menyebabkan mortalitas pada seluruh ukuran ikan mas dan koi (Hedrick et al., 2000 dan Perelberg et al., 2003). CyHV-1 hanya dapat menyebabkan kematian pada Ikan Mas dan Koi yang berumur kurang dari 2 bulan (Sano et al., 1985 dalam Hedrick et al., 2005). Selain itu, bukti lainnya adalah tidak terjadi pembentukan papilloma pada ikan-ikan yang dapat bertahan hidup dari infeksi KHV.
Perbandingan-perbandingan DNA genomik dan polipeptida virion dari KHV terhadap CyHV-1 menunjukkan bahwa virus-virus tersebut memiliki kemiripan tetapi merupakan agen-agen yang berbeda nyata. Perbedaan ini dapat ditunjukkan melalui deteksi terhadap masing-masing virus dengan menggunakan uji-uji PCR yang dikembangkan oleh beberapa peneliti (Gray et al., 2002; Bercovier et al., 2005). Gray et al. (2002), telah mengembangkan uji PCR melalui pembuatan disain primer untuk mendeteksi KHV. Primer set SphI-5, forward
(5’-GACACCACATCTGCAAGGAG-3’) dan reverse
(5’-GACACATGTTACAATGGTGGC-3’), untuk mengamplifikasi produk dengan ukuran fragmen DNA 290 bp, terbukti dapat mendeteksi KHV. Hal ini ditunjukkan dengan munculnya pita DNA yang tampak jelas pada ukuran fragmen 290 bp. Sedangkan pada isolat-isolat virus lainnya seperti Channel Catfish Virus (CCV) dan Cyprinid Herpesvirus (CHV), pita-pita DNA tersebut tidak tampak (Gambar 3). Uji PCR ini cukup sensitif untuk mendeteksi 100 femtograms atau sekitar 600 kopi DNA dari DNA genomik KHV (Gray et al., 2002).
Gambar 3 PCR primer set yang didisain Gray et al. (2002) spesifik untuk mendeteksi KHV, hal ini tampak dari pita DNA pada ukuran fragmen 290bp.
2.2. Variasi Genetik
Variasi genetik merupakan ciri-ciri yang paling esensial pada seluruh organisme hidup, yang merupakan salah satu cara untuk beradaptasi secara
progresif terhadap perubahan lingkungan alamiah (Walker, 2000). Mutasi dan rekombinasi menyebabkan adanya variasi genetika (Trun dan Trempy, 2004).
Menurut Trun dan Trempy (2004), mutasi adalah suatu perubahan fisikal pada satu atau lebih dari satu pasang nukleotida dalam DNA, dan dapat hanya mempengaruhi satu pasang nukleotida atau dapat mempengaruhi ratusan kilo basa nukleotida. Pengaruh mutasi tergantung pada tempat dimana mutasi tersebut terjadi dalam DNA. Mutasi dapat diklasifikasikan dalam dua kelompok, yaitu yang terjadi pada nukleotida tunggal dan banyak nukleotida. Mutasi yang mempengaruhi nukleotida tunggal disebut microlesions, sedangkan yang mempengaruhi banyak nukleotida disebut sebagai macrolesions.
Beberapa tipe microlesions adalah mutasi titik dan mutasi frameshift. Mutasi titik adalah perubahan yang terjadi pada satu pasang basa nukleotida. Perubahan tersebut dapat berupa substitusi basa dari satu purin dengan satu purin (A menjadi G atau G menjadi A), atau satu pirimidin dengan satu pirimidin (T menjadi C atau C menjadi T). Mutasi titik ini disebut juga transisi. Jika mutasi titik tersebut berupa substitusi basa dari satu purin dengan satu pirimidin atau satu pirimidin dengan satu purin, mutasi ini disebut juga suatu transverse. Tipe microlesions lainnya adalah mutasi frameshift, yang berupa insersi/penyisipan atau delesi/penghapusan satu pasang basa tunggal dalam suatu gen. Beberapa mutasi frameshift juga dapat diklasifikasikan sebagai macrolesions, jika mutasi tersebut berupa insersi/penyisipan atau delesi/penghapusan yang terjadi pada basa dalam jumlah banyak. Tipe mutasi macrolesions termasuk didalamnya meliputi delesi/penghapusan, duplikasi, insersi/penyisipan, dan penyusunan kembali
seperti inverse/pembalikan dan translokasi. Seluruh mutasi tersebut melibatkan perubahan-perubahan besar dalam urutan nukleotida (Trun dan Trempy, 2004)..
Variasi genetik juga dapat terjadi pada virus. Variasi viral tersebut dapat terjadi melalui sejumlah mekanisme yang meliputi penyusunan/pengaturan kembali yang utama pada struktur genom, dan pengorganisasiannya dapat terjadi melalui rekombinasi genetik. Selain itu, dapat juga melalui duplikasi gen, pertukaran gen, penghapusan gen, dan penyisipan gen. Namun demikian, bentuk variasi yang paling umum adalah mutasi melalui substitusi nukleotida (Walker, 2000).
Variasi genetika yang terjadi dapat disebabkan virus-virus tersebut harus menghadapi perubahan lingkungan yang terjadi secara terus menerus. Seperti halnya virus-virus melintas dari satu inang ke inang lainnya, mereka harus berhadapan dengan respon pertahanan dan sistem imunologis inang. Penghindaran dari pertahanan inang merupakan suatu ciri pokok strategi bertahan pada seluruh virus (Walker, 2000).
Terkait dengan KHV, sejak pertama kali terjadinya wabah, perkembangan KHV telah dilaporkan melalui berbagai penelitian maupun forum ilmiah. Berdasarkan pada penelitian – penelitian tersebut diketahui bahwa isolat–isolat KHV dari berbagai Negara seperti USA, Israel, Indonesia, Malaysia, Thailand, dan Negara–Negara Eropa memiliki perbedaan (Sano et al., 2007). Penelitian Stone et al. (2007) melaporkan berbagai varian KHV yang terdeteksi di Eropa. Selanjutnya, penelitian Aoki et al. (2007) menemukan adanya variasi genetik 3 isolat KHV yang berasal dari Jepang, Amerika Serikat, dan Israel. Ukuran genom KHV yang teridentifikasi untuk masing-masing varian adalah 295,271 bp (varian
1 ), 295,146 bp (varian 2) dan 295,138 (varian 3). Berdasarkan pada preparasi DNA melalui Restriction Endonuclease Digestion dengan Notl atau XbaI menghasilkan profil yang identik dari ketiga strain tersebut. Genom strain KHV memiliki pengulangan langsung (direct repeat) sebesar 22 kbp pada tiap–tiap terminal (22,437 bp untuk varian 1, 22,469 bp untuk varian 2 dan 22,485 bp untuk varian 3). Genom-genom varian tersebut memiliki tingkat kesamaan yang cukup tinggi pada level sekuen. Sebagai contoh, substitusi nukleotida tunggal (tidak termasuk duplikat terminal pengulangan/terminal repeat) varian 1 berbeda dengan varian 2 dan 3 pada loki 181 dari 217 loki. Hal tersebut berarti terdapat perbedaan pada setiap rata–rata 1,5 kbp. Disisi lain, dari sebanyak 36 nonconserved loki, varian 3 berbeda dengan varian 1 dan 2 pada 32 loki dan varian 2 berbeda dengan varian 1 dan 3 pada 4 loki. Selain tinjauan pada genom dan loki tersebut, ketiga strain KHV menunjukkan perbedaan pada open reading frame atau dikenal sebagai ORF. Kejadian tersebut diduga karena adanya insersi dan delesi yang terjadi pada satu atau dua strain menyebabkan kerusakan pada titik pengkodean (coding region).
Variasi genetik KHV dari beberapa isolat di Eropa, Israel dan Amerika jauh sebelumnya pada tahun 2003 telah diteliti oleh Gilad et al. (2003). Pada penelitian tersebut diketahui bahwa sebanyak 6 dari 7 isolat yang diteliti tidak memiliki perbedaan yang signifikan berkaitan dengan polipeptida virionnya. Pada isolat KHV D-081 dari Israel diketahui memiliki polipeptida tambahan dengan ukuran 162 dan 41 kDa. Penelitian tersebut juga menemukan adanya perbedaan antara isolat KHV yang berasal dari Israel dengan isolat Amerika. Isolat-isolat selanjutnya yang ditemukan di Israel maupun Amerika dengan lokasi geografis
yang berbeda-beda memiliki polipeptida virion dan RFLP yang identik atau mirip dengan isolat KHV yang terlebih dulu diisolasi (Gilad et al., 2002). Sejalan dengan hasil penemuan tersebut, Banks (1993) mengemukakan bahwa meskipun terdapat variasi minor yang terlihat pada isolat virus dalam satu spesies, namun isolat yang berasal dari lokasi geografis yang sejenis akan membentuk kelompok yang bersifat relative homogeny. Penelitian yang mendalam berkaitan dengan sequencing amplikon KHV dan jumlah variasi lokasi geografis asal genom di masa mendatang akan sangat berguna dalam membedakan beberapa isolat KHV berdasarkan lokasi geografis.
2.3. Inang
Ikan mas dan koki (Cyprinus carpio) merupakan inang KHV (Perelberg et al., 2003; Hedrick et al., 2005; Ishioka et al., 2005; Shapira et al., 2005; Waltzek et al., 2005). KHV ini diketahui dapat menyerang seluruh ukuran ikan. Meskipun demikian, ikan-ikan berukuran kecil lebih sensitif terhadap KHV daripada yang berukuran lebih besar ( Perelberg et al., 2003).
Kerentanan ikan terhadap infeksi KHV dan cara transmisinya dapat berpatokan pada hasil penelitian Perelberg et al. (2003) dengan menggunakan berbagai jenis ikan cyprinids yaitu Cyprinus carpio, Oreochromis niloticus, Bidyanus bidyanus, Hypophthalmichthys molitrix, Carassius auratus, Ctenopharyngodon idella. Hasil penelitiannya menunjukkan hanya Cyprinus carpio saja yang rentan terhadap infeksi KHV, dengan tingkat kematian mencapai 72 % setelah terpapar virus, sedangkan jenis-jenis ikan lainnya tidak terpengaruh dan tetap bertahan hidup. Melalui uji kohabitasi yang dilakukan Perelberg et al. (2003) menunjukkan hanya ikan mas saja yang dapat
mentransmisikan KHV ke ikan mas lainnya yang sehat. Sedangkan ikan-ikan dari strain-strain yang resisten tetap bertahan hidup dan tidak menularkan KHV pada ikan mas yang sehat.
2.4. Mekanisme dan Perubahan Patologis Infeksi KHV
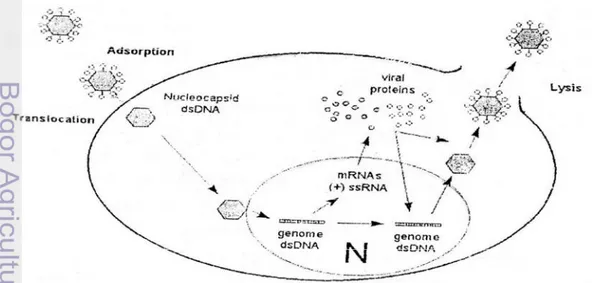
Proses infeksi herpesvirus pada sel inang dimulai dengan terjadinya perlekatan atau adsorpsi partikel virus pada reseptor yang ada di permukaan sel inang. Adsorpsi virus pada permukaan sel segera diikuti oleh masuknya virus-virus yang mengandung genom ds DNA ke dalam sitoplasma melalui proses endocytosis. Selanjutnya nucleocapsid ditransportasikan sepanjang matriks
cytoskeletal menuju membran inti kemudian masuk ke dalam inti/nukleus. Setelah memasuki inti, terjadi proses replikasi virus dengan langkah-langkah biosintesisnya menurut urutan sebagai berikut: 1) Transkripsi untuk pembuatan messenger RNA (mRNA) dari DNA virus asal (parent) yang menginfeksi sel (sesudah uncoating). 2) mRNA tersebut berpindah ke ribosom dalam sitoplasma sel dan diterjemahkan (translated) menjadi enzim dan protein-protein lainnya (early protein = protein awal) yang melakukan sintesis asam nukleat untuk virus baru. 3) Replikasi DNA virusdalam inti.4) Transkripsi lanjutan untuk pembuatan mRNA lagi dari DNA-parent dan virus baru (progeny). 5) Penerjemahan (translation) mRNA yang dibentuk kemudian (late mRNA) menjadi protein (late protein) sebagai bagian dari komponen virus dan sebagai enzim yang sama dengan early enzyme. 6) Perakitan (assembly) virus baru (progeny virus) di dalam
inti sel. 7) Pelepasan virus yang matang (mature virus) dari sel. Herpesvirus selain keluar secara biasa melalui sitoplasma dimana virus-virus ini memperoleh
amplop, dapat juga berpindah langsung ke sel terdekat tanpa harus terlebih dahulu keluar sel yang terinfeksi.
Metode transfer antar sel tersebut memungkinkan virus menyebar dalam tubuh inang walaupun terdapat banyak antibodi di dalam cairan tubuh di luar sel. Hal inilah yang menyebabkan terjadinya infeksi virus secara laten atau kronis selama berbulan-bulan sampai bertahun-tahun pada inang yang terlihat sehat (Malole, 1988; Walker, 2000). Strategi dasar replikasi virus dengan genome ds DNA dapat dilihat pada Gambar 4. Terkait dengan KHV, Hedrick et al. (2000) dan Perelberg et al. (2003) berdasarkan hasil penelitiannya menyimpulkan bahwa KHV pertama kali masuk dan menginfeksi ikan melalui insang dan atau usus. Mekanisme infeksi KHV menurut laporan Pikarsky et al. (2004) menyebutkan bahwa virus pertama kali masuk ke dalam tubuh ikan melalui insang, selanjutnya bereplikasi di dalam insang. Aktivitas replikasi tersebut mempengaruhi struktur insang sehingga terlihat mengalami nekrosis dan kelukaan pada lapisan mukosanya. Kerusakan insang yang parah merupakan salah satu faktor munculnya gejala klinis pada ikan.
Berdasarkan hasil penelitiannya, Gray et al. (2002) melaporkan bahwa KHV menyebar secara sistemik pada ikan yang terinfeksi. Hal tersebut dibuktikan melalui analisis PCR dan DNA hybridization, yang mendeteksi DNA KHV pada jaringan insang, gastrointestinal, dan hati ikan yang terinfeksi. Pada jaringan otak, DNA KHV terdeteksi lemah.
Studi yang dilakukan beberapa peneliti dengan menggunakan pengujian patologik mikroskopik dan uji-uji PCR kuantitatif juga menunjukkan jaringan-jaringan target KHV meliputi insang, ginjal, limpa, kulit, otak, usus, dan hati (Hedrick et al., 2000; Gray et al., 2002; Gilad et al., 2003; Gilad et al., 2004). Hasil penelitian Gilad et al. (2004) menemukan konsentrasi DNA KHV tertinggi terdapat pada insang, ginjal, limpa, dengan jumlah genom yang ekuivalen secara konsisten yaitu mulai dari 108 sampai 109 setiap 106 sel-sel inang. Level DNA KHV yang tinggi juga ditemukan pada mucus, hati, usus, dan otak. Ikan koi yang dapat bertahan hidup dari infeksi KHV pada 62 – 64 hari setelah terpapar virus, masih mengandung kopi genom KHV dalam jumlah yang lebih rendah (sampai dengan 1,99 x 102per 106sel-sel inang) pada insang, ginjal, atau otak.
Gejala klinis ikan yang terinfeksi KHV sangat variatif dan umumnya tidak spesifik. Gejala-gejala yang ditemukan antara lain adalah ikan berenang ke permukaan untuk mengambil udara atau ikan mengumpul di tempat-tempat air masuk. Ikan kelihatan megap-megap karena frekuensi pernafasannya tinggi. Selain itu, seringkali ditemukan juga ikan bergerak kehilangan arah dan berenang dengan gerakan yang tidak teratur, sebelum akhirnya mengalami kematian (Gray et al., 2002). Kematian ikan berlangsung sangat cepat, sekitar 24 – 48 jam setelah
Hasil pengamatan terhadap ikan yang terserang KHV secara umum menunjukkan tanda-tanda produksi lendir (mucus) berlebih sebagai respon fisiologis terhadap kehadiran patogen, selanjutnya produksi lendir menurun drastis sehingga tubuh ikan terasa kasat. Pada tahap awal infeksi, insang ikan menunjukkan bercak-bercak putih kecil di bagian ujung-ujung lembaran insang dan warna insang masih terlihat normal dan cerah. Infeksi lebih lanjut ditandai dengan warna ujung-ujung lembaran insang menjadi pucat putih keabu-abuan disamping bercak-bercak putih menjadi lebih jelas dan meluas. Perkembangan infeksi selanjutnya menunjukkan sebagian besar lembaran-lembaran insang mengalami nekrosis atau kematian sel-sel insang. Secara keseluruhan insang mengalami kerusakan, terjadi penempelan diantara lembaran-lembaran insang, geripis, dan akhirnya membusuk. Pendarahan (hemorrhage) juga terjadi di sekitar pangkal dan ujung sirip serta permukaan tubuh lainnya, bahkan selanjutnya sirip menjadi rapuh dan geripis. Sering juga ditemukan adanya kulit yang melepuh, atau bahkan luka yang diikuti dengan infeksi sekunder oleh bakteri, jamur, dan parasit (Perelberg et al., 2003; Taukhid et al., 2004; Hedrick et al., 2005). Gejala klinis yang tampak pada ikan yang terinfeksi KHV dapat dilihat pada Gambar 5.
Ikan yang terserang penyakit KHV juga menunjukkan perubahan pada organ-organ internalnya. Hati (liver) terlihat membengkak, terdapat bercak – bercak putih yang sebenarnya adalah nekrosis, tekstur lembek, pucat, terdapat petechiae, selanjutnya mengalami kerusakan. Ginjal membengkak dan terlihat berwarna pucat. Studi yang dilakukan beberapa peneliti menunjukkan bahwa ikan yang terinfeksi KHV mengalami disfungsi hati dan sistem osmoregulasi,
hypoprotein, serta imunosupresif sehingga rentan terhadap infeksi patogen sekunder (Hedrick et al., 2000; Perelberg et al., 2003; Taukhid et al., 2004).
Secara histologis, ikan-ikan yang terserang KHV menunjukkan adanya kerusakan jaringan atau lesi yang serius terutama pada kulit, insang, dan organ dalamnya (hati, ginjal, limpa dan sistem pencernaan). Pada jaringan insang terjadi hyperplasia dan hypertrophy terutama pada sel-sel epitel lamella sekunder sehingga terjadi fusi antar lamella sekunder yang berdekatan (Gambar 6). Hal tersebut terjadi karena adanya proliferasi dan pembengkakan sel-sel epitel lamella sekunder yang tidak terkontrol akibat induksi virus-virus yang menginfeksi (Perelberg et al., 2003).
Selanjutnya, kerusakan atau perubahan-perubahan histologis tersebut antara lain dapat dilihat dengan ditemukannya semacam eosinophilic intracytoplasmic inclusion body (EICB-like) dan nekrosis serta intranuclear inclusion bodies pada sel-sel epithelium jaringan insang (epithelium branchial). Perubahan berikutnya adalah ditemukannya koloni sel-sel bakteri yang terdapat di dalam suatu ruangan yang terbentuk akibat adanya fusi antar lamela sekunder yang berdekatan (Perelberg et al., 2003; Taukhid et al., 2004.
Gambar 5 Gejala klinis ikan yang terserang KHV, tampak pendarahan dan luka pada permukaan tubuh, sirip geripis, insang busuk dan mengalami erosi (sumber: BRKP, 2004)
Organ limpa (spleen) mengalami nekrosis di beberapa lokasi pada sel-sel atau jaringan parensimnya. Pada sel-sel parensim limpa ada sebagian inti selnya yang mengalami pembengkakan (hipertrofi) dan mengakibatkan terjadinya marjinalisasi kromatin (Gilad et al., 2002).
Gambar 6 Jaringan insang yang terinfeksi KHV, menunjukan hiperplasia dan fusi Lamela sekunder (Sumber: Perelberg et al., 2003)
Perubahan histologis dari organ ginjal ditandai oleh perubahan-perubahan yang terjadi antara lain pada sel-sel hematopoietik di jaringan interstitial pada bagian anterior ginjal yang mengalami nekrosis dan di dalam inti selnya terdapat badan inklusi (Hedrick et al., 2000; Perelberg et al., 2003).
Hedrick et al. (2000), telah melakukan penelitian untuk mengetahui efek virus terhadap sel. Dalam penelitian tersebut digunakan beberapa jenis cell lines yaitu koi fin-1 (KF-1), epithelioma papulosum cyprini (EPC), dan fathead minnow (FHM) dari Prenephales promelas. Virus yang digunakan berasal dari hasil ekstraksi organ-organ ginjal, limpa (spleen), dan insang, yang berasal dari ikan yang secara klinis terinfeksi. Ekstrak tersebut selanjutnya diinokulasikan pada KF-1, EPC, FHM. Hasil pengamatan setelah 1 – 2 minggu pasca inokulasi menemukan adanya aktivitas atau efek virus pada cell lines atau cytopathic
effects (CPE) pada KF-1 dan EPC. Efek tersebut meliputi terbentuknya vakuola-vakuola pada sel-sel kultur / jaringan dan terbentuknya fusi antar sel yang merupakan pengaruh dari serangan virus.
Pengamatan yang dilakukan Hedrick et al. (2000) dengan menggunakan mikroskop elektron pada jaringan organ yang terinfeksi, menemukan adanya perubahan antara lain pada sel-sel jaringan insang. Perubahan tersebut berupa pembengkakan sel, dan inti selnya mengalami hipertrofi yang diikuti dengan terjadinya difus atau penyebaran kromatin. Selanjutnya pada inti sel yang mengalamihipertrofi tersebut ditemukan adanya virion-virion, baik pada jaringan insang, hati, maupun limfosit dalam pembuluh darah di hati. Virion atau partikel virus tersebutberbentuk heksagonalyang merupakan tipikal Herpesvirus.
2.5. Pengaruh Lingkungan
Salah satu faktor lingkungan yang sangat berpengaruh terhadap perkembangan KHV adalah suhu. Oleh karena itu, memelihara ikan mas dan koi (Cyprinus carpio) pada suhu tertentu dapat membatasi pengaruh dari penyakit ini.
Hampir seluruh wabah penyakit KHV terjadi selama musim semi dan gugur pada saat suhu air sekitar 18-26°C (Hedrick et al, 2000; Perelberg et al., 2003). Pada suhu air yang lebih rendah virus dapat menginfeksi ikan tanpa menginduksi gejala klinis penyakit, tetapi pada suhu air yang memungkinkan perkembangan KHV, gejala klinis akan tampak dan selanjutnya dapat menyebabkan mortalitas (Gilad et al., 2004 dalam Hedrick et al., 2005). Infeksi KHV umumnya lebih serius pada suhu air antara 22-27°C (OATA, 2001 dalam Taukhid et al., 2004), menginfeksi ikan mas dan koi semua umur dengan ikan
Berkaitan dengan suhu, suatu penelitian dengan menggunakan sel koi fin (KF-1) telah dilakukan oleh Gilad et al. (2003), untuk mengetahui perkembangan KHV pada suhu yang berbeda. Hasil penelitian tersebut menunjukkan bahwa suhu sangat berpengaruh terhadap perkembangan KHV. Virus ini menginduksi fusi sel dan vakuolasi sitoplasmik pada sel-sel KF-1 dalam waktu 5 hari setelah inokulasi KHV pada suhu 20°C. Efek sitopatik yang meluas sangat jelas terlihat setelah 7-10 hari, dan berkembang pesat ke seluruh sel setelah 14 hari. Hasil penelitian tersebut juga menunjukkan bahwa KHV dapat tumbuh pada kisaran suhu 15-25°C, dengan suhu terbaik untuk replikasi KHV pada 20-25°C. Pada suhu 30°C dan 40°C, tidak ditemukan adanya pertumbuhan KHV, dan hanya pertumbuhan minimal yang terjadi pada suhu 10°C.
Berdasarkan kisaran suhu KHV, maka dapat dikembangkan suatu cara untuk mengendalikan penyakit melalui penginfeksian ikan dengan KHV pada suhu yang memungkinkan perkembangannya, dan kemudian merubah suhu pada kisaran yang dapat menghambat pertumbuhan KHV untuk menghindari munculnya gejala klinis penyakit dan untuk menginduksi imunitas ikan terhadap penginfeksian kembali. Tampaknya merubah suhu air dibawah atau diatas batas toleransi KHV (sebagai contoh 30°C atau 13°C) akan menghambat munculnya gejala klinis penyakit (Ronen et al., 2003 dalam Hedrick et al., 2005; Gilad et al., 2004).
Cara pemaparan ikan terhadap KHV dan perubahan suhu air yang tinggi, telah dilakukan dalam uji coba skala besar di Israel untuk menghasilkan ikan-ikan yang resisten secara alamiah terhadap KHV. Ketika ikan-ikan tersebut kebal terhadap penginfeksian kembali, maka ikan-ikan tersebut merupakan carrier
potensial dari KHV (Hedrick et al., 2005). Gilad et al. (2004) menemukan ikan-ikan yang diinfeksi pada suhu 13°C tidak menunjukkan gejala klinis tetapi mengandung DNA KHV yang dapat dideteksi dengan taqman PCR.
2.6. Sebaran Geografis
Sejak awal tahun 1995–1996 dilaporkan telah terjadi wabah penyakit baru yang menyerang ikan mas dan koi (Cyprinus carpio), tetapi secara formal baru dilaporkan terjadi di Jerman pada tahun 1997 (Bretzinger et al., 1997 dalam Hedrick et al., 2005). Penyebab aktual dari penyakit tersebut belum teridentifikasi hingga tahun 1998 menyusul investigasi yang dilakukan pada saat terjadinya wabah penyakit pada ikan mas dan koi di Israel dan Amerika Serikat (Hedrick et al., 2000). Hasil investigasi menunjukkan keberadaan suatu virus herpes yang selanjutnya disebut Koi herpesvirus atau KHV, pada ikan koi sakit asal Israel dan Amerika Serikat yang berhasil diisolasi dengan menggunakan suatu cell line yang baru dikembangkan dari koi fin (KF-1). Virus hasil isolasi tersebut menunjukkan dapat menginduksi karakteristik penyakit yang sama dan mortalitas tinggi seperti pada kejadian wabah alamiahnya melalui infeksi percobaan pada ikan koi di laboratorium (Hedrick et al., 2000).
Perkembangan selanjutnya menunjukkan penyebaran yang cepat dari KHV. Wabah penyakit KHV dilaporkan telah menyebabkan mortalitas yang tinggi pada ikan mas dan koi (Cyprinus carpio) di seluruh dunia. Negara-negara tersebut meliputi Amerika Serikat, beberapa negara Eropa, Afrika Selatan, Cina, Taiwan, Indonesia, dan Jepang ( Waltzek dan Hedrick, 2004; Sano et al., 2004 dalam Hedrick et al., 2005). Haenen dan Engelsma (2004) melaporkan serangan
yang meliputi Belgia (tahun 1999), Inggris (tahun 2000), Belanda (tahun 2002), Denmark (Juli tahun 2002), Perancis (tahun 2003), Austria (wabah pertama terjadi pada musim panas tahun 2003), Switzerland (tahun 2003), Luxemburg (tahun 2003), dan Italia (tahun 2003). Serangan KHV di negara-negara Asia meliputi Cina (tahun 2001), Indonesia (Maret tahun 2002), Taiwan (Januari tahun 2003), Jepang (Mei tahun 2003). Selanjutnya, KHV juga sudah menyerang ikan mas dan koi di Afrika Selatan (tahun 2003).
Serangan KHV di Indonesia pertama kalinya terjadi pada bulan Maret tahun 2002 di Blitar Jawa Timur. Wabah terjadi pada ikan koi yang baru datang dari Surabaya. Ikan koi ini diimpor ke Surabaya dari Cina melalui Hongkong pada bulan Desember 2001–Januari 2002. Wabah terjadi setelah hujan deras dengan total kematian mencapai 80–95%. Ikan yang sakit memperlihatkan gejala klinis berupa lepuh pada kulit. Oleh karena itu penyakit ini mula-mula dikenal sebagai penyakit melepuh. Meskipun ada gejala kerusakan insang pada ikan yang terkena wabah, tetapi gejala ini belum banyak diperhatikan. Blitar dikenal sebagai sentra produksi ikan koi, dengan lebih dari 5000 petani ikan koi. Koi dari Blitar termasuk yang sakit selanjutnya menyebar ke berbagai daerah termasuk ke Jawa Tengah, Jawa Barat dan Jakarta yang merupakan pangsa pasar utama (Balai Penelitian Perikanan Air Tawar, 2002; Sunarto et al., 2004; Taukhid et al., 2004).
Serangan berikutnya terjadi pada ikan mas di Subang, Jawa Barat pada bulan April 2002. Gejala klinis penyakit ini pada ikan mas di Subang sama dengan gejala klinis pada ikan koi di Blitar, yaitu insang rusak. Wabah ini menyebabkan kematian ratusan ton ikan mas, sehingga terjadi kelebihan supply
ikan konsumsi. Petani dari daerah lain membeli ikan mas sakit ini sehingga penyakit menyebar cepat ke daerah lain (Sunarto et al., 2004).
Pada bulan Mei-Juni 2002 terjadi serangan berikutnya di Waduk Cirata. Waduk Cirata merupakan salah satu dari tiga waduk di Sungai Citarum yang meliputi Waduk Saguling (4.425 unit karamba apung), Cirata (33.000 unit karamba terapung) dan Jatiluhur (2000 unit karamba apung). Beberapa minggu sebelum terjadi wabah, ada pembudidaya ikan yang memasukkan ikan dari Subang ke waduk Cirata. Sebelumnya tidak pernah terjadi wabah semacam ini. Kematian masal yang biasanya terjadi karena umbalan (up-welling) yang terjadi hampir setiap tahun terutama pada permulaan musim hujan (Oktober, November, Desember). Ikan yang sakit menunjukkan gejala klinis berupa nafsu makan menurun, lemah, beberapa gejala klinis lain yang tidak selalu ditemukan pada ikan sakit kecuali insang rusak (Sunarto et al., 2004).
Pada bulan Februari 2003, serangan penyakit ini sudah mencapai Lubuk Linggau, Sumatera Selatan. Gejala klinis ikan sakit di Sumatera ini sama persis dengan gejala klinis pada wabah yang terjadi di Jawa. Wabah di Lubuk Linggau ini diduga terjadi karena kontaminasi ikan mas sakit dari waduk Cirata, Jawa Barat. Selanjutnya wabah menyebar dari Lubuk Linggau ke daerah sekitarnya termasuk ke Bengkulu di selatan dan Jambi di sebelah barat (Sunarto et al., 2004). Pada bulan September 2004, wabah terus menyebar ke Sumatera Barat yang menyebabkan kematian masal ikan mas sebanyak 150 ton (Sunarto et al., 2004). Kasus serangan penyakit ini juga dilaporkan terjadi di Sumatera Utara yang menyebabkan kematian masal ikan mas di sekitar perairan Danau Toba (Balai Karantina Ikan Polonia, 2004). Pada bulan September tahun 2004 juga,
wabah ini telah menyebar ke Kalimantan Selatan melalui kontaminasi induk ikan mas dari Punten, Jawa Timur ke Kalimantan (Sunarto et al., 2004; Taukhid et al., 2004).
Berdasarkan hasil pemantauan hama dan penyakit ikan karantina (HPIK) yang dilakukan oleh unit-unit pelaksana teknis (UPT-UPT) karantina ikan di seluruh Indonesia, pada tahun 2010 KHV sudah ditemukan pada 17 provinsi di Indonesia yaitu DKI Jakarta, Jawa Barat, Jawa Tengah, DI Yogyakarta, Jawa Timur, Bali, Nusa Tenggara Timur, Lampung, Bengkulu, Jambi, Riau, Sumatera Barat, Sumatera Selatan, Sumatera Utara, Kalimantan Selatan, Sulawesi Tengah, dan Sulawesi Utara (Pusat Karantina Ikan, 2010).
Berdasarkan uraian di atas dapat diketahui penyebaran penyakit KHV tergolong sangat cepat. Bermula dari satu lokasi dan seterusnya menyebar secara lokal di sekitar lokasi tersebut serta menyebar secara jarak jauh. Dari setiap satu lokasi yang terserang penyakit, selanjutnya penyakit menyebar secara lokal di sekitarnya. Penyebaran penyakit ini di tempat-tempat pemeliharaan ikan mas dan koi (Cyprinus carpio) terjadi segera setelah pemasukan ikan sakit dari luar.
Pada sistem kolam air deras di sungai, penyakit ini menyebar ke hilir (ke bawah) tapi tidak ke hulu (bagian atas). Hal ini merupakan indikasi bahwa penyakit dapat menyebar melalui aliran air (Sunarto, 2004). Hasil penelitian Perelberg et al. (2003) menemukan partikel KHV masih dapat bertahan hidup di luar inang (dalam air) dan masih infektif sekurangnya selama 4 jam.
Beberapa mekanisme penularan penyakit KHV ini adalah : 1) menyebar di dalam farm melalui kontak langsung antara ikan sakit dan ikan sehat, melalui bangkai ikan sakit dan melalui air; 2) penularan lokal antar farm melalui air yang
terkontaminasi, dan kemungkinan juga penularan melalui peralatan yang terkontaminasi; 3) penularan jarak jauh, terutama melalui pemindahan ikan dari daerah wabah ke daerah bebas (Hedrick et al., 2000; Perelberg et al., 2003; Sunarto et al., 2004; Taukhid et al., 2004).