An improved method to evaluate the
o
-diphenol oxidase activity of soil
P. Perucci
a,*, C. Casucci
a, S. Dumontet
baDipartimento di Biotecnologie Agrarie ed Ambientali, Facolta` di Agraria, Universita` degli Studi di Ancona, Via Brecce Bianche, 60131 Ancona, Italy bDipartimento di Produzione Vegetale, Facolta` di Agraria, Universita` degli Studi della Basilicata, Via N. Sauro 85, 85100 Potenza, Italy
Accepted 5 May 2000
Abstract
Phenolic compounds are intermediary transformation products of many naturally occurring substances and synthetic chemicals, including crop residues, organic wastes, pesticides and industrial products. They are suitable substrates for the phenolases and peroxidases produced by the soil microflora. Although measurement of the enzymatic oxidation of phenolic compounds is an important measure of the soil micro-flora’s capacity to degrade potentially recalcitrant organics, the proposed methods to perform this measurement are few and questionable. In this paper a new, reliable and simple method to measure theo-diphenol oxidase activity in whole soil sample is proposed. The method is based on a spectrophotometric determination of a red compound (4-(N-proline)-o-benzoquinone) developed from enzymatic oxidation of catechol in the presence of proline.q2000 Elsevier Science Ltd. All rights reserved.
Keywords:o-Diphenol oxidase; Kinetic parameters; Soil enzyme activity
1. Introduction
The formation of humic substances is one of the least understood aspects of humus chemistry, yet one of the most intriguing. Studies on this subject are many (Steven-son, 1982; Malik, 1996) and research on this topic has theo-retical and practical implications (Tate, 1987). Several pathways have been proposed for the formation of humic substances during the decay of plant and animal remains in soil. The classical theory (Waksman, 1936) is that humic substances are modified lignin compounds but the majority of present-day researchers favour a mechanism involving quinones.
Many studies have demonstrated the importance of phenolic products and acids in the formation of humic substances (Schnitzer and Khan, 1972; Flaig et al., 1975; Haider et al., 1975), which are released during microbiolo-gical hydrolysis from many naturally occurring substances and synthetic chemicals (including crop residues, organic wastes, pesticides and industrial products). The relevance of microbial mediated processes in the transformation of phenolic compounds in soil is also widely acknowledged (Hartley and Whitehead, 1985).
Usually, phenolic products are unstable in soil and can be subject to oxidative conversion to quinones which
polymer-ise into humus-like macromolecules in the presence or absence of amino compounds. The oxidative conversion can occur either by abiotic reactions (Lehmann et al., 1987; Lehmann and Cheng, 1988) or by enzymatic oxida-tive processes (Martin and Haider, 1980; Bollag et al., 1982). Nevertheless, phenolic compounds in soil are mainly transformed by oxidative processes catalysed by phenolases and peroxidases produced by the soil microflora (Sjoblad and Bollag, 1981). Claus and Filip (1990) have demon-strated the importance of white-rot fungi and actinomycetes in the production of phenoloxidases such as laccase, tyrosi-nase and peroxidase.
The most important problems in the study of the oxida-tion of phenolic compounds in soils arise from the reducoxida-tion of substrate availability, caused by sorption to the soil components and the interference with the enzymic catalysis. In particular, Lehmann and Cheng (1988) studying the reac-tivity and the oxidation in soil of phenolic acids, found that a rapid chemical reaction occurred immediately upon contact of catechols with soil, which was followed by biochemical processes leading to a partial degradation and/or stabilisa-tion of reacstabilisa-tion products. In addistabilisa-tion, Claus and Filip (1988) reported the inhibiting effect of clay minerals and other soil-related adsorbents on the behaviour of phenoloxidases.
Quinones, which are formed by oxidation of phenolic compounds, are highly reactive and able to form various condensation products, especially in the presence of compounds with free amino groups (Hartley and
0038-0717/00/$ - see front matterq2000 Elsevier Science Ltd. All rights reserved.
PII: S 0 0 3 8 - 0 7 1 7 ( 0 0 ) 0 0 1 6 8 - 1
www.elsevier.com/locate/soilbio
* Corresponding author. Fax:139-71-2204858.
Whitehead, 1985). The polymerisation of mono-, di-, and trihydroxyphenols and quinones, in either the absence or the presence of amino groups (Mason and Peterson, 1955; Berry and Boyd, 1984) can lead to a covalent incorporation into humic macromolecules. Jackson and Kendal (1949) and Mason and Peterson (1955) in studying the enzymatic oxidation of catechol under aerobic conditions, obtained a red compound produced from the further reaction of proline with the quinone. Yamaguchi et al. (1970) and Mayaudon et al. (1973) used the Mason–Peterson method in order to evaluate the o-diphenoloxidase activity (o-DPO activity, EC 1.10.3.1) in mushrooms and in purified soil extracts, respectively. Yamaguchi et al. (1970) found that the red compound, which developed after the enzymatic oxidation of catechol and the reaction with proline, was stable within pH 6–7 with a molar absorptivity of 5.0×103at 525 nm.
Although numerous methods have been proposed to esti-mate the activity of phenolases in pure cultures of mush-rooms, fruit extracts, vegetables and crustaceans, studies dealing with estimations in soil are few. Mayaudon et al. (1973) and Bollag et al. (1987) used purified soil extracts to determine the phenolase activity but only Duxbury and Tate (1981) have proposed a method to evaluate the soil poly-phenol oxidase activity. Their method is questionable as the oxygen consumption during the assays can be due to both the overall oxidative systems present in soil (laccase, perox-idase, tyrosinase) and abiotic chemical oxidation. For this reason, this method cannot be used for the specific determi-nation ofo-DPO activity in soil.
This paper proposes a new, rapid and reliable method for assaying the soil o-DPO activity using spectrophotometric determination of the red compound derived from the reac-tion between the products ofo-DPO activity and proline.
2. Materials and methods
2.1. Soils
Moist samples of soil A (clayey, mesic, mixed calcic
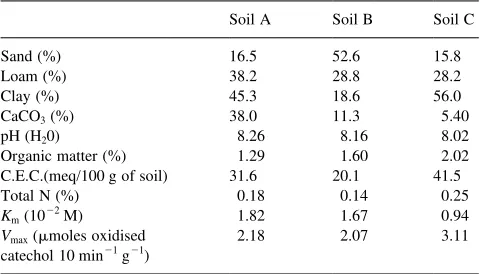
Haploxcrept), soil B (sandy-loam, mesic, mixed typic Haplustept) and soil C (clayey, mesic, mixed entic Haploxe-roll), (0–20 cm layer), were air-dried at 258C and sieved (2 mm) to remove plant material, soil macrofauna and stones. After sieving, the soil samples were allowed to equi-librate by storage at 208C for 3 d before use. The physical and chemical characteristics (pH, C.E.C., organic matter, total N and particle size distribution), determined according to ASA-SSSA methods (Page, 1982; Klute, 1986), are reported in Table 1.
2.2. Assay procedure
Reagent solutions of 0.1 M phosphate (0.1 M, pH 6.5), containing 0.2 M of catechol or 0.2 M of proline, were oxygenated for 3 min and incubated for 10 min at 308C. Then, 1 g of fresh soil was added to 3 ml of reagent solution (obtained by mixing 1.5 ml of catechol solution with 1.5 ml of proline solution) and 2 ml of phosphate buffer (0.1 M, pH 6.5). The mixture was incubated for 10 min at 308C and the reaction stopped by cooling in an ice-bath and adding 5 ml of ethanol. The mixture was centrifuged at 5000gat 48C for 5 min. The absorbance of the supernatant fraction was measured at 525 nm (Varian model Cary 210 double grating spectrophotometer). Assays without soil and without cate-chol were carried out simultaneously as controls.
Enzyme activity is expressed as mmoles of catechol oxidised 10 min21g21soil (on dry weight basis).
2.3. Optimum pH and temperature and stability of red compound
The pH optimum of theo-DPO activity in soil was deter-mined by adding the substrate catechol and proline in various phosphate buffer solutions (0.1 M) adjusted to pH values ranging from 5.5 to 8.0. Soil was then added to the solutions, and the o-DPO activity was measured as previously described.
The temperature optimum of soil o-DPO activity was determined by allowing the substrates (catechol and proline in equimolecular concentration) to equilibrate in the buffer solution at pH 6.5 for 10 min at temperatures ranging from 10 to 508C. Soil samples were added to the solutions and incubated at the corresponding temperatures for 10 min. Theo-DPO activity was measured as previously described. To evaluate the stability of the red compound under experimental conditions, the absorbance values were moni-tored every 10 min for 2 h. The red compound was found to be stable in all experimental conditions.
2.4. Michaelis–Menten parameters
The Michaelis–Menten constants (Km) and maximum
enzyme velocities (Vmax) were determined by using catechol
and proline (molar ratio 1:1) in buffered phosphate solution at seven different concentrations, varying from 0.02 to 0.4 M and computed from the Hanes–Wolf linear P. Perucci, C. Casucci / Soil Biology & Biochemistry 32 (2000) 1927–1933
1928 Table 1
Physico-chemical and chemical properties and biochemical parameters of soils
Organic matter (%) 1.29 1.60 2.02 C.E.C.(meq/100 g of soil) 31.6 20.1 41.5 Total N (%) 0.18 0.14 0.25
Km(1022M) 1.82 1.67 0.94
Vmax(mmoles oxidised
catechol 10 min21g21)
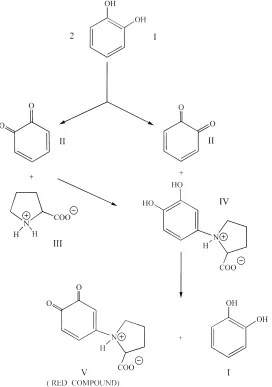
Fig. 1. Stoichiometry of reaction: (I) catechol; (II)o-benzoquinone; (III), proline; (IV) 4-(N-proline)-2-hidroxyphenol; (V) 4-(N-proline)-o-benzoquinone.
transformation of Michaelis–Menten equation:
S=V Km=Vmax1 S=Vmax
2.5. Abiotic chemical oxidation of catechol in soil
Assays of o-DPO activity were performed in sterilised soil samples to evaluate the abiotic chemical oxidation of catechol in soil under aerobic conditions, in the presence and absence of native organic matter. Sterilised soil samples were obtained by autoclaving fresh soil for 30 min at 1208C or by fumigation with CHCl3for 24 h (Jenkinson and Ladd
1981). The oxidation of native organic matter was achieved
by treating samples with H2O2in a water bath at 408C, until
the evolution of CO2had ceased.
2.6. Statistical analysis
In order to test the reproducibility of the this method, there were five replicates for each treatment. Anova was performed and the overall variability coefficient was calcu-lated for each treatment. The coefficient of variability ranged from 1.00 to 5.58%.
3. Results and discussion
The stoichiometry of the reaction between catechol and P. Perucci, C. Casucci / Soil Biology & Biochemistry 32 (2000) 1927–1933
1930
proline up to the formation of the red compound is reported in Fig. 1. This shows that one mole of the red compound (v) is produced from one mole of oxidised catechol. Therefore, the o-DPO activity was measured as mmole of catechol oxidised per 10 min per g of soil.
The o-DPO activity versus catechol concentration for each soil sample is shown in Fig. 2. According to these results it appears that the reaction (at a substrate concentra-tion of 0.2 M) did not reach its maximum. Tests carried out at higher catechol substrate concentrations showed slightly higher Vmax values. However, at high substrate
concentra-tions the abiotic chemical oxidation of the reagent takes place. This reaction produces a yellow product, which poly-merises to form a brown product (probably due to the condensation of quinones) which makes the subsequent measurement of the red compound difficult. Consequently, the 0.2 M substrate concentration is considered the maxi-mum concentration at which the test can be carried out without interference from abiotic catalysis.
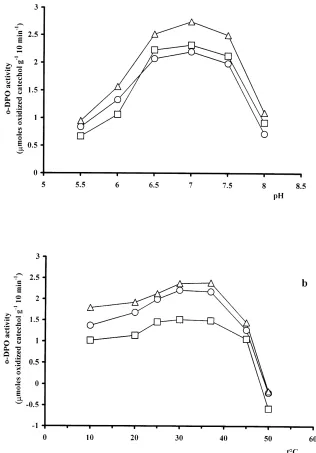
The behaviours of theo-DPO activity at different pH and temperature values after 10 min incubation are reported in Fig. 3a and b. The optimum pH and temperature to assayo -DPO activity were between 6.5–7.5 and from 30–378C. Tests carried out at pH 6.5 and 308C showed an o-DPO activity from 8–10% lower than that at pH 7.0 and 378C. However, pH 7.0 and 378C were not employed because a reduced stability of catechol was measured. In fact, follow-ing the oxidative abiotic reaction by dissolved oxygen in the reaction means, catechol is transformed to quinone, which condenses to form a black polymer that interferes with the absorbance of the red coloured product.
In order to investigate the validity of the proposed method, additional tests were carried out. The following combinations of different substrates and buffer were used: (i) catechol without proline; (ii) different substrates, such as l-DOPA and p-cresol1proline in the same phosphate buffer solution; (iii) crude enzyme extracts obtained from
soil by means of a phosphate buffer solution (0.1 M, pH 6.5 or 7.0) extraction (10 g of fresh soil was added to 10 ml of phosphate buffer solution, the mixture was then agitated on a horizontal shaker for 1 h and allowed to stand for 12 h at 08C before centrifugation at 2500g); (iv) acetate buffer (0.1 M, pH 6.5 or 7.0). In combination (i), no red colour developed after incubation, but after centrifugation and addition of proline a red colour appeared in the supernatant fraction. This confirms the oxidation of the catechol in the reaction mixture. In all the other tests the o-DPO activity was significantly less (30–70%) than that measured using the proposed procedure.
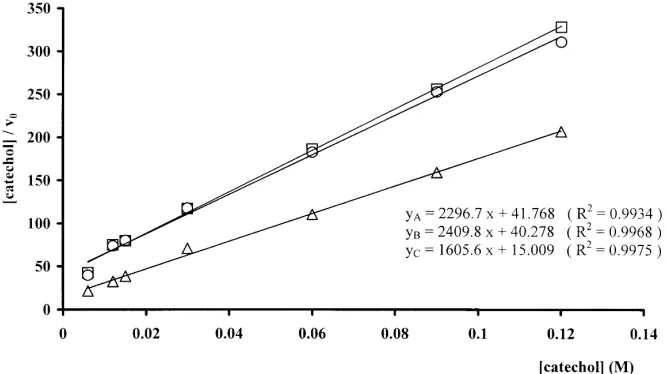
In Fig. 4 the Hanes–Wolf plots are shown and Table 1 reports the parameters of these linear regressions. The Km
values obtained have to be considered as ‘apparent’ values as the soil enzymic oxidation of catechol is always coupled to chemical oxidation.
It is well known that phenolic compounds in soil are rapidly adsorbed by clay minerals and hydroxides and under aerobic conditions undergo rapid chemical oxidation (Lehmann et al., 1987; Lehmann and Cheng, 1988; Claus and Filip 1990). For this reason, the soils used in this experi-ments were chosen because of their different particle size distribution (Table 1).
Measurements of o-DPO activity, using sterilised soil samples with or without native organic matter, were carried out in order to confirm that abiotic chemical oxidation occurs. Sterile soil achieved by autoclaving or by fumiga-tion, showed similar levels of catechol oxidation.
The enzymatic activity is obtained by subtracting the abiotic activity from the overall catechol oxidation. The data obtained in such way (Fig. 5a–c) are in agreement with the findings of Claus and Filip (1988) who found differ-ent behaviour of phenoloxidases in presence of clay and other soil-related adsorbents. These findings underline the importance of the soil’s physico-chemical characteristics in abiotic catechol oxidation.
In conclusion, it is recommend that sterilised soil controls should be used when using oxidation of catechol to evaluate soilo-DPO activity.
Acknowledgements
We thank Dr Andrea Onofri of the Dipartimento di Scienze Agro-Ambientali e della Produzione Vegetale, Universita` di Perugia, for his help with the statistical analysis.
References
Berry, D.F., Boyd, S.A., 1984. Oxidative coupling of phenols and aniline by peroxidase: structure–activity relationships. Soil Society Science of American Journal 48, 565–569.
Bollag, J.M., Liu, S.Y., Minard, R.D., 1982. Enzymatic oligomerization of vanillic acid. Soil Biology & Biochemistry 14, 157–163.
Bollag, J.-M., Chen, C.M., Sarkar, J.M., Loll, M.J., 1987. Extraction and purification of a peroxidase from soil. Soil Biology & Biochemistry 19, 61–67.
Claus, H., Filip, Z., 1990. Effects of clays and other solids on the activity of phenoloxidases produced by some fungi and actinomicetes. Soil Biol-ogy & Biochemistry 22, 483–488.
Claus, H., Filip, Z., 1988. Behaviour of phenoloxidases in the presence of
P. Perucci, C. Casucci / Soil Biology & Biochemistry 32 (2000) 1927–1933
1932
clays and other soil-related adsorbents. Applied Microbiology and Biotechnology 28, 506–511.
Duxbury, J.M., Tate III, R.L., 1981. The effect of soil depth and crop cover on enzymatic activities in Pahokee Muck. Soil Science Society of American Journal 45, 322–328.
Flaig, W., Beutelspacher, H., Rietz, E., 1975. Chemical composition and physical properties of humic substances. In: Gieneking, J.E. (Ed.). Soil Components, vol. 1. Springer, Berlin, pp. 1–211.
Haider, K., Martin, J.P., Filip, Z., 1975. Humus biochemistry. In: Paul, E.A., McLaren, A.D. (Eds.). Soil Biochemistry, vol. 4. Marcel Dekker, New York, pp. 195–244.
Hartley, R.D., Whitehead, D.C., 1985. Phenolic acids in soils and their influence on plant growth and soil microbial processes. In: Vaughan, D., Malcolm, R.E. (Eds.). Soil Organic Matter and Biological Activity, Martinus Nijhoff: Junk Publishers, Dordrect, pp. 109–149.
Jackson, H., Kendal, L.P., 1949. The oxidation of catechol and homocate-chol by tyrosinase in the presence of amino-acids. Biochemical Journal 44, 477–487.
Jenkinson, D.S., Ladd, J.N., 1981. Microbial biomass in soil: measurement and turnover. In: Paul, E.A., Ladd, J.N. (Eds.). Soil Biochemistry, vol. 5. Marcel Dekker, New York, pp. 415–471.
Yamaguchi, M., Hwang, P.M., Campbell, J.D., 1970. Latento-diphenol oxidase in mushrooms (Agaricus biporus). Canadian Journal of Biochemistry 28, 198–202.
Klute, A., 1986. Methods of Soil Analysis. Part 1. 2nd ed. American Society of Agronomy, Madison.
Lehmann, R.G., Cheng, H.H., Harsh, J.B., 1987. Oxidation of phenolic acids by iron and manganese oxides. Soil Science Society of American Journal 51, 352–356.
Lehmann, R.G., Cheng, H.H., 1988. Reactivity of phenolic acids in soil and formation of oxidation products. Soil Science Society of American Journal 52, 1304–1309.
Malik, K.A., 1996. Soil organic matter. In: Rashid, A., Memon, K.S., Bashir, E., Bantel, R. (Eds.). Soil Science, National Book Foundation, Islamabad, pp. 235–258 (chap. 8).
Mayaudon, J., El Halfawi, M., Chalvignac, M.A., 1973. Proprietes des diphenol oxydases extraites des sols. Soil Biology & Biochemistry 5, 369–383.
Martin, J.P., Haider, K., 1980. A comparison of the use of phenolase and peroxidase for the synthesis of model humic acid-type polymers. Soil Science Society of American Journal 44, 983–988.
Mason, H.S., Peterson, E.W., 1955. The reaction of quinones with prota-mine and nucleoprotaprota-mine:N-terminal proline. Journal of Biological Chemistry 212, 485–493.
Page, A.L., 1982. Methods in Soils Analysis. Part 2. 2nd ed. American Society of Agronomy, Madison.
Schnitzer, M., Khan, S.U., 1972. Humic Substances in the Environment. Dekker, New York.
Sjoblad, R.D., Bollag, J.-M., 1981. Oxidative coupling of aromatic compounds by enzymes from soil microorganisms. In: Paul, E.A., Ladd, J.N. (Eds.). Soil Biochemistry, vol. 5. Marcel Dekker, New York, pp. 113–152.
Stevenson, F.J., 1982. Humus Chemistry: Genesys, Composition, Reac-tions, Wiley, New York, p. 443.
Tate III, R.L., 1987. Soil Organic Matter. Biological and ecological effects, Wiley, New York, p. 291.