MAKALAH GENETIKA MOLEKULER SENTRAL DOGMA
OLEH FITRIA DELA
1410421006
DOSEN PENGAMPU : Dr. Dewi Imelda Roesma, M.Si.
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS ANDALAS
BAB I PENDAHULUAN 1.1 Latar Belakang
DNA sebagai bahan genetik karena DNA dapat mewariskan sifat-sifat organisma induk, sudah diidentifikasi pada pertengahan abad 20 (Greenspan and Baxter 1994). Genom adalah sepotong DNA/segment DNA yang menyandi protein mengandung semua informasi genetik yang dimilikinya. Dengan penemuan ini ditemukan bagaimana informasi genetik diwariskan dan diekspresikan. Mekanisme molekuler dari pewarisan melibatkan proses yang dikenal sebagai replikasi, dimana rantai DNA induk berfungsi sebagai cetakan untuk sintesis salinan DNA (Murray et al. 2000)
Setiap kita yang ingin memahami bagaimana metabolisme sel dan pewarisan sifat dari induk (parental) ke generasi berikutnya adalah dengan memahami mekanisme kerja DNA. Berbagai eksperimen dan kajian sampai pada kesimpulan bahwa mekanisme kerja DNA adalah replikasi, transkripsi dan translasi. Tiga proses ini dikenal dengan sebutan dogma sentral. Replikasi adalah proses menyalin secara utuh 2 untai DNA menjadi 2 untai yang baru.Transkripsi adalah proses menyalin salah satu untai DNA menjadi mRNA sedangkan translasi adalah proses penterjemahan mRNA menjadi polipeptida (Yuwono, 2013).
Materi genetik (DNA) yang terdapat pada suatu sel selalu dalam keadaan aktif karena senantiasa melakukan replikasi, transkripsi, translasi, reparasi (perbaikan) dan rekombinasi. Proses penyimpanan dan pemindahan informasi genetik dinyatakan dalam suatu dalil yang disebut dogma sentral, yang ditemukan oleh Francis Crick dan George Gamov pada tahun 1957. Prinsip dogma sentral bahwa DNA menjadi penentu jenis RNA yang selanjutnya akan diterjemahkan menjadi suatu protein (Yuwono, 2013).
Bahan genetik yang ada pada setiap jasad akan mengalami proses perbanyakan sebagai salah satu tahapan sangat penting dalam proses pertumbuhan sel atau perbanyakan partikel virus. Proses perbanyakan bahan genetik dikenal sebagai proses replikasi. Replikasi bahan genetik dapat dikatakan sebagai proses yang mengawali pertumbuhan sel, meskipun sebenarnya pertumbuhan merupakan suatu resultan banyak
proses yang saling berkaitan satu sama lain. Replikasi bahan genetik diikuti oleh pembentukan sel-sel anakan yang membawa duplikat bahan genetik hasil replikasi. Oleh karena itu, kesalahan dalam proses replikasi bahan genetik dapat mengakibatkan perubahan pada sifat-sifat sel anakan (Yuwono, 2005).
Pada transkripsi sinte-sa mRNA (messenger RNA) dari DNA template dilakukan oleh enzim RNA polymerase. Mula-mula RNA po-lymerase berikatan dengan bagi-an DNA template yang disebut promotor. Kemudian RNA poly-merase akan bergerak dengan a-rah 5‟→3‟, transkripsi akan di-mulai pada tempat yang disebut initiation site dan berakhir pada termination site. Sedangkan translasi (atau sintesa protein) adalah proses penerjemahan “bahasa” asam nukleat (mRNA) menjadi “ba-hasa” protein oleh ribosom dan tRNA. Mula-mula ribosom dan tRNA akan berikatan pada start codon (urutan basa nukleotida: AUG) di mRNA. Kemudian a-kan terjadi elongasi dan berakhir pada stop codon. Satu segmen DNA yang mengkode suatu pro-tein disebut open reading frame (orf).
1.2 Rumusan Masalah
1. Apakah yang dimaksud dengan Sentral Dogma? Bagaimanakah pembagian serta tahapan dari Sentral Dogma
3. Apakah yang dimaksud dengan replikasi? Bagaimanakah tahap dari replikasi? 4. Apakah yang dimaksud dengan transkripsi? Bagaimanakah tahap dari transkripsi? 5. Apakah yang dimaksud dengan translasi? Bagaimanakah tahap dari translasi? 6. Apa itu RNA coding dan noncoding?
1.3. Tujuan
1. Untuk mengetahui pengertian dari Sentral Dogma serta tahapan nya. 2. untuk mengetahui pengertian dari replikasi serta tahapannya
3. untuk mengetahui pengertian dari transkripsi serta tahapannya 4. untuk mengetahui pengertian dari translasi serta tahapannya
BAB II ISI
Dogma sentral biologi menjelaskan mengenai proses perubahan gen dari DNA menjadi RNA, dan RNA menjadi protein. Dogma ini menjelaskan bagaimana proses pembacaan materi genetik menjadi protein yang berperan di setiap tahap metabolisme di dalam tubuh suatu organisme (Bettelheim et al, 1984) Sebagai pernbawa informasi genetika, DNA rnempunyai dua fungsi utama: 1) rnembuat kopi yang tepat dari pada dirinya sendiri pada waktu proses repllkasi atau duplikasi dan 2) rneneruskan koda-koda informasi yang dimiliki ke.nRNA (tnessenger RNA) pada waktu proses transkripsi. Dengan demikian mRNA kelaknya dapat menterjemahkan (mengtranslasikan) informasi-informasi "bahasa dalam 4 huruf" dari pada asam nukleat ke dalam "bahasa dalam 24 huruf" darl pada protein. Konsep ini (gambar 1) merupakan dasar yang terkenal sebagai Dogma Sentral yang dlkemukakan oleh Crick (2) pada tahun 1958 (Soedigdo, 1973).
Dogma yang berlaku universal ini menyatakan bahwa sekali informasi telah diteruskan menjadi protein, maka tidak dapat dikembalikan menjadi bentuk asalnya (DNA). Aliran informasi dari asam nukleat ke asam nukleat memang memungkinkan, tetapi aliran informasi dari protein ke asam nukleat atau dari protein ke protein tidak memungkinkan. Dogma sentral terdiri dari tiga tahap yaitu replikasi, transkripsi dan translasi. Tahap replikasi dilakukan untuk memasok DNA pada setiap organisme, sedangkan tahap transkripsi bertujuan untuk menulis ulang DNA dalam bentuk mRNA (messenger RNA). Tahap translasi untuk menterjemahkan mRNA tersebut menjadi suatu protein (Yuwono, 2013).
Gambar 1. Tahap dalam sentral dogma
Secara umum, replikasi bahan genetik merupakan proses pengkopian rangkaian molekul bahan genetik (DNA atau RNA) sehingga dihasilkan molekul anakan yang sangat identik. Meskipun konsep dasar replikasi antara struktur bahan genetik yang satu dengan yang lainnya adalah serupa, namun diketahui ada banyak perbedaan dalam hal mekanisme rincinya. Sebagai contoh, bahan genetik yang berupa molekul RNA mempunyai mekanisme replikasi rinci yang berbeda dengan replikasi molekul DNA. Pada kelompok virus, misalnya , replikasi bahan genetiknya terjadi di dalam sel inang yang sebenarnya merupakan jasad hidup yang lain dari jasad virus itu sendiri. Hal ini dapat terjadi karena virus merupakan jasad parasit obligat. Di lain pihak, replikasi DNA pada prokaryot dan eukaryot terjadi di dalam sel jasad hidup yang bersangkutan. Selain itu perbedaan struktural molekul bahan genetik, misalnya antara DNA lingkar (Circular DNA) dengan DNA linear juga berimplikasi pada perbedaan mekanisme replikasi (Yuwono, 2005).
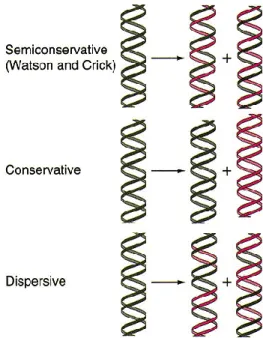
Beberapa hipotesa diajukan untuk menjelaskan proses sintesa/replikasi DNA ini yaitu model konservatif, semikonservatif dan dispersif. Pada model konservatif disintesa masing-masing satu untai lama dan satu untai baru, kemudian kedua untai tersebut mensintesa komplemennya. Model ini tidak sesuai dengan struktur DNA yang ada. Demikian pula model dispersif, tidak mungkin terjadi sintesa secara berselang-seling antar yang lama dan yang baru semacam suatu hybrid. Sifat komplementer pada model DNA untai ganda (double helix) dari Watson dan Crick memberi kesan bahwa replikasi DNA terjadi secara semikonsevatif. Dengan demikian, jika masing-masing untai pada molekul induk DNA untai ganda terpisah dari komplemennya saat replikasi, setiap bagian tersebut akan berfungsi sebagai cetakan (template), yang dengan cetakan ini disintesis sebuah untai komplementer yang baru (Yuwono, 2013)
Gambar 2. Tiga Hipotesa tentang Replikasi DNA
Replikasi DNA dimulai pada tempat-tempat khusus yang disebut pangkal replikasi (origin of replication). Pangkal replikasi yaitu satu bagian DNA yang mempunyai urutan nukleotida yang spesifik . Protein yang memulai replikasi DNA mengenali urutan ini dan menempel pada DNA, memisahkan kedua untaian dan membuka sebuah „gelembung‟ replikasi. Tahap pembukaan DNA untai ganda dikatalis oleh 3 macam enzim yaitu :
1. Helikase adalah sejenis enzim yang berfungsi membuka untai ganda di cabang replikasi, dan memisahkan kedua untai lama.
2. Enzim untai destabilizing protein , atau single stranded DNA binding protein (SSB), molekul dari protein pengikat untai tunggal kemudian berjajar disepanjang untai-untai lama yang tidak berpasangan menjaga agar untai-untai ini tetap terpisah selama mereka bertindak sebagai cetakan untuk sintesis untai-untai komplementer yang baru
3. DNA girase, enzim ini mengkatalis pembukaan untai ganda sebelum proses replikasi dimulai.
Replikasi DNA kemudian berjalan dalam dua arah sampai seluruh molekul tersebut disalin. Setiap kromosom eukariot mempunyai ratusan atau ribuan pangkal replikasi. Gelembung replikasi terbentuk dan akhirnya menyatu, sehingga mempercepat
penyalinan molekul DNA yang sangat panjang ini. Di setiap ujung gelembung replikasi terdapat cabang replikasi (replication fork), suatu daerah berbentuk huruf Y dimana untai DNA baru mulai memanjang (Yuwono, 2013).
Terdapat beberapa komponen-komponen penting dalam replikasi DNA. Replikasi bahan genetik ditentukan oleh beberapa komponen utama, yaitu: 1) DNA cetakan, yaitu molekul DNA atau RNA yang akan direplikasi. 2) molekul deoksi ribonukleotida yaitu dATP, dTTP, dCTP, dan dGTP. Deoksi ribonukleotida terdiri atas tiga omponen yaitu basa purin atau pirimidin, gula 5-karbon (deoksiribosa) dan gugus fosfat. 3) enzim polimerase, yaitu enzim utama yang mengkatalisis proses polimerasi nukelotida menjadi untaian DNA. Pada bakteri Escherichia coli terdapat tiga macam DNA polimerase yaitu DNA polimerase I, DNA polimerase II dan DNA polimerase III. Pada jasad eukaryot terdapat lima macam DNA polimerase yaitu DNA polimerase α, DNA polimerase δ, DNA polimerase ε, DNA polimerase β dan DNA polimerase γ. 4) Enzim primase yaitu enzim yang mengkatalisis sintesis primer untuk memulai replikasi DNA. Pada bakteri E. Coli kompleks enzim ini disebut primosom yang terdiri atas beberapa macam protein. 5) enzim pembuka ikatan untaian DNA induk, yaitu helikase dan enzim lain yang membantu proses tersebut yaitu enzim girase. 6) molekul protein yang menstabilkan untaian DNA yang sudah terbuka yaitu protein SSB (single strand bonding protein). 7) enzim DNA ligase yaitu suatu enzim yang berdungsi untuk menyambung fragmen-fragmen DNA (Yuwono, 2005).
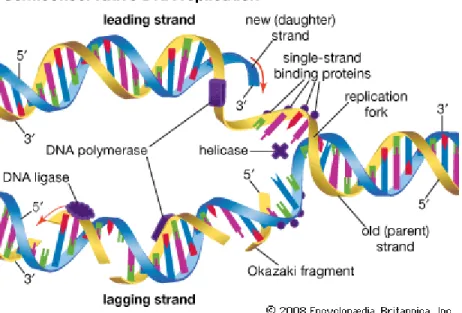
Enzim utama yang berpe-ran dalam replikasi adalah DNA polymerase. DNA polymerase mensintesa DNA baru dengan arah 5‟→3‟. DNA polymerase tidak dapat memulai pembuatan untaian DNA baru, enzim ini hanya dapat menambahkan nu-kleotida pada 3‟-OH yang sudah ada. Oleh karena itu, untuk memulai untai yang baru harus ada primer (biasanya berupa RNA) di mana DNA polymerase dapat menempelkan nukleotida yang pertama. Karena untaian DNA berpasangan secara anti-paralel, maka pada untai di-mana pembentukan untai DNA barunya itu dari 5‟ ke 3‟ akan terjadi sintesa DNA secara ber-sinambungan dan disebut lead-ing strand, sedangkan pada untai yang lainnya akan terjadi sintesa DNA dengan terputus-putus dan disebut lagging strand.
Re-plikasi dimulai pada tempat yang disebut ori (origin of repli-cation) dan berakhir pada termi-nator (Lucianus, 2003).
Gambar 3. bagian Replikasi DNA semikonservatif
Replikasi DNA berlangsung dalam beberapa tahap yaitu: (1) denaturasi (pemisahan) untaian DNA induk, (2) peng-”awal”-an (initiation, inisiasi) sintesis DNA, (3) pemanjangan untaian DNA, (4) ligasi fragmen fragmen DNA, dan (5) peng-“akhir”-an (termination, terminasi) sintesis DNA. Sintesis untaian DNA baru akan dimulai setelah kedua DNA induk terpisah membentuk garpu replikasi, pemisahan dilakukan oleh enzim DNA helikase. Sintesis DNA berlangsung dengan orientasi 5‟-P→3‟-OH. Oleh karena ada dua untaian DNA cetakan yang orientasinya berlawanan, maka sintesis kedua untaian DNA baru juga berlangsung dengan arah geometris yang berlawanan namun semuanya teteap dengan orientasi5‟→3‟. Keadaan semacam ini menimbulkan perbedaan dalam hal meanisme sintesis antara keua untaian DNA yang baru (Yuwono, 2005).
Proses replikasi diawali dengan pembukaan untaian ganda DNA pada titik-titik tertentu di sepanjang rantai DNA. Proses pembukaan rantai DNA ini dibantu oleh beberapa jenis protein yang dapat mengenali titik-titik tersebut, dan juga protein yang mampu membuka pilinan rantai DNA. Setelah cukup ruang terbentuk akibat pembukaan untaian ganda ini, DNA polimerase masuk dan mengikat diri pada kedua rantai DNA
yang sudah terbuka secara lokal tersebut. Proses pembukaan rantai ganda tersebut berlangsung disertai dengan pergeseran DNA polimerase mengikuti arah membukanya rantai ganda. Monomer DNA ditambahkan di kedua sisi rantai yang membuka setiap kali DNA polimerase bergeser. Hal ini berlanjut sampai seluruh rantai telah benar-benar terpisah
Pada model replikasi biasa umumnya replikasi berlangsung ke satu arah sehingga hanya ada satu untaian DNA awal dan satu untaian DNA lambat. Pada kenyataannya telah diketahui bahwa replikasi juga dapat berlangsung ke dua arah yang berlawanan, yiatu dikenal sebagai replikasi dua arah (bidirectional replication). Dalam replikasi dua arah akan terbentuk dua garpu replikasi yang bergerak ke arah yang berlawanan. Pada kedua garpu replikasi tersebut juga terjadi sintesis DNA baru secara kontinu dan diskontinu sehingga ada dua untaian DNA awal dan dua untaian DNA lambat (Yuwono, 2005).
Inisiasi (peng-„awal‟-an) replikasi DNA adalah proses permulaan sintesis untaian DNA yang sebelumnya didahului oleh sintesis molekul primer. Proses inisiasi berlangsung dengan mekanisme yang berbeda antara suatu jasad dengan jasad yang lain. Pada bakteri E. coli sintesis primer dikatalisis oleh kompleks protein yang disebut primosom. Primosom terdiri atas enzim primase/DnaG (berperan dalam sintesis molekul primer). Protein PriA, PriB, PriC, DnaTm, DnaB dan DnaC. Pada E. coli diketahui ada dua tipe atau sistem reaksi inisiasi, yaitu (1) sistem фX dan (2) sistem oriC. Sistem фX adalah sistem inisiasi replikasi DNA virus фX174, sedangkan sistem oriC aladah sistem inisiasi yang melibatkan ori pada E. coli (Yuwono, 2005).
Struktur untai ganda ini akan mempengaruhi replikasi DNA. DNA polimerase menambahkan nukleotida hanya pada ujung 3' yang bebas dari untai DNA yang sedang terbentuk, tidak pernah pada ujung 5'. Jadi untai DNA baru dapat memanjang hanya pada arah 5'→ 3'. Di sepanjang salah satu untai cetakan, DNA polimerase dapat mensintesa untai komplementer yang kontinu memanjangkan DNA yang baru dengan arah 5'→3'. Untai DNA yang dibuat dengan metode ini disebut leading strand. Untuk memanjangkan untai baru DNA yang lain, polimerase harus bekerja disepanjang cetakan jauh dari cabang replikasi. Untai DNA yang disintesis dalam arah ini disebut lagging
strand. Berbeda dengan leading strand, yang memanjang terus menerus, lagging strand pertama kali disintesis sebagai serangkaian segmen. Potongan ini disebut fragmen Okazaki, sesuai dengan nama ilmuwan Jepang yang menemukannya. Panjang fragmen-fragmen ini sekitar 100 sampai 200 nukleotida pada eukariot. Proses
ini mengandung satu untai utuh DNA anak mengikuti DNA induk dan satu untai lagi fragmen berupa DNA anak. Fragmen anak ini kemudian dirangkaikan menjadi satu untai utuh oleh enzim DNA ligase sehingga akhirnya satu DNA untai ganda menghasilkan 2 DNA anak untai ganda dan seterusnya (Yuwono, 2013).
Setelah dilakukan inisiasi dan polimerasi akhirnya proses replikasi DNA akan diakhiri dengan proses terminasi atau pengakhiran replikasi. Pada prokaryot replikasi genom berbentuk lingkar akan berakhir pada waktu kedua garpu replikasi bertemu pada suatu titik namun pada eukaryot keadaannya menjad lain karena struktur genomnya linear sehingga ada komplikasi terminasi replikasi pada ujung-ujung kromosom. Titik tempat pengakhiran replikasi disebut sisi terminasi. Pada E. coli ada enam sisi terminasi yaitu TerA, TerB, TerC, TerD, TerE, dan TerF. Urutan konsensus sisi terminasi adalah sebagai berikut AATTAGTATGTTGTAACTAAANT. Urutan basa DNA tersebut diperlukan untuk menghentikan garpu replikasi yang mendekati daerah tersebut dari ujung 5‟ (Yuwono, 2005).
Kesalahan proses replikasi molekul DNA hanya terjadi satu dalam 1 miliar nukleotida, tetapi kesalahan pemasangan awal antara nukleotida yang sudah ada pada untai cetakan dapat mencapai 100.000 kalinya atau sebesar 10.000 pasang basa. Sel memiliki mekanisme reparasi yaitu perbaikan salah pasang (mismatch repair ) yang akan memperbaiki kesalahan-kesalahan yang terjadi ketika DNA disalin. Selama replikasi DNA, DNA polimerase sendirilah yang melakukan perbaikan salah-pasang. Polimerase ini mengoreksi setiap nukleotida terhadap cetakannya begitu nukleotida ditambah pada untaian. Dalam rangka mencari nukleotida yang pasangannya tidak benar, polimerase memindahkan nukleotida tersebut kemudian melanjutkan kembali sintesis. Protein lain selain DNA polimerase juga melakukan perbaikan salah-pasang (Yuwono, 2013).
Transkripsi merupakan tahapan penting dalam sintesis protein atau ekspresi gen. Proses transkripsi terjadi pada nukleus (prokaryotik: nukleoid) di mana DNA diterjemahkan menjadi kode-kode dalam bentuk basa nitrogen membentuk rantai RNA yang bersifat single strain. Namun, pada rantai RNA yang terbentuk basa Timin digantikan dengan basa Urasil. Pada prokaryotik, rantai RNA langsung ditranslasikan sebelum transkripsi selesai. Sedangkan pada eukaryotik, rantai di bawa menuju sitoplasma (ribosom) untuk ditranslasi menjadi produk gen. Pembentukan RNA pada proses transkripsi melibatkan enzim RNA polymerase. Fungsi dasar kedua yang harus dijalankan oleh DNA sebagai materi genetik adalah fungsi fenotipik. Artinya, DNA harus mampu mengatur pertumbuhan dan diferensiasi individu organisme sehingga dihasilkan suatu fenotipe tertentu. Fungsi ini dilaksanakan melalui ekspresi gen, yang tahap pertamanya adalah proses transkripsi, yaitu perubahan urutan basa molekul DNA menjadi urutan basa molekul RNA. Dengan perkataan lain, transkripsi merupakan proses sintesis RNA menggunakan salah satu untai molekul DNA sebagai cetakan (templat) nya (Warianto, 2011).
Transkripsi terdiri dari tiga tahap, yaitu: (a) Inisiasi (permulaan). Transkripsi diawali oleh promoter, yaitu daerah DNA tempat RNA polimerase melekat. Promoter mencakup titik awal transkripsi dan biasanya membentang beberapa pasang nukleotida di depan titik awal tersebut. Fungsi promoter selain menentukan di mana transkripsi dimulai, juga menentukan yang mana dari kedua rantai ganda DNA yang digunakan sebagai cetakan. (b) Elongasi (pemanjangan). Ketika RNA bergerak di sepanjang DNA, pilinan rantai ganda DNA tersebut terbuka secara berurutan kira-kira 10-20 basa DNA. Enzim RNA polimerase menambahkan nukleotida ke ujung 3‟ dari molekul RNA yang dibentuk di sepanjang rantai ganda DNA. Setelah sintesis RNA berlangsung, rantai ganda DNA akan terbetuk kembali dan RNA baru akan terlepas dari cetakannya. (c) Terminasi (pengakhiran). Transkripsi berlangsung hingga RNA polimerase mentranskripsi urutan DNA yang dinamakan terminator. Terminator merupakan urutan DNA yang berfungsi untuk mengakhiri proses transkripsi. Pada prokariotik, transkripsi berhenti pada saat RNA polimerase mencapai titik terminasi. Pada eukariotik, RNA
polimerase terus melewati titik terminasi, 10-35 nukleotida, RNA yang telah terbentuk terlepas dari enzim tersebut (Kusnadi, 2010).
Interaksi antara RNA polimerase eukariot dan faktor transkripsi merupakan suatu contoh betapa pentingnya interaksi protein-protein dalam mengontrol transkripsi. Salah satu faktor transkripsi adalah TATA box. Pada sel eukariot enzim yang mentranskripsi gen pengkode-protein menjadi pra mRNA ialah RNA polimerase II. Enzim ini memulai sintesis RNA pada promoter yang biasanya berupa TATA box yaitu suatu urutan nukleotida TATAAAA. TATA ini terletak kira-kira 25 nukleotida upstream jauhnya dari titik awal transkripsi. RNA polimerase II tidak dapat mengenali TATA dan tanda-tanda khusus lain pada promoter. Protein lain dari faktor transkripsi yang membantu mengenali TATA ini, mengikatkan diri pada DNA sebelum RNA polimerase memulai transkripsi (Yuwono, 2013).
Jika inisiasi berhasil, RNA polimerase melepaskan faktor s, dan bersama-sama dengan DNA dan RNA nasen (RNA yang baru disintesis), akan membentuk kompleks terner atau kompleks yang terdiri atas tiga komponen. Dengan adanya kompleks terner ini RNA polimerase dapat berjalan di sepanjang molekul DNA. Artinya, promoter akan ditinggalkannya untuk kemudian ditempati oleh holoenzim RNA polimerase berikutnya sehingga terjadi reinisiasi transkripsi. Bagian DNA yang mengalami pembukaan heliks, atau disebut dengan gelembung transkripsi (transcription bubble), akan terlihat bergeser di sepanjang molekul DNA sejalan dengan gerakan RNA polimerase. Panjang bagian DNA yang mengalami pembukaan heliks tersebut relatif konstan, yakni sekitar 17 pb sedangkan ujung 5’ molekul RNA yang disintesis akan membentuk heliks hibrid dengan pita antisens DNA sepanjang lebih kurang 12 pb. Ukuran ini ternyata tidak mencapai satu putaran heliks. RNA polimerase E. coli bergerak dengan kecepatan rata-rata 40 nukleotida per detik. Akan tetapi, angka ini
dapat bervariasi sesuai dengan urutan lokal DNA (urutan DNA yang telah dicapai oleh RNA polimerase). Tetap dipertahankannya bagian DNA yang mengalami pembukaan heliks menunjukkan bahwa RNA polimerase membuka heliks DNA di depan gelembung transkripsi dan menutup heliks DNA di belakangnya. Dengan demikian, heliks hibrid RNA-DNA harus berputar setiap kali terjadi penambahan nukleotida pada
RNA nasen (Warianto, 2011).
Satu gen tunggal dapat ditranskripsi secara simultan oleh beberapa molekul RNA polimerase yang saling mengikuti seperti barisan truk dalam satu konvoi. Untai RNA yang sedang tumbuh memperlihatkan jejak dari setiap polimerase, dengan panjang setiap untai baru yang mencerminkan sejauh mana enzim itu telah berjalan dari titik awalnya. Transkripsi berlangsung sampai RNA polimerase menyalin urutan DNA
yang disebut terminator. Terminator merupakan suatu urutan RNA yang berfungsi sebagai sinyal pengakhiran transkripsi. Pada sel prokariot, transkripsi biasanya berhenti tepat pada akhir sinyal terminasi. Ketika polimerase mencapai titik tersebut polimerase melepas RNA dan DNA. Sebaliknya pada sel eukariot transkripsi akan berhenti setelah polimerase melewati sinyal terminasi yaitu suatu urutan AAUAAA di dalam pra-mRNA sejauh kira-kira 10-35 nukleotida. Pra30 mRNA ini akan dipotong hingga terlepas dari enzim tersebut. Tempat pemotongan pada RNA juga merupakan tempat untuk penambahan ekor-poli A (Yuwono, 2013).
Terminasi menggunakan protein rho selain karena adanya struktur tusuk konde, terminasi transkripsi dapat juga terjadi dengan bantuan suatu protein khusus yang dinamakan protein rho (ρ). Rho merupakan protein heksamer yang akan menghidrolisis ATP dengan adanya RNA untai tunggal. Protein ini nampak terikat pada urutan sepanjang 72 basa pada RNA, yang diduga lebih disebabkan oleh pengenalan suatu struktur spesifik daripada karena adanya urutan konsensus. Rho bergerak di sepanjang RNA nasen menuju kompleks transkripsi. Pada kompleks transkripsi ini rho memungkinkan RNA polimerase untuk berhenti pada sinyal terminator tertentu. Sinyal-sinyal terminator ini, seperti halnya Sinyal-sinyal terminator yang tidak bergantung kepada rho, lebih dikenali oleh RNA daripada oleh DNA cetakannya. Adakalanya terminator tersebut juga berupa struktur tusuk konde tetapi tidak dikuti oleh urutan poli U (Warianto, 2011)
TRANSLASI
Translasi adalah proses penerjemahan urutan nukleotida yang ada pada molekul mRNA menjadi rangkaian asam-asam amino yang menyusun suatu polipeptida atau protein.
Perlu dipahami bahwa hanya molekul mRNA yang ditranslasi, sedangkan rRNA dan tRNA tidak ditranslasi. Molekul mRNA merupakan transkrip (salinan) urutan DNA yang menyusun suatu gen dalam bentuk ORF (open reading frame), kerangka baca terbuka). Molekul rRNA adalah salah satu molekul penyusun ribosom, yakni organel tempat berlangsungnya sintesis protein, sedangkan tRNA adalah pembawa asam-asam amino yang akan disambungkan menjadi rantai polipeptida. Suatu ORF dicirikan oleh: (1) kodon inisiasi translasi yaitu urutan ATG (pada DNA) atau AUG (pada mRNA) (2) serangkaian urutan nukleotida yang menyusun banyak kodon dan (3) kodon terminasi translasi yaitu RAA (UAA pada mRNA), TAG (UAG pada mRNA) atau TGA (UGA pada mRNA) perlu diingat bahwa pada RNA tidak ada basa thymine (T) melainkan dalam bentuk uracil (U) (Yuwono, 2013).
Translasi berlangsung di dalam sitoplasma dan ribosom. Translasi merupakan proses penterjemaahan sutu kode genetik menjadi protein yang sesuai. Kode genetik tersebut berupa kodon di sepanjang molekul RNAd, sebagai penterjemaahnya RNAt. RNAt membawa asam amino dari stoplasma ke ribosom. Molekul RNAt membawa asam amino spesifik pada salah satu ujungnya yang sesuai dengan triplet nukleotida pada ujung RNAt lainnya yang disebut antikodon. Misalnya, perhatikan kodon RNAd UUU yang ditranslasi sebagai asam amino fenilalanin. RNAt pembawa fenilalanin mempunyai antikodon AAA yang komplemen dengan UUU agar terjadi reaksi penambahan fenilalanin pada rantai polipeptida sebelumnya. RNAt yang mengikat diri pada kodon RNAd harus membawa asam amino yang sesuai ke dalam ribosom. Melekatnya asam amino pada RNAt dibantu oleh enzim aminoasil-RNAt sintetase (aminoacyl-tRNA synthetase). Ribosom memudahkan pelekatan antara antikodon RNAt dengan kodon RNAd selama sintesis protein. Ribososm tersususn atas subunit besar dan subunit kecil yang dibangun oleh protein-protein dan molekul-molekul RNAt (Kusnadi, 2010).
Kita dapat membagi translasi menjadi 3 tahap yaitu insiasi, elongasi dan terminasi. Semua tahapan ini memerlukan faktor-faktor protein yang membantu mRNA, tRNA dan ribosom selama proses translasi. Untuk inisiasi dan elongasi rantai dibutuhkan sejumlah energi yang disediakan oleh GTP (guanin triphospat) yaitu suatu
molekul yang mirip ATP. Tahap inisiasi dari translasi membawa bersama-sama mRNA, sebuah tRNA yang memuat asam amino pertama dari polipeptida, dan dua subunit ribosom. Pertama subunit ribosom kecil mengikatkan diri pada mRNA dan tRNA inisiator khusus. Pada tahap elongasi, asam-asam amino ditambahkan satu persatu pada asam amino pertama. Tiap penambahan melibatkan partisipasi beberapa protein yang disebut faktor elongasi dan terjadi dalam siklus tiga tahap yaitu:
1. Pengenalan kodon. Kodon mRNA pada tempat A dari ribosom membentuk ikatan hidrogen dengan antikodon molekul tRNA yang baru masuk yang membawa asam amino yang tepat. Faktor elongasi membawa tRNA ke tempat A. Langkah ini juga membutuhkan hidrolisis GTP.
2. Pembentukan ikatan peptida. Molekul rRNA dari subunit ribosom besar, berfungsi sebagai ribozim, mengkatalis pembentukan ikatan peptida yang menggabungkan polipeptida yang memanjang dari tempat P ke asam amino yang baru tiba di tempat A. Pada tahap ini, polipeptida memisahkan diri dari tRNA tempat pelekatannya semula, dan asam amino pada ujung karboksilnya berikatan dengan asam amino yang dibawa oleh tRNA di tempat A.
3. Translokasi. Molekul tRNA di tempat A, sekarang terikat pada polipeptida yang sedang tumbuh, ditranslokasikan ke tempat P. Saat RNA berpindah tempat, antikodonnya tetap berikatan dengan hidrogen pada kodon mRNA; mRNA bergerak bersama -sama dengan antikodon ini dan membawa kodon berikutnya untuk ditranslasi pada tempat A. Sementara t RNA yang tadinya berada pada tempat P bergerak ketempat E dan dari tempat ini keluar dari ribosom. Langkah translokasi membutuhkan energi yang disediakan oleh hidrolisis GTP. mRNA bergerak melalui ribosom ke satu arah saja, mulai dari ujung 5' hal ini sama dengan ribosom yang bergerak 5'→3' pada mRNA. Hal yang penting disini adalah ribosom dan mRNA bergerak relatif satu sama lain, dengan arah yang sama, kodon demi kodon. Siklus Elongasi menghabiskan waktu kurang dari 1/10 detik dan terus diulang saat tiap asam amino ditambahkan pada rantai hingga polipeptidanya lengkap.
Asam amino Kodon RNA
Alanin GCU GCC GCA GCG
Arginin CGU CGC CGA CGG AGA AGG
Asam aspartat GAU GAC
Asam glutamat GAA GAG
Asparagin AAU AAC
Fenilalanin UUU UUC
Glisin GGU GGC GGA GGG
Histidin CAU CAC
Isoleusin AUU AUC AUA
Leusin CUU CUC CUA CUG UUA UUG
Lisin AAA AAG
Metiosin AUG
Prolin CCU CCC CCA CCG
Serin UCU UCC UCA UCG
Sistein UGU UGC
Tirosin UAU UAC
Treonin ACU ACC ACA ACG
Triptofan UGG
Valin GUU GUC GUA GUG
Mulai (Start) AUG GUG
Berhenti (Stop) UAA UAG UGA
Translasi akan berakhir pada waktu salah satu dari ketiga kodon terminasi (UAA, UGA, UAG) yang ada pada mRNA mencapai posisi A pada ribosom. Dalam keadaan normal tidak ada aminoasil-tRNA yang membawa asam amino sesuai dengan ketiga kodon tersebut. oleh karena itu, jika ribosom mencapai salah satu dari ketiga kodon terminasi tersebut, maka proses translasi berakhir. Pada E. coli ketiga sinyal penghentian proses translasi tersebut dikenali oleh suatu protein yang disebut release factors (RF)
misalnya RF1 yang mengenali kodon UAA atau UAG atau RF2 yang mengenali kodon UAA atau UGA. Sebaliknya pada eukaryot hanya ada satu release factor yaitu eRF yang mengenali ketiga kodon terminasi tersebut (Yuwono, 2005).
CODING dan NON CODING RNA
Gena adalah unit heriditas suatu organisme hidup. Gen ini dikode dalam material genetik organisme, yang kita kenal sebagai molekul DNA, atau RNA pada beberapa virus, dan ekspresinya dipengaruhi oleh lingkungan internal atau eksternal seperti perkembangan fisik atau perilaku dari organisme itu. Gena tersusun atas daerah urutan basa nukleotida baik yang mengkode suatu informasi genetik (coding-gene region as exon) dan juga daerah yang tidak mengkode informasi genetik (non-coding-gene region as intron), hal ini penting untuk pembentukan suatu protein yang fungsinya diperlukan ditingkat sel, jaringan, organ atau organisme secara keseluruhan.
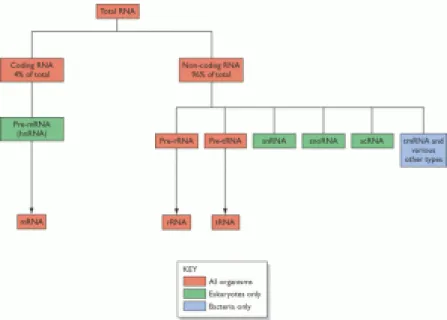
Menurut penelitian Jacob dan Monod (1961). mengidentifikasi peranan molekul antara yang labil keberadaannya maka terbangun suatu hubungan konsepsional penterjemahan informasi dari urutan basa DNA ke dalam urutan asam amino protein atau struktur primer protein. Pada proses transkripsi menghasilkan tiga jenis RNA yaitu mRNA (massengerRNA), tRNA (transfer RNA), rRNA (ribosomal RNA). dalam pembagian berdasarkan fungsinya terdapat 2 pembagian utama yaitu coding RNA dan non-coding RNA. Berikut adalah bagan dari pembagian total RNA :
Gambar 4. bagan RNA dalam sel
Coding RNA terdiri dari transkripton dan hanya terdiri dari satu kelas molekul yaitu: Messenger RNA (mRNAs), yang mentraskrip gen pengkode protein dan karenanya ditranslasi menjadi protein dalam tahapan selanjutnya pada ekspresi genom. mRNA ini sendiri berfungsi sebagai cetakan dalam sintesis protein, dan hanya mRNA yang akan diterjemahkan dalam protein, selain itu mRNA juga memiliki umur yang sangat pendek yaitu akan segera terdegradasi setelah sintesis.
Tipe yang kedua dari RNA adalah non-coding, ada dua tipe yaitu:
1. Transfer RNA (tRNA) yaitu molekul kecil yang terlibat dalam dalam sintesis protein.tRNA membawa asam amino dalam bentuk yang di aktifkan ke dalam ribosom untuk pembentukan ikatan peptida dalam suatu urutan yang ditentukan oleh mRNA sebagai cetakan.
2. Ribosomal RNA (rRNA) merupakan salah satu komponen utama Ribosom dan merupakan tipe RNA yang jumlahnya paling melimpah dalam sel serta menyusun sekitar 80% dari total RNA pada bakteri yang sedang aktif membelah. molekul ini merupakan struktur tempat berlangsungnya sintesis protein. Eukariot mempunyai tipe non-coding RNA lain yang distribusinya lebih terbatas, dan RNA non-codingnya pendek. tipe RNA non-coding ini biasanya di bagi dalam 3 kategori sesuai dengan lokasi utamanya di dalam sel yaitu :
·Small nuclear RNA (snRNA), disebut juga U-RNA karena kaya akan nukleotida uridin yang terlibat dalam pemrosesan mRNA.
·Small nucleolar RNA (snoRNA), yang berperan penting dalam pemrosesan molekul rRNA.
·Small cytoplasmic RNA (scRNA), merupakan kelompok molekul yang sebagian fungsinya sudah diketahui dan sebagian lagi masih misteri.
Perbedaan antara coding RNA dan non-coding RNA adalah :
1. Pada coding RNA terdiri dari transkripton dan hanya terdiri dari dari satu kelas molekul yaitu : Messenger RNA (mRNAs) sedangkan pada non-coding RNA baik pada prokariot maupun eukariot terdapat dua tipe yaitu: Ribosomal RNA (rRNA) dan Transfer RNA (tRNA).
2. mRNA, tRNA maupun rRNA sama-sama berperan dalam proses translasi tetapi hanya mRNA yang diterjemahkan dalam protein.
3. mRNA berfungsi sebagai cetakan sintesis protein, dan tRNA berfungsi sebagai pembawa asam amino ke ribosom untuk dirangakai dengan urutan yang sesuai dengan sekuens nukleotida pada mRNA yang sedang di translasi, sedangkan rRNA merupakan komponen penyusun ribosom yang merupakan struktur tempat berlangsungnya sintesis protein.
4. pada RNA non-coding yang lebih terbatas distribusinya di bagi menjadi 3 kategori sesuai dengan lokasi utamanya dalam sel yaitu: small nuclear RNA (snRNA), small nukeolar RNA (snoRNA), small cytoplasmic RNA (scRNA) (Brown, 2000).
BAB III PENUTUP 3.1 Kesimpulan
Dari uraian diatas, dapat diambil kesimpulan sebagai berikut:
1. Sentral Dogma adalah proses penyimpanan dan pemindahan informasi genetik melalui 3 tahap yaitu replikasi, transkripsi dan translasi.
2. Replikasi adalah proses pengkopian rangkaian molekul bahan genetik (DNA atau RNA) sehingga dihasilkan molekul anakan yang sangat identik.
3. Tahapan proses replikasi diantaranya adalah denaturasi (pemisahan) untaian DNA induk, inisiasi (sintesis DNA), pemanjangan untaian DNA, ligasi fragmen-fragmen DNA dan terminasi (sintesis DNA)
4. Transkripsi adalah proses sintesis protein dimana DNA diterjemahkan menjadi kode-kode dalam bentuk basa nitrogen membentuk rantai RNA yang bersifat single strain.
5. Tahapan dalam proses transkripsi diantaranya adalah inisiasi, elongasi dan terminasi.
6. Translasi adalah proses penerjemahan urutan nukleotida yang ada pada molekul mRNA menjadi rangkaian asam-asam amino yang menyusun suatu polipeptida atau protein.
7. Tahapan dalam proses translasi diantaranya adalah inisiasi, elongasi dan terminasi. 8. Dalam pembagian berdasarkan fungsinya terdapat 2 pembagian utama yaitu coding
RNA dan non-coding RNA. Coding RNA terdiri dari transkripton dan hanya terdiri dari satu kelas molekul yaitu messenger RNA (mRNAs), sedangkan non coding RNA terdiri dari tRNA dan rRNA.
3.2 Saran
Adapun saran untuk makalah ini adalah sebaiknya penjelasan mengenai Sentral Dogma lebih diperbanyak kembali dan uraian tahapan tahapan dalam sentral dogma yang lebih diperjelas.
DAFTAR PUSTAKA
Bettelheim, F.A., & Landesberg, J. 1984. Laboratory Experiments for General, Organic, & Biochemistry. (edisi keempat). John Wiley & Sons Inc. New Jersey.
Brown, T. A. 2000. Genomes. 2nd edition. Department of Biomolecular Sciences, UMIST. Manchester. UK
Greenspan, F.S dan Baxter, J.D. 1994. Basic and Clinical Endocrinology. Appleton and Lange. USA.
Kusnaidi. 2010. Substansi Genetika. Direktori FPMIPA UPI. Bandung.
Lucinus, J. 2003. Induksi Genetika Molekular Virus. JKM vol. 3 No. 1. UK. Maranatha. Bandung.
Murray, R.K., D.K. Granner, P.A. Mayes and V.W. Rodwell. 2000. Biokimia Harper. Edisi 25. Buku Kedokteran. EGC. Jakarta.
Soedigdo, P. 1973. Tinjauan Ulang Mengenai Biokimia DNA Dan RNA Serta Biosintesa Protein. Proceedings ITB Vol. 7, No. 2. ITB. Bandung.
Yuwono. 2013. Bioinformatika: Sebuah Pengantar. Fakultas Kedokteran Universitas Sriwijaya. Palembang.
Yuwono, Tribowo. 2005. Biologi Molekular.Erlangga. Jakarta.