TINJAUAN PUSTAKA
Tomato infectious chlorosis virus (TICV)
Tomato infectious chlorosis virus (TICV) pertama kali ditemukan pada tahun 1993 di daerah Irvine Orange, California. Pengamatan pertama kali dilakukan oleh Bill Glover (Crops Production Service-Riverside menunjukkan adanya gejala serangan TICV pada pertanaman tomat). Tanaman yang sakit ini menunjukkan gejala menguning pada bagian di antara tulang daun (interveinal yellowing) dan nekrosis. Survei untuk mengetahui kejadian penyakit yang disebabkan oleh TICV dilakukan di daerah dekat Irvine dan sebelah selatan Irvine pada musim semi tahun 1994. Survei penyakit ini dilakukan kembali di Orange, San Diego dan Carlsbad. Pada bulan Juli 1994 Dr. Bryce Falk menemukan gejala serangan TICV yang sama pada pertanaman tomat di daerah Yolo, California (pertanian organik Universitas California). TICV diisolasi dari pertanaman tomat di lapang dan rumah kaca di kampus Davis. Penyakit kemudian ditemukan di pembibitan tomat komersial rumah kaca, 15 mil dari Davis bagian utara dan California Tengah (daerah San Benito) (Duffus et al. 1996). Penyakit ini menyebar dan menimbulkan kerugian yang sangat besar di Negara penghasil tomat seperti Yunani (Dovas et al. 2002), Italia dan Jepang (Hartono et al. 2003), Taiwan (Tsai et al. 2004) dan Spanyol (Font et al. 2004).
Survei lapang terhadap penyakit ini telah dilakukan di Indonesia pada tahun 2005 sampai awal 2006, yaitu di daerah Magelang, Jawa Tengah dan Purwakarta, Jawa Barat. Penyakit yang disebabkan TICV ini sering disebut dengan penyakit ungu oleh petani di Magelang. Rata-rata intensitas penyakit ini di Magelang mencapi 30% sampai 80% (Hartono dan Wijonarko 2007). Serangan TICV telah ditemukan di dataran tinggi di Kecamatan Cikajang, Kabupaten Garut, Jawa Barat (Fitriasari 2010).
TICV adalah salah satu anggota Genus Crinivirus. Crinivirus berasal dari kata ‘crinis’ (Bahasa Latin, yang artinya rambut) yang berarti ‘virus yang partikelnya tampak seperti benang yang sangat panjang’. Virion terdiri atas kapsid, kapsidnya tidak punya envelope, kapsid atau nukleokapsid memanjang
dengan simetri helix. Menurut Wisler et al. (1996) partikel virus TICV, yang dilihat pada siapan murni hasil ekstraksi tanaman tomat sakit, berbentuk seperti benang (threadlike), memanjang (filamentous), dan lentur (flexuous). Partikel TICV memiliki panjang 850 – 900 nm (Duffus et al. 1996, Liu et al. 2000).
Gambar 1 Partikel TICV berbentuk seperti benang (threadlike), memanjang (filamentous) dan lentur (flexuous) (Liu 2000).
Genom TICV bersegmen (segmented). TICV mempunyai dua genom (bipartite), positif sense. Single stranded (ss) RNA, yaitu RNA 1 (7.8 kb) dan RNA 2 (7.4 kb). Pada RNA genom tersebut terdapat beberapa open reading frame (ORF) yang menyandi beberapa protein struktural dan non struktural. RNA 1 mengandung ORF yang mengkode dua jenis protein yang terlibat dalam replikasi virus, sedangkan RNA 2 mengandung ORF untuk sebuah protein kecil yang hidrofobik (small hydrophobic protein), heat shock protein 70 homologue (HSP70h), sebuah protein berukuran sekitar 60 kDa, dan dua jenis protein mantel yaitu main capsid protein (CP) dan minor capsid protein (CPm) (Wintermantel et al. 2009).
Gambar 2 Organisasi genom TICV (Wintermantel et al. 2009).
Infeksi TICV pada tanaman tomat menyebabkan daun-daun tomat klorosis, yaitu menguning di antara tulang daun (interveinal yellowing) yang berasosiasi dengan berkurangnya kemampuan fotosintesisnya. Pada perkembangan selanjutnya daun-daun menjadi rapuh (leaf brittleness), mengalami nekrotik pada beberapa bagian dan warna bagian yang nekrotik menjadi merah keunguan (bronzing), kebugaran (vigor) tanaman menjadi sangat berkurang, dan apabila menghasilkan buah maka ukurannya jauh lebih kecil dari normal dan proses pematangannya terganggu, serta mudah gugur (early senescence) sehingga sangat menurunkan bahkan meniadakan nilai ekonomi tanaman yang terinfeksi (Duffus et al. 1996; Dalmon et al. 2005).
TICV tidak dapat ditularkan secara mekanis, tetapi dapat ditularkan dengan serangga vektor (kutu kebul rumah kaca (Trialeurodes vaporariorum Westwood.) (Hemiptera: Aleyrodidae)) secara semipersisten. Berdasarkan ketidakmampuan virus tumbuhan melakukan sirkulasi dalam vektor, virus semipersisten sama non-persisten. Sebaliknya, berdasarkan kemampuan penularannya virus semipersisten lebih menyerupai virus persisten, yaitu membutuhkan waktu yang relative lama. Menurut Wisler et al. (1998a) TICV memiliki periode persistensi selama 4 hari. Periode makan akuisisi di atas 48 jam. TICV dapat ditularkan dengan waktu yang terbatas antara 1-9 hari tergantung dari virusnya.
TICV memiliki inang yang luas. TICV mampu menyerang 26 spesies dari delapan keluarga yang berbeda. Inang TICV mencakup beberapa tanaman penting
yang meliputi: famili Solanaceae (Tomat (Lycopersicon esculentum Mill., tomatillo (Physalis ixocarpa Brot.), Physalisalkekengi L., P. floridana. Rybd., P. wrightii Gray., dan kentang (Solanum tuberosum L.), Nicotiana benthamiana Domin., N. clevelandii Gray., N. glauca Graham., petunia (Petunia hybrida Vilm.)) ; Chenopodiaceae (Chenopodium capitatum L., C. murale L.) ; Compositae (artichoke (Cynara scolymus L.), Cynara scolymus L., lettuce (Lactuca sativa L.), Picris echioides L., Senecio vulgaris L., Sonchus oleraceus L., Zinnia elegans Jacq) ; Cruciferae (Capsella bursa-pastoris (L.) Medic)) ; Geraniaceae (Erodium cicutarium (L.)L’Her., Geranium dissectum L.) ; Leguminosae (Trifolium subterraneum L.) ; Malvaceae ( Anoda cristata (L.) Schlecht.) ; Umbelliferae (Conium maculatum L.) (Duffus et al. 1996; Wisler et al. 1996; Li et ai. 1998).
Deteksi TICV pada tomat sangat diperlukan dalam strategi pengendalian. Seiring dengan pesatnya kemajuan pada bidang bioteknologi, metode deteksi virus tumbuhan juga berkembang sangat cepat. Teknik RT-PCR dikembangkan untuk melakukan analisis terhadap molekul RNA hasil transkripsi yang terdapat dalam jumlah sangat sedikit di dalam sel. Oleh karena PCR tidak dapat dilakukan dengan menggunakan RNA sebagai cetakan, maka terlebih dahulu dilakukan proses transkripsi balik (reverse transcription) terhadap molekul RNA sehingga diperoleh molekul cDNA (complementary DNA). Molekul cDNA tersebut kemudian digunakan sebagai cetakan dalam proses PCR. Teknik RT-PCR ini sangat berguna untuk mendeteksi ekspresi gen, untuk amplifikasi RNA sebelum dilakukan kloning dan analisis, maupun untuk diagnosis agensia infektif maupun penyakit genetik (Yuwono 2006).
Teknik RT-PCR memerlukam enzim transkriptase balik (reverse transcriptase). Enzim transkriptase balik adalah enzim DNA polymerase yang menggunakan molekul RNA sebagai cetakan untuk menyintesis molekul DNA (cDNA) yang komplementer dengan molekul RNA tersebut. Beberapa enzim transkriptase balik yang dapat digunakan antara lain mesophilic viral reverse transcriptase (RTase) yang dikode oleh Avian myoblastosis virus (AMV) maupun oleh Moloney murine leukemia virus (M-MuLV), dan Tth DNA polymerase. RTase yang dikode oleh AMV maupun M-MuLV bersifat sangat prosesif dan
mampu menyintesis cDNA sampai sepanjang 10 kb, sedangkan Tth DNA polymerase mampu menyintesis cDNA sampai sepanjang 1-2 kb (Yuwono 2006).
Teknik deteksi TICV dengan RT-PCR telah dilakukan oleh para peneliti menggunakan beberapa primer. Hartono & Wijonarko (2007) dan Jaquemond (2008) telah melakukan deteksi TICV dengan teknik RT-PCR menggunakan primer HSP-70 h, Fitriasari (2010) mendeteksi TICV dengan primer CP, serta Andini (2011) dan Nurulita (2011) dengan primer CPm.
Teknik serologi juga merupakan salah satu cara deteksi dan identifikasi suatu patogen dalam suatu inang. Teknik serologi dengan hibridisasi dot blot menggunakan label digoxigenin TICV probe cRNA, ELISA (Enzyme linked immunosorbent assay), western blot telah dikembangkan dan digunakan untuk mendeteksi TICV pada tanaman tembakau (Nicotiana clevelandii), Physalis wrightii, dan tomat di laboratorium (Duffus et al. 1996; Wisler 1996, dan Li et al. 1998). Uji serologi merupakan pengujian berdasarkan sifat protein suatu virus. Pada dasarnya uji serologi adalah suatu uji yang memerlukan antigen dan antibodi, yang kemudian reaksi akan terjadi antara antigen dan antibodi dalam suatu substrat (Bos 1994 ; Wahyuni 2005).
Prinsip dari uji serologi adalah reaksi spesifik antara antibodi dengan antigen yang membentuk kompleks antigen-antibodi. Antibodi yang digunakan dalam teknik serologi untuk deteksi virus tanaman diperoleh melalui penyuntikan hewan (imunisasi) dengan antigen yang berasal dari hasil pemurnian virus. Darah hewan yang sudah mengandung antibodi terhadap virus yang disuntikkan tersebut kemudian akan diproses lebih lanjut untuk memperoleh antiserum yang spesifik terhadap virus yang bersangkutan. Adanya kompleks antigen-antibodi dapat diperhatikan dengan pengujian in vitro. Teknik serologi yang lazim digunakan untuk diagnosis virus tumbuhan didasarkan pada interaksi antigen-antibodi yang berupa ikatan primer, yaitu ELISA, DIBA, TBIA atau ikatan sekunder yaitu uji presipitasi atau aglutinasi (Akin 2006).
Kloning
Salah satu teknologi DNA rekombinan yang dikembangkan saat ini adalah kloning gen. Menurut Glick & Pasternak (2003) kloning gen adalah sejumlah eksperimen yang bertujuan memindahkan DNA dari satu organism ke organism lain. Eksperimen DNA rekombinan secara umum meliputi: (1) ekstraksi DNA sisipan dari organisme donor, (2) pemotongan dan penyambungan secara enzimatis ke DNA vektor untuk membentuk molekul DNA rekombinan baru, (3) pemindahan hasil konstruksi vektor kloning-DNA sisipan ke dalam suatu sel inang dan pemeliharaan di dalam sel tersebut, dan (4) penyeleksian sel-sel inang yang membawa konstruksi DNA.
Prinsip dari ekstraksi DNA dalam proses kloning adalah menghancurkan dinding sel, baik secara mekanis atau enzimatis; melisis sel dengan menambahkan deterjen (seperti: SDS; membersihkan debris sel menggunakan pelarut organik fenol dan chloroform-isoamilalkohol; dan mengendapkan DNA dari lisat jernih dengan menambahkan etanol dan garam natrium (Old & Primrose 2003).
DNA sisipan dan DNA vektor dipotong menjadi fragmen linear. Pemotongan DNA merupakan kerja enzim restriksi yang bersifat spesifik sehingga menghasilkan DNA dengan potongan unik, baik berujung tumpul (blunt-end) ataupun lancip (sticky-end). Bakteri menghasilkan enzim yang menghancurkan DNA fag sebelum fag ini sempat mengadakan replikasi dan mengarahkan sintesis partikel fag baru. DNA bakteri sendiri terlindung dari enzim ini, hal ini dikarenakan DNA mempunyai gugus metil tambahan yang menghalangi kerja degradatif enzim. Enzim-enzim degradatif ini disebut endonuklease restriksi dan disintesis oleh banyak spesies bakteri. Jenis-jenis enzim restriksi antara lain: Hindlll, Kpnl, Sacl, BamHl, spel, BstXl, EcoRl, EcoV,Notl, Xhol, Nsil, Xbal dan Apal (Brown 2003; Glick & Pasternak 2003). Penyambungan DNA sisipan dengan DNA vektor dilakukan oleh enzim ligase. Konstruksi DNA sisipan-vektor plasmid ditransfer ke sel inang melalui proses transformasi. Prinsip transformasi adalah membuat suatu kondisi yang mempengaruhi sel hidup sehingga dapat menarik dan membiarkan molekul DNA
asing masuk kedalam sel melalui membran sel dari lingkungannya (sel kompeten). Sel kompeten dibuat dengan menurunkan suhu pertumbuhan sel beberapa lama, lalu memberikan kejutan panas. Kemungkinan DNA asing masuk kedalam sel menjadi lebih besar jika pada lingkungannya terdapat ion-ion divalen Ca2+ dan Mg2+. Suatu inang yang baik hendaknya memenuhi prasyarat: pertumbuhan cepat, non patogenik, mampu menangkap molekul DNA dan stabil dalam kultur memiliki enzim yang sesuai untuk replikasi vektor rekombinan, mempunyai informasi genetik selengkap mungkin, dan mempunyai genotip spesifik untuk efektifitas hasil kloning (Glick & Pasternak 2003)
Sistem inang E.coli popular digunakan. Galur E.coli DH5α adalah E.coli yang dimutasi pada bagian lacZ (lacZ∆M15) sehingga dapat dimanfaatkan sebagai penseleksi transforman, Jika galur ini ditransformasikan oleh plasmid yang membawa daerah regulator operon lac yaitu gen penyandi ß-galaktosidase- dan suatu segmen pendek DNA penyadi ujung animo terminal plasmid tersebut berkombinasi dengan produk ß-galaktosidase tidak lengkap yang dihasilkan galur lacZ∆M15, menghasilkan ß-galaktosidase yang fungsional. Peristiwa penggabungan potongan protein lacZ menjadi lacZ fungsional ini disebut komplementasi-α. Enzim ß-galaktosidase fungsional ini dapat diinduksi oleh IPTG. Fenotip ini dapat diamati sebagai warna biru yang dihasilkan dari reaksi dengan substrat kromogenik X-gal (5-bromo-4chloro-3indoly-ß-D-galactoside). Telah dirancang tepat pada bagian hilir lacZ suatu multiple cloning region atau multiple cloning sites (MCS), yaitu suatu daerah sempit sebagai situs penyisipan suatu fragmen DNA. Jika DNA terklon pada daerah tersebut, maka aktivitas fungsional lacZ di plasmid terganggu, sehingga tidak dihasilkan ß-galaktosidase yang fungsional, akibat substrat tidak bereaksi menghasilkan warna biru. Prinsip seleksi koloni biru putih bermanfaat untuk membedakan transforman dengan koloni lainya (Glick & Pasternak 2003). Seleksi transforman hanya menggambarkan masuk tidaknya konstruksi DNA ke dalam inang. Untuk membedakan rekombinasi yaitu koloni yang membawa konstruksi DNA dengan plasmid non rekombinasi perlu dilakukan uji ekspresi klon gen pada media tertentu.
Keberhasilan transformasi dipengaruhi oleh: jenis plasmid yang digunakan, suhu, jumlah dan ukuran DNA, lama perlakuan, adanya enzim nuclease pada sel inang, lama dan cara pemberiannya kejutan panas, spesifitas panas, kekuatan ion, konformasi dan konsentrasi DNA. Untuk menghindari religasi vektor plasmid maka alkalin fosfatase dapat digunakan sehingga tidak muncul transforman yang tidak mengandung insert (Glick & Pasternak 2003).
Ekspresi Gen
Proses ekspresi gen merupakan proses transformasi informasi genetik melalui transkripsi dan translasi, untuk pembentukan protein atau enzim. Protein dan enzim sangat penting dalam proses metabolisme, sehingga ekspresi gen sebenarnya merupakan proses pengendalian metabolisme oleh gen (Jusuf 2009).
Secara umum dikenal dua sistem regulasi ekspresi gen, yaitu regulasi positif dan negatif. Regulasi ekspresi gen melibatkan suatu operon lac. Operon lac adalah operon yang dibutuhkan dalam transpor dan metabolisme dari laktosa di E.coli. Operon ini diregulasi oleh berbagai faktor seperti adanya glukosa dan laktosa. Gen struktural pada operan lac tersebut baru akan aktif bila ada induksi dari laktosa. Sistem regulasinya terjadi pada tahapan transkripsinya karena energi yang diperlukan akan menjadi lebih sedikit dan efisien. Bila tidak ada laktosa, gen lacI akan menghasilkan protein represor yang mengikat operator lac dan mencegah terjadinya transkripsi karena enzim RNA polimerase tidak lagi dapat melekat di situs tersebut. Akan tetapi, saat laktosa ditambahkan ke dalam mediumnya, represor LacI akan terlepas karena terikat pada alolaktosa lalu transkripsi ketiga gen struktural akan berjalan (Kimball 2006).
Setiap protein rekombinan yang diekspresikan pada E. coli dapat mengganggu dalam fungsi sel secara normal, dan bahkan ada yang beracun bagi bakteri. Vektor plasmid sebagai pembawa DNA sisipan adalah molekul DNA yang telah terkarakterisasi dengan baik, yang memungkinkan introduksi molekul DNA rekombinan ke sel inang yang sesuai, serta memungkinkan bertahan stabil dalam sel iang. Sebuah vektor plasmid harus memiliki fungsi (1) origin of replication (ori) dan gen-gen lain yang memungkinkan molekul ini bereplikasi
sebagai elemen ekstra kromosomal, (2) penanda seleksi, biasanya merupakan gen-gen penyandi resistensi terhadap senyawa toksik, seperti antibiotik, dan (3) situs-situs enzim restriksi unik sebagai situs kloning sisipan DNA. Plasmid yang berkualitas tinggi biasanya berukuran kecil, berbentuk sirkular, dan mempunyai banyak jumlah kopi (Glick & Pasternak 2003).
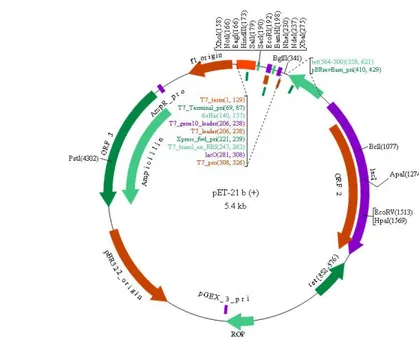
Salah satu pendekatan untuk mengendalikan ekspresi adalah dengan menggunakan vektor ekspresi yang mengandung T7 lac promoter (Studier et al. 1990). Sistem pET adalah alat ekspresi protein yang kuat, karena dapat mengatur ekspresi protein dengan T7 / T7 lac promoter, pLysS atau host pLysS E (Novagen 2003). Vektor pET-21b merupakan vektor ekspresi yang memiliki promoter T7, yang semakin optimal dengan adanya elemen operator yang mengandung runutan operator lac yang mampu meningkatkan ikatan repressor lac dan memastikan rendahnya represi promoter T7.
pET-21b berukuran sekitar 5.4 kb yang memiliki promoter T7, lacO, synthetic ribosome-binding site (RBS), ATG (start codon), runutan 6xhis-tag, multiple cloning site (MCS) dan stop codons (Gambar 5). Vektor ini memiliki situs yang resisten terhadap ampisilin. Plasmid pET-21b dapat ditransformasi dalam sel E. coli BL21(DE3)pLysS. Ekspresi protein diinduksi dengan penambahan isopropyl-thio-D-galactoside (IPTG).
Vektor pET-21b menghasilkan 6xhis-tag pada ujung C dari protein yang terekspresi. His-tag ini memudahkan dalam proses purifikasi karena afinitasnya terhadap resin nickel-nitrilotriacetic (Ni-NTA) (Qiagen 2003). pET-21b mempunyai kelebihan antara lain: mampu mengatur transkripsi gen target, hanya membutuhkan induser dengan konsentrasi yang kecil untuk dapat mengekspresikan gen target, dan hampir semua sel dari gen target terekspresi (Novagen 2003).
Menurut Glick & Pasternak (2003), penyisipan gen dalam suatu vektor tidak memberikan jaminan bahwa gen yang bersangkutan akan diekspresikan. Proses-proses dalam ekspresi suatu protein antara lain: (1) transkripsi, (2) translasi, (3) proses proteolitik dan degradasi, (4) lokalisasi seluler, (5) pelipatan protein, dan (6) pertumbuhan sel.
Laju ekspresi gen asing sangat tergantung pada karakteristik organisme inang yang digunakan. Pada umumnya produksi protein rekombinan masih menggunakan E.coli sebagai inang. Keunggulan produksi protein rekombinan dengan E. coli adalah produksi cepat dan murah, informasi genetik, karakteristik biologi molekuler, biokimia, dan fisiologinya paling banyak diketahui dan diteliti. Namun demikian, penggunaan sistem inang dengan E. coli mempunyai kelemahan. Kelemahan tersebut antara lain: (1) Sinyal transkripsi dan translasi spesies lain tidak dikenali dengan baik oleh inang E. coli, sehingga ekspresi gen-gen heterolog di E. coli lemah, (2) sulit mempelajari fungsi gen-gen dengan lintasan metabolic dan pengaturan yang tidak terdapat di E. coli, seperti degradasi hidrokarbon, dan (3) kemungkinan toksisitas dari produk-produk gen-gen heterolog terhadap sel E. coli. Selain hal-hal tersebut di atas, ada masalah serius pada ekspresi protein rekombinan pada E. coli, yaitu degradasi protein produk secara cepat dan seringkali protein rekombinan terakumulasi dalam sel inang dalam bentuk agregat kompak, bersifat inaktif tak larut, yang disebut badan inklusi (inclusion bodies). Hal ini terjadi akibat keterbatasan E. coli membentuk struktur tiga dimensi protein secara benar dalam proses pelipatan pasca translasi (Glick & Pasternak 2003).
Ekspresi gen pada E. coli untuk produksi antibodi suatu virus tanaman telah dilakukan oleh Kubota et al. (2011) untuk membuat antibodi terhadap Cucurbit
chlorotic yellow virus (CCYV), Fajardo et al. (2007) Grapevine leafroll associated virus 3 (GLRaV-3), Abouzid et al. (2002) terhadap empat Begomovirus, yaitu Bean golden mosaic virus (BGMV) isolat Brazil, Cabbage leaf curl virus (CabLCV), Tomato yellow leaf curl virus (TYLCV), dan Tomato mottle virus (ToMV), Cotillon et al. (2005) Cucurbit yellow stunting disorder crinivirus (CYSDV), dan Nickel et al. (2004) Apple stem grooving virus (ASGV). Keunggulan penyediaan antibodi dengan metode ini antara lain: protein yang dihasilkan bersifat spesifik sehingga tidak bereaksi terhadap protein tanaman, dapat tersedia antigen dalam jumlah yang mencukupi setiap saat apabila diperlukan untuk produksi antiserum, (Cotillon et al. 2005).