Brain Research 888 (2001) 176–179
www.elsevier.com / locate / bres
Short communication
Inhibition of astrocyte TNF
a
expression by extracellular potassium
*
Hee-Yoon Chang, Liwei L. Hua, Anna Morgan, Sunhee C. Lee
Department of Pathology(F-717), Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY 10461, USA
Accepted 10 October 2000
Abstract
TNFa and IL-6 are cytokines of great interest, given the numerous biological activities and the documented expression in several central nervous system (CNS) pathologies. In this report, we have examined cultures of IL-1- or IL-1 / IFNg-activated human fetal astrocytes as a model to study mechanisms of cytokine regulation in the inflamed CNS. Since one of the major functions of astrocytes is
1
spatial buffering of K ions, we examined the effect of high extracellular KCl on astrocyte cytokine expression by ribonuclease protection
1
assay and ELISA. Results demonstrate that astrocyte TNFaproduction was potently inhibited by K with 44 and 89% inhibition at 25 and 55 mM K1, respectively. In contrast, astrocyte IL-6 inhibition required higher concentrations of K1 ($75 mM). These results demonstrate a novel role for astrocyte potassium channel activity in modulation of glial cytokine production. 2001 Elsevier Science B.V. All rights reserved.
Theme: Endocrine and autonomic regulation
Topic: Neural-immune interactions
Keywords: Brain; Hyperkalemia; Cytokine; Inflammation
Astrocytes play an important role in the maintenance of migration into the CNS parenchyma [2,8,9,14]. This is an optimal microenvironment for neuronal growth and pertinent to inflammatory diseases of the CNS such as
1
functional activity. For example, excess K ions excreted multiple sclerosis and HIV-1 encephalitis, in which as-by overly active neurons into the extracellular space is trocyte-activating cytokines such as IL-1, TNFa, and IFNg
1
shunted by astrocytes to an area of lesser K -concen- have been shown to exist. Astrocytes are also important tration, a process often referred to as ‘spatial buffering’ sources for cytokines such as TNFa and IL-6 in the CNS. [15]. In this manner, astrocytes may modulate neuronal Because one of the characteristics of diseased CNS is its excitability and diminish spreading depression. This role change in ionic environment [4,6], we sought to determine may be of particular importance in pathological conditions if hyperkalemic conditions could modulate inflammatory such as ischemia, hypoxia or epileptic seizures, where the activation of astrocytes. Our earlier study showed that
1
extracellular K concentration can rise to 80 mM from the potassium and potassium channel inhibitors could modu-3.5 mM observed under resting conditions [16]. Recent late the expression of inducible nitric oxide synthase studies have shown the presence of voltage-gated ion (iNOS) in astrocytes [13]. In this report, we demonstrate channels in astrocytes and their important role as that potassium is a potent inhibitor of astrocyte TNFa modulators of astrocyte activation and signaling [1]. production.
Mounting evidence has led to the emerging understand- Human fetal astrocytes were isolated and cultivated as ing of astrocytes as immunomodulatory cells. As an described [10]. Briefly, cerebral tissue from second trimes-integral part of the blood–brain-barrier, astrocytes recog- ter fetal abortuses were dissociated by trituration and nize inflammatory cells and cytokines and mediate their enzymatic digestion, and plated as mixed cultures compris-ing astrocytes, neurons and microglia [10]. At 2 weeks, culture media containing microglia were removed, and the remaining adherent cells were subjected to trypsinization *Corresponding author. Tel.: 11-718-430-2666; fax: 1
1-718-430-and replating. 8867.
E-mail address: [email protected] (S.C. Lee). This procedure was repeated every 2 weeks until highly
H.-Y. Chang et al. / Brain Research 888 (2001) 176 –179 177
enriched astrocytes (.99% glial fibrillary acidic protein: GFAP1) were obtained. Culture medium was DMEM (5 mM KCl, 110 mM NaCl, 44 mM NaHCO ) with 4.5 g / l3 of glucose, 4 mM ofL-glutamine, and 25 mM of HEPES buffer (from Life Technologies: Bethesda, MD: Catalogue
[11995) supplemented with 5% FCS (Gemini: Calabasas,
CA), penicillin (100 U / ml) / streptomycin (100 mg / ml) and fungizone (0.25mg / ml). High potassium medium with normal osmolarity was prepared by purchasing a custom-made DMEM identical to the astrocyte media described above, but without KCl or NaCl (Life Technologies) and then adding back desired amounts of KCl and NaCl [13].
1 1
The concentrations of K and Na in each medium were determined with ion-specific electrodes (Multichannel Analyzer: Roche Hitachi [ 914) and the osmolarity by
freezing point depression.
To determine the expression of cytokine mRNA by astrocytes, ribonuclease protection assay (RPA) was per-formed [7]. Highly enriched astrocytes were stimulated with 10 ng / ml each of IL-1b6IFNg (PeproTech: Rocky Hill, NJ) for 16 h, then RNA was extracted by TRIZOL (Molecular Research Center, Cincinnati, OH). Ten (10)mg of total RNA was analyzed by RPA using a custom-made probe set from PharMingen following the manufacturer’s instructions. Densitometry was performed using Ambis Imagequant software. For ELISA, triplicate astrocyte cultures in 96 well plates were stimulated with cytokines for 16 h (for TNFa) or 48 h (for IL-6), then cytokine levels in culture supernatants were measured by commer-cial ELISA kits (Immunotech). The concentrations of stimulating cytokines and duration of stimulation were determined based on our previous studies [11].
To determine if high potassium media induced astrocyte toxicity, cell viability was determined by the lactate dehydrogenase assay. Mild cytotoxicity (|15%) was
in-1
duced at 100 mM K but not at lower concentrations, as
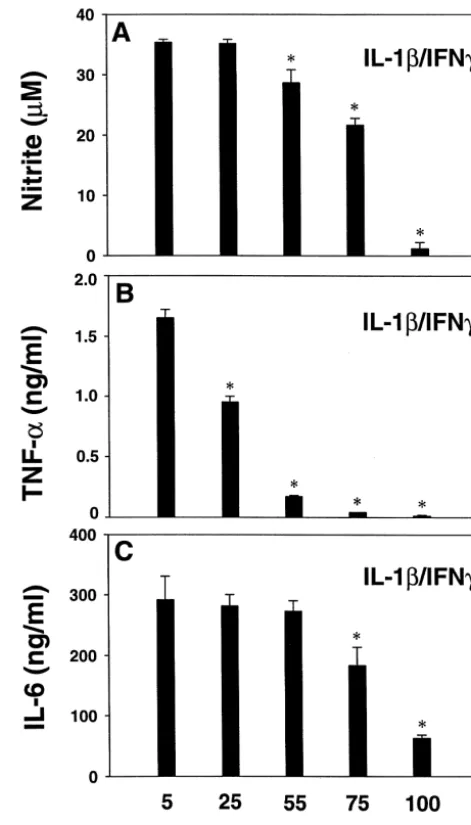
Fig. 1. Effect of high KCl on IL-1b/ IFNg-induced astrocyte TNFa, IL-6 reported [13]. and nitrite production. Human fetal astrocyte cultures were treated with The results of cytokine assays are illustrated in Figs. 1 10 ng / ml each of IL-1band IFN-gin triplicates in medium containing 5 (control), 25, 50, 75, or 100 mM KCl, then levels of TNFa (16 h) and and 2. The results of nitrite assays are shown for
com-IL-6 (48 h) were measured by ELISA. For comparison, nitrite levels in parison [13]. In human fetal astrocytes, maximal induction
the 48 h culture supernatants were determined by the Griess reaction, as of cytokines and nitrite required stimulation with IL-1 /
described [13]. Data are mean6S.D. and are representative of three
1
IFNg, as reported [3,11]. We tested whether high extracel- independent experiments. * P,0.05 vs. 5 mM K by Student’s t-test. lular KCl could modulate IL-1 / IFN-g- (Fig. 1) or IL-1- The results demonstrate a dose-dependent inhibition of astrocyte TNFa
1 1
production by K . Inhibition of IL-6 required higher K . (Fig. 2) induced cytokine production in astrocytes. The
1
results indicate that high extracellular K inhibited IL-1 / IFNg-induced astrocyte TNFaproduction in a
dose-depen-dent manner, with approximately 40, 90, and 98% inhibi- IL-1b-induced TNFa was inhibited dose-dependently at
1 1
tion at 25, 55, and 75 mM KCl (P,0.05 vs. 5 mM K : 25, 55, and 75 mM K , similar to that observed with
1
Fig. 1B). IL-6 was also inhibited by K , but only at 75 and IL-1 / IFNg stimulation (Fig. 2B). In contrast,
IL-1b-in-1
100 mM (Fig. 1C). Nitrite was inhibited by KCl at 55 mM duced IL-6 levels were increased at 25, 55 and 75 mM K
1
or higher (Fig. 1A), as reported [13]. We further tested but reduced at 100 mM K (Fig. 2C). IL-1-induced nitrite
1 1
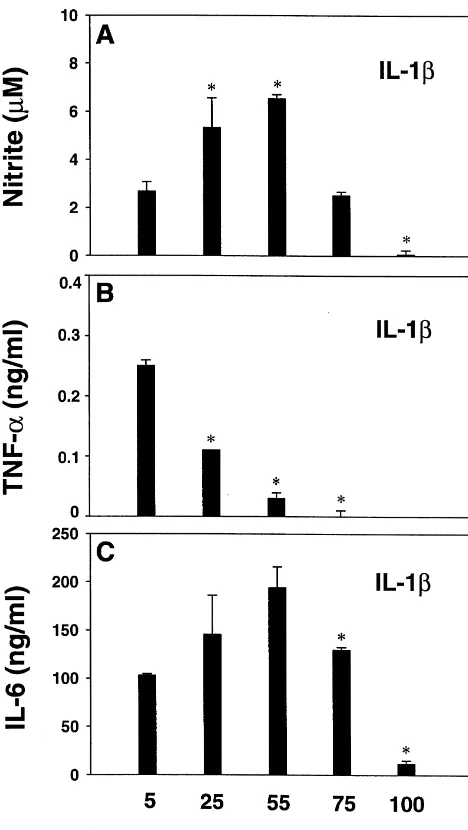
whether K exerted similar inhibitory effects on cytokine levels were also increased at 25 and 55 mM K , as induction by IL-1b alone (Fig. 2). reported [13]. Thus, both IL-1- or IL-1 / IFNg-induced Stimulation of astrocytes with IL-1balone induced high astrocyte TNFa production was inhibited at physiological nanogram amounts of IL-6 but picogram amounts of TNFa levels of hyperkalemic conditions, while IL-6 was not
1
178 H.-Y. Chang et al. / Brain Research 888 (2001) 176 –179
TNFa mRNA expression (n53), while inhibiting protein expression (Fig. 1).
In this report, we sought to determine the influence of neural molecules on astrocyte cytokine expression, to gain insight into how the neural environment may affect astrocyte activation in vivo. One of the important functions of astrocytes is spatial buffering of potassium. Excessive extracellular levels of potassium locally produced under physiological and pathological conditions can affect as-trocyte function. We demonstrated that excess extracellular
1
K in human fetal astrocyte cultures significantly inhibits IL-1b- or IL-1b/ IFNg-stimulated TNFa generation. The results suggest that TNFa production and function can become markedly limited in a high potassium CNS-en-vironment. The results with IL-1 vs. IL-1 / IFNg on IL-6 production are interesting because they demonstrate that alterations in potassium levels can exert dual effects on astrocyte IL-6 expression, while consistently inhibiting
1
TNFa. In this regard, regulation of IL-6 by K resembles that of iNOS, which also varied depending on the stimula-tion (IL-1 vs. IL-1 / IFNg) [13]. It is also noted that
1
whereas 25 mM K significantly and consistently inhibited the level of TNFa protein production (n53), no such inhibition was detected at the mRNA level. These data are
1
consistent with the action of K at multiple steps of TNFa transcription and translation. Furthermore, IL-1 /
IFNg-in-1
duced astrocyte ICAM mRNA was not altered by K , even at extremely high doses, demonstrating that the expression of inflammatory molecules are differentially
1
regulated by K in astrocytes.
The mechanism by which astrocytes involved in CNS
1
injury regulate extracellular K depends on the com-position of ion channels residing in the plasma membrane of these gliotic cells. MacFarlane and Sontheimer [12] have studied an in vitro glial scar model and shown that ‘scarring’ induces a rapid shift in potassium channel Fig. 2. Effect of high KCl on IL-1b-induced astrocyte TNFa, IL-6 and
composition. A reduction in inwardly rectifying potassium nitrite production. Astrocytes were treated as described in Fig. 1 legend,
1
channels (K ) was noted as well, as an increase in K except that stimulating cytokines consisted only of IL-1b. The results IR
1
show that whereas TNFaproduction was potently inhibited by K , IL-6 conductance through delayed rectifiers (K ), shunting D
1 1
and nitrite were elevated by 25 and 55 mM K . *P,0.05 (vs 5 mM K ) potassium out of astrocytes. These findings have implica-by Student’s t-test, or Mann–Whitney Rank Sum Test when Equal
tions for our study on astrocyte TNFa generation. Under
Variance Test failed. 1
normokalemic conditions, the increased shunting of K out of astrocytes through KD channels would conceivably in IL-1 / IFNg-treated cultures required much higher con- provide a condition favorable for TNFa induction. Yet centrations than those required for TNFa inhibition. various pathological conditions associated with elevated
1 1
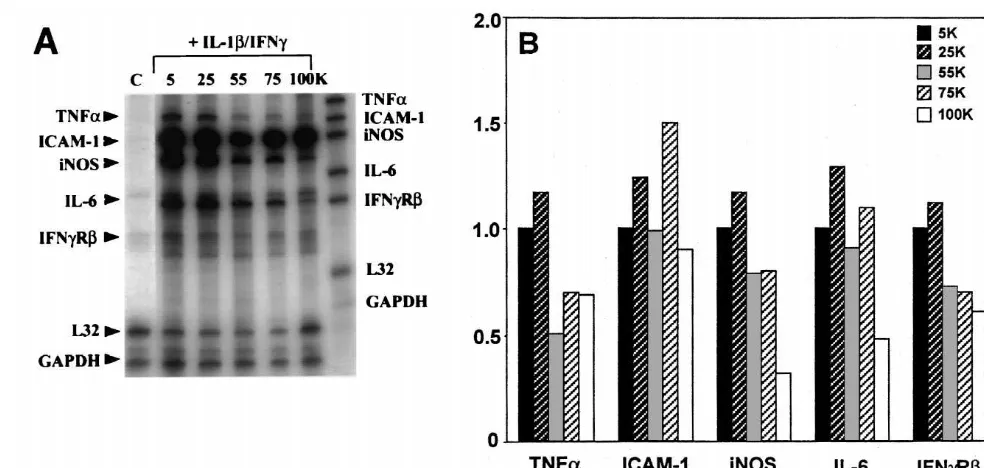
We further performed RPA to determine if K modu- extracellular K would limit KD activity, leading to
1
lates cytokine mRNA expression in astrocytes. A repre- intracellular K accumulation, and subsequent inhibition sentative blot and the densitometric values are illustrated in of TNFa induction. The fact that the extracellular
con-1
Fig. 3A and B, respectively. Messenger RNA for TNFa, centration of K used to inhibit TNFain our experiments intercellular adhesion molecule-1 (ICAM-1), inducible was within the physiological norm implies that a subtle nitric oxide synthase (iNOS), IL-6, and theb-chain of the change in potassium concentrations may indeed affect IFNg receptor (IFNgRb) were induced by IL-1 / IFNg astrocyte TNFa expression.
treatment of astrocytes, and the levels were inhibited by Cytokines TNFa and IL-6 are induced under a number
1
55, 75 and 100 mM K . One exception was mRNA of pathologic conditions of the CNS and are pivotal in the expression for ICAM-1, which remained unchanged with regulation of inflammatory and cytotoxic responses [2,5].
1
ob-H.-Y. Chang et al. / Brain Research 888 (2001) 176 –179 179
Fig. 3. Effect of high KCl on IL-1b/ IFNg-induced astrocyte TNFa, ICAM-1, iNOS, IL-6, and IFNgR mRNA expression. Astrocytes treated for 16 h as described in Fig. 1 legend were analyzed for mRNA expression for TNFa, ICAM-1, iNOS, IL-6 and IFNgRbby a multiprimer RPA kit, as described in the text. A representative blot out of four is shown in (A), and the densitometric ratios to the house-keeping genes (L321GAPDH) are shown in (B). Data are
1
expressed as amounts relative to those obtained with 5 mM K medium. Control (C) denotes unstimulated culture.
adult rat brain cortex during anoxia, Acta Physiol. Scand. 99 (1977) served with IL-6, TNFa appears to exert primarily toxic
412–420. and proinflammatory responses in the CNS. Thus the
[5] S.J. Hopkins, N.J. Rothwell, Cytokines and the nervous system I: differential regulation of TNFa and IL-6 by potassium expression and recognitions, Trends Neurosci. 18 (1995) 83–88. found in our study may be a mechanism by which neural [6] K.A. Hossmann, S. Sakaki, V. Zimmermann, Cation activities in cells interact with immune system components to limit the reversible ischemia of the cat brain, Stroke 8 (1977) 77–81.
[7] L.L. Hua, S.C. Lee, Distinct patterns of stimulus-inducible extent of neural damage. These findings may have
rele-chemokine mRNA accumulation in human fetal astrocytes and vance to the pathogenesis of several CNS disorders,
microglia, Glia 30 (2000) 74–81.
1
especially those in which altered K levels have been [8] S.C. Lee, C.F. Brosnan, Molecular biology of glia: astrocytes, in:
implied. W.C. Russell (Ed.), Molecular Biology of Multiple Sclerosis, John
Wiley, 1997, pp. 71–96.
[9] S.C. Lee, D.W. Dickson, C.F. Brosnan, Interleukin-1, nitric oxide and reactive astrocytes, Brain Behav. Immun. 9 (1995) 345–354. Acknowledgements
[10] S.C. Lee, W. Liu, C.F. Brosnan, D.W. Dickson, Characterization of human fetal dissociated CNS cultures with an emphasis on mi-We are grateful to Wa Shen for tissue culture and Dr. croglia, Lab. Invest. 67 (1992) 465–475.
1 1
Janet Piscitelli for determination of K and Na levels in [11] S.C. Lee, W. Liu, D.W. Dickson, C.F. Brosnan, J.W. Berman, Cytokine production by human fetal microglia and astrocytes: culture media. Tissue was provided by the Human Fetal
differential induction by LPS and IL-1b, J. Immunol. 150 (1993) Tissue Repository at the Albert Einstein College of
2659–2667.
Medicine. This study was supported by MH55477 and the [12] S.N. MacFarlane, H. Sontheimer, Electrophysiologic changes that National Multiple Sclerosis Society Research Grant 2771. accompany reactive gliosis in vitro, J. Neurosci. 17 (1997) 7316–
7329.
[13] A.C.A. Morgan, H.Y. Chang, J.S.H. Liu, L.L. Hua, S.C. Lee, High extracellular potassium modulates nitric oxide synthase expression References
in human astrocytes, J. Neurochem. 74 (2000) 1903–1912. [14] L. Mucke, M. Eddleston, Astrocytes in infectious and immune-[1] B.A. Barres, W.J. Koroshetz, L.L.Y. Chun, D.P. Corey, Ion channel mediated diseases of the central nervous system, FASEB J. 7 (1993)
expression by white matter glia: The type 1 astrocytes, Neuron 5 1226–1232.
(1990) 527–544. [15] R.K. Orkland, J.G. Nicolls, S.W. Kuffler, Effect of nerve impulses [2] E.N. Benveniste, Cytokine actions in the central nervous system, on the membrane potential of glial cells in the central nervous
Cytokine Growth Factor Rev. 9 (1998) 259–275. system of amphibia, J. Neurophysiol. 29 (1966) 788–806. [3] M. Downen, T.D. Amaral, L.L. Hua, M.L. Zhao, S.C. Lee, Neuronal [16] H. Sontheimer, Voltage-dependent ion channels in glial cells, Glia 11
death in cytokine-activated primary human brain cell culture: role of (1994) 156–172. tumor necrosis factor-alpha, Glia 28 (1999) 114–127.