Produktivitas Media LB dan MTM terhadap Aktivitas Enzim
Xilanolitik dari Kultur Campuran Escherichia coli BL21
Rekombinan
Rahmat Eko SanjayaDepartemen Kimia, Fakultas Sains dan Teknologi, Universitas Airlangga, Surabaya 60115, Indonesia
Program Studi Pendidikan Kimia, Fakultas Keguruan dan Ilmu Pendidikan, Universitas Lambung Mangkurat, Banjarmasin, Indonesia
Email: rahmat.eko.sanjaya@gmail.com
Sofijan Hadi
Departemen Kimia, Fakultas Sains dan Teknologi, Universitas Airlangga, Surabaya 60115, Indonesia
Email: sofijanh@yahoo.com
Ni Nyoman Tri Puspaningsih
Departemen Kimia, Fakultas Sains dan Teknologi, Universitas Airlangga, Surabaya 60115, Indonesia
Laboratorium Proteomik, Lembaga Penyakit Tropis, Universitas Airlangga, Surabaya 60115, Indonesia
Email: nyomantri@yahoo.com
Abstrak
Media pertumbuhan merupakan salah satu faktor utama yang menentukan tingkat aktivitas enzim terhadap substrat. Enzim xilanolitik rekombinan dari kultur campuran E. coli BL21/pET-Abfa dan E. coli BL21/pET-XylB dalam media terdefinis i modifikasi (MTM) memberikan nilai aktivitas sebesar 0.094 ± 0.0031 U mL-1 dengan
menggunakan IPTG dan 0.067 ± 0.0201 U mL-1 tanpa menggunakan IPTG. Pada
media Luria Bertani (LB) tanpa menggunakan IPTG, enzim xilanolitik rekombina n dari kultur campuran memberikan aktivitas enzimatik sebesar 0.183 ± 0.0015 U mL -1. Pengujian aktivitas enzim xilanolitik dari kultur campuran pada media MTM tanpa
IPTG, dilakukan pada substrat alami (oat spelt xylan, beechwood xylan, corn cob xylan) dan substrat sintetik (pNP-X dan pNP-A). Aktivitas tertinggi pada substrat alami diperoleh pada substrat oat spelt xylan dengan nilai aktivitas sebesar 0.061 ± 0.0079 U mL-1 dan pada substrat spesifik pNP-X dan pNP-A masing- masing adalah
2.940 ± 0.0630 U mL-1 dan 2.353 ± 0.4131 U mL-1. Hasil ini mengindikasikan bahwa
jenis media pertumbuhan memberikan pengaruh terhadap aktivitas enzim xilanolit ik dari kultur campuran dan nilai aktivitas terhadap substrat pNP-X dan pNP-A mengindikasikan adanya sinergisitas antara α-L-arabinofuranosidas e (E. coli BL21/pET-Abfa) dengan β-xilosidase (E. coli BL21/pET-XylB).
Kata kunci: enzim xilanolitik, Escherichia coli BL21 rekombinan, media LB, media MTM
Abstract
Growth medium is one of the main factors that determine the level of activity of the enzyme to the substrate. Recombinant xylanolytic enzyme from mixed cultures of E. coli BL21/pET-Abfa and E. coli BL21/pET-XylB in defined medium modification (DMM) have mean activity value 0.094 ± 0.0031 U mL-1 using IPTG and 0.067 ±
0.0201 U mL-1 without IPTG. In Luria Bertani (LB) without IPTG, the recombinant
xylanolytic enzyme from mixed cultures provide the enzymatic activity 0.183 ± 0.0015 U mL-1. Activity of recombinant xylanolytic enzyme from mixed cultures tested on
natural substrates (oat spelt xylan, beechwood xylan, corn cob xylan) and synthetic substrates (pNP-X and pNP-A). The highest activity on the natural substrate was obtained on oat spelt xylan with the activity mean value 0.061 ± 0.0079 U mL-1 and
the specific substrate pNP-X and pNP-A, respectively were 2.940 ± 0.0630 U mL-1
and 2.353 ± 0.4131 U mL-1. These results indicate that the type of growth medium
have effect on xylanolytic enzyme activity from mixed cultures and activity mean value against pNP-X and pNP-A indicates a synergy between α-L-arabinofuranosidase (E. coli BL21/pET-Abfa) and β-xylosidase (E. coli BL2/pET-XylB).
Keywords: xylanolitic enzyme, recombinant Escherichia coli BL21, LB medium, MTM medium
I. PENDAHULUAN
Industri berbasis bioteknologi mengalami peningkatan minat dalam beberapa dekade terakhir. Peningkatan minat tersebut disebabkan oleh proses industri yang ramah lingkungan (green industry) dan dapat diperbaharui (renewable). Enzim merupakan salah satu objek dalam kegiatan industri berbasis bioteknologi. Salah satu jenis enzim yang banyak digunakan dalam kegiatan industri adalah enzim degradatif. Enzim hemiselulase memiliki aktivitas degradatif terhadap hemise lulosa yang merupakan salah satu bahan baku dalam kegiatan industri berbasis biomasa. Kandungan hemiselulosa relatif lebih tinggi dibandingkan dengan komponen lain, yaitu sebesar 50% terdapat pada jerami gandum, selulosa (30%) dan lignin (15%) (Horward et al., 2003).
Xilan merupakan struktur utama penyusun hemiselulosa pada kayu keras. Produk hidrolisis xilan dapat dimanfaatkan sebagai sumber xilitol melalui proses fermentasi. Sebesar 50-60% xilitol diperoleh dari fraksi xilan atau 8-15% dari total material biomasa berlignoselulosa (Winkelhausen & Kuzmanova, 1998). Xilanase merupakan golongan enzim yang mendegradasi xilan melalui reaksi hidrolisis ikatan β-1,4 glikosidik menjadi monomer-monomer gula penyusunnya. Komplek enzim ini meliputi endo-β-1,4-xilanase (EC 3.2.1.8), β-xilosidase (EC 3.2.1.37) dan beberapa enzim lain seperti α-L-arabinofuranosidase (EC 3.2.1.55), α-glukuronidase (EC 3.2.1.139), asetil xilan esterase (EC 3.1.1.72), asam ferulat esterase (EC 3.1.1.73) dan asam p-coumaric esterase (EC 3.1.1.x) (Saha, 2003). Masing-masing enzim tersebut memiliki peran spesifik dalam degradasi xilan. Produk hasil degradasi xilan dapat dimanfaatkan sebagai prebiotik, bioetanol, pakan ternak dan xilitol serta produk fermentasi lainnya yang banyak digunakan oleh industri (Wong et al., 1988)
Enzim α-L-arabinofuranosidase memiliki kemampuan untuk produksi arabinosa yang berperan sebagai antigycemic agent, produksi senyawa antimetastatic dan anticarcinogenic, pakan ternak serta bersinergi dengan enzim xilanotik lain untuk industri kertas dan pulp serta bioetanol (Numan & Bhose, 2006; Saha, 2000). Enzim xilanolitik lainnya adalah β-xilosidase yang bersama dengan xilanase lainnya berperan sebagai agen biobleaching kertas dan pulp (Terrasan et al., 2013) serta industri roti dan pembuatan bir (Harris & Ramalingan, 2010; Polizeli et al., 2006). Gen penyandi xilanase termostabil telah berhasil diisolasi dari Geobacillus thermoleovarans IT-08, yang meliputi gen penyandi β-D-xilosidase (GbtXyl43 B, GenBank No. DQ387047), β-D-xilosidase (GbtXyl43A, GenBank No. DQ345777) dan α-L-arabinofuranosidase (abfa51, GenBank No. DQ387046) (Puspaningsih, 2004; Ratnadewi A. I. et al., 2013). Ketiga gen tersebut telah berhasil dikarakterisas i dan diekspresikan masing-masing dalam sistem pET101/D-TOPO dengan sel inang E. coli BL21.
Tim peneliti dari Laboratorium Proteomik Universitas Airlangga, Surabaya, melaporkan bahwa media terdefinisi yang mengandung sumber karbon, nitrogen dan garam anorganik berupa garam fosfat dan sulfat serta dengan modifikasi berupa
penambahan ekstrak bahan alam sebagai pelarut dapat memproduksi enzim α-L-arabinofuranosidase rekombinan (Puspaningsih, 2011). Oleh karena itu, perlu
dilihat perbandingan produktivitas antara media LB dan MTM sebagai langkah awal dalam pengembangan media produksi. Produksi enzim rekombinan tunggal dari satu kultur hanya akan menghasilkan satu jenis enzim dengan jumlah yang relatif besar. Untuk mendapatkan enzim campuran β-xilosidase (XylB) dan α-L-arabinofuranosidase (Abfa) maka proses produksi dapat dilakukan dengan menggabungkan kedua jenis kultur. Selain itu, produksi enzim β-xilosidase dalam E.coli BL21/pET-xylB dan α-L-arabinofuranosidase dalam E. coli BL21/pET-abfa yang dilakukan secara bersamaan dapat meningkatkan efisiensi produksi.
II. METODE
Preparasi inokulum. Satu ose biakan E. coli BL21/pET-XylB dan E. coli BL21/pET-Abfa diambil dari media padat LB dan masing-masing diinokulasikan ke dalam Erlenmeyer 100 mL yang berisi 25 mL media inokulum yang telah ditambahkan 25 µL ampisilin 1 mg/mL. Inokulum diinkubasi dalam shaker incubator (Gerhardf) dengan kecepatan 150 rpm selama 9-10 jam pada suhu 370C. Sebelumnya,
media disterilisasi menggunakan autoclave (TOMY) selama 15 menit pada suhu 1210C.
Perbandingan produktivitas media. Tiga persen (v/v) inokulum hasil pencampuran E. coli/pET-XylB dan E. coli/pET-Abfa (1:1), dimasukkan masing-masing ke dalam Erlenmeyer 250 mL yang berisi 50 mL media produksi MTM dan LB. Sebelumnya, ke dalam masing- masing media produksi ditambahkan 50 µL ampisilin 1 mg mL-1. Kultur diinkubasi pada temperatur 370C dengan kecepatan 150
rpm selama 16 jam. Media MTM dengan IPTG, dilakukan penambahan 50 µL IPTG 1 M pada jam ke 2 inkubasi atau pada OD600 antara 0.4 – 0.6 dan dilanjutkan kembali
inkubasi hingga 16 jam. Setelah 16 jam, kultur dipanen dan dilakukan penentuan aktivitas enzim xilanolitik dengan substrat alami berupa oat spelt xylan. Penentuan produktivitas media dilakukan duplo. Aktivitas enzim xilanolitik hasil produksi dengan media MTM diuji terhadap substrat alami (oat spelt. beechwood, xilan dari tongkol jagung) dan substrat spesifik (pNP-A dan pNP-X). Uji aktivitas enzim xilanolitik terhadap berbagai substrat dilakukan triplo pengukuran.
Isolasi enzim rekombinan. Kultur disentrifugasi dengan kecepatan 3500 rpm selama 25 menit pada temperatur 40C(Hermle Z 400 K). Pelet sel dicuci dengan NaCl 0.9%
dan suspensi pelet disentrifugasi kembali. Pelet sel dilarutkan kembali dalam buffer fosfat sitrat 50 mM pH 6 dan dilisis menggunakan ultrasonikator(JY 92-IIDN). Lisat kemudian disentrifugasi dan supernatan diinkubasi pada suhu 500C dalam penangas
air selama 30 menit untuk menghilangkan protein non termofil. Hasil inkubas i disentrifugasi kembali dan supernatan hasil sentrifugasi merupakan protein xilanolit ik rekombinan.
Uji aktivitas enzim xilanolitik. Aktivitas ekstrak kasar enzim ditentuka n dengan mengukur banyaknya gula pereduksi atau para-nitrofenol yang dihasilkan dari hidrolisis substrat. Seratus mikroliter substrat ditambah 100 μL enzim diinkubasi pada temperatur 600C selama 30 menit. Hasil inkubasi ditambah dengan 600 μL DNS
(Miller, 1959), didihkan dalam penangas air dan dinginkan. Absorbansi dibaca dengan Spektrofotometer UV-Vis (Shimadzu UV-1800)pada λ = 550 nm(Puspanings i h, 2004). Kontrol yang digunakan diperlakukan sama dengan kondisi di atas tanpa ada proses inkubasi.Standar xilosa dibuat dengan berbagai variasi konsentrasi dari larutan stok xilosa 10 mg/mL dan diuji dengan menggunakan metode DNS (Miller, 1959). Satu unit aktivitas enzimdidefinisikan sebagai jumlah enzim yang melepaskan 1 µmo l gula reduksi per menit pada kondisi percobaan.
Unit aktivitas enzim juga ditentukan dengan menggunakan substrat spesifik berupa pNP-A dan pNP-X. Lima puluh mikroliter enzim direaksikan dengan 450 µL substrat spesifik, kemudian inkubasi selama 30 menit pada temperatur 600C. Reaksi
dihentikan dengan menambahkan 50 µL Na2CO3 0,4 M. Absorbansi dibaca pada λ =
405 nm (Puspaningsih, et al., 2008). Satu unit aktivitas didefinisikan sebagai jumla h enzim yang menghasilkan 1 µmol p-nitrofenol per menit pada kondisi percobaan.Standar p-nitrofenol dibuat dengan berbagai variasi konsentrasi dari larutan stok p-nitrofenol 10 mM.
III. HASIL DAN PEMBAHASAN
Media Luria Bertani (LB) merupakan media komplek yang digunakan sebagai media komersil untuk produksi dan pertumbuhan berbagai jenis bakteri, baik bakteri
wild type maupun rekombinan. Media LB yang terdiri atas yeast extract, tripton dan NaCl memberikan yield sel dan produk yang baik untuk banyak spesies bakteri, termasuk E. coli (Sezonovet al., 2007). Bakteri yang ditumbuhkan pada media LB, dipaksa untuk menggunakan asam amino yang terkandung dalam media sebagai sumber utama untuk proses metabolisme. Media LB merupakan media komplek yang kaya sumber nitrogen dan minim sumber karbon yang berupa gula fermentas i. Kandungan gula fermentasi dalam media LB berada pada konsentrasi kurang dari 0.1 mM (Sezonov et al., 2007).
Penggunaan media LB sebagai media produksi untuk skala laboratorium terbukti memberikan hasil yang memuaskan. Akan tetapi, jika digunakan dalam kegiatan industri untuk produksi skala besar, penggunaan media LB tidak ekonomis dan relatif mahal. Umumnya untuk skala besar, produksi menggunakan media terdefinisi (defined medium) dengan komposisi media yang telah diketahui secara pasti. Tim peneliti dari Laboratorium Proteomik Universitas Airlangga telah mengembangkan media alternatif untuk produksi enzim rekombinan dalam sel inang E. coli BL21. Media tersebut merupakan media terdefinisi dengan modifikasi atau media terdefinisi modifikasi (MTM). MTM terdiri atas campuran sumber karbon, sumber nitogen dan garam anorganik serta ekstrak bahan alam sebagai pelarut.
Perbandingan produktivitas media LB dan MTM untuk produksi enzim xilanolitik dari kultur campuran E. coli BL21 rekombinan terlihat seperti pada Tabel1. Penelitian ini juga menentukan pengaruh dari penggunaan induser berupa Isopropil-β-D-tiogalaktosida(IPTG) terhadap aktivitas enzim xilanolitik. Hasil penelitia n menunjukkan bahwa untuk produksi kultur campuran, media LB memberikan hasil berupa aktivitas enzim yang relatif lebih besar dibandingkan dengan produksi menggunakan MTM, meskipun tanpa penggunaan IPTG.
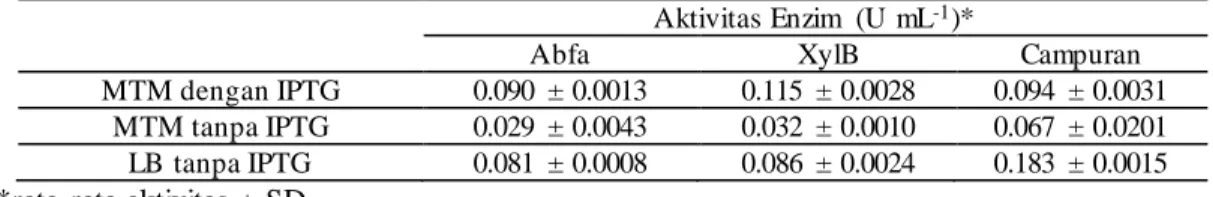
Tabel 1. Aktivitas enzim xilanolitik pada media MTM dan LB
Aktivitas Enzim (U mL-1)*
Abfa XylB Campuran
MTM dengan IPTG 0.090 ± 0.0013 0.115 ± 0.0028 0.094 ± 0.0031 MTM tanpa IPTG 0.029 ± 0.0043 0.032 ± 0.0010 0.067 ± 0.0201 LB tanpa IPTG 0.081 ± 0.0008 0.086 ± 0.0024 0.183 ± 0.0015
*rata-rata aktivitas ± SD
Aktivitas enzim xilanolitik sebesar 0.183 U mL-1 diperoleh dari kultur campuran
dengan menggunakan media LB. Aktivitas ini lebih besar dua kali lipat dibandingk a n dengan aktivitas xilanolitik dari kultur campuran menggunakan media MTM dengan IPTG, yaitu sebesar 0.094 U mL-1. Sementara itu, aktivitas dari kultur campuran
dengan MTM tanpa IPTG menunjukkan nilai yang relatif rendah, yaitu sebesar 0.067 U mL-1. Akan tetapi, jika dilihat dari segi penggunaan induser IPTG dalam media
MTM, penggunaan IPTG tidak memiliki pengaruh yang signifikan.
Penggunaan kultur campuran memiliki tujuan untuk meningkatkan proses hidrolisis substrat akibat kerja sinergis antara enzim α-L-arabinofuranosidase (Abfa)
dan β-xilosidase (XylB). Hasil penelitian menunjukkan bahwa produksi enzim secara berasamaan dalam sebuah kultur campuran memberikan hasil yang relatif lebih tinggi dibandingkan dengan produksi secara individu. Aktivitas enzim Abfa dan XylB dengan media MTM tanpa IPTG berturut-turut adalah 0.029 U mL-1 dan
0.032 U mL-1, lebih kecil dibandingkan aktivitas xilanolitik ketika diproduksi secara
bersamaan. Sehingga produksi enzim yang berasal dari kultur campuran dapat dilakukan.
Aktivitas yang dihasilkan dari kultur campuran dengan menggunakan MTM, baik dengan atau tanpa IPTG, memberikan hasil yang tidak jauh berbeda. Sehingga terlihat bahwa IPTG yang digunakan tidak memiliki pengaruh yang signifik a n terhadap aktivitas enzim dengan media pertumbuhan MTM. Penggunaan media LB sebagai media produksi, meskipun tanpa IPTG, memberikan hasil yang lebih besar dibandingkan dengan MTM, baik dengan maupun tanpa IPTG. Senyawa IPTG memiliki peran sebagai induser yang memberikan efek berupa peningkatan ekspresi atau over ekspresi dari enzim rekombinan yang tersisipi dalam sistem plasmid. Sehingga, meskipun tanpa keberadaan IPTG, sistem ekspresi tetap berjalan dalam keadaan paling dasar (basal level). Selain itu, sistem pET-101/DTOPO menggunak a n promotor dan sistem operon lac untuk ekspresi protein heterolognya. Sistem lac merupakan sistem ekpresi yang tidak terlalu ketat, sehingga tanpa keberadaan IPTG masih dimungkinkan untuk terjadinya ekspresi dalam keadaan dasar. Selain itu, penambahan laktosa yang memiliki kemiripan struktur dengan IPTG dapat menjadi induser alternatif untuk over ekspresi xilanase rekombinan.
Aktivitas enzim yang lebih tinggi dihasilkan dengan menggunakan media produksi LB, meskipun media LB tanpa menggunakan IPTG. Media LB merupakan media kaya sumber nitrogen yang berasal dari asam amino hasil ekstraksi ragi dan susu. Media MTM merupakan media yang menggunakan gula fermentasi sebagai sumber karbon dan garam ammonium sebagai sumber nitrogen. Selain itu, penggunaan ekstrak bahan alam sebagai pelarut menambah sumber karbon dan sumber nitrogen pada media. Keberadaan gula fermentasi yang berlebih dapat menekan sistem ekspresi gen, sehingga secara otomatis berpengaruh terhadap aktivitas enzim. Hal ini dikenal dengan istilah represi katabolit glukosa atau efek glukosa. Konsentrasi gula fermentasi lainnya dalam keadaan berlebih dapat menekan ekspresi protein rekombinan (Eiteman & Altman, 2006; Rinas et al., 1989). Meskipun MTM memiliki nilai aktivitas yang lebih rendah dari pada LB, MTM menjanjik a n sebagai media alternatif untuk produksi enzim rekombinan. Hal ini terlihat dari adanya aktivitas enzim rekombinan yang diproduksi dengan menggunakan media MTM.
Enzim hasil produksi dari kultur campuran dengan menggunanakan media MTM, diuji terhadap beberapa substrat dan hasilnya terlihat seperti pada Tabel 2. Enzim xilanolitik diuji terhadap substrat beechwood xylan, oat spelt xylan, corn cob
xylan, pNP-A dan pNP-X. Terhadap substrat-substrat tersebut, enzim xilanolit ik memberikan hasil yang positif.
Tabel 2. Aktivitas enzim xilanolitik terhadap beberapa substrat
Substrat Aktivitas (U mL-1)*
Beechwood xylan Oat spelt xylan Corn cob xylan pNP-A pNP-X 0.047 ± 0.0102 0.061 ± 0.0079 0.020 ± 0.0057 2.353 ± 0.4131 2.940 ± 0.0630 *rata-rata aktivitas ± SD
Aktivitas enzim terhadap substrat spesifik menunjukkan bahwa, baik Abfa maupun XylB tidak saling menekan dan sama-sama memberikan aktivitas meskipun diproduksi dalam kultur campuran. Hasil ini membuktikan bahwa dalam proses hidrolisis substrat terjadi sinergisitas antara Abfa dan XylB. Sinergisitas tersebut turut menigkatkan kemampuan hidrolisis enzim terhadap substarat komplek lainnya.
Pertumbuhan bakteri merupakan parameter awal untuk mengetahui produktivitas sebuah media produksi. Terdapat perbedaan laju pertumbuhan spesifik, µm, antara pertumbuhan E. coli BL21 dalam media LB dan media MTM. Pertumbuha n E. coli BL21 dengan media MTM memiliki laju pertumbuhan spesifik sebesar 0.795 h-1 dan laju pertumbuhan spesifik pada media LB sebesar 0.249 h-1. Kedua media
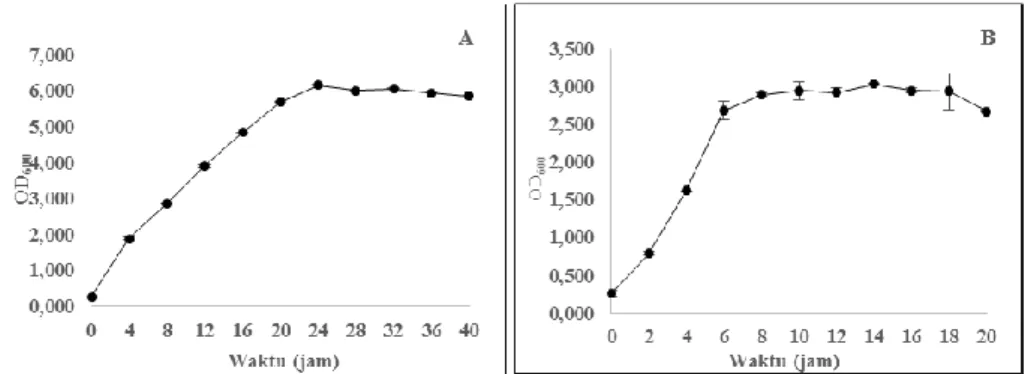
tersebut menunjukkan perbedaan laju pertumbuhan spesifik yang sangat besar. Perbedaan ini disebabkan oleh keberadaan gula pereduksi (gula fermentasi) yang terdapat dalam media MTM. Gula fermentasi dalam konsentrasi yang relatif tinggi dalam media pertumbuhan akan menghasilkan laju pertumbuhan yang tinggi (Luli & Strohl, 1990). Akibatnya, fase logaritmik E. coli BL21 akan berakhir dalam waktu yang relatif singkat. Konsentrasi gula fermentasi yang besar dengan laju pertumbuha n spesifik yang tinggi dalam sistem aerob akan menghasilkan produk samping berupa senyawa organik yang bersifat asam dan dikenal dengan istilah efek Crabtree (Rinas et al., 1989; Doelle et al., 1982). Perbandingan pertumbuhan antara E. coli BL21 pada media MTM dan LB, terlihat seperti pada Gambar 1.
Gambar 1. Kurva pertumbuhan E. coli BL21 rekombinan dengan menggunakan media LB (A) dan media MTM (B).
Asam asetat merupakan produk samping utama yang terbentuk dari proses fermentasi secara aerob menggunakan media yang mengandung gula fermentas i. Asam asetat selain sebagai produk samping proses fermentasi aerob, juga dapat menekan pertumbuhan sel dan ekspresi gen rekombinan (Eiteman & Altman, 2006; Rinas et al., 1989; Tripathi et al., 2009) yang mempengaruhi produktivitas sel E. coli BL21 rekombinan dalam menghasilkan enzim xilanolitik. Terlihat bahwa pada konsentrasi gula fermentasi yang relatif tinggi dalam media MTM (data tidak ditunjukkan), baik pertumbuhan sel maupun aktivitas enzim mengalami penuruna n. Penurunan aktivitas enzim menunjukkan bahwa keberadaan produk samping yang umumnya berupa asetat mempengaruhi proses biosintesis protein yang berakibat pada penurunan aktivitas enzim xilanolitik.
Keberadaan asetat dalam media produksi sebagai produk samping, memberika n peran dalam menghambat pertumbuhan sel dan mempengaruhi ekspresi gen rekombinan. Asetat dalam media produksi dengan konsentrasi 5 g L-1 hingga 10 g L -1 dapat menghambat pertumbuhan sel (Koh et al., 1992; Pan et al., 1987; Luli &
Strohl, 1990) dan menghambat pembentukan protein serta produk rekombinan lainnya (Rinas et al., 1989; Shimizu et al., 1988; Jensen & Carlsen, 1990), bahkan tercatat pada konsentrasi 0.5 g L-1, asetat dapat menghambat pertumbuhan E. coli (Nakano et al., 1997). Pembentukan asetat disebabkan oleh ketidakseimbangan metabolis me glukosa, yaitu keadaan jumlah karbon yang masuk ke dalam sel melebihi tuntuta n untuk proses biosintesis (Gleiser & Bauer, 1981; El-Mansi & Holms, 1989).
Sel E. coli memiliki batas untuk dapat mengkonsumsi semua sumber karbon atau gula yang tersedia dalam media pertumbuhan. Penggunaan gula fermentasi yang berlebih dalam sistem batch, meskipun memberikan hasil yang optimum, menyebabkan produksi kurang efisien dari segi bahan. Oleh karena itu, rekayasa proses berupa pemanfaatan sistem fed-batch untuk produksi dengan menggunak a n gula fermentasi merupakan salah satu alternatif. Penggunaan fed-batch juga dapat meminimalisir terbentuknya produk samping yang dapat mempengaruhi pertumbuha n dan produksi enzim rekombinan (Shimizu et al., 1988; Tripathi et al., 2009; Luli & Strohl, 1990).
Media MTM untuk pertumbuhan E. coli BL21 rekombinan menghasilkan OD600
maksimum sebesar 2.896 hingga 3.883 dan media LB mampu menghasilk a n pertumbuhan hingga mencapai OD600 sebesar 6.160. Perbedaan ini disebabkan oleh
pertumbuhan sel yang lambat untuk media LB, sehingga nilai OD600 yang dapat
dicapai oleh media LB lebih besar daripada media MTM. Akan tetapi, lamanya waktu logaritmik yang dialami E. coli BL21 rekombuinan dengan media LB, mengakibatk a n waktu produksi yang lebih lama. Media MTM, meskipun hanya mampu mencapai setengah dari nilai OD600 maksimum media LB, memiliki waktu produksi yang relatif
singkat. Hal ini mengindikasikan bahwa media MTM memiliki kemampuan sebagai media alternatif untuk pertumbuhan E. coli BL21 rekombinan.
III. KESIMPULAN
Berdasarkan data dan penjelasan di atas, dapat disimpulkan bahwa media MTM memiliki potensi untuk menjadi media alternatif dalam produksi enzim xilanolitik dari kultur campuran Escherichia coli BL21 rekombinan. Produksi dengan menggunak a n media MTM tidak memerlukan IPTG sebagai induser untuk meningkatkan aktivita s enzim xilanolitik rekombinan. Media MTM dan LB memiliki laju pertumbuha n spesifik yang berbeda. Media MTM memiliki laju pertumbuhan spesifik yang lebih tinggi dari pada laju pertumbuhan spesifik media LB.
IV. UCAPAN TERIMAKASIH
Terimakasih kepada Staff dan Tim Peneliti dari Laboratorium Proteomik, Lembaga Penyakit Tropis, Universitas Airlangga yang telah menyediakan tempat untuk melakukan penelitian ini.
Terimakasih kepada Prof. Dr. Ni Nyoman Tri Puspaningsih, M.Si yang telah memberikan dukungan, fasilitas dan bantuan dana dalam menyelesaikan penelit ia n ini.
DAFTAR PUSTAKA
Doelle, H. W., Ewings, K. N., & Hollywood, N. W., 1982, Regulation of glucose metabolism in bacterial system, Advanced Biochemistry Engineering, 1-35. Eiteman, M. A., & Altman, E., 2006, Overcoming acetate in Escherichia coli
recombinant protein fermentations, TRENDS in Biotechnology, 24(11), 530-536.
El-Mansi, E. M., & Holms, W. H., 1989, Control of carbon flux to acetate excretion during growth of Escherichia coli in batch and continuous cultures, Journal of General Microbiology, 2875-2883.
Gleiser, I. E., & Bauer, S., 1981, Growth of E. coli W to high cell concentration by oxygen level linked control of carbon source. Biotechnology and Bioengineering, 1015-1021.
Harris, A. D., & Ramalingan, C., 2010, Xylanases and its application in food industr y: a review, Journal of Experimental Science, 1(7), 01-11.
Horward, R. L., Abotsi, E., Jansen van Rensburg, E. L., & Howard, S., 2003, Lignocellulose biotechnology: issues of bioconversion and enzyme production, African Journal of Biotechnology, 2(12), 602-619.
Jensen, E. B., & Carlsen, S., 1990, Production of recombinant human growth hormone in Escherichia coli: expression of different precursors and physiological effects of glucose, acetate, and salts, Biotechnology and Bioengineering., 1-11. Koh, B. T., Nakashimada, U., Pfeiffer, M., & Yap, M. G., 1992, Comparison of acetate
inhibition on growth of host and recombinant E. coli K12 Strain, Biotechnology Letter, 1115-1118.
Luli, G. W., & Strohl, W. R., 1990, Comparison of growth, acetate production and acetate inhibition of Escherichia coli strains in batch and fed-batch fermentation, Applied and Environmental Microbiology, 56(4), 1004-1011. Miller, G. L., 1959, Use of DinitrosaIicyIic Acid reagent for determination of reducing
sugar, Analytical Chemistry, 31(3), 426-428.
Nakano, K., Rischke, M., Sato, S., & Marki, H., 1997, Influence of acetic acid on the growth of Escherichia coli K12 during high-cell density cutivation in a dialys is reactor. Applied Microbiology and Biotechnology, 597-601.
Numan, M. T., & Bhose, N. B., 2006, α-L-arabinofuranosidases: the potential application in biotechnology, Journal of Industrial Microbiology & Biotechnology, 33, 247-260.
Pan, J. G., Rhee, J. S., & Lebeaulut, J. M., 1987, Physiological constraints in increasing biomass concentration of Escherichia coli B in fed-batch culture, Biotechnology Letter, 89-94.
Polizeli, M. L., Rizzatti, A. C., Monti, R., Terenzi, H. F., Jorge, J. A., & Amorim, D. S., 2006, Xylanases from fungi: properties and industrial applications, Applied Microbiology and Biotechnology, 577-591.
Puspaningsih, N. N. T., 2004, Pencirian enzim xilanolitik dan kloning gen penyandi xilosidase dari Bacillus thermoleovarant IT-08,Disertasi, Institut Pertanian Bogor.
Puspaningsih, N. N., 2011, Produksi "Excelzyme" sebagai Bahan Campuran "Biodex" Pendegradasi Bahan Organik, Riset Kerjasama Unair-PT. Pupuk Kaltim, Universitas Airlangga, Surabaya
Ratnadewi, A. A., Fanani, M. Z., Kurniasih, S. D., Sakka, M., Warsito, E. B., Sakka, K., Nurachman, Z., Puspaningsih, N. N. T., 2013, β-D-xylosidase from Geobacillus thermoleovarans IT-08: biochemical characterization and bioinformatics of the enzyme, Applied Biochemistry and Biotechnology, 170(8), 1950-1964.
Rinas, U., Kracke-Helm, H.-A., & Schugerl, K., 1989, Glucose as a substrate in recombinant strain fermentation technology, Applied Microbiology and Biotechnology, 163-167.
Saha, B. C., 2003, Hemicellulose bioconversion, Journal of Industrial Microbiology & Biotechnology, 30, 279-291.
Saha, B. C., 2000, α-L-arabinofuranosidases: biochemistry, molecular biology and application in biotechnology, Biotechnology Advances, 8, 403-423.
Sezonov, G., Joseleau-Petit, D., & D'Ari, R., 2007, Escherichia coli Physiology in Luria-Bertani Broth, Journal of Bacteriology, 189(23), 8746-8749.
Shimizu, N., Fukuzono, S., Fujimori, K., Nishimura, N., & Odawara, Y., 1988, Fed-batch of recombinant Escherichia coli with inhibitory substance concentratio n monitoring, Journal of Fermentation Technology, 187-191.
Terrasan, C. R., Temer, B., Sarto, C., Junior, F. G., & Carmon, E. C., 2013, Xylanase and β-xylosidase from Penicillium janczewskii: production, physico-chemica l properties and application of the crude extract to pulp biobleaching, BioResource, 8(1), 1292-1305.
Tripathi, N. K., Sathyaseelan, K., Jana, A. M., & Rao, P. L., 2009, High yield production of heterologus proteins with Echerichia coli, Defence Science Journal, 137-146.
Winkelhausen, E., & Kuzmanova, S., 1998, Microbial conversion of D-xylose to xylitol, Journal of Fermentation and Bioengineering, 1-14.
Wong, K. K., Tan, L. U., & Saddler, J. N., 1988, Multiplicity of β-1,4-Xylanase in microorganisms: functions and applications, Microbiological Reviews, 52(3), 305-317.