TINJAUAN PUSTAKA
Tanah Ultisol dan Permasalahannya
Pada wilayah beriklim tropika basah seperti di Indonesia, kemasaman tanah yang tinggi merupakan suatu masalah utama yang sering ditemui. Curah hujan ≥ 2.000 mm per tahun, temperatur rata-rata 27 C, mengakibatkan tercucinya kation-kation basa, sehingga tanah umumnya didominasi oleh oksida aluminium dan besi yang tinggi. Hal ini mengakibatkan tanah bereaksi masam. Masamnya tanah ini dapat terjadi sebagai akibat kemampuan ion Al dalam menghidrolisis air sehingga dibebaskan ion H . Ion H+ merupakan anasir penyebab tanah menjadi masam. Salah satu jenis tanah yang bersifat masam adalah Ultisol.
Luas tanah Ultisol di Indonesia adalah 45.8 juta ha atau sekitar 24 luas daratan Indonesia (Subagyo et al. 2000). Pemanfaatan tanah Ultisol untuk produksi banyak menghadapi masalah, dimana terdapat horizon argilik dengan kepadatan yang tinggi di dekat permukaan tanah yang mengakibatkan hambatan terhadap laju perkolasi air hujan ataupun penetrasi akar tanaman, sehingga apabila terjadi hujan, lapisan tanah bagian atas akan cepat mengalami jenuh air, bersifat masam, jumlah basa-basa yang dapat ditukar tergolong sangat rendah (Hidayat dan Mulyani 2002). Selanjutnya dikatakan bahwa pada reaksi tanah sangat masam (pH < 4.5) kelarutan Al dapat ditukarkan meningkat sehingga menaikan kejenuhan Al. Konsepsi pokok dari tanah Ultisol adalah tanah yang telah mengalami proses hancuran lanjut (ultimate) dan pencucian berat oleh curah hujan yang tinggi, berwarna merah kuning, berpenampang > 2 m, dan terdapat lapisan argilik dari akumulasi liat (Subagyo et al. 2000). Tanah Ultisol dengan kejenuhan Al lebih dari 30% dan pH kurang dari 4.5 akan menimbulkan cekaman Al bagi tanaman (Subagyo et al. 2000). Bentuk-bentuk Al dalam larutan tanah tergantung tingkat kemasamannya. Pada keadaan reaksi tanah sangat masam (pH 4.5), Al menjadi sangat larut terutama dalam bentuk Al 3+ yang beracun bagi tanaman (Rout et al. 2001; Vitorello et al. 2005). Akibat keracunan pertumbuhan akar menjadi terhambat dan akhirnya menurunkan kemampuan akar menyerap hara mineral dan air (Matsumoto et al. 1996; Samuel et al. 1997). Gejala umum yang paling nyata terlihat bila keracunan Al adalah terhambatnya pertumbuhan dan perpanjangan
akar, baik akar primer, akar lateral maupun bulu akar. Tanaman yang keracunan Al mempunyai akar yang pendek, percabangan sedikit, akar adventif lebih banyak tumbuh pada pangkal akar, serta akar primer berkembang melebar ke arah apikal meristem sehingga terlihat gemuk (Rout et al. 2001). Pertumbuhan akar yang demikian sulit melakukan penetrasi ke lapisan sub soil, menyebabkan penyerapan hara dan air menjadi lebih rendah (Marschner 1995).
Pada tanah masam, fosfat yang dibebaskan baik dari proses pelapukan mineral apatit, dekomposisi bahan organik ataupun pupuk, akan segera diikat oleh liat serta aluminium, besi, ataupun kalsium, sehingga fosfat tidak tersedia bagi tanaman karena berubah menjadi garam yang mengendap dan tidak larut air (Syekhfani 1999). Radjagukguk (1983) mengemukakan bahwa salah satu ciri tanah mineral masam adalah rendahnya kandungan P dan fiksasi P yang tinggi.
Taksonomi, Karakteristik dan Habitat FMA
Mikoriza merupakan suatu struktur khas pada sistem perakaran yang terbentuk karena adanya simbiosis mutualistik antara fungi (myces) dan akar (rhiza) dari tumbuhan tingkat tinggi. Brundrett (2004) mendifinisikan mikoriza secara luas, yang mencakup seluruh keragaman mikoriza sebagai suatu asosiasi simbiotik yang esensial bagi satu atau kedua mitra, antara suatu fungi dan akar dari suatu tumbuhan hidup, yang terutama bertanggung jawab untuk transfer hara. Bentuk struktur khas mikoriza dapat dibedakan berdasarkan cara infeksinya pada perakaran tanaman inang, yaitu (1) endomikoriza, merupakan struktur mikoriza yang terbentuk sampai ke dalam sel korteks akar, (2) ektomikoriza, merupakan struktur mikoriza pada lapisan luar akar yang bentuknya berupa jala hartig; dan (3) ektendomikoriza, merupakan struktur mikoriza yang tidak hanya dapat membentuk jala hartig di permukaan akar, tetapi dapat menembus sel korteks (Smith dan Read 1997). Salah satu simbion fungi yang banyak membentuk struktur endomikoriza pada tanaman pertanian adalah FMA.
Berdasarkan hasil tes analisa DNA filum Glomeromycota dikenali ada dua belas genus yaitu Archaeospora, Geosiphon, Paraglomus, Gigaspora, Scutellospora,
Acaulospora, Kuklospora, Intraspora, Entrophospora, Diversipora, Pacispora dan Glomus (Schubler et al. 2001, Oehl dan Sieverding 2006). Dalam INVAM (2006)
dinyatakan bahwa FMA adalah salah satu tipe fungi mikoriza dan termasuk kedalam golongan endomikoriza termasuk kedalam golongan Glomeromycota, dengan ordo Glomales yang mempunyai dua sub-ordo, yaitu Gigasporineae dan Glomineae. Gigasporineae dengan famili Gigasporaceae mempunyai dua genus yaitu Gigaspora dan
Scutellospora. Glomineae mempunyai empat famili yaitu Glomaceae dengan genus Glomus, famili Acaulosporaceae dengan genus Acaulospora dan Entrophospora, Paraglomaceae dengan genus Paraglomus dan Archaeosporaceae dengan genus Archaespora. Identifikasi FMA dapat dilakukan berdasarkan morfologi sporanya,
ataupun dengan menggunakan teknik molekuler. Perbedaan morfologinya dapat dilihat dari perkembangan spora, bentuk spora, ukuran spora, warna spora, pola lapisan dinding spora dan reaksi warnanya, ornamentasi pada dinding spora, isi spora, perkecambahan spora dan hifa (Simanungkalit 2007).
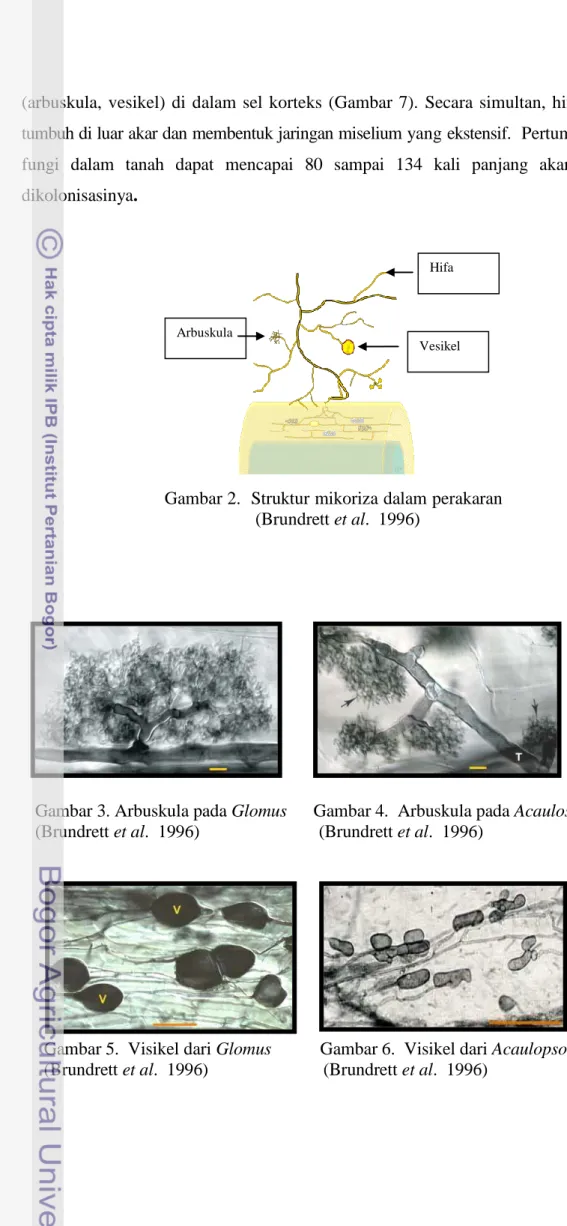
Fungi mikoriza arbuskula bersifat obligat, tidak mampu melengkapi daur hidupnya tanpa tanaman inang. Sporanya dapat berkecambah dan tumbuh tanpa tanaman inang akan tetapi pertumbuhannya sangat terbatas. Fungi mikoriza arbuskula memiliki beberapa karakteristik yaitu perakaran inang yang terkena infeksi tidak membesar, tetap mempunyai rambut-rambut akar sehingga penampilannya tidak berbeda dengan akar-akar yang tidak terinfeksi. Hal tersebut disebabkan karena fungi hanya membentuk struktur hifa tipis pada permukaan akar, tidak setebal mantel seperti pada ektomikoriza. Karakteristik lain yang merupakan ciri khas FMA adalah adanya struktur berbentuk percabangan hifa yang disebut arbuskula (arbuscules) dan ada juga yang membentuk struktur berbentuk oval yang disebut vesikula (vesicules), hifa koil dan spora (pada beberapa spesies) fungi mikoriza arbuskula dalam asosiasinya dengan tanaman juga membentuk organ/struktur diluar akar tanaman yaitu hifa eksternal, vesikula eksternal, dan spora. Arbuskula mengisi sebagian besar volume sel dan merupakan organ tempat pertukaran hara antara fungi dan tanaman. Vesikula berfungsi sebagai organ penyimpan cadangan makanan. Jenis FMA yang membentuk arbuskula dan vesikel adalah jenis Glomus, Sclerocitis, Acaulospora
dan Entrophospora (Gambar 2), sedangkan jenis lainnya seperti Gigaspo ra dan Scutellospora hanya membentuk struktur arbuskula (Brundrett et al. 1996).
Bentuk arbuskula dan vesikel pada masing-masing spesies berbeda (Gambar 3, 4, 5, 6). Struktur infeksi mikoriza arbuskula dicirikan oleh formasi struktur
(arbuskula, vesikel) di dalam sel korteks (Gambar 7). Secara simultan, hifa juga tumbuh di luar akar dan membentuk jaringan miselium yang ekstensif. Pertumbuhan fungi dalam tanah dapat mencapai 80 sampai 134 kali panjang akar yang dikolonisasinya.
Gambar 2. Struktur mikoriza dalam perakaran (Brundrett et al. 1996)
Gambar 3. Arbuskula pada Glomus Gambar 4. Arbuskula pada Acaulospora (Brundrett et al. 1996) (Brundrett et al. 1996)
Gambar 5. Visikel dari Glomus Gambar 6. Visikel dari Acaulopsora (Brundrett et al. 1996) (Brundrett et al. 1996)
Arbuskula
Vesikel Hifa
Secara umum proses kolonisasi FMA pada akar tanaman melewati empat tahap, yaitu (1) induksi perkecambahan spora dan pertumbuhan hifa, (2) kontak antara hifa dan permukaan akar yang menyebabkan pengenalan dan pembentukan apresorium, (3) penetrasi hifa ke dalam akar, dan (4) perkembangan struktur hifa interseluler.
.
Gambar 7. Struktur infeksi FMA pada sel-sel kortek akar (Diouf et al. 2003)
Fungi mikoriza arbuskula memiliki selang ekologis yang luas dan dapat dijumpai pada ekosistem semak, sabana (Cuenca dan Lovera 1992), arid (Allen dan Allen. 1992), semi arid (Lee et al. 1996), daerah temperate, tropika (Muthukumar et al. 1996), di daerah antartika (Phipps Taylor 1996), ekosistem gambut alami (Astianti Ekamawati 1996) dan gambut yang sudah terbuka (Ervayenri 1998; Kartika 2006), hutan hujan tropika (Janos Hartsorn 1997), padang rumput (Nadarajah dan Nawawi l997) serta daerah pantai (Setiadi 2000; Swasono 2006). Fungi mikoriza arbuskula dapat diisolasi dari tanah asam hingga alkalin pH 2.7- 9.2. Menurut Sieverding (1991), FMA yang hidup baik pada pH <5.0 adalah Entrophospora columbiana, pada pH >5.0 meliputi Glomus mosseae dan Gigaspora margarita serta pada pH 4.0 – 8.0 terdiri dari Acaulospora myriocarpa, A longula, A morrowae, A scrobiculata, G
aggregatum, G versiforme dan Scutellospora pellucida. Gigaspora gigantean
toleran terhadap kejenuhan Al tinggi. Hasil penelitian Heijne et al. (1996) menunjukkan bahwa infeksi FMA Glomus fasciculatum menurun dengan menurunnya pH tanah pada perakaran tanaman Arnica Montana L, Hietacium
dimulai dari propagul (spora dan residu akar) atau dari akar yang berdekatan dengan tanaman yang sama atau berbeda spesies tanaman. Propagul mampu menginfeksi akar tanaman inang karena adanya sinyal berupa eksudat flavanoid dari akar. Perkembangan infeksi FMA di akar berhubungan dengan pembentukan eksudat gula dan asam organik. Fungi mikoriza arbuskula dengan cepat mengkonversi dan mentransfer hasil fotosintat tanaman inang ke dalam senyawa karbon yang spesifik sebagai lipid atau glikogen (Gianinazzi-Pearson dan Gianinazzi 1983).
Mikoriza arbuskula dapat berasosiasi dengan hampir 90% jenis tanaman. Tiap jenis tanaman juga dapat berasosiasi dengan satu atau lebih jenis FMA. Beberapa penelitian menunjukkan bahwa setiap jenis tanah dan jenis tanaman memiliki jenis FMA yang berbeda, seperti di sawah tadah hujan Laladon Bogor ditemukan 2 isolat Gigaspora dan 5 isolat Glomus (Hanafiah 2001); pada lahan penanaman padi di tanah podsolik ditemukan 1 isolat Acaulospora, 4 isolat
Glomus (Iriani 2003); pada tanah PMK bekas hutan ditemukan 4 isolat Glomus, 5
isolat Acaulospora; pada tanah PMK bekas karet ada 7 isolat Glomus dan 2 isolat
Acaulospora (Kartika 2006), pada kawasan pantai Samas sekitar tegakan Tridax procumbens terdapat 2 isolat Glomus dan 1 isolat Gigaspora (Swasono 2006).
Terbentuknya simbiosis antara tanaman dan FMA sangat tergantung pada jenis FMA, genotip tanaman, faktor iklim dan kondisi tanah serta interaksi keempat faktor. Tanaman yang ketergantungannya tinggi terhadap fosfat akan cenderung untuk berasosiasi dengan mikoriza. Intensitas infeksi FMA dipengaruhi oleh berbagai macam faktor meliputi pemupukan, nutrisi tanaman, pestisida, intensitas cahaya, musim, kelembaban tanah, pH, kepadatan inokulum dan tingkat kerentaan tanaman (Fakuara 1988). Faktor lingkungan yang mempengaruhi perkembangan dan pertumbuhan hifa diantaranya: (1) oksigen, penurunan konsentrasi oksigen dapat menghambat perkecambahan spora FMA dan kolonisasi akar (Setiadi 1992), (2) suhu, suhu tanah yang tinggi umumnya dapat meningkatkan kolonisasi dan sporulasi FMA yang lebih tinggi (Gunawan 1993), (3) cahaya, besarnya intensitas cahaya berimplikasi pada banyak sedikitnya pembentukan FMA. Hal ini disebabkan karena cahaya matahari berperan dalam pembentukan karbohidrat melalui asimilasi karbon yang selanjutnya FMA akan
menggunakan karbon tersebut sebagai sumber energi bagi pertumbuhannya (Fakuara 1988), (4) keasaman tanah. Setiadi (1992) menyatakan bahwa perkembangan spora FMA sangat dipengaruhi oleh pH tanah, pH optimum untuk
Glomus sp antara 5.5-9.5, Gigaspora sp berkisar 4-6. Setiadi (2004)
menyimpulkan bahwa produksi spora pada kultur spora tunggal ditentukan oleh jenis media, tanaman inang, dan ukuran wadah. Media zeolit, tanaman inang dan wadah berukuran 250 ml merupakan kombinasi terbaik untuk menghasilkan jumlah spora tertinggi.
Peran FMA dalam Meningkatkan Serapan Hara
Fungi mikoriza arbuskula berperan penting dalam ekosistem alami maupun ekosistem yang telah dikelola. Beberapa penelitian menunjukkan bahwa simbiosis FMA dengan tanaman dapat meningkatkan serapan hara khususnya P, meningkatkan ketahanan tanaman terhadap patogen tular tanah, dan terhadap cekaman kekeringan. Beberapa cara FMA dalam meningkatkan serapan hara adalah 1) perluasan areal permukaan karena adanya hifa eksternal yang berukuran lebih kecil (1/10) dibandingkan dengan akar, 2) mempercepat pergerakan P ke akar melalui peningkatan afinitas P ke akar sehingga mengurangi konsentrasi ambang yang diperlukan P untuk berdifusi, 3) dengan merubah lingkungan rhizosfer secara kimia misalnya melalui pelepasan asam organik, dan peningkatan aktifitas fosfatase dan 4) meningkatkan produksi fitohormon yang dapat merubah fenotipe akar yaitu dengan pembentukan akar dengan orde yang lebih tinggi serta membuat umur akar menjadi lebih lama sehingga meningkatkan kapasitas penyerapan hara total (Orcutt dan Nielsen 2000). Secara fungsional, FMA dapat membantu penyerapan hara terutama P melalui tiga tahap yaitu 1) P diserap oleh hifa dari larutan tanah, 2) translokasi P melalui hifa, dan 3) transfer P melewati interfase ruang fungi (Cruz et al. 2004). De La Cruz (1988) menunjukkan bahwa FMA dapat mengefisienkan kira-kira 50% kebutuhan fosfat, 40 nitrogen dan 25 kalium. Hasil penelitian Mieke et al. (2003) menunjukkan bahwa efisiensi pupuk P pada tanaman kentang dapat ditingkatkan dengan pemberian inokulum fungi mikoriza yaitu meningkat 23.5 Unsur-unsur lain yang meningkat adalah Magnesium (Liu et a1. 2002). Pada tanaman cabai merah, inokulasi Gigaspora
margarita dapat membantu penyerapan P sebesar 30.95 (Haryantini Santoso 2001). Hasil penelitian Guntoro (2003) menunjukkan inokulasi FMA dan bakteri Azospirillum pada turfgrass C. Dactlon var. Tifdwarf dapat meningkatkan serapan hara dan meningkatkan efisiensi pemupukan.
Produksi hifa sangat bervariasi antara FMA, tetapi variasi ini tidak selalu mencerminkan besarnya serapan P (Sanders et al. 1977). Demikian pula dengan sebaran hifa bervariasi pada masing-masing spesies FMA (Jakobsen
el a1.1992). Acaulospora laevis menyebar 81 mm setelah empat minggu
sedangkan dua fungi lainnya hanya menyebar 31 mm panjang hifa. Panjang hifa eksternal berkisar antara 1-30 m/cm3. Selain itu umur hifa juga mempengaruhi serapan P. Bertambahnya umur hifa, maka semakin aktif dalam reaksi metabolisme, namun demikian umumnya hanya berkisar 5-7 hari setelah inisiasi (Friese dan Allen 1991). Serapan P oleh hifa sangat dipengaruhi oleh kedekatan hifa dan kualitas hifa. Semakin dekat dengan sumber P, maka serapan P akan semakin tinggi, demikian pula hifa yang cukup berumur akan lebih mampu menyerap P dibandingkan hifa yang belum matang
Mekanisme Serapan Fosfat
Kadar P total di dalam tanah umumnya rendah, dan berbeda-beda menurut jenis tanah. Tanah-tanah muda dan perawan biasanya lebih tinggi daripada tanah-tanah tua. Begitu juga penyebarannya di dalam profil tanah berbeda, terlihat bahwa kadar P-anorganik makin bertambah dengan dalamnya lapisan kecuali bentuk P-organik. Sirkulasi P di alam, dalam bentuk fosfat berbeda dengan sirkulasi karbon dan nitrogen. Fosfor terdapat dalam tanah, batuan dan organisme hidup dan tidak dalam atmosfer. Total fosfor yang terdapat dalam bumi cukup besar, pada kerak bumi terdapat 2x1018 kg atau 0.12%. Sumber P tanah meliputi 0.04% atau 1.2 x 1 013 kg.
Unsur hara fosfat di tanah dijumpai dalam tiga bentuk, yaitu P inorganik terlarut yang terdapat dalam larutan tanah, P inorganik tak larut yang terdapat pada kristal jattice, dan senyawa P organik. Bentuk P organik yang sering dijumpai adalah fosfolipid, asam nukleat dan fosfat inisitol. Pada umunya P
organik merupakan 20-50% dari total P tanah. Makin lanjut perkembangan tanah makin besar persentase P organik terhadap P total. Mineralisasi P organik menghasilkan H2PO4- yang segera diikat oleh komponen tanah. Bentuk P inorganik
dapat dibedakan menjadi (1) P aktif, yang meliputi P-Ca, P-Al dan P-Fe dan (2) P tidak aktif, yang meliputi „occlude-P”, dan mineral fosfat primer. (Soepardi 1983). Fosfat mobil dalam tanaman, tetapi tidak mobil dalam tanah (McWilliams 2003). Fosfat relatif tidak mudah bergerak di dalam tanah dan mengalami difusi yang lambat menuju akar tanaman yaitu berkisar 10-12 sampai 10-15 m2 detik-1,sehingga tingginya serapan P tanaman menyebabkan terjadinya daerah deplesi/pengurasan P di sekitar akar (Schachtman et al. 1998; Smith 2002). Rata-rata konsentrasi P dalam larutan tanah adalah 0.05 ppm dan sangat bervariasi diantara jenis tanah. Konsentrasi P dalam larutan yang diperlukan tanaman bervariasi dari 0.003 sampai 0.3 ppm dan bergantung pada spesies tanaman dan tingkat produksinya (Havlin et
al. 1999), sedangkan kemampuan tanaman mengambil P dari larutan tanah juga
bervariasi tergantung pada spesies (genotipe) dan dipengaruhi oleh interaksi antara tanaman dan mikroba (Kaeppler et al. 2000). Sifat khas dari P yaitu sukar larut dalam air namun larut dalam asam lemah, hal ini juga menjadi salah satu kendala rendahnya ketersediaan P tanah (Syekhfani 1999; Hardjowigeno 2007). Sifat khas P tersebut mengakibatkan tanaman memperoleh P melalui mekanisme difusi. Gardner et al. (1991) menyebutkan pada tanaman jagung, P yang diserap tanaman melalui mekanisme difusi 32 kali lebih banyak dari P yang diperoleh tanaman melalui mekanisme aliran massa dan kontak langsung. Fosfor organik dalam tanah tidak langsung tersedia untuk tanaman, tetapi terlebih dahulu harus dimineralisasi menjadi P anorganik yang selanjutnya berada dalam reaksi kesetimbangan dalam larutan tanah (Smith et al. 2003). Ion fosfat dalam tanah terdapat dalam tiga fraksi yaitu dalam larutan tanah, dapat ditukarkan (tidak stabil), dan tidak larut (stabil). Hanya P dalam larutan yang tersedia bagi tanaman. Sebagai unsur penyusun ATP dan ADP setiap organisme tanah, maka P juga dimanfaatkan oleh berbagai organisme, sehingga tanaman budidaya juga harus berkompetisi dengan berbagai organisme lain untuk mendapatkannya (Rao et al. 1999; Gardner et al. 1991).
karena hifa eksternal dari fungi mikoriza berperan sebagai system perakaran. Hifa eksternal menyediakan permukaan yang lebih efektif (lebih ekstensif dan lebih baik penyebarannya) dalam menyerap hara dari tanah yang kemudian dipindahkan ke tanaman inang. Hifa disini berperan sebagai “jalan bebas hambatan” untuk gerakan fosfat melalui zona deplesi di sekeliling akar. Dari hasil pengukuran laju gerakan P pada akar yang bermikoriza adalah kira -kira 6 kali laju akar tidak bermikoriza (Gunawan 1993). Fosfor yang diserap oleh FMA diubah menjadi polyfosfat yang kemudian ditranslokasikan ke komponen FMA dalam jaringan akar. Proses pengangkutan berlangsung sangat efisien dan hampir 1.000 kali lebih cepat daripada laju difusi ion P melalui tanah (Barea et al. 1980). Polifosfat dalam arbuskula, diubah menjadi orthofosfat yang kemudian masuk kedalam sitoplasma FMA. Dari sini P melewati dinding sel FMA dan memasuki tanaman inang melalui proses difusi. Cooper dan Tinker (1978) menyatakan pergerakan P dari medium ke tanaman dapat dibagi dalam tiga proses yaitu: 1) serapan P oleh hifa dari tanah, 2) translokasi P melalui hifa dan 3) transfer P melewati interfase ruang fungi.
De La Cruz (1981) melaporkan bahwa ektomikoriza mampu mengektrak senyawa berunsur hara secara langsung dari bahan organik, dan mengubahnya menjadi senyawa organik di dalam jaringan mikoriza selama proses metabolismenya, serta selanjutnya mengirimkan senyawa organik tersebut ke tanaman inang. Antibus et al. (1981) mengatakan bahwa ektomikoriza mampu mengubah fosfat organik tak larut menjadi tersedia bagi tanaman. Aktivitas ini terkonsentrasi pada selubung mikoriza, dimana pada selubung ini terjadi peningkatan aktivitas fosfatase.
Gunawan (1993) mengatakan ada beberapa mekanisme serapan fosfat dari sumber tidak larut: 1) Fosfat organik dapat dipakai oleh tanaman bermikoriza melalui aksi enzim fosfatase. Aktifitas fosfatase pada umumnya disebabkan oleh adanya mikroorganisme. Asam fosfatase yang dihasilkan oleh hifa FMA yang sedang aktif tumbuh dan peningkatan aktivitas fosfatase pada permukaan akar menyebabkan Pi dibebaskan oleh fosfat pada daerah dekat permukaan sel, sehingga dapat diserap melalui serapan akar. 2) Sumber
fosfat organik dapat dilarutkan oleh adanya asam-asam organik. Banyak fungi menghasilkan asam oksalat yang dapat mengkelat ion Ca atau membuang ion tersebut dari larutan tanah dalam bentuk kalsium oksalat. Asam oksalat dapat juga memacu pelepasan ion fosfat yang diabsorpsi oleh hidroksi besi dan alumunium. 3) Beberapa bakteri pelarut fosfat dan fungi tanah dapat melepaskan P ke dalam pool P labil di tanah, yang selanjutnya dapat diserap oleh tanaman bermikoriza.
Potensi mikoriza dalam membantu tanaman menyerap P bergantung pada kondisi P tanah. Pada tanah yang difisiensi P, simbiosis mikoriza dengan tanaman menguntungkan, sedangkan pada tanah yang ketersediaan P–nya tinggi, simbiosis tersebut tidak menguntungkan (Swift 2004). Level P tanah lebih dari 140 mg/kg (140 ppm), infeksi mikoriza akan menurun, sedangkan apabila level P tanah 50 mg/kg (50 ppm) perkembangan mikoriza tinggi . Rahim (2002) menyatakan bahwa keefektivan mikoriza berbeda setiap tanaman dan lingkungan
Aspek Mutu Benih
Diskripsi Benih Cabai Merah
Benih cabai berukuran kecil (3-5 mm), berwarna kuning dan berbentuk pipih seperti ginjal (buah pinggang). Berat 1000 biji kering berkisar antara 3-6 gram. Bagian-bagian benih cabai adalah embrio, endosperma, poros embrio (hipokotil, radikula, plumula), kotiledon, dan kulit benih.
Embrio cabai merah dikelilingi oleh endosperma. Endosperma berfungsi sebagai cadangan makanan untuk embrio dan kecambah muda. Endosperma terletak didepan radikal dan terdiri dari 7-9 sel (Walkins et al. 1985). Cadangan makanan benih sebagian besar tersusun protein dan lipid (Chen dan Lott 1992). Sel endosperma dibatasi oleh epidermis internal, epidermis berbentuk angular, mempunyai butiran-butiran minyak dan aleuron yang mengandung kristaloid.
Walkins et al., (1985) menyatakan bahwa dinding sel yang tebal dari endosperma cabai kemungkinan berfungsi sebagai penyimpan polisakarida yang mengandung mannan. Pada benih cabai, perubahan endosperma yang tampak dari
luar (secara visual) terlihat terjadi satu hari sebelum radikula membesar dan menembus kulit benih. Perubahan ini dibarengi dengan hilangnya integritas endosperma dan berkurangnya ketebalan pada endosperma di depan radikula, tetapi pada bagian endosperma lain tidak mengalami perubahan. Sel-sel pada bagian paling luar dari endosperma yang berbatasan dengan kulit biji dan sel-sel protoderm embrio diseliputi oleh kutikula. Bagian-bagian benih pada Gambar 8.
Gambar 8. Irisan melintang benih cabai: C: kotiledon, E: endosperma; H: hipokotil; R: radikula; SC: kulit biji (Bosland dan Votava 1999)
Viabilitas dan Vigor Benih
Mutu benih adalah salah satu faktor penentu keberhasilan pertanaman secara ekonomis. Petani Jepang mengatakan bahwa benih adalah setengah kesuksesan. Menurut Ilyas (2003) mutu benih yang tinggi dicirikan oleh (1) tingkat kemurnian tinggi, (2) daya berkecambah tinggi, (3) vigor tinggi (4) bebas dari penyakit seedborne.
Daya berkecambah benih didefinisikan sebagai muncul dan berkembangnya struktur terpenting dari embrio benih serta kecambah tersebut menunjukkan kemampuan untuk berkembang menjadi tanaman normal pada kondisi lingkungan yang menguntungkan. Vigor benih menurut ISTA adalah sejumlah sifat yang menentukan taraf potensi dari aktivitas dan penampilan benih selama perkecambahan dan pertumbuhan bibit. Menurut AOSA (1983) vigor adalah keadaan yang menentukan potensi tumbuh cepat dan merata serta berkembangnya bibit normal pada kisaran kondisi lapang yang luas. Benih yang vigor adalah benih yang kuat yang memiliki ciri-ciri: (1) tahan simpan, (2) berkecambah cepat dan merata, (3) bebas dari penyakit terbawa benih, (4) tahan terhadap gangguan mikroorganisme, (5) bibit tumbuh kuat, (6) bibit secara maksimum dapat memanfaatkan persediaan bahan makanan dalam benih,
sehingga dari padanya dapat tumbuh jaringan baru, (7) laju tumbuh atau penambahan bobot kering dari bibit yang sudah berfotosintesis lebih tinggi, (8) menghasilkan tanaman yang berproduksi tinggi dalam waktu tertentu. Vigor benih tidak hanya mengukur sifat tunggal, tetapi merupakan sejumlah sifat yang menggambarkan beberapa karakteristik yang berhubungan dengan penampilan suatu lot benih yaitu: a) kecepatan dan keserempakan berkecambah dan pertumbuhan kecambah, b) kemampuan munculnya titik tumbuh kecambah pada kondisi lingkungan yang tidak sesuai untuk pertumbuhan, dan c) kemampuan benih untuk berkecambah setelah mengalami penyimpanan (Ditjen Hortikultura, 2006). Sadjad (1993) menyatakan bahwa pengujian vigor mencakup dua yaitu: (1) pengujian kekuatan tumbuh dan (2) pengujian daya simpan. Kekuatan tumbuh benih dapat dicerminkan oleh kecepatan tumbuh dan ketahanan terhadap kekeringan. Benih yang cepat tumbuh mengindikasikan lebih mampu menghadapi kondisi lapang yang sub optimum.
Peranan P Dalam Benih
Fosfor merupakan unsur esensial untuk setiap sel hidup. Unsur ini terlibat langsung dalam berbagai metabolisme energi dan biosintesis asam nukleat dan membran, yang digunakan dalam proses fotosintesa, respirasi dan regulasi sejumlah enzyme (Raghothama 1999). Di dalam tanaman, fosfor terdapat sebagai fosfit ataupun P elemen. Fosfor inorganik banyak terdapat di dalam cairan sel yang merupakan komponen dari sistem penyangga tanaman. Fosfor dalam bentuk organik, antara lain terdapat sebagai: (1) fosfolipid, yang merupakan komponen membram sitoplasma dan khloroplas, (2) fitin, yang merupakan simpanan fosfat dalam biji, (3) gula fosfat, yang merupakan senyawa-senyawa dalam berbagai proses metabolisme tanaman, (4) nukleo protein, komponen utama DNA dan RNA dari inti sel, (5) ATP, ADP, dan AMP yang merupakan sumber energi untuk metabolisme, (6) NAD dan NADP, keduanya koenzim penting dalam proses reduksi dan oksidasi (Sutcliffe dan Baker 1974).
Fitin sebagai bentuk utama P dalam benih merupakan cadangan P yang tidak larut dan tidak dapat digunakan secara langsung oleh sel-sel tanpa perombakan menjadi P anorganik dengan enzim fitase. Fitin merupakan senyawa campuran dari potasium, magnesium dan kalsium dari asam myoinositol heksafosfat yang tersimpan dalam bentuk elemen fosfat dan mineral mikronutrien. Fitin ditransformasikan dari inositol
pada saat pemasakan benih dalam proses pembentukan benih (Noggle dan Fritz 1979). Senyawa fitin berfungsi sebagai cadangan fosfor dan untuk pemeliharaan energi dalam benih, sebab P dapat bergabung dengan dinukleotida (ADP) menjadi ATP.
Kadar P di dalam benih ditentukan oleh jenis tanah (kondisi tanah), varietas dan iklim (Fathan et al. 1988). Setiap varietas memiliki kadar P yang berbeda-beda. Kandungan asam fitat pada serealia berkisar antara 0.5% - 0.8%, sedangkan pada legum berkisar antara 0.4% - 7.5%. Menurut Sadjad (1983) fitin dalam benih sangat menentukan kemampuan benih untuk mempertahankan viabilitasnya agar tetap tinggi. Karena itu masa sebelum dan sesudah antesis sangat mempengaruhi kondisi benih, karena pada masa itulah fitin dibentuk dan disimpan dalam organ penyimpanan makanan. Kandungan fitin yang lebih tinggi menjadikan vigor benih lebih baik sehingga kemampuannya untuk tumbuh dan berkembang pada lingkungan yang optimum dan sub optimum lebih tinggi pula.