TINJAUAN PUSTAKA
Bakteri Tahan Krom
Banyak mikroorganisme dapat hidup dan bereproduksi dalam habitat yang terkontaminasi logam berat seperti Cr(VI). Kelompok Pseudomonas, Enterobacteria, Streptomyces spp., dan Corynebacteria yang diisolasi dari sedimen sungai Ottawa yang tercemar limbah industri mampu hidup dalam medium yang mengandung 100 µg Cr(VI) /L, dengan Pseudomonas sebagai kelompok yang paling banyak ditemukan (Luli, et.al., 1983). Dari lindi tempat pembuangan akhir sampah, Larashati (2004) mengisolasi bakteri Bacillus pumilus, Bacillus firmus, dan Bacillus brevis yang tahan terhadap kromat dalam konsentrasi tinggi dan mampu menyisihkan kromat dari medium. B. pumilus dan B. firmus resisten terhadap kromat sampai konsentrasi 500 mg/L, sedangkan Bacillus brevis resisten sampai konsentrasi kromat mencapai 400 mg/L. Bakteri jenis Bacillus banyak ditemukan di tempat pembuangan sampah dan ada yang memiliki resistensi dan kemampuan reduksi terhadap Cr(VI).
Kemampuan mikroorganisme untuk hidup dalam lingkungan yang terkontaminasi logam berat disebabkan oleh sejumlah mekanisme pertahanan yang dikembangkan mikroorganisme dalam mengatasi toksisitas logam berat. Gadd (1990) menyebutkan bahwa mekanisme pertahanan tersebut dapat melalui:
1. Presipitasi atau pembentukan kompleks ekstraseluler.
2. Menurunkan permeabilitas logam atau transport logam melewati membran sel.
3. Kompartementalisasi intraseluler, antara lain melalui penumpukan dalam vakuola.
4. Detoksifikasi melalui sejumlah reaksi kimia dalam sel.
Sedangkan menurut Bontidean, et.al. (2000) umumnya daya tahan bakteri terhadap berbagai jenis logam berat disebabkan adanya suatu faktor penentu yang memberikan resitensi terhadap satu atau sejumlah kecil logam berat. Mungkin karena bakteri adalah sel sederhana dengan kompartemen yang sederhana, pada umumnya resistensi bersifat sederhana untuk satu atau beberapa logam. Mekanisme resistensi pada bakteri meliputi pengaliran logam keluar sel bakteri
(efluks logam), modifikasi spesies logam, pengurungan logam, atau kombinasi dari mekanisme tadi. Mayoritas faktor penentu resistensi logam bersifat terinduksi oleh ion logam. Sistem metaloregulator yang telah diidentifikasi hingga saat ini adalah untuk kation atau oksianion Ag, As, Cd, Cr, Cu, Fe, dan Hg (Bontidean, et. al., 2000).
Terdapat indikasi bahwa resistensi terhadap kromium berkorelasi dengan keberadaan plasmid pada beberapa isolat (Bopp, et.el., 1983; Luli, et.al., 1983; Ohtake, et.al. 1987). Pseudomonas fluorescens LB300 yang diisolasi dari sedimen sungai Hudson yang terkontaminasi kromium dapat tumbuh dalam medium yang mengandung lebih dari 1,5 mg K2CrO4 per mL atau 200 kali lebih
resisten dibandingkan bakteri lain yang diisolasi dari lokasi yang sama. Resistensi terhadap Cr(VI) ini diperoleh dari plasmid pLHB1 (Bopp, et.al.,1983). Penelitian selanjutnya terhadap bakteri yang sama, yaitu Pseudomonas fluorescens LB300, mendapatkan bahwa adanya plasmid pLBH1 berhubungan dengan penurunan penyerapan Cr(VI). Strain sensitif-Cr(VI) P. fluorescens LB303 yang tidak memiliki plasmid, menyerap Cr(VI) 2,2 kali lebih banyak dibandingkan P. fluorescens LB300 yang resisten-Cr(VI). Selain itu, sensitivitas seluler terhadap kromat dipengaruhi oleh sulfat (Ohtake, et.al., 1987).
Hasil penelitian Wang, et.al. (1990) mendapatkan bahwa pada Enterobacter cloacae HO1 yang diisolasi dari lumpur aktif menggunakan CrO4
2-sebagai akseptor elektron pada kondisi anaerob dengan cara mereduksinya. Hal ini ditandai oleh:
1. Pertumbuhan bakteri dalam kondisi anaerob yang disertai dengan penurunan krom dalam medium.
2. Aktivitas reduksi-kromat pada sel yang sedang tumbuh dihambat oleh oksigen.
3. Reduksi terjadi lebih cepat dalam sel yang ditumbuhkan dalam medium gliserol dan asetat dibandingkan sel yang ditumbuhkan dalam medium glukosa.
Banyak lingkungan asam yang terjadi secara alami maupun yang diakibatkan oleh kegiatan manusia kaya akan krom dan logam-logam lainnya. Pada kondisi yang demikian, toksisitas logam seringkali muncul karena
logam-logam terdapat dalam bentuk yang bioavailable. Penelitian Cummings dan kawan-kawan (2007) mendapatkan bahwa bakteri pereduksi Fe(III) Acidiphilum cryptum memiliki toleransi yang tinggi terhadap beberapa logam berat termasuk Cr(VI) dan mampu mereduksi Cr(VI) melalui dua cara. Cara pertama, Cr(VI) direduksi secara langsung oleh komponen seluler yang sensitif terhadap Hg2+ dan tergantung pH (enzim). Reduksi Cr(VI) secara enzimatis ini tidak berhubungan dengan konservasi energi, tetapi lebih ditujukan sebagai strategi detoksifikasi. Cara kedua, Cr(VI) direduksi secara tidak langsung melalui perantaraan reaksi reduksi enzimatis Fe(III) menjadi Fe(II) yang secara cepat mentranspor elektronnya ke Cr(VI) sehingga tereduksi menjadi Cr(III) dalam tiga kali transfer elektron. Cara kedua tersebut dapat ditemui pada respirasi mikroba seperti Shewanell alga strain BrY (Wielinga, et.al., 2001). Pada bakteri tersebut reduksi besi merupakan proses penerimaan elektron terminal (TEAP, terminal electron accepting process) yang dominan dan reduksi tak langsung terhadap kromat oleh hasil samping respirasi tampaknya merupakan jalur reduktif utama seperti pada reaksi berikut: ¾ C3H5O3 + 3Fe(OH)3 ¾ C2H3O2 + 3Fe2+ + ¾ HCO3 + 2H2O + 5¼ OH (1) 3Fe2+ + HCrO4 + 8H2O 3Fe(OH)3 + Cr(OH)3 + 5H+ (2)
Reduksi Krom (VI) secara Enzimatis pada Beberapa Bakteri
Beberapa penelitian pada sejumlah bakteri yang berbeda mendapatkan bahwa akumulasi kromium tampaknya melibatkan proses reduksi enzimatis ekstraseluler atau intraseluler, atau keduanya dengan salah satu proses sebagai proses reduksi utamanya seperti isolat Pseudomonad CRB5 (Mc Lean, 2000). Berbagai spesies bakteri mengembangkan sistem reduksi Cr(VI) yang berbeda-beda, bahkan pada spesies yang sama seperti Pseudomonas putida PRS2000 (Ishibashi, et.al., 1990) dan Pseudomonas putida MK1 (Park, et.al., 2000) memiliki enzim terlarut yang sifatnya sangat berbeda yang digunakan untuk mereduksi Cr(VI).
Pseudomonas putida PRS2000 yang dipelajari oleh Ishibashi dan kawan-kawan (1990) diketahui memiliki aktivitas enzim kromium reduktase dalam ekstrak sel, sementara pada fraksi membran sel tidak ditemukan aktivitas pereduksi krom (VI). Enzim terlarut ini bersifat heat labile dimana pemanasan pada 50oC selama 10 menit dapat menurunkan aktivitas hingga 40-50%, sementara pH optimum diperoleh pada kisaran 6,5 – 7,5. Ekstrak enzim dari sel P. putida PRS2000 membutuhkan NADH atau NADPH sebagai donor elektron untuk mereduksi kromat. Walau konsentrasi sulfat dalam medium mempengaruhi sensitivitas seluler terhadap Cr(VI) (sebagai CrO42-) (Ohtake, et.al., 1987),
reduksi kromat tidak dipengaruhi oleh sulfat dan oksianion lain (SO32-, MoO42-,
VO42-, PO42-, dan NO3-), serta Cr3+. Hg2+ dan Ag+ merupakan inhibitor kuat bagi
enzim ini.
Enzim solubel pereduksi Cr(VI) yang didapatkan dari ekstrak periplasma sel P. putida MK1 (Park, et.al., 2000) memiliki perbedaan dari P. putida PRS2000, dimana enzim MK1 memiliki pH optimum 5,0 dan suhu optimum 80oC. Selain itu, enzim MK1 ini dihambat oleh sulfat, sedangkan enzim PRS2000 tidak terhambat. Karena kromat dengan mudah melewati membran sel dan di dalam sitoplasma bersifat toksik, maka sifat enzim MK1 sebagai protein periplasmik memberi keuntungan bagi sel karena detoksifikasi kromat dapat langsung terjadi begitu kromat memasuki sel bakteri.
Proses reduksi enzimatis ekstraseluler ditemukan di bakteri Enterobacter cloacae HO1, Escherichia coli ATCC 33456, dan Ochrobactrum anthtopi. Wang, et.al. (1990) mendapatkan bahwa, pada Enterobacter cloacae HO1 dalam kondisi anaerobik, aktivitas tertinggi enzim yang dapat mereduksi Cr(VI) terdapat di fraksi membran sel. Aktivitas enzim kromat-reduktase membran paling tinggi (yaitu sebesar 5,47 µg CrO42-/menit/mg protein) terjadi ketika fenazin metosulfat
yang telah direduksi oleh asam askorbat ditambahkan sebagai donor elektron ke dalam medium. Pemanasan membran sel pada 100oC selama satu menit menyebabkan hilangnya aktivitas reduksi (Wang, et.al, 1990). Penelitian terhadap E. coli ATCC 33456 yang dilakukan Shen dan Wang (1993) mendapatkan bahwa reduksi Cr(VI) menjadi Cr(III) sebagian besar terjadi dalam medium eksternal. Sementara penelitian terhadap O. anthropi (Li, et.al., 2008) mendapatkan bahwa
reduksi enzimatis ekstraseluler mendominasi reduksi Cr(VI) dan hanya sebagian kecil saja Cr(VI) ditemukan di dalam sel dan tereduksi menjadi Cr(III). Akumulasi kromium dipermukaan sel menyebabkan permukaan sel menjadi kasar.
Reduksi enzimatis Cr(VI) menjadi Cr(III) secara intraseluler ditemukan pada P. ambigua G-1 (Suzuki, et.al.,1992), P. putida (Ishibashi et al., 1990; Park, et.al., 2000; Ackerley, et.al., 2004), dan P.fluorescens (Mc Lean, 2000 dan 2001) yang menggunakan enzim reduktase terlarut yang terdapat di dalam sitoplasma untuk mereduksi kromat baik secara aerobik maupun anaerobik. Walau demikian laju reduksi tertinggi terjadi di dalam kondisi anaerobik. Beberapa enzim yang diketahui memiliki aktivitas pereduksi-Cr(VI) adalah glutation reduktase, aldehida oksidase, dan sitokrom P-450 (Suzuki, et.al., 1992).
Suzuki, et.al. (1992) mendapatkan bahwa ekstrak sel P. ambigua G-1 membutuhkan 3 mol NADH sebagai donor elektron untuk mereduksi secara enzimatis 1 mol Cr(VI) menjadi Cr(III). Dalam reaksi ini, Cr(V) terbentuk sebagai senyawa antara sehingga Suzuki dan kawan-awan menyimpulkan bahwa reaksi reduksi Cr(VI) adalah reaksi dua tahap. Pertama, Cr(VI) menerima satu elektron dari satu molekul NADH dan menghasilkan senyawa antara Cr(V). Tahap kedua, senyawa antara Cr(V) menerima dua elektron dari dua molekul NADH sehingga terbentuk Cr(III). Tahap pertama terjadi lebih cepat dari tahap kedua. Pada kedua tahap ini, NADH berubah menjadi NAD+.
Isolat Pseudomonad CRB5 yang berkerabat dekat dengan P. fluorescens mereduksi Cr(VI) menjadi Cr(III) menggunakan enzim terlarut yang kemungkinan besar terdapat di sitoplasma atau di periplasma, sementara pada fraksi membran plasma tidak ditemukan aktivitas kromat reduktase (Mc Lean, 2000; Mc Lean and Beveridge, 2001). Aktivitas enzim reduktase ini juga ditemukan dalam medium setelah 48 jam inkubasi. Hal ini mungkin merupakan suatu proses detoksifikasi ekstraseluler dimana enzim disekresikan ke lingkungan yang terkontaminasi Cr(VI) atau keberadaannya disebabkan oleh lisis sel yang mati. Isolat CR5 tidak membutuhkan donor elektron eksternal seperti NADH, tetapi isolat ini dapat menggunakan sumber elektron endogenus. Walau demikian,
reduksi minor Cr(VI) yang berasosiasi dengan membran ditemukan dalam kondisi anaerob non-pertumbuhan dengan adanya donor elektron organik.
Tabel 1 menyajikan nilai Km dan Vmax enzim pereduksi Cr(VI) dari
beberapa bakteri. Enzim pereduksi Cr(VI) dari bakteri P. putida, yang telah dipurifikasi menggunakan kolom Superose 12 HR 10/30, memiliki nilai Km dan
Vmax yang lebih tinggi dibandingkan bakteri lainnya. Metoda pemurnian enzim
mempengaruhi nilai Km dan Vmax enzim.
Tabel 1. Nilai Km dan Vmax dari enzim peredusi Cr(VI) yang diisolasi dari
beberapa jenis bakteri.
Bakteri Lokasi Enzim Metoda Purfikasi Donor Elektron Km Vmax Pustaka
P. putida sitoplasma Superose
12 HR 10/30 NADH 374 µM 1,72 µmol/menit/mg protein Park, et.al., 2000 Bacillus sp. sitoplasma (NH4)2SO4 NADH 14 µM 3,8 nmol/menit/mg protein Elangovan, et.al., 2006 V. harveyi sitoplasma Sephadex
G-75 NADH 5,4 µM 10 nmol/menit/mg protein Kwak, et.al., 2003 P.ambigua sitoplasma Sephadex
G-75 NADH 13 µM 27 nmol/menit/mg protein Kwak, et.al., 2003
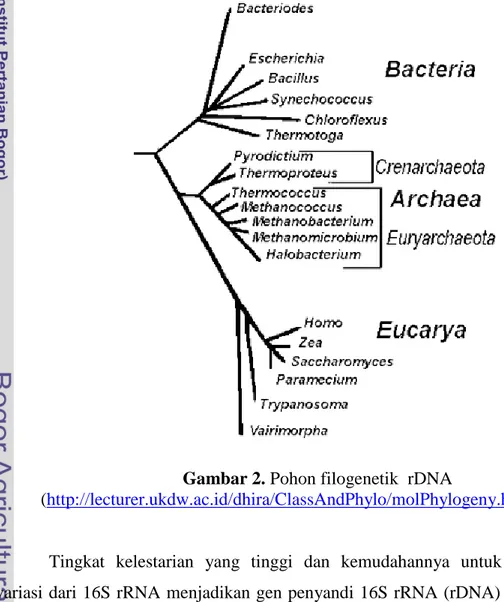
16S RNA Ribosom (rRNA)
RNA ribosom (rRNA) merupakan komponen inti dari mesin pembentuk protein pada semua makhluk hidup yaitu ribosom. Molekul rRNA bersifat ubikuitus dengan fungsi yang identik pada seluruh organisme (Pangastuti, 2006). Ribosom merupakan kompleks ribonukleoprotein yang terdiri atas dua subunit yaitu subunit kecil (small subunit) dan subunit besar (large subunit). Terdapat tiga jenis RNA dalam ribosom prokaryot berdasarkan nilai koefisien sedimentasi, yaitu 5S, 16S, dan 23S rRNA. 16S rRNA merupakan bagian dari subunit kecil ribosom dan berperan penting dalam pengenalan ujung 5’-mRNA serta memposisikannya pada letak yang tepat dalam ribosom.
RNA ribosom pada semua sel memiliki daerah yang urutan basanya sangat lestari dan beberapa daerah yang urutan basanya variatif. Perbandingan urutan basa yang lestari berguna untuk mengkonstruksi pohon filogenetik karena
mengalami perubahan dengan relatif lambat dan mencerminkan kronologi evolusi bumi. Sebaliknya, urutan basa yang bersifat variatif dapat digunakan untuk melacak keragaman dan menempatkan galur-galur dalam satu spesies (Pangastuti, 2006).
Dari tiga jenis rRNA yang dimiliki prokaryot, gen penyandi 16S rRNA paling sering digunakan sebagai penanda molekuler karena memiliki ukuran basa yang paling ideal dari segi analisis statistika dibandingkan gen penyandi 5S rRNA dan 23S rRNA. Molekul 5S rRNA memiliki ukuran basa yang terlalu pendek sehingga tidak ideal untuk analisis statistika, sementara molekul 23S rRNA memiliki srtuktur sekunder dan tersier yang cukup panjang sehingga menyulitkan analisis (Pangastuti, 2006).
Gambar 2. Pohon filogenetik rDNA
(http://lecturer.ukdw.ac.id/dhira/ClassAndPhylo/molPhylogeny.html)
Tingkat kelestarian yang tinggi dan kemudahannya untuk mengukur variasi dari 16S rRNA menjadikan gen penyandi 16S rRNA (rDNA) secara luas digunakan pada penentuan taksonomi, filogeni (hubungan evolusioner), dan
memperkirakan laju penyebaran bakteri (Weisburg, et.al., 2001). Penelitian mengenai kekerabatan evolusioner di antara mikroorganisme berdasarkan perbandingan urutan basa 16S rDNA dipelopori oleh Carl Woese (Krane and Raymer, 2003). Berdasarkan urutan basa 16S rDNA, Woese mengajukan tiga Domain dalam sistem klasifikasi yaitu Archaea, Bacteria, dan Eucarya (Gambar 2). Dua bakteri dapat dikelompokkan dalam satu genus bila memiliki kemiripan maksimum (maximum identity) > 93% (Hagstrom, et.al., 2002), sementara dua bakteri dianggap sebagai satu spesies bila memiliki kemiripan maksimum >97% (Hagstrom, et.al., 2002; Schloss and Handelsman, 2004; Pangastuti, 2006).
Reaktor Pengolah Limbah Pelapisan Krom
Cr(VI) mudah tereduksi oleh Fe(II) di alam (Moore and Ramamoorthy, 1984). Mineral besi yang melimpah, seperti pirit dan mineral besi-sulfur lainnya, memegang peranan penting dalam siklus geokimia sejumlah unsur-unsur renik termasuk kromium (Houda, et.al.,2007).
Laboratorium Pengendalian Pencemaran di Puslit Limnologi LIPI telah mengembangkan reaktor kolom pengolahan limbah cair Cr(VI) yang berasal dari produk akhir limbah pelapisan logam berdasarkan reaksi reduksi-oksidasi antara krom dan besi. Disain reaktor tersebut dapat dilihat pada Gambar 8. Spesifikasi reaktor kolom pengolahan limbah Cr(VI) sebagai berikut:
Dimensi : Diameter : 16 cm
Tinggi : 80 cm
Volume void : 16 L Media Reaktor : tinggi 70 cm Komposisi: Fe(0) diameter : 10 mm Karakteristik: Porosity Fe : 10,7349%
Bulk sample : 1,7945 gram/cm3 Q (Loading rate) : 4 Liter/hari
HRT : 0,5 day
Gambar 3. Disain reaktor kolom dan Instalasi reaktor kolom pengolahan limbah cair pelapisan logam dan limbah dari kegiatan analisis Laboratorium Hidrokimia P2-Limnologi LIPI.Keterangan: 1. Influent; 2. Pompa; 3. Flowmeter; 4a. Reaktor kolom 1; 4b. Reaktor kolom 2; 5a. Effluent 1 (tertutup); 5b. Effluent 2 (tertutup); 6a. Gas Buangan (terbuka, larutan NaCl); 6b. Gas Buangan (terbuka, larutan NaCl); 7. Gas Port; 8. Sampling Port
Prinsip reaktor pengolah limbah ini adalah berdasarkan reaksi reduksi-oksidasi antara Cr(VI) dan Fe(0) yang menghasilkan Cr(III) dan Fe(II) dan Fe(III). Ketika Fe(0) terendam air anoksik maka akan terjadi korosi secara elektrokimia dan elektron-elektron dilepaskan pada suatu area (bagian anoda). Sementara pada bagian katoda ion-ion hidrogen dihasilkan dari disosiasi air untuk memproduksi atom-atom hidrogen atau molekul-molekul pada daerah katoda:
Fe0 Fe2+ + 2e- anoda 2H+ + 2e- H2(g) katoda
____________________________________ Fe0 + 2H+ Fe2+ + H2(g) reaksi
Fe0 Fe3+ + 3e- anoda
CrO42- + 4H2O+ 3e- Cr(OH)3 + 5OH- katoda
__________________________________________________ Fe0 + CrO42- + 4H2O Fe3+ + Cr(OH)3 + 5OH- reaksi
Reaksi redoks melibatkan oksidasi Fe0 dengan oksigen terlarut dan reduksi ion kromat dengan kation Fe(II), berdasarkan persamaan berikut:
2Fe0 + O2 +H2O 2Fe2+ +4OH- (1)
Fe2+ + CrO42- + 4H2O Fe(OH)3 +Cr(OH)3 + 2OH- (2)
Reaksi ini meningkatkan pH pada sistem dan membentuk Cr(III) dalam bentuk hidroksida tak larut (Crx Fe1-x )(OH)3 sebagai hasil reduksi kromat.
Reaktor pengolahan ini menggunakan dua jenis materi Fe(0) yang memiliki ukuran partikel berbeda. Pada reaktor kolom 1, Fe(0) yang digunakan berupa partikel halus dan pada reaktor kolom 2, Fe(0) yang digunakan berbentuk memanjang. Bahan materi besi Fe(0) diperoleh dari produk sisa pembubutan besi. Ukuran partikel Fe(0) yang berbeda menyebabkan adanya perbedaan porositas, Fe(0) berbentuk partikel halus memiliki porositas 4,7911 % dan bulk sample 1,6605 gram/cm3 (hasil analisis Laboratorium Geologi – Badan Geologi) (Gambar 9).
Gambar 4. Ukuran Fe(0) yang digunakan pada reaktor kolom; (a) 1 cm, (b) memanjang 1-2 cm dan (c) partikel halus
Rahayu dan Tontowi (2000) menggunakan FeSO4 untuk mereduksi
Cr(VI) dalam air limbah industri tekstil, industri penyamakan kulit, dan industri
pembuatan shampoo (konsentrasi Cr(II) 0,275 – 1,22 mg/L) dengan persentase penyisihan mencapai 100%,. Pada sistem ini, FeSO4 selain berperan sebagai
pereduksi senyawa ini juga bertindak sebagai koagulan. Sementara Houda dan kawan-kawan (2007) menggunakan mineral besi pirit untuk mereduksi Cr(VI) sebagai kromat. Houda mendapatkan bahwa laju penyisihan kromat dipengaruhi oleh konsentrasi awal pirit dan kromat, pH, suhu, dan jenis pirit. Penyisihan kromat meningkat dalam kondisi asam (pH 3) dan suhu tinggi (37oC).
Walaupun sistem pengolahan limbah krom(VI) yang memanfaatkan reaksi redoks antara Fe0 – Cr6+ tersebut relatif murah dan bahannya mudah didapat, metoda ini memiliki kelemahan. Dari penelitian pendahuluan didapatkan bahwa pada reaktor tersebut penurunan konsentrasi Cr6+ ternyata disertai kenaikan konsentrasi yang tinggi dari Fe2+ dan Fe3+, serta pembentukan gas H2S.
Konsentrasi Fe2+ terlarut dalam inlet reaktor meningkat dari 0,181 mg/L menjadi 68,062 mg/L dalam outlet reaktor, sementara Fe3+ terlarut yang awalnya 0,984 mg/L meningkat menjadi 50,838 mg/L. Selain itu, pada outlet reaktor terbentuk H2S sebesar 2,562 mg/L. Walaupun besi relatif kurang toksik dibandingkan
krom(VI), toksisitas besi tetap dapat muncul bila terjadi akumulasi besi dalam konsentrasi tinggi.