T
he notothenioid fishes form a monophyletic radiation that dominates the fish fauna of the Antarctic continental shelf. This perciform suborder (Notothenioidei) has about 95 species divided among five families. Four of the families are predominantly or exclusively Antarctic; however, one family (Bovichtidae) is only found outside the Antarctic. Hence, the term Antarctic notothenioids refers to the suborder Notothenioidei minus the family Bovichtidae.The radiation of Antarctic fishes
Recent reviews1,2 summarize our understanding of the origins of the evolutionary radiation, and how it relates to the Antarctic environ-ment past and present. The develo-ping consensus is that Antarctic notothenioid success can be at-tributed to one key evolutionary innovation – glycopeptide
anti-freeze1(Box 1). Protection from freezing is essential in shal-low ice-covered sea, because teleost blood is less salty than seawater and freezes at a correspondingly higher tempera-ture. Thus, in shallow Antarctic habitats, freezing protec-tion alone might be sufficient to explain the dominance of notothenioid fishes and the virtual absence of competition from other teleost fish groups. Elasmobranch fishes employ an osmoregulatory strategy providing them with sufficient freezing protection; however, they are only poorly represented in Antarctic waters. Eastman1suggests that there might be other habitat or trophic factors that explain their low representation in the Antarctic fish fauna. Recent molecular evidence has shown that the anti-freeze glycoprotein evolved from pancreatic trypsinogen3, thus providing an exciting and rare example of how an old protein gene can be changed to generate a new gene for an entirely new protein with a new function. Indeed, if the notothenioid antifreeze is the ‘key evolutionary inno-vation’ of the group, it presents us with a unique case of being able to directly relate an extensive radiation, and faunal dominance of a group of fishes through organismal adaptation, to protein evolution.
Adaptive radiations are important case studies in evolu-tionary biology. However, within the fishes we have only a few clear-cut examples. The best known is the radiation of cichlid fishes in the African Great Lakes; however, there is also a significant radiation of cottid fishes in Lake Baikal4. It is interesting to briefly compare the notothenioid radiation with the better known cichlid radiations. Like the notothe-nioids, the cichlids have evolved to play a wide variety of eco-logical roles5; however, the cichlid faunas of Lakes Malawi
and Victoria have evolved more rapidly6, thus generating a higher species richness. These cichlid radiations have also provided a rich source of data against which to debate issues of evolution and adaptation, such as the relation-ships between microevolution and macroevolution6, speciation pro-cesses7,8and adaptive radiation9,10. The intrinsic scientific inter-est of Antarctica and the multi-national nature of Antarctic sci-ence has meant that surprising progress has been made in under-standing the biology and evolu-tion of Antarctic fishes. In the past five years there have been over 150 papers on Antarctic fishes: 30 on ecological questions; over 80 on anatomical, physiological and molecular adaptations to the environment (particularly low temperature); 15 on systematics; and 20 on evolution. From this and previous work, the phylogen-etic relationships of the group are relatively well known and the timescale over which the radiation has occurred is also well constrained. Thus, the monophyletic radiation of Antarctic notothenioids is an excellent system with which to explore issues of adaptation and evolutionary change, comparable to and complementary with the cichlid radi-ations. The main thrust of this article is to single out the issue of adaptive loss and regain, which has recently become evident in the evolution of Antarctic fishes. Disap-tation and its partial reversal are apparent in the lateral-line sensory systems of species that have become second-arily pelagic, and in the haemoglobinless channichthyids.
Disaptation and recovery in the
evolution of Antarctic fishes
John Montgomery and Kendall Clements
The radiation of notothenioid fishes provides an excellent system to explore issues of evolution
and adaptation. Most studies emphasize adaptation to the extreme Antarctic environment; however, recent work provides cogent examples of disaptation or evolutionary
loss of function. The nature and extent of regressive change is revealed by subsequent adaptive recovery. Ancestral notothenioids were
benthic but some became secondarily pelagic through the retention of larval characters. Paedomorphosis has produced detrimental changes in lateral-line sensory systems that have been made good by compensatory adaptation. In the icefish family, compensatory adaptation has followed the loss of the oxygen-binding pigments haemoglobin and myoglobin.
John Montgomery and Kendall Clements are at the School of Biological Sciences, University of Auckland,
Private Bag 92019, Auckland, New Zealand ([email protected];
Box 1. Glossary
Allometry: the relationship between the sizes of various parts of an organism. Antifreeze: Antarctic fishes have a glycopeptide antifreeze, which is secreted into the blood and other body fluids. The effect of the antifreeze is to lower the freezing point of the blood and prevent the build-up of ice crystals. Crista: a molecular layer that overlies the primary sensory nucleus of the lateral-line system. The eminentia and crista form part of an adaptive filter that cancels self-generated noise in the mechanosensory lateral-line system. Eminentia: part of the brain associated with the cerebellum. The main cells in this area are the granule cells whose axons form the parallel fibres of the crista. Myoglobin: an oxygen-binding pigment located in muscle cells.
Secondary pelagicism and paedomorphosis Generally, it is agreed that notothenioids are derived from bottom-dwelling ances-tors without swimbladders1. Typical benthic fishes with no swimbladder and no special buoyancy adapta-tions in water weigh about 5% of their weight in air. The thornfish (Bovichtus variegatus), often taken as a functional outgroup for the Antarctic notothenioids, and the New Zealand black cod (Notothenia angustata) weigh in at close to that. By con-trast, Antarctic notothenioids all weigh considerably less than 5% of their weight in water (Fig. 1). This is especially true of the pelagic
Pagothenia borchgrevinki,
Pleuragramma antarcticum
and Dissostichus mawsoni. Eastman11 has detailed the distinctive characters of
Pleuragramma, including those that relate to their pelagic lifestyle (Table 1). To his list, we could also add silveryness and modest lateral compres-sion – two characters also found in pelagic notothenioids and typical of many midwater teleosts. Both are persistent larval characteristics that contribute to midwater camouflage.
The suite of characters associated with secondary pelagicism make an interesting case study for understand-ing evolutionary processes. As Eastman11has noted, many of these characters are paedomorphic (Box 1), a view re-inforced by osteological studies12. In general, notothenioids have pelagic larval stages and it appears that secondary pelagicism has evolved from the retention of larval charac-ters. The mechanism underlying paedomorphosis is thought to be a change in the timing of developmental events11,13. Changes at such a fundamental level will impact on a large interrelated character set, with the result that it makes little sense to try to tease apart individual traits as adaptations. In addition, only a subset of these characters
Fig. 1.Phylogeny of selected notothenioid taxa. Dots on the cladogram show the multiple evolution of a pelagic lifestyle. The cladogram is based on Klingenberg and Ekau13, and the character distribution is from Eastman11.
Habitat and buoyancy (weight in water) data are from Eastman1.
Trends in Ecology & Evolution
Bovichtus variegatus
Dissostichus mawsoni
Notothenia angustata
Trematomus pennellii
Trematomus loennbergii
Trematomus newnesi
Trematomus bernacchii
Trematomus hansoni
Trematomus nicolai
Pagothenia borchgrevinki
Pleuragramma antarcticum Pelagic Pelagic
Cryopelagic Benthic Benthic Benthic Benthic Benthic Benthic
Epibenthic
Semipelagic
Habitat
5.87 ± 0.17
0.01 ± 0.01
4.43 ± 0.14
3.04 ± 0.11
2.28 ± 0.17
2.62
3.37 ± 0.09
2.91 ± 0.07
3.13 ± 0.08
2.75 ± 0.08
0.57 ± 0.07
Wt in H2O/ wt in air
100 ( SE)
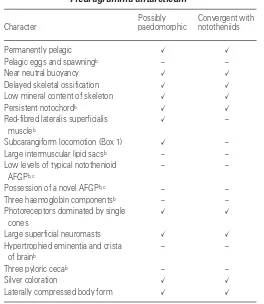
Table 1. Distinctive characters of adult Pleuragramma antarcticuma
Character
Possibly Convergent with paedomorphic nototheniids
Permanently pelagic ✓ ✓
Pelagic eggs and spawningb – –
Near neutral buoyancy ✓ ✓
Delayed skeletal ossification ✓ ✓
Low mineral content of skeleton ✓ ✓
Persistent notochordb ✓ ✓
Red-fibred lateralis superficialis ✓ – muscleb
Subcarangiform locomotion (Box 1) ✓ – Large intermuscular lipid sacsb – –
Low levels of typical notothenioid – – AFGPb,c
Possession of a novel AFGPb,c – –
Three haemoglobin componentsb – –
Photoreceptors dominated by single ✓ ✓ cones
Large superficial neuromasts ✓ ✓
Hypertrophied eminentia and crista – – of brainb
Three pyloric cecab – –
Silver coloration ✓ ✓
Laterally compressed body form ✓ ✓
aExcept for large superficial neuromasts, all characters are apomorphic when
polarized relative to Trematomus bernacchii. Check marks indicate characters that might be paedomorphic in Pleuragramma and also characters that are convergent in other water column clades of nototheniids.
bAutapomorphic (unique derived or diagnostic) characters for Pleuragramma.
cAFGP, antifreeze glycopeptide.
Box 2. Definitions
Adaptation:an organismal character produced by natural selec-tion for a particular biological role33–35.
Aptation:an organismal character that confers use to the organism, regardless of whether its phylogenetic origin featured natural selection33,34.
Disaptation:an organismal character whose use to the organism is demonstrably inferior to that of a phylogenetically antecedent character33,35.
will contribute to performance enhancement within the defined ‘selection regime’ (sensuLauder et al.14).
Certain character changes might be correlated with the paedomorphic change, but be functionally neutral or even disadvantageous; this evolutionary loss of function is termed disaptation (Box 2). For example, incomplete canal formation in the lateral line can be caused by an early arrest of develop-ment. This leaves canal neuromasts exposed on the surface of the body without the benefit of the front-end mechanical filter that the canals usually provide15 (Fig. 2). In essence, the boundary layer generated within the canals protects the sense organ from low-frequency self-generated noise. The loss of the canals is an example of a change induced by an altered developmental programme with potentially detrimental func-tional implications16. It is interesting that Pleuragramma behaves as a sit-and-wait predator11, and that the eminentia and crista of the brain are hypertrophied (Box 1; Table 1). These characteristics would, to some extent, compensate for the loss of canals. Sit-and-wait predatory behaviour reduces self-induced noise at the level of the receptor, and the eminentia and crista form part of a sophisticated adaptive fil-ter that cancels self-induced noise17. It would appear that loss of the lateral-line canals, incidental to paedomorphic change, might have been compensated for by changes in behaviour and central sensory processing mechanisms.
Mapping the occurrence of secondary pelagicism on a notothenioid cladogram indicates that it has evolved at least three times across the group1,11,13(Fig. 1). Thus, on the basis of multiple independent evolutionary origins it is reasonable to talk of the retention of larval characters in notothenioids as an adaptation to the pelagic habitat. One of these secondarily pelagic groups includes the genus
Dissostichus. What is remarkable about the two
Dissostichus species is their large size in comparison with other notothenioids (Fig. 3). One interpretation is that the changes in development lead-ing to paedomorphosis have resulted in an altered growth profile. Whatever the de-velopmental mechanism, the increased size of D. mawsoni
has functional implications for certain organ systems. The mechanosensory lateral line again provides an exam-ple. The mechanical filtering effect of lateral-line canals depends on canal diameter – a large canal loses the ability to act as a filter. Allometric (Box 1) growth of lateral-line canals in D. mawsoni
would produce wide canals with little or no attenuation of low-frequency noise. Com-pensatory adaptation again appears to have occurred with the incorporation of a narrow section of canal in the region of the receptor that reasserts the low-frequency attenuation and has the added benefit of producing a high-frequency amplification15.
Loss of respiratory pigments in the icefish
Loss of haemoglobin and myoglobin (Box 1) in the family Channichthyidae also provides compelling examples of disaptation and recovery. Loss of haemoglobin is an autapomorphy (unique derived or diagnostic character) for Channichthyidae. The cold temperature of Antarctic waters increases oxygen solubility, such that haemoglobin loss is
Fig. 2.Lateral view of the head of Trematomus hansoni showing the pore openings of the lateral-line system. Trematomus hansoni is a benthic notothenioid with an adult body length of approximately 200 mm. The trunk lateral-line system is visible, as are the pore openings of the infraorbital canal line (below the eye) and other head lines. The pores open into canals within which are the sensory neuromasts. The canals act as a mechanical filter, attenuating low frequency noise. If the canals are lost by the early arrestment of development, as happens particularly in the trunk system of pelagic species, then this front-end filter is lost.
not lethal but is detrimental. This point is made in studies of red-blooded notothenioids, which can successfully survive haemoglobin poisoning with carbon monoxide18, but pre-sumably with a reduced scope for activity. Further evidence that the loss of haemoglobin is detrimental comes from mol-ecular studies that provide evidence for positive selection of haemoglobin in red-blooded Antarctic fishes19. For example,
Gymnodraco acuticeps (Bathydraconidae, sister taxa to Channichthyidae) has a higher rate of nonsynonymous substitutions than synonymous substitutions in the DNA sequences of its betaglobin gene. This is taken as evidence of positive selection on the haemoglobin gene with its asso-ciated corollary that loss would represent a disaptation.
Recent molecular evidence suggests that the abro-gation of haemoglobin synthesis in icefishes probably resulted from a single mutational event in the ancestral channichthyids that deleted the entire betaglobin gene and the 59end of the linked alphaglobin gene20–22. The strongest evidence that this loss represented a disaptation comes from the compensatory adaptive changes that have subse-quently occurred in icefishes. For example, compared with red-blooded notothenioids, the relative mass of the saclike ventricle is about three times larger and blood volume is about two to four times larger than that of other teleosts1,23. In addition, systemic resistance is estimated as being less than 10% that of rainbow trout24. These compensatory changes mean that the heart functions as a volume, rather than a pressure, pump, and that large volumes of blood are circulated at high flow rates and low pressures to meet the metabolic demands of the tissues25,26.
Myoglobin is a respiratory pigment found in muscle, which enhances oxygen delivery to the tissue. Recent evi-dence27–29shows that myoglobin in icefishes is expressed only in the cardiac ventricle and that this expression occurs in many, but not all, icefish species. Phylogenetic and molecular analysis demonstrates that mutations resulting in the loss of myoglobin expression have oc-curred independently at least three times, and by two dis-tinctly different mechanisms. In Chaenocephalus aceratus
and Pagetopsis macropterus, mature messenger RNA (mRNA) encoding myoglobin is apparently not produced, despite the presence of the myoglobin gene in the DNA of these species. In Champsocephalus gunnari, mature myo-globin mRNA is detectable but this message is not trans-lated to produce myoglobin protein at detectable levels.
Multiple loss might be expected if the myoglobin was nonfunctional. However, Sidell and co-workers have recently presented molecular30 and physiological28,31evidence indi-cating that retention of myoglobin is under positive selection pressure, and that it does play a functional role in the icefish heart. For example, in myoglobin-expressing species myoglo-bin cDNA sequences are highly conserved. By comparison, the two species expressing mutant myoglobin RNA exhibited a high degree of sequence variation.
The performance of isolated perfused hearts from myo-globin-containing and myoglobinless icefishes provides the best demonstration of the functional role of myoglobin in the icefish heart. Myoglobin-containing hearts of Chionodraco rastrospinosus are capable of maintaining cardiac output at higher after-load challenges than the myoglobinless hearts of C. aceratus. However, poisoning the myoglobin of
C. rastrospinosushearts dropped their performance to well below that of C. aceratus, which provides evidence not only that myoglobin is functional where it occurs in icefish hearts, but also that compensatory adaptive changes have occurred in the hearts of myoglobinless species. The nature of these compensatory adaptive changes is yet to be determined.
Disaptation and recovery
Readaptation implies an adaptive change that compen-sates for an earlier detrimental loss (Box 2). In the case of paedomorphic change or other macroevolutionary change involving developmental mechanisms, it is easy to under-stand how other interrelated structures could be im-pacted, at times negatively. Paedomorphic change is not an uncommon occurrence in evolution. There are several other fish examples where members of benthic taxa have reradiated into the pelagic habitat by the retention of lar-val characters. For example, the pelagic New Zealand blennioid triplefin Obliquichthys is thought to be paedo-morphic32. The Baikal radiation of cottid fishes might also provide examples. It will be instructive to study these other paedomorphic groups with respect to associated loss and regain of particular functional attributes.
In the case of the repeated loss of single functional pro-teins, such as myoglobin, it is less obvious how these detri-mental changes could become fixed in the population, although genetic drift in small populations is a possible sug-gestion. Whatever the mechanism of detrimental change, be it pleiotropy (Box 1) or genetic drift, the Antarctic fish radi-ation provides explicit examples of the nature and extent of regressive change or disaptation in evolution. The best evi-dence of loss is the subsequent adaptive recovery, where the compensatory adaptive change both illuminates the loss and demonstrates the propensity of adaptive evolution.
Conclusions
It is possible to argue that the low competitive environ-ment under which the notothenioid radiation has occurred has allowed a tolerance of disaptation, and that this phe-nomenon might be higher in this radiation than elsewhere. However, it could equally well be argued that disaptation can only be adequately studied where there is a good phyl-ogeny on which to map both the disaptation and the recov-ery. A study of other such radiations is required to address the issue of the extent to which disaptation, then recovery, is a general phenomenon in evolution.
References
1 Eastman, J.T. (1993) Antarctic Fish Biology: Evolution in a Unique Environment, Academic Press
2 Clarke, A. and Johnston, I.A. (1996) Evolution and adaptive radiation of
antarctic fishes. Trends Ecol. Evol. 11, 212–218
3 Chen, L. et al. (1997) Evolution of antifreeze glycoprotein gene from a
trypsinogen gene in Antarctic notothenioid fish.Proc. Natl. Acad. Sci. U. S. A. 94, 3811–3816
4 Eastman, J.T. and Clarke, A. (1998) A comparison of adaptive
radiations of Antarctic fish with those of nonAntarctic fish. In Fishes of Antarctica: A Biological Overview (Di Prisco, G. et al., eds), pp. 3–26, Springer-Verlag
5 Carroll, R.L. (1997) Patterns and Processes of Vertebrate Evolution, Cambridge University Press
6 Johnson, T.C. et al. (1996) Late Pleistocene dessication of Lake Victoria and rapid evolution of cichlid fishes.Science 273, 1091–1093
7 Martens, K. (1997) Speciation in ancient lakes. Trends Ecol. Evol.
12, 177–182
8 Seehausen, O. and Bouton, N. (1997) Microdistribution and fluctuations in niche overlap in a rocky shore cichlid community in Lake Victoria. Ecol. Freshw. Fish. 6, 161–173
9 Bouton, N. et al. (1999) Local adaptations in populations of rock-dwelling haplochromines (Pisces: Cichlidae) from southern Lake Victoria. Proc. R. Soc. London Ser. B 266, 355–360
10 Rüber, L. et al. (1999) Replicated evolution of trophic specializations in an endemic cichlid fish lineage from lake Tanganyika. Proc. Natl. Acad. Sci. U. S. A. 96, 10230–10235
12 Voskoboynikova, O.S. (1994) Rates of individual development of the bony skeleton of eleven species of the family Nototheniidae. J. Ichthyol. 34, 108–120
13 Klingenberg, C.P. and Ekau, W. (1996) A combined morphometric and phylogenetic analysis of an ecomorphological trend: pelagization in Antarctic fishes (Perciformes: Nototheniidae). Biol. J. Linn. Soc. 59, 143–177
14 Lauder, G.V. et al. (1993) Adaptations and history. Trends Ecol. Evol. 8, 294–297
15 Montgomery, J.C. et al. (1994) Aspects of structure and function in the anterior lateral line of six species of antarctic fish of the suborder Notothenioidei. Brain Behav. Evol. 44, 299–306
16 Coombs, S. and Montgomery, J.C. (1994) Function and evolution of superficial neuromasts in an antarctic notothenioid fish. Brain Behav. Evol. 44, 287–298
17 Montgomery, J.C. and Bodznick, D. (1994) An adaptive filter cancels self-induced noise in the electrosensory and lateral line
mechanosensory systems of fish.Neurosci. Lett. 174, 145–148
18 di Prisco, G. et al. (1992) Antarctic fish survive exposure to carbon monoxide. Experientia 48, 473–475
19 Bargelloni, L. et al. (1998) Antarctic fish hemoglobins: evidence for adaptive evolution at subzero temperature.Proc. Natl. Acad. Sci. U. S. A. 95, 8670–8675
20 Cocca, E. et al. (1995) Genomic remnants of alpha-globin genes in the hemoglobinless Antarctic icefishes. Proc. Natl. Acad. Sci. U. S. A. 92, 1817–1821
21 Cocca, E. et al. (1997) Do the hemoglobinless icefishes have globin genes? Comp. Biochem. Physiol. A Physiol. 118, 1027–1030
22 Zhao, Y. et al. (1998) The major adult alpha-globin gene of Antarctic teleosts and its remnants in the hemoglobinless icefishes: calibration of the mutational clock for nuclear genes.J. Biol. Chem. 273, 14745–14752
23 Acierno, R. et al. (1995) Blood volume in the hemoglobinless Antarctic teleost Chionodraco hamatus (Lonnberg). J. Exp. Zool. 272, 407–409
24 Egginton, S. and Rankin, J.C. (1998) Vascular adaptations for a low pressure/high flow blood supply to locomotory muscles of Antarctic icefish. In Fishes of Antarctica: A Biological Overview (di Prisco, G. et al., eds), pp. 185–196, Springer-Verlag
25 Tota, B. et al. (1997) The heart of the Antarctic icefish as paradigm of cold adaptation. J. Thermal Biol. 22, 409–417
26 Feller, G. and Gerday, C. (1997) Adaptations of the hemoglobinless Antarctic icefish (Channichthyidae) to hypoxia tolerance. Comp. Biochem. Physiol. A Physiol. 118, 981–987
27 Sidell, B.D. et al. (1997) Variable expression of myoglobin among the hemoglobinless Antarctic icefishes. Proc. Natl. Acad. Sci. U. S. A. 97, 3420–3424
28 Sidell, B.D. (1998) Intracellular oxygen diffusion: the roles of myoglobin and lipid at cold body temperature. J. Exp. Biol. 201, 1118–1127
29 Sidell, B.D. and Vayda, M.E. (1998) Physiological and evolutionary aspects of myoglobin expression in the haemoglobinless Antarctic icefishes. In Cold Ocean Physiology (Porter, H.O. and Playle, R.C., eds), pp. 122–142, Cambridge University Press
30 Vayda, M.E. et al. (1997) Conservation of the myoglobin gene among Antarctic notothenioid fishes. Mol. Mar. Biol. Biotech. 6, 207–216
31 Acierno, R. et al. (1997) Myoglobin enhances cardiac performance in antarctic icefish species that express the protein. Am. J. Physiol. 273, R100–R106
32 Fricke, R. (1994) Tripterygiid Fishes of Australia, New Zealand and the Southwest Pacific Ocean (Teleostei), Koeltz Scientific Books
33 Larson, A. and Losos, J.B. (1996) Phylogenetic systematics of adaptation. In Adaptation (Rose, M.V. and Lauder, G.V., eds), pp. 187–219, Academic Press
34 Gould, S.J. and Vrba, E.S. (1982) Exaptation – a missing term in the science of form. Paleobiology 8, 4–15
35 Baum, D.A. and Larson, A. (1991) Adaptation reviewed: a phylogenetic methodology for studying character macroevolution. Syst. Zool. 40, 1–18
S
elf-promoting or selfish gen-etic elements (SGEs) have long been of interest to evolu-tionary biologists, because their existence emphasizes the impor-tance of a gene level perspective when interpreting selection and adaptation. They spread because they encode traits beneficial to their own transmission, even if they impart costs to the organ-isms carrying them1,2. They in-clude ‘ultracompetitive’ alleles, supernumerary B chromosomes, organelles and intracellular para-sites1–5. These factors manipulate gametogenesis or host reproduc-tion by a variety of means to enhance their own transmission; for instance, by inducing meiotic drive (Box 1) or cytoplasmic in-compatibility (CI; Box 1), and byfeminizing, killing or sterilizing males (Box 2). Early popu-lation genetic models predicted that, in the absence of sup-pression, many such elements should spread through populations to high frequency or fixation, sometimes with
disastrous consequences for the host population. For instance, sex ratio distorters at fixation should cause population (and thus their own) extinction owing to the absence of one sex. Other SGEs, such as those causing autosomal meiotic drive or CI, will lose their effect at fixation and, therefore, might become impossible to de-tect. Although this has almost certainly happened in some instances2, many empirical stud-ies suggest that polymorphism for SGEs is widespread in natural populations and that SGEs are maintained at relatively low fre-quencies (Table 1). Recent em-pirical and theoretical studies are now beginning to shed light on this problem.
Selection for suppressors
Because the spread of SGEs creates genetic conflict with other elements of the host genome, selection should favour genes that modify or resist their action2. Generally,
Persistence of selfish genetic elements:
population structure and conflict
Melanie J. Hatcher
Selfish genetic elements are vertically transmitted factors that spread by obtaining a transmission advantage relative to the rest of the genome of their host organism, often with a
cost to overall host fitness. In many cases, conventional population genetics theory predicts them spreading through populations, reaching fixation and becoming undetectable or
sometimes driving the population extinct. However, in several well studied systems, these
genetic elements are known to persist at relatively low, stable frequencies. Recent research suggests that several processes might explain these observations, including population structure, intragenomic conflict and coevolution.
Melanie Hatcher is at the School of Biology, Miall Building, University of Leeds, Leeds, UK LS2 9JT