RESPON BEBERAPA GENOTIPE PADI TERHADAP
PERIODE KEKERINGAN PADA SISTEM SAWAH
HERMAN WAFOM TUBUR
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011
Dengan ini saya menyatakan bahwa tesis yang berjudul ”Respon Beberapa Genotipe Padi terhadap Periode Kekeringan pada Sistem Sawah” merupakan gagasan dan karya saya bersama pembimbing yang belum pernah dipublikasikan dalam bentuk apapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, Juli 2011
Herman W Tubur NRP. A252080051
HERMAN W TUBUR. Responses of Rice Genotypes to Drought Periods on Low Land Rice System. Under direction of M A. CHOZIN, E. SANTOSA and A. JUNAEDI.
The objective of this experiment was to identify responses of IR64, Ciherang, IPB-97-F-15, Menthik Fragrant, Rokan, Way Apo Buru, Jatiluhur and Silugonggo to drought. The experiment was conducted under plastic house with massive walls 4x3 m (LxW) with split plot design experiment with four replications. Drought treatments were induced at 3, 6, 9 WAT (weeks after transplanting) and control (standard rice growing). The results showed that drought at 3 and 6 WAT more reduced growth rate and yield. Yield reduction on drought at 3 and 6 WAT caused by increased of unfilled spikelet. The highest of plant canopy showed by Jatiluhur, whereas Silugonggo showed the highest on number of leaf, tiller and productive tillering than other genotypes. Jatiluhur and Silugonggo have highest percentage of flowering, whereas longest panicle showed by IPB-97-F-15. Jatiluhur showed the highest production of grain yield and harvest with lowest of unfilled spikelet percentage. There are differences in responses 8 rice genotypes to drought periods in the low land rice system. Based on the parameters of productive tillers, scores leaf rolling and leaf desication, and drought resistance index for yield, all eight genotypes can be grouped into 3 groups: Way Apo Buru Jatiluhur were tolerant; Rokan and Menthik Fragrant were sensitive; IR64, Ciherang, IPB-97-F-15 and Silugonggo were moderate to drought.
HERMAN W TUBUR. Respon Beberapa Genotipe Padi terhadap Periode Kekeringan pada Sistem Sawah. Dibimbing oleh M. A. CHOZIN, E. SANTOSA, dan A. JUNAEDI.
Tujuan penelitian ini adalah untuk mengidentifikasi respon beberapa genotipe padi terhadap periode kekeringan pada sistem sawah. Penelitian dilakukan di kebun percobaan Institut Pertanian Bogor dalam rumah plastik pada 16 unit petak tanam dengan ukuran 4 m x 3 m, tiap petak dibatasi dinding tembok dengan kedalaman 0.8 m. Penelitian disusun menggunakan rancangan split plot design dengan 4 ulangan. Perlakuan pada petak utama adalah periode pengeringan mulai 3 MST, 6 MST dan 9 MST serta kontrol (tanpa pengeringan) dan perlakuan anak petak terdiri dari 8 genotipe padi yaitu IR64, Ciherang, Way Apo Buru, IPB-97-F-15, Jatiluhur, Silugonggo dan Menthik Wangi.
Periode pengeringan mulai 3 MST dan 6 MST nyata menghambat pertumbuhan vegetatif, hasil dan komponen hasil yang ditunjukkan dengan rendahnya tinggi tanaman, jumlah daun, jumlah anakan, jumlah anakan produktif, persen pembungaan dan panjang malai, bobot basah dan kering tajuk. Pengeringan saat 3 MST dan 6 MST juga meningkatkan persen gabah hampa dan menurunkan bobot gabah per rumpun, bobot 1000 butir dan indeks panen.
Tanaman tertinggi ditunjukkan oleh Jatiluhur dan IPB-97-F-15 sementara jumlah daun tertinggi ditunjukkan oleh Silugonggo. Jumlah anakan tertinggi ditunjukkan oleh IR 64, Silugonggo dan Rokan. Jumlah anakan produktif tertinggi ditunjukkan oleh Silugonggo. Persen pembungaan tertinggi ditunjukkan oleh Jatiluhur dan Silugonggo dan malai terpanjang ditunjukkan oleh IPB-97-F-15. Bobot 1000 gabah terendah ditunjukkan oleh Rokan sementara indeks panen tertinggi ditunjukkan oleh Jatiluhur, Silugonggo dan Ciherang. Bobot basah dan bobot kering tajuk tertinggi ditunjukkan oleh Rokan, Menthik Wangi dan IPB-97-F-15.
Terdapat perbedaan respon 8 genotipe padi terhadap periode kekeringan pada sistem sawah. Berdasarkan parameter jumlah anakan produktif, skor penggulungan dan skor kekeringan daun, dan indeks ketahanan kekeringan untuk daya hasil, kedelapan genotipe tersebut dapat dikelompokan menjadi 3 kelompok, Jatiluhur dan Way Apo Buru termasuk kelompok yang toleran; Rokan dan Menthik Wangi termasuk kelompok yang peka; IR64, Ciherang, IPB-97-F-15 dan Silugonggo termasuk pada kelompok agak toleran (moderat) terhadap kekeringan.
©
Hak cipta milik IPB, tahun 2011
Hak Cipta dilindungi oleh Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
Dilarang mengumpulkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apapun tanpa izin IPB.
PERIODE KEKERINGAN PADA SISTEM SAWAH
HERMAN WAFOM TUBUR
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011
Nama : Herman Wafom Tubur
NRP : A252080051
Mayor : Agronomi dan Hortikultura
Disetujui Komisi Pembimbing
Prof. Dr. Ir. M. A. Chozin, M.Agr
Ketua
Dr. Edi Santosa, SP., M.Si Dr. Ir. Ahmad Junaedi, M.Si
Anggota Anggota
Ketua Mayor
Agronomi dan Hortikultura
Dekan Sekolah Pascasarjana IPB
Dr. Ir. Munif Ghulamahdi, MS. Dr. Ir. Dahrul Syah, M.Sc. Agr.
Puji dan syukur penulis panjatkan ke Hadirat Tuhan Yang Maha Kuasa atas kasih dan kekuatan-Nya sehingga penulisan tesis ini dapat diselesaikan. Ucapan terima kasih penulis sampaikan kepada Prof. Dr. Ir. M. A. Chozin, M.Agr., Dr. E. Santosa, SP., M.Si dan Dr. Ir. A. Junaedi, M.Si selaku komisi pembimbing atas pengertian, arahan, kesabaran dan motivasinya. Teriring ucapan terima kasih kepada penyelenggara program BPPS dan TIM peneliti IMHERE B.2.C. yang tergabung dalam penelitian “Studi Fisiologi dan Pengembangan Tanaman untuk Adaptasi pada Kondisi Kekeringan dan Rendah Methan” atas segenap perhatian dan dukungannya baik moril ataupun materil.
Dalam menempuh pendidikan, pelaksanaan penelitian hingga penyelesaian tesis, penulis juga menyampaikan terima kasih atas kesabaran, pengertian dan dukungan doa dari mama, istri, anak dan keluarga.
Penulis juga menyampaikan terima kasih kepada pimpinan IPB, Dekan Program Pasca Sarjana, Ketua Mayor Agronomi dan Hortikultura serta teman-teman seangkatan 2008 dan senior atas dorongan dan motivasi yang diberikan.
Akhir kata, besar harapan penulis kiranya tesis ini bermanfaat untuk memperkaya khasanah ilmu pengetahuan.
Bogor, Juli 2011
Penulis dilahirkan di Jayapura, Papua pada tanggal 14 Maret 1981 dari ayah Musa W Tubur (alm.) dan Ibu Erna sebagai anak pertama. Penulis menikah dengan Oliva Asem dan dikaruniai seorang putra, Jeylandro P M Tubur Asem pada 13 April 2009.
Tahun 1999 penulis lulus dari SMU Negeri 3 Jayapura, dan pada tahun yang sama penulis melanjutkan studi S1 di Universitas Negeri Papua pada Program Studi Agronomi, Jurusan Budidaya Pertanian, Fakultas Pertanian dan lulus pada tahun 2004. Penulis mengabdi sebagai staf pengajar di Universitas Negeri Papua pada Program Studi Agronomi sejak tahun 2005, dan pada tahun 2008 penulis mendapat kesempatan untuk melanjutkan studi Program Magister Sains pada program studi Agronomi dan Hortikultura IPB dengan dukungan BPPS. Penulis juga mendapat kesempatan untuk menjadi bagian dalam tim Peneliti I-MHERE B2.C tahun 2010 yang diketuai oleh Dr. Ir. A. Junaedi, M.Si.
Halaman DAFTAR TABEL DAFTAR GAMBAR DAFTAR LAMPIRAN PENDAHULUAN ... 1 Latar Belakang ... 1 Perumusan Masalah ... 2 Tujuan Penelitian ... 3 Hipotesis ... 4 TINJAUAN PUSTAKA ... 5
Cekaman Kekeringan pada Lingkungan Tumbuh Padi ... 5
Mekanisme Adaptasi Tanaman Padi terhadap Kekeringan ... 6
Sifat Perakaran dan Hubungannya dengan Cekaman Kekeringan 9 Pertumbuhan Pucuk dan Cekaman Kekeringan ... 12
Hubungan Ketersediaan Air dan Pertumbuhan Tanaman ... 15
BAHAN DAN METODE ... 18
Lokasi dan Waktu ... 18
Bahan dan Alat ... 18
Rancangan Penelitian ... 18
Pelaksanaan Penelitian ... 19
Parameter Pengamatan ... 21
Analisis Data ... 23
HASIL DAN PEMBAHASAN ... 24
Hasil Pertumbuhan Vegetatif ... 24
Hasil dan Komponen Hasil ... 29
Sensitivitas Kekeringan ... 33
Potensial Air Tanah ... 36
Pembahasan Pertumbuhan Vegetatif ... 37
Hasil dan Komponen Hasil ... 39
Sensitivitas Kekeringan ... 42
Potensial Air Tanah ... 43
KESIMPULAN DAN SARAN ... 46
Kesimpulan ... 46
Saran ... 47
DAFTAR PUSTAKA ... 48
No Halaman
1. Hubungan antara satuan Bar (atm), cm air dan pF ... 16 2. Nilai skor penggulungan daun dan tingkat kekeringan ... 22
3. Rata-rata tinggi tanaman, jumlah daun dan jumlah anakan 8
genotipe padi saat 11 MST ... 28
4. Rata-rata jumlah anakan produktif. persen pembungaan dan bobot gabah per rumpun 8 genotipe padi pada berbagai perlakuan
kekeringan ... 30
5. Rata-rata panjang malai, persen gabah hampa, bobot gabah per
rumpun, dan bobot 1000 gabah. ... 31
6. Rata-rata bobot basah dan bobot kering tajuk, dan indeks panen
pada perlakuan kekeringan dan genotipe ... . 33
7. Rata-rata indeks ketahanan kekeringan untuk daya hasil 8
genotipe padi pada berbagai periode pengeringan ... . 34
8. Skor penggulungan dan kekeringan daun 8 genotipe pada
1. Model perlakuan periode kekeringan saat 3 MST, 6 MST, 9 MST
dan kontrol ... 20
2. Perkembangan tinggi tanaman padi pada berbagai perlakuan
periode kekeringan ... 24
3. Perkembangan tinggi tanaman 8 genotipe padi ... 25
4. Perkembangan jumlah daun padi pada berbagai perlakuan periode
kekeringan ... . 26
5. Perkembangan jumlah daun 8 genotipe padi ... 26
6. Pekembangan jumlah anakan padi pada berbagai periode
pengeringan ... 27
7. Perkembangan jumlah anakan 8 genotipe padi ... 28
8. Perkembangan potensial air tanah pada berbagai perlakuan
kekeringan yang diukur pada kedalaman tanah 10 cm ... 36
9. Perkembangan potensial air tanah pada berbagai perlakuan
No Halaman
1. Rekapitulasi sidik ragam tinggi tanaman, jumlah daun dan jumlah
anakan saat 4-11 MST ... 55
2. Rekapitulasi sidik ragam hasil dan komponen hasil ... 56
3. Korelasi antar parameter ... 57
4. Kelembaban dan suhu dalam rumah plastik ... 58
5. Hasil Analisis sifat fisik dan kimia tanah pada lahan percobaan ... 60
6. Deskripsi Genotipe padi yang diuji ... 61
PENDAHULUAN
Latar Belakang
Padi (Oryza sativa L.) merupakan salah satu sumber pangan utama yang dikonsumsi oleh hampir tiga milyar penduduk dunia. Padi juga merupakan salah satu komoditi pangan yang mampu memenuhi 32% kebutuhan kalori (Sarwar dan Kanif 2005; Bouman et al. 2007).
Luas lahan padi dunia diperkirakan mencapai 148 juta ha, dimana 79 juta ha diantaranya merupakan lahan padi dengan sistem irigasi (irrigated lowland rice), sementara padi tadah hujan dataran rendah (low land rice) dan dataran tinggi (up land rice) masing-masing mencapai 54 juta ha dan 14 juta ha. Dari jumlah total produksi padi dunia, 75% diantaranya dihasilkan dari sistem padi beririgasi, sementara 19% dan 4% masing-masing disumbangkan dari padi dataran rendah tadah hujan dan dataran tinggi tadah hujan (Maclean et al. 2002).
Menurut Dawe (2005) dan Tuong et al. (2004), 56% dari luas lahan tanaman budidaya beririgasi di dunia berada di wilayah Asia, dimana 40-46% merupakan lahan padi dengan sistem irigasi. Kebutuhan air untuk tanaman padi diperkirakan dua hingga tiga kali lebih tinggi dibandingkan tanaman budidaya lainnya. FAO (2004) melaporkan bahwa rata-rata pemakaian air untuk satu kali musim tanam padi berkisar antara 900-2250 mm, sedang menurut Bouman et al. (2007) rata-rata pemakaian air untuk padi sawah mencapai 1300-1500 mm, dimana 25-50% dari asupan air hilang akibat perkolasi dan perembesan.
Tingginya tingkat penggunaan air pada budidaya padi dihadapkan pada masalah kelangkaan air yang turut berdampak pada kehilangan hasil dan penurunan produksi padi. Hanson et al. (1990) melaporkan pengaruh kekeringan telah meluas hingga mencapai 50% luas lahan padi dunia. Pandey dan Bhandari (2007) melaporkan bahwa pada kurun waktu 1970-2002 dampak kekeringan menyebabkan kehilangan hasil padi di Thailand mencapai 0.7 juta ton, China 1.2 juta ton dan India mencapai 5.4 juta ton, sementara dampak kekeringan terhadap luasan lahan padi Di Indonesia pada periode tahun 2001-2006 rata-rata
mencapai 29222 ha dan mengalami puso akibat kekeringan mencapai 3872 ha (Purwani 2006).
Kelangkaan air dan kekeringan disebabkan oleh meningkatnya persaingan dalam penggunaan air antar sektor dan perubahan iklim. Tuong dan Bouman (2003) mengestimasi bahwa hingga tahun 2025 kelangkaan air dan kekeringan akan meluas pada 15-20 juta ha lahan padi di sebagian besar wilayah Asia. Kekeringan yang terus meluas tentu akan berpengaruh terhadap penurunan produksi dan pemenuhan kebutuhan pangan bagi populasi penduduk yang terus meningkat.
Untuk meminimalkan dampak kelangkaan air dan kekeringan terhadap produksi padi maka beberapa upaya yang dapat dilakukan antara lain perlunya mengoptimalkan produksi tanaman per satuan unit evapotranspirasi melalui perbaikan manajemen teknik agronomi, minimalisasi penggunaan air pada tahap persiapan lahan dan persiapan tanaman, menekan kehilangan air akibat perkolasi, perembesan, evaporasi dan aliran permukaan, serta melakukan seleksi varietas/genotipe padi yang toleran terhadap kekeringan (Guera et al. 1998).
Perumusan Masalah
Kekeringan merupakan penurunan kelembaban tanah atau potensial air tanah yang disebabkan oleh evapotranspirasi tanah dan tanaman pada zone perakaran yang dapat menghambat fungsi fisiologis tanaman dan turut berpengaruh terhadap menurunnya pertumbuhan dan hasil tanaman (Takane et al. 1995).
Kekeringan memiliki kisaran yang luas, namun dilihat dari tipe lingkungan tumbuh padi, maka resiko cekaman kekeringan pada sistem budidaya padi tadah hujan umumnya lebih tinggi dibanding padi dengan sistem irigasi, terutama disebabkan oleh pendeknya periode musim hujan (Maclean et al. 2007).
Respon tanaman padi terhadap kekeringan berkaitan dengan waktu dan periode kekeringan yang berasosiasi dengan fase tumbuh (Fukai dan Cooper 1995). Hal ini menunjukkan bahwa stres kekeringan pada fase tumbuh padi yang berbeda akan menunjukkan respon yang berbeda. Tanaman padi memiliki 3 fase tumbuh utama yaitu vegetatif, reproduktif dan pematangan (De Datta 1981;
Takane et al. 1995). Wopereis et al. (1996) menjelaskan stress kekeringan pada fase awal vegetatif berpengaruh pada petumbuhan anakan dan penundaan pembungaan, sedangkan pada fase reproduktif terutama pada masa pembungaan diketahui sangat sensitif terhadap kekeringan, diikuti dengan fase gametogenesis (booting) dan pengisian bulir. Kekeringan pada fase reproduktif juga berpengaruh pada meningkatnya persen gabah hampa dan menurunkan bobot gabah yang menyebabkan penurunan hasil.
Genotipe padi pada lahan kering umumnya lebih resisten terhadap kekeringan dibandingkan genotipe padi sawah (Bouman et al. 2007), walaupun demikian terdapat variasi tingkat resistensi, pertumbuhan dan daya hasil tiap genotipe terhadap kondisi kekeringan. Takane et al. (1995) menjelaskan pengujian dan identifikasi karakter morfofisiologi genotipe padi terhadap kekeringan dan relevansinya dengan pertumbuhan dan hasil perlu dilakukan untuk pengembangan genotipe yang toleran kekeringan. Salah satu pendekatan dalam pengembangan genotipe toleran kekeringan adalah melalui identifikasi karakter morfologi tanaman pada kondisi kekeringan pada sejumlah populasi di suatu lingkungan target (Fukai dan Cooper 1995).
Beberapa pengujian untuk memperoleh varietas toleran kekeringan pada beberapa genotipe padi sawah ataupun gogo telah dilakukan di tingkat lapangan dengan stres kekeringan yang terjadi secara alami (Fukai dan Cooper 1995; Boonjung dan Fukai 1996), namun pengujian genotipe padi pada sistem sawah dengan kondisi kekeringan yang dapat dikontrol hingga kini belum banyak dilakukan. Oleh karena itu perlu dilakukan pengujian untuk mengetahui respon beberapa varietas padi terhadap periode kekeringan pada sistem sawah.
Tujuan Penelitian
Penelitian ini bertujuan:
1. Mengetahui respon beberapa genotipe padi terhadap periode kekeringan pada sistem sawah.
2. Mengetahui tingkat sensitifitas kekeringan beberapa genotipe padi terhadap periode kekeringan pada sistem sawah.
Hipotesis
Hipotesis yang diajukan:
1. Perlakuan kekeringan, genotipe dan interaksinya diduga akan berpengaruh terhadap pertumbuhan dan hasil padi.
2. Pada kondisi cekaman kekeringan genotipe padi gogo akan menunjukkan pertumbuhan dan hasil yang lebih baik dibandingkan genotipe padi sawah.
TINJAUAN PUSTAKA
Cekaman Kekeringan pada Lingkungan Tumbuh Padi
Karakteristik lingkungan tumbuh tanaman padi dapat dikelompokan menjadi 3 tipe yaitu padi dataran rendah dengan sistem irigasi (irrigated lowland rice), padi dataran rendah tadah hujan (rainfed lowland rice) dan padi dataran tinggi tadah hujan (rainfed upland rice) (Bouman et al. 2007). Padi dataran tinggi lebih dikenal juga dengan istilah padi ladang atau padi gogo sementara padi dataran rendah dikenal dengan istilah padi sawah.
Resiko cekaman kekeringan pada sistem budidaya padi tadah hujan umumnya lebih tinggi dibanding padi dengan sistem irigasi, terutama disebabkan oleh pendeknya periode musim hujan. Rata-rata produktivitas padi tadah hujan dataran rendah mencapai 2.3 ton per ha sementara padi dataran tinggi cendrung lebih rendah yaitu 1 ton per ha (Maclean et al. 2002).
Beberapa wilayah di Asia yang mengalami dampak kerusakan lahan tertinggi akibat kekeringan antara lain Di India dengan tingkat kerusakan lahan mencapai 20 juta ha dan Thailand mencapai 7 juta ha. Rata-rata kehilangan hasil padi akibat kekeringan tahunan selama periode waktu 1970-2002 mencapai 5.4 juta ton di India, 1.2 juta ton di China, dan 0.7 juta ton di Thailand (Pandey dan Bhandari 2007). Di Indonesia dampak kekeringan terhadap luasan lahan padi pada periode 2001-2006 rata-rata mencapai 29222 ha dan mengalami puso akibat kekeringan mencapai 3872 ha (Purwani 2006).
Penanaman padi di wilayah Asia umumnya mengikuti pola hujan bimodal oleh karenanya stres kekeringan selama musim tanam dapat dikelompokan menjadi 3 tipe yaitu : stres kekeringan di awal musim tanam (early stress), stres kekeringan tengah musim tanam (Mild-Intermitten stress) dan stres kekeringan akhir musim tanam (late stres) (Chang et al. 1979).
Stres kekeringan pada awal musim tanam (early stress) umumnya terjadi pada masa semai atau pembenihan (Chang et al. 1979), dimana masa kekeringan terjadi bersamaan dengan waktu transplanting. Selain beresiko terhadap bibit muda yang rentan terhadap stres kekeringan, dapat juga memperlambat proses
transplanting sehingga benih yang digunakan telah berumur tua sehingga berpengaruh terhadap menurunnya hasil (Maurya dan O’Toole 1986).
Tipe stres pada tengah musim (mild-stress intermiten) terjadi pada saat periode pertumbuhan anakan sampai pembungaan yang berdampak pada kehilangan hasil yang tinggi. Suplai air yang terbatas selama periode tumbuh ini menyebabkan pelayuan daun, menurunkan indeks luas daun, penutupan stomata dan berdampak pada penurunan berat kering dan hasil padi (Boonjung 1993).
Stres akhir (late stress) umumnya terjadi ketika kultivar berumur panjang karena stres kekeringan yang terjadi bersamaan dengan periode pembungaan dan pengisian gabah di awal musim kering. Pada kondisi stres akhir, kultivar berumur pendek memiliki kemungkinan terhindar dari stres kekeringan (Chang et al. 1979).
Mekanisme Adaptasi Tanaman Padi terhadap Kekeringan
Tanaman padi diketahui lebih peka terhadap kondisi kekeringan dibanding beberapa jenis tanaman sereal lainnya. Tingkat kepekaaan tanaman padi terhadap kekeringan dipengaruhi oleh anatomi daun dan akar yang berasosiasi dengan pola penyerapan dan pelepasan air (Lafitte dan Bennet 2002). Daun padi lebih sempit dan tidak terjadi diferensiasi sel mesofil menjadi jaringan palisade dan parenkim spongi. Jumlah stomata pada daun padi 10 kali lebih banyak dibandingkan jenis rerumputan yang tumbuh di daerah kering. Padi memiliki ukuran stomata yang kecil dan terdapat perbedaan frekuensi stomata antar kultivar akibat pengaruh lingkungan. Untuk jenis padi dataran rendah diperkirakan 30% air ditranspirasikan setelah pembungaan melalui penikel (Bouman et al. 2007).
Umumnya tanaman padi dapat beradaptasi pada kondisi terendam penuh (complete subemergence) antara 3-4 hari walaupun demikian beberapa varietas terutama pada kelompok padi sawah di daerah bercurah hujan tinggi dapat bertahan pada kondisi genangan lebih dari 10 hari (Maclean et al. 2002).
Padi sangat sensitif terhadap periode penggengangan yang pendek. Kondisi kandungan air tanah yang rendah dapat menyebabkan penggulungan daun, mengurangi permukaan luas daun, menurunkan laju fotosintesis dan ukuran sink
sehingga berpengaruh terhadap pertumbuhan dan hasil (Yoshida 1981; Bouman dan Tuong 2001).
Dampak langsung stres kekeringan terhadap tanaman adalah penutupan stomata yang mulai terjadi pada kondisi potensial air daun mencapai -0.75 MPa sehingga menurunkan laju transpirasi secara bertahap (Dingkuhn et al. 1989b). Disamping penutupan stomata, stres kekeringan juga menginduksi penggulungan daun (leaf rolling), penurunan luas daun dan intersepsi cahaya, mempercepat pengguguran daun sehingga mengurangi total fotosintesis dan produksi biomass (Turner et al. 1986a).
Diketahui fase tumbuh padi dari perkembangan penikel hingga masa anthesis adalah fase yang paling peka terhadap stres air (O’Toole 1982). Boonjung (1993) menunjukkan bahwa periode stres kekeringan selama 15 hari setelah perkembangan malai menyebabkan produksi gabah menurun hingga 2% per hari dimana nilai potensial air daun di pagi hari kurang dari -1.0 MPa.
Dilihat dari fase tumbuh maka fase pembungaan terutama pada masa anthesis adalah periode yang sangat sensitif terhadap stres kekeringan karena dapat meningkatan sterilitas spikelet (Cruz dan O’Toole 1984; Ekanayake et al. 1989). Pada kondisi iklim tropika, 13 hari setelah pembungaan akan terjadi pengisian bulir dalam 2 tahap, tergantung pada heterogenitas bulir dalam satu malai dan jumlah malai dalam suatu populasi tanaman (Yoshida 1981), stres air pada masa pengisian bulir dapat membatasi laju fotosintesis namun mobilisasi asimilat dapat terjadi untuk mengkompensasi source sehingga penghambatan pengisian bulir dapat ditekan (O’Toole 1982).
Diketahui terdapat 4 mekanisme kemampuan adaptasi tanaman padi terhadap kekeringan yaitu: menghindari kekeringan (drought escape); menghindari dehidrasi (dehydration avoidance), toleran kekeringan (dehydration tolerance) dan kemampuan memperbaiki sistem tumbuh setelah melewati periode stres kekeringan (drought recovery) (Arrandeau 1989).
Mekansime penghindaran terhadap kekeringan (drougth escape) berkaitan dengan umur dan fenologi tanaman (O’Toole 1982). Kultivar berumur pendek umumya dapat menghindari stres kekeringan dibandingkan kultivar berumur panjang (Arrandeau 1989).
Mekanisme dehydration avoidance berkaitan dengan kemampuan tanaman mempertahankan status air tanaman terutama potensial air daun tetap tinggi dengan cara menyerap air lebih banyak atau menggunakan air yang lebih lambat sehingga air menjadi lebih tersedia selama periode kekeringan berikutnya (Arrandeau 1989). Dua cara ini sangat bermanfaat pada beberapa tanaman budidaya termasuk tanaman padi gogo, sementara pada sistem penanaman padi sawah cendrung lebih banyak faktor pembatas seperti penghambatan pada sistem perakaran pada kedalaman tanah tertentu dan tingkat kehilangan air yang lebih banyak karena perkolasi, perembesan dan evaporasi (Fukai dan Cooper 1995). Penghindaran dari dehidrasi akibat kekeringan dapat terjadi pada kondisi lingkungan padi gogo ataupun padi dataran rendah karena menurunnya kebutuhan evaporasi atau pemanasan yang terjadi di pucuk. Status air tanaman ditentukan oleh keseimbangan air yang diserap melalui sistem akar dan kebutuhan air oleh pucuk. Dengan demikian potensial air tanaman yang tinggi dapat meningkatkan mekanisme pucuk untuk mengurangi kebutuhan air (Fukai dan Cooper 1995).
Mekanisme dehydration tolerance berkaitan dengan kemampuan tanaman meningkatkan metabolisme walaupun potensial air daun rendah. Arrandeau (1989) menunjukkan bahwa terdapat hubungan antara translokasi asimilasi dengan kemampuan toleransi tanaman padi terhadap dehidrasi. Ketika stres kekeringan terjadi selama masa pengisian gabah, tanaman padi yang lebih toleran terhadap dehidrasi biasanya dapat lebih meningkatkan aktivitas metabolisme untuk beberapa tambahan waktu. Ini berarti laju translokasi asimilasi dapat terus berlanjut untuk pengisian gabah. Dibandingkan dengan tanaman sereal lainnya, padi diketahui lebih banyak menyimpan asimilat untuk pengisian gabah (Weng et al. 1982).
Mekanisme drought recovery berkaitan dengan kemampuan tanaman memulihkan pertumbuhan setelah melewati periode kekeringan tertentu (Arrandeau 1989). Mekanisme ini penting ketika kekeringan terjadi pada awal pertumbuhan dan perkembangan tanaman hal ini ditunjukkan pada beberapa genotipe yang mampu menghasilkan lebih banyak anakan dan memproduksi gabah setelah melewati periode kekeringan (Fukai dan Cooper 1995). Kemampuan tanaman untuk memperbaiki sistem metabolisme akibat kekeringan
berhubungan dengan kemampuannya untuk mempertahankan daun tetap hijau selama periode kekeringan. Mempertahankan daun tetap hijau ketika stres kekeringan terjadi selama masa inisasi malai sangat penting karena daun yang tetap hijau dapat memberikan lebih banyak asimilat bagi perkembangan malai dengan demikian produksi spikelet turut meningkat (Lilley dan Fukai 1994b).
Sifat Perakaran dan Hubungannya dengan Cekaman Kekeringan
Tanaman padi, jagung dan sorgum yang tumbuh di daerah dataran tinggi memiliki pola kepadatan dan panjang akar yang sama dan kemampuan menyerap air dapat mencapai kedalaman 60 cm. Walaupun demikian kemampuan penyerapan air pada tanaman padi lebih rendah dibandingkan tanaman sorgum, hal ini menyebabkan padi lebih peka terhadap stres air dibandingkan sorghum (Fukai dan Inthapan 1988; Intahapan dan Fukai 1988). Boonjung (1993) juga menjelaskan perkembangan sistem perakaran padi sangat sensitif terhadap rendahnya kandungan air tanah.
Tanaman padi memiliki variasi genotipe dalam sistem perakaran yang berasosiasi dengan resistensi kekeringan. O’Toole (1982) menjelaskan mekanisme adaptasi tanaman padi terhadap kekeringan yang berkaitan dengan sistem perakaran mencakup tiga hal : 1. Kondisi ketersediaan air tanah yang rendah akan meningkatkan panjang dan kepadatan akar, meningkatkan rasio akar : pucuk dan konduktansi akar; 2. Kondisi air tanah yang rendah menyebabkan peningkatan penetrasi akar pada berbagai faktor pembatas tanah baik itu faktor fisik ataupun kimia tanah; dan 3. Terdapat kemungkinan terjadinya penyesuaian osmotik pada akar.
Tingginya penyesuaian osmotik pada akar memungkinkan tanaman lebih banyak menyerap air tanah. Tanaman akan lebih toleran terhadap kekeringan dan memiliki kemampuan untuk memperbaiki sistem pertumbuhan setelah melewati masa stres kekeringan. Walaupun demikian belum banyak penelitian yang menunjukkan pengaruh penyesuaian osmotik pada akar dan hubungannya dengan resistensi tanaman terhadap kekeringan (Fukai dan Cooper 1995).
Terdapat variasi yang luas diantara galur padi yang memiliki kepadatan panjang akar kurang dari 30 cm. Secara umum galur dengan densitas panjang
akar dibawah 30 cm memiliki sistem perakaran yang lebih panjang (Fukai dan Cooper 1995), lebih lanjut Yosida dan Hasegawa (1982) menjelaskan bahwa rasio panjang akar yang lebih tinggi dibandingkan pucuk adalah indeks resistensi tanaman padi terhadap kekeringan (avoidance) sebab sistem perakaran yang lebih panjang dan lebih padat lebih memiliki kemampuan untuk menyerap air dan laju transpirasi akan lebih rendah dengan rasio akar terhadap pucuk yang kecil.
Hasil skrining yang dilakukan Yosida dan Hasegawa (1982) pada 1081 galur menemukan bahwa rasio akar yang lebih dalam terhadap pucuk cendrung memiliki jumlah anakan yang sedikit. Karakter ini sesuai dengan ciri genotipe padi tradisional lahan kering. Selama sistem perkaran padi tersusun atas akar nodal, galur yang memiliki banyak anakan akan lebih memiliki lebih banyak akar, terutama pada anakan sekunder dan tersier yang tumbuh lebih lambat akan lebih banyak memiliki akar pendek. Hal ini juga menunjukkan bahwa galur yang memiliki sedikit perkembangan anakan berasosiasi dengan tingginya kepadatan perakaran pada kedalaman tanah. Beberapa akar diantaranya memiliki diameter yang luas dan karenanya memiliki pembuluh xylem dengan banyak axial konduktansi.
Penelitian hubungan antara panjang akar pada kedalaman tertentu dan jumlah air yang diekstrak dari suatu permukaan tanah sangat jarang dilakukan. Puckridge dan O’Toole (1981) menemukan bahwa Kultivar Kinandang Patong dengan sistem akar yang dalam dapat mengekstraksi lebih banyak air pada kedalaman 40-70 cm dibandingkan kultivar IR20 dan IR36 yang memiliki akar dangkal. Hasil yang sama juga diperoleh oleh Mambani dan Lal (1983b,c). Lilley dan Fukai (1994) menunjukkan juga bahwa variasi ekstraksi air pada 4 kultivar padi dataran tinggi secara langsung berhubungan dengan kepadatan panjang akar yang tinggi.
Panjang akar berpengaruh juga terhadap tingginya potensial air daun dan menunda terjadinya kematian daun selama kekeringan (Mambani dan Lal 1983a; Cruz dan O’Toole 1985; Ekanayake et al. 1985a). Liley dan Fukai (1994b) menjelaskan bahwa terdapat indikasi bahwa kultivar dengan akar yang panjang memiliki penampilan yang lebih baik dibandingkan kultivar lain terutama pada kondisi stres ditengah periode tumbuh (mild stress), tapi tidak terdapat hubungan
langsung antara total panjang akar dan hasil gabah ketika hanya terdapat satu periode kekeringan yang panjang. Hal ini juga menunjukkan bahwa galur dengan sistem perakaran yang luas dapat lebih bertahan ketika terjadi beberapa periode kekeringan seperti misalnya stres kekeringan intermitten.
Galur dengan panjang akar mencapai 40-80 cm pada daerah dataran tinggi sangat bermanfaat untuk sistem pertanaman padi tadah hujan. Pada penelitian yang dilakukan di daerah dataran rendah dimana stres air diberikan selama 45-75 hari setelah transplanting, hasil gabah pada 30 galur tidak berkaitan dengan karakteristik perakaran pada kondisi sistem pertanaman aeroponik (Ingram et al. 1990). Walaupun demikian pada beberapa lahan tadah hujan dataran rendah dimana tidak dilakukan praktek pelumpuran, pada kondisi tekstur tanah liat berpasir, variasi genotipe panjang akar pada kedalaman tanah akan terekspresi dan akar yang panjang dan luas adalah karakter yang bermanfaat (Fukai dan Cooper 1995).
Galur padi dengan sistem perakaran yang panjang diketahui memiliki sifat toleran terhadap kondisi kemasaman tanah, galur ini diketahui juga memiliki penampilan yang baik pada kondisi kesuburan tanah rendah (Cattivelli et al. 2008). Karakteristik perakaran lain yang dianggap penting dan mempengaruhi laju aliran air dari akar ke pucuk adalah sifat resistensi akar axial. Tanaman padi seringkali tidak mampu mengekstrak air melalui lapisan tanah yang sangat dalam karena meningkatnya resistensi axial yang disebabkan oleh meningkatnya jarak pucuk dan kecilnya diameter akar. Yambao et al. (1992) mengobservasi bahwa terdapat variasi genotipe pada ketebalan akar yang berasosiasi dengan diameter xylem, walaupun peningkatan diameter pembuluh xylem tidak secara langsung meningkatkan resistensi padi terhadap kekeringan.
Gomathinayagam et al. (1989) menjelaskan bahwa pertumbuhan akar seminal dapat digunakan untuk melakukan skrining untuk akar pada tanaman padi terutama yang berkaitan dengan kemampuan resistensi dorongan akar (root push resistance). Tanaman yang memiliki resistensi dorongan akar cendrung mampu mempertahankan potensial air daun tetap tinggi (Ekanayake et al. 1985b). Resistensi dorongan akar dapat digunakan untuk menghitung sistem perakaran
dan hubungannya dengan penapisan terhadap resistensi kekeringan (O’Toole dan Bland 1987).
Pertumbuhan Pucuk dan Cekaman Kekeringan
O’Toole (1982) menjelaskan bahwa terdapat 3 mekanisme adaptasi tanaman terhadap kekeringan dan hubungannya dengan pertumbuhan pucuk yaitu: akumulasi asam amino atau regulator pertumbuhan; mekanisme drought avoindance dan penyesuaian osmotik (osmotic adjustment).
Berkaitan dengan akumulasi asam amino, Dingkuhn et al. (1991a), menemukan bahwa terdapat perbedaan antara genotipe dalam akumulasi asam abcisic (ABA), tapi perbedaan tersebut tidak berhubungan dengan sifat fisiologi suatu galur dibawah kondisi stres air. Disamping itu stres kekeringan juga menginduksi akumulasi prolin yang berkorelasi positif dengan penyesuaian osmotik.
Padi diketahui memiliki sedikit lapisan lilin pada epicuticular dan memiliki konduktansi kutikula yang tinggi dibandingkan jenis serealia lainnya (O’Toole dan Seiber 1979). Hal ini mengindikasikan padi akan mengalami kehilangan air walaupun stomata menutup dan mengakibatkan kematian daun yang lebih cepat.
O’Toole (1982) menjelaskan terdapat variasi genotipe yang luas dalam hal kuantitas epicuticular lilin, walaupun demikian peran epicuticular lilin selama pertumbuhan pada kondisi stres air dan hubungannya dengan proses pemulihan tanaman belum banyak diketahui. Pada penelitian terkini, ditemukan bahwa terdapat variasi konduktansi epidermal pada beberapa galur tapi tidak ada indikasi yang menunjukkan bahwa konduktansi epidermal dapat mempertahankan daun tetap hijau pada periode waktu yang lama (Cattivelli et al. 2008).
Konduktansi difusi dan penggulungan daun merupakan bagian dari mekanisme avoidance kekeringan pada beberapa genotipe (O’Toole dan Cruz, 1980). Variasi konduktansi difusi dan penggulungan daun berkaitan dengan kemampuan tanaman mengekstrak air dari tanah, yang cendrung merujuk pada perbedaan sistem akar. Kemampuan akar mengekstrak air dari tanah turut menentukan status air daun. Galur yang dapat mempertahankan potensial air daun tetap tinggi dapat meningkatkan konduktansi daun dan menurunkan tingkat
penggulungan daun (O’Toole dan Moya 1978). Walaupun demikian hubungan antara penggulungan daun dan potensial air daun bervariasi pada tiap galur dan terdapat kemungkinan lain yang disebabkan adanya pengaruh penyesuaian osmotik, galur dengan penyesuaian osmotik yang tinggi cendrung dapat meningkatkan potensial turgor sel pada kondisi potensial air daun rendah (Turner et al. 1986a; Fukai dan Inthapan 1988). Hubungan ini juga terjadi pada beberapa jenis tanaman serealia lainnya akan tetapi bukti penelitian yang kuat sangat terbatas untuk tanaman padi (Cattivelli et al. 2008).
Penyesuaian osmotik berpengaruh terhadap penggulungan daun pada beberapa galur padi (Hsiao et al. 1984). Dingkuhn et al. (1989a) menemukan adanya korelasi antara penggulungan daun dan potensial air pada kondisi stres di tengah periode pertumbuhan padi (mild stress), yang menunjukkan bahwa penggulungan daun berkorelasi positif terhadap meningkatnya potensial air daun. Hal ini berbeda pada beberapa galur yang memiliki kemampuan untuk mempertahankan potensial air daun tetap tinggi dengan derajat penggulungan daun yang kecil. Secara umum penggulungan daun adalah salah satu strategi tanaman untuk tumbuh dan berhubungan dengan mekanisme avoidance untuk menjaga potensial air daun tetap tinggi.
Penyesuaian osmotik (osmotic adjustment) adalah suatu mekanisme adaptif tanaman dalam merespon efek lingkungan tumbuh yang beragam, seperti kekeringan, salinitas dan temperatur rendah, akumulasi larutan dalam sel dan penurunan potensial osmotik demikian halnya terhadap kondisi stres air (Steponkus et al. 1982; Turner et al. 1986b). Potensial osmotik yang rendah menyebabkan peningkatan turgor sel seiring dengan turunnya potensial air. Penyesuaian osmotik terjadi sangat cepat pada tanaman padi terutama karena stres air yang diukur melalui potensial air daun dibandingkan dengan tanaman sorghum dan jagung (Fukai dan Inthapan 1988). Penyesuaian osmotik pada padi sangat berperan penting terutama pada fase awal periode kekeringan.
Dengan penyesuaian osmotik, tekanan turgor yang tinggi relatif dapat dipertahankan walaupun terjadi pengurangan potensial air di daun (Cutler et al. 1980). Penyesuaian osmotik dapat menunda penggulungan dan kematian daun dapat (Hsiao et al. 1984), walaupun demikian tekanan turgor tidak selalu penting
sebagai faktor penentu respon tanaman terhadap kekurangan air tanah. Pendapat terkini menunjukkan bahwa respon pucuk terhadap kekeringan secara langsung melalui signal akar (root signal) (Ludlow et al. 1989). Hingga kini belum ada pengujian pada tanaman padi dan hubungannya dengan signal akar yang kuat dengan kecepatan respon suatu spesies terhadap kekeringan (Cattivelli et al. 2008).
Steponkus et al. (1982) menunjukkan bahwa terdapat variasi yang kecil untuk penyesuaian osmotik pada 4 kultivar padi yang memiliki penyesuaian osmotik maksimum antara 0.3-0.5 MPa. Pada studi lapangan, terdapat variasi dalam penyesuaian osmotik dengan nilai maksimum 0.5 MPa pada Kultivar padi dataran rendah dan lebih rendah pada kultivar padi dataran tinggi pada 7 kultivar yang berbeda (Turner et al. 1986b). Perbedaan yang ada berkaitan dengan perkembangan pola stres air pada kultivar ini dan kemampuan penyesuaian yang luas pada kultivar dataran rendah berkaitan dengan paparan stres kumulatif yang luas.
Belum banyak bukti dan kajian yang mengindikasikan bahwa variasi genotipe dalam penyesuaian osmotik memiliki pengaruh yang positif terhadap pertumbuhan dan hasil padi. Henderson et al. (1993) menunjukkan adanya indikasi beberapa pengaruh positif penyesuaian osmotik dalam menetralkan rendahnya potensial air daun sehingga tetap mendukung retensi daun tetap hijau (leaf green retention). Karena penyesuaian osmotik pada tanaman padi berlangsung sangat cepat dan penyesuaian osmotik maksimum terjadi selama periode kekeringan, maka penyesuaian osmotik dapat efektif sebagai buffer dalam melawan kematian akibat stres sedang atau stres intermitten (Fukai dan Cooper 1995).
Beberapa argumen menunjukkan bahwa pada beberapa kondisi, penyesuaian osmotik tidak menunjukkan pengaruh yang positif terhadap pertumbuhan tanaman dan hasil gabah (Munns 1988). Dengan demikian, upaya peningkatan resistensi kekeringan, dalam kaitannya dengan penyesuaian osmotik perlu diuji dibawah kondisi stres yang bervariasi, sebelum digunakan sebagai kriteria seleksi pada program pemuliaan tanaman.
Pengaruh mekanisme penyesuaian osmotik terhadap hasil lebih banyak diketahui pada sorghum dibandingkan padi. Beberapa penelitian lapang dengan menggunakan varietas hibrida komersial pada level penyesuaian osmotik yang berbeda menunjukkan adanya hubungan antara tekanan osmotik dan hasil tanaman pada kondisi stres sebelum anthesis atau selama masa pengisian gabah (Wright et al. 1983; Ludlow et al. 1990; Santamaria et al. 1990). Produksi gabah yang dicapai pada kondisi stres air dan pada kondisi air tersedia akan meningkat secara linear disertai meningkatnya penyesuaian osmotik maksimum pada varietas hibrida.
Dibandingkan dengan varietas hibrida yang memiliki penyesuaian osmotik rendah maka varietas yang memiliki penyesuaian osmotik tinggi mampu mengekstrak lebih banyak air, dan terjadi peningkatan pada produksi gabah dan translokasi asimilat selama masa pre-anthesis. Tanaman yang memiliki mekanisme penyesuaian osmotik yang tinggi mampu menghasilkan 0.5 ton per ha produksi gabah dibandingkan tanaman dengan mekanisme penyesuaian osmotik lebih rendah (Fukai dan Cooper 1995). Pada kondisi air yang terbatas, kelompok tanaman yang mampu memproduksi gabah tertinggi memiliki retensi warna hijau daun yang lebih baik selama masa pengisian gabah dan lebih banyak memproduksi gabah pertanaman (Cattivelli et al. 2008).
Dalam suatu penelitian yang dilakukan pada padi yang ditumbuhkan dalam pot dalam lingkungan yang dikontrol menunjukkan bahwa pada umur tanaman 25 hari selama siklus kekeringan menunjukkan kisaran nilai maksimum penyesuaian osmotik antara 0,4-1,7 MPa pada 59 galur dengan latar belakang lingkungan adaptasi yang berbeda (Cattivelli et al. 2008).
Hubungan Ketersediaan Air dan Pertumbuhan Tanaman
Tanaman sering dipengaruhi oleh berbagai bentuk stres seperti kekeringan, temperatur rendah, garam, genangan, panas dan logam berat yang berpengaruh langsung terhadap kondisi pertumbuhan dan perkembangan tanaman (Jaleel et al. 2009). Kekeringan merupakan salah satu bentuk stres yang berkaitan dengan fenomena meteorologi dan berasosiasi dengan rendahnya curah hujan dan ketersediaan air (Jaleel et al. 2009).
Terdapat 3 tipe kekeringan yaitu kekeringan meteorologi, hidrologi, sosial ekonomi dan pertanian. Kekeringan meteorologi berkaitan dengan rendahnya curah hujan selama periode waktu tertentu. Kekeringan hidrologi berkaitan dengan rendahnya ketersediaan air di daerah sungai, danau dan daerah aquifers yang diantaranya dimanfaatkan untuk kepentingan irigasi, wisata ataupun transportasi. Kekeringan sosial ekonomi merupakan refleksi hubungan antara suplai dan kebutuhan air untuk beragam kebutuhan sosial dan ekonomi (WMO 2006). Tipe kekeringan pertanian berkaitan rendahnya ketersediaan air tanah untuk mendukung pertumbuhan tanaman. Kekeringan dalam konsep pertanian juga dapat diartikan sebagai penurunan kelembaban potensial tanah yang disebabkan oleh evapotranspirasi tanah dan tanaman pada daerah perakaran tanaman dan berpengaruh pada pertumbuhan dan hasil (Takane et al. 1995).
Kekeringan berkaitan juga dengan ketersediaan air terutama dalam menunjang pertumbuhan tanaman. Air bagi tanaman berperan sebagai unsur hara, pelarut unsur hara dan penyusun sel tanaman. Ketersediaan air yang rendah secara mendasar menurunkan pertumbuhan, perkembangan dan produksi tanaman (Gardner 1991; Ashari 2006). Tanaman menyerap dari dalam tanah dipengaruhi oleh organ akar dan sifat fisik tanah yang berkaitan dengan gaya-gaya adhesi, kohesi dan gravitasi (Hardjowigeno 2007).
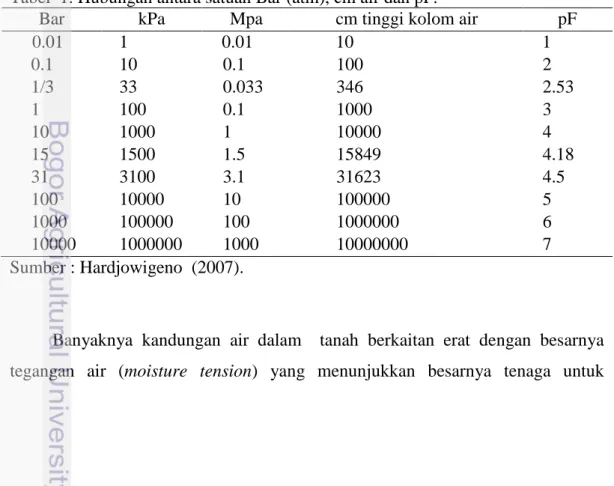
Tabel 1. Hubungan antara satuan Bar (atm), cm air dan pF.
Bar kPa Mpa cm tinggi kolom air pF
0.01 1 0.01 10 1 0.1 10 0.1 100 2 1/3 33 0.033 346 2.53 1 100 0.1 1000 3 10 1000 1 10000 4 15 1500 1.5 15849 4.18 31 3100 3.1 31623 4.5 100 10000 10 100000 5 1000 100000 100 1000000 6 10000 1000000 1000 10000000 7 Sumber : Hardjowigeno (2007).
Banyaknya kandungan air dalam tanah berkaitan erat dengan besarnya tegangan air (moisture tension) yang menunjukkan besarnya tenaga untuk
menahan air tersebut dalam tanah. Tegangan diukur dalam bar, kPa (kilo pascal), atmosfir, cm air, atau logaritma dari cm air yang disebut pF. Satuan bar dan atmosfer sering dianggap sama karena 1 atm = 1.0127 bar (Tabel 1).
Kandungan air pada kapasitas lapang ditunjukkan oleh kandungan air pada titik layu permanen adalah pada tegangan 15 bar (1500 kPa), air yang tersedia bagi tanaman adalah air yang terdapat pada tegangan 1/3 - 15 bar atau 33 kPa-1500 kPa.
Keadaan tanah yang cukup lembab menunjukkan jumlah air terbanyak yang dapat ditahan oleh tanah terhadap gaya tarik gravitasi kondisi ini disebut dengan kapasitas lapang. Air dalam tanah dapat dibedakan dalam 2 kelompok yaitu air higroskopik dan kapiler. Sebagian besar air tersedia dalam tanah dan dapat diserap oleh tanaman disebut air kapiler, sedangkan kondisi dimana kandungan air tanah cukup namun tanaman tidak dapat menyerap air tersebut disebut air higroskopis (Ashari 2006; Hardjowigeno 2007).
Suatu kondisi dimana tanaman tidak lagi mampu menyerap air dan menjadi layu disebut dengan titik layu (wilting point). Respon titik layu dapat bersifat balik namun juga tidak balik dan bervariasi antar spesies. Respon titik layu yang dapat balik berkaitan dengan kemampuan tanaman untuk tetap bertumbuh saat pemberian air dilakukan setelah melewati periode kekeringan tertentu, sementara respon layu permanen umumnya tanaman tidak dapat melakukan aktivitas tumbuh, daun tetap mengalami pelayuan walaupun telah diberikan air (Hardjowigeno 2007), kondisi ini akan berujung pada kematian tanaman.
BAHAN DAN METODE
Lokasi dan Waktu
Penelitian dilakukan di kebun percobaan Institut Pertanian Bogor, Sawah Baru Babakan Darmaga, selama 4 bulan, dari bulan Mei-September 2010.
Bahan dan Alat
Bahan-bahan yang digunakan dalam penelitian ini antara lain bibit padi yang terdiri dari 8 genotipe : IR64 dan Ciherang (padi sawah varietas unggul), IPB-97-F-15 (padi sawah tipe baru), Way Apo Buru (padi type amphibi), Jatiluhur, Silugonggo (padi gogo), Menthik Wangi (padi sawah varietas lokal) dan Rokan (padi hibrida). Alat-alat yang digunakan tensiometer, meteran, termohigrometer, timbangan analitik, oven, trai semai dan kamera.
Rancangan Penelitian
Rancangan penelitian yang digunakan adalah rancangan split plot dengan dua faktor perlakuan yaitu faktor periode kekeringan (petak utama) yang terdiri dari periode pengeringan mulai 3 minggu setelah tranplanting, pengeringan mulai 6 MST, pengeringan mulai 9 MST dan kontrol (tanpa pengeringan). Faktor genotipe (anak petak) yang terdiri dari 8 genotipe yaitu : IR64 dan Ciherang (padi sawah varietas unggul), IPB-97-F-15 (padi sawah tipe baru), Way Apo Buru (padi type amphibi), Jatiluhur, Silugonggo (padi gogo), Menthik Wangi (padi sawah varietas lokal) dan Rokan (padi hibrida). Kombinasi faktor perlakuan periode kekeringan dan genotipe menghasilkan 32 kombinasi perlakuan yang diulang 4 kali sehingga terdapat 128 satuan percobaan.
Model linier rancangan split plot:
Yijk = µ + Kk + αi + δik + βj + γik+ (αβ)ij + εijk
Yijk : Nilai pengamatan faktor perlakuan kekeringan ke-i, dan perlakuan
faktor genotipe ke-j dan blok ke-k
Kk : Pengaruh pengelompokan ke-k; k= 1, 2, 3, 4
αi : Pengaruh faktor kekeringan (petak utama) ke-i =; i=1,…4
βj : Pengaruh faktor varietas (anak petak) ke-j. j=1,2,…8
δik : Komponen galat dari faktor kekeringan (petak utama) γik : Komponen galat dari faktor varietas (anak petak)
(αβ)ij : Pengaruh interaksi antara faktor kekeringan dan faktor varietas
εijk : Pengaruh galat dari interaksi antara faktor kekeringan dan faktor
varietas
Pelaksanaan Penelitian
Persiapan rumah plastik dan petak tanam
Rumah plastik dibangun dengan ukuran 20 m x 15 m, tinggi rangka bangunan kurang lebih 2.2-4.5 m. Dalam rumah plastik dibuat bak tanam dengan ukuran 400 x 300 cm sebanyak 16 bak dengan sekat dinding tembok sedalam 0.8 m. Jarak petak antar perlakuan 35 cm dan jarak petak antar ulangan 35 cm. Pada tiap bak tanam dilengkapi jaringan pipa inlet berdiameter 1 inchi dan outlet berdiameter 2 inchi. Sebelum dilakukan penanaman, terlebih dahulu dilakukan penggenangan selama 5 hari dan 2 kali pengolahan tanah.
Persiapan benih dan penanaman
Untuk menyeragamkan daya kecambah, benih dioven selama 48 jam pada suhu 450 C. Setelah benih dioven, selanjutnya benih dari masing-masing genotipe ditimbang sebanyak 35 g dan direndam dalam air selama 5 jam. Setelah dilakukan perendaman, benih untuk tiap genotipe disemai pada bak semai hingga tanaman berumur 12 hari.
Pada tiap petak percobaan ditanami 8 genotipe, tiap genotipe terdiri dari 30 tanaman dalam 2 barisan tanaman dengan jarak tanam 20 cm x 20 cm, dan jarak tanam antar genotipe 25 cm. Pada kedua sisi petak ditanam tanaman pinggir. Jumlah populasi per petak adalah 270 populasi, dan jumlah total populasi tanaman untuk seluruh percobaan adalah 4320 tanaman.
Pemeliharaan tanaman
Pemiliharaan tanaman dilakukan pemupukan dalam 3 tahap menggunakan pupuk dasar 37.5 kg N/ha, 36 kg P2O5/ha, dan 60 kg K2O/ha diberikan 1 minggu
setelah tanam (MST) dan untuk pemupukan kedua dan ketiga diberikan 37.5 kg N/ha pada 5 MST dan 9 MST. Pengendalian hama dan penyakit dilakukan secara kimia sesuai kondisi dan kebutuhan di lapangan.
Pengaturan pengairan untuk perlakuan pengeringan
Untuk pengaturan perlakuan pengeringan dilakukan penghentian pemberian air pada tiap petak tanam sesuai perlakuan periode pengeringan. Untuk perlakuan periode pengeringan mulai 3 MST (minggu setelah tanam) pemberian air dihentikan saat tanaman berumur 3 MST hingga panen, periode pengeringan 6 MST pemberian air dihentikan ketika tanaman berumur 6 MST hingga panen dan periode pengeringan 9 MST pemberian air dihentikan ketika tanaman berumur 9 MST hingga panen, sedangkan untuk perlakuan kontrol pemberian air terus dilakukan hingga menjelang panen (Gambar 1). Pada penggenangan awal tinggi muka air dipertahankan 2.5 cm dari permukaan tanah.
Gambar 1. Model perlakuan periode kekeringan saat 3 MST, 6 MST, 9 MST dan Kontrol (Warna hitam menunjukkan pemberian air dihentikan).
Parameter Pengamatan
Pertumbuhan vegetatif
Parameter pertumbuhan vegetatif yang diamati adalah tinggi tanaman, jumlah daun dan jumlah anakan dihitung tiap minggu sejak tanaman berumur 2 MST hingga 11 MST. Pengukuran tinggi tanaman dilakukan dari permukaan tanah hingga daun tertinggi, namun saat setelah membentuk malai pengukuran tinggi dilakukan hingga malai tertinggi. Jumlah daun dan jumlah anakan dihitung per rumpun dari tiap tanaman sample dengan menggunakan alat bantu hand counter. Daun yang dihitung adalah daun yang masih hijau. Jumlah anakan dihitung per rumpun dari tiap tanaman contoh.
Hasil dan komponen hasil
Hasil dan komponen hasil yang diamati adalah jumlah anakan produktif, dihitung sejak pembentukan malai berdasarkan jumlah anakan yang menghasilkan malai. Persen pembungaan, ditentukan berdasarkan jumlah tanaman yang menghasilkan bunga pada tiap genotipe. Umur berbunga (hari), umur berbunga diamati pada saat tiap genotipe mulai menghasilkan bunga. Panjang malai (cm), diukur pada saat sebelum panen dari pangkal hingga ujung malai dari 3 malai pada tiap tanaman contoh. Bobot gabah per rumpun (g) ditentukan dengan menimbang total gabah di setiap rumpun setelah kering angin selama 3 hari. Persen gabah hampa (%), dihitung setelah panen dengan membandingkan bobot gabah hampa terhadap gabah isi. Bobot 1000 butir (g), ditentukan dengan dengan menimbang 1000 gabah bernas dari setiap rumpun setelah dijemur 3 hari. Bobot basah dan bobot kering tajuk tanaman (g), bobot basah tajuk dihitung dengan ditimbang pada saat panen dan bobot kering tajuk dihitung dengan menimbang berat kering tanaman setelah dikeringkan dalam oven pada suhu 800 C selama 48 jam.
Indeks panen ditentukan berdasarkan persamaan : Indeks Panen = Bobot kering gabah Bobot kering tajuk
Sensitifitas kekeringan (Drougth sensitivity)
Penentuan sensitifitas kekeringan dilakukan berdasarkan indeks ketahanan kekeringan untuk daya hasil, skor penggulungan daun dan kekeringan daun. Indeks ketahanan kekeringan ditentukan dengan membandingkan bobot gabah per rumpun (g) tiap genotipe pada perlakuan kontrol terhadap bobot gabah per rumpun tiap genotipe pada tiap perlakuan kekeringan. Persamaan Indeks ketahanan kekeringan untuk daya hasil = 1 - (Kn - Hnj) / Kn
Kn : Daya hasil genotipe ke-n (1,2,3,…8) pada perlakuan kontrol
Hnj : Daya hasil genotipe ke-n (1,2,3,….8) pada perlakuan kekeringan ke-j
(1,2,3,4).
Tingkat penggulungan daun dan kekeringan daun dilakukan secara visual mengacu pada sistem standar evaluasi untuk tanaman padi (2002) berdasarkan nilai skor 1-9 (Tabel 2).
Tabel 2. Nilai skor penggulungan daun dan tingkat kekeringan.
Skor Penggulungan daun Skor Kekeringan
0 Daun sehat/daun tidak
menggulung
0 Tidak ada gejala kekeringan 1 Daun mulai menunjukkan
lipatan
1 kekeringan mulai tampak
pada ujung daun
3 Daun melipat- bentuk huruf V 3 Kekeringan pada ujung daun
meningkat hingga ¼ dari bagian daun
5 Daun melipat- bentuk huruf U 5 ¼ - ½ jumlah total daun pada
tanaman kering 7 Pinggiran daun saling
bersentuhan-bentuk huruf O
7 Lebih dari 2/3 dari jumlah total daun pada tanaman mengalami kekeringan penuh
9 Daun menggulung penuh 9 Seluruh bagian tanaman
kering Sumber : IRRI (2002). .
Lingkungan Tumbuh
Parameter lingkungan tumbuh yang diamati adalah suhu, kelembaban harian, potensial air tanah mulai 1 minggu setelah perlakuan kekeringan menggunakan tensiometer. Penentuan potensial air tanah untuk tiap pengamatan dilakukan pada durasi 15-20 menit. Analisis sifat kimia tanah yaitu pH-H2O dan
dapat ditukar (K, Na, Ca, Mg-dd) dan kemasaman dapat tukar (Al dan H-dd) dilakukan sebelum penanaman.
Analisis Data
Data yang diperoleh disajikan dalam bentuk tabulasi dan analisis statistik menggunakan analisis sidik ragam dengan alat bantu SAS versi 9.1. Jika terdapat perlakuan yang berpengaruh nyata berdasarkan uji F, untuk melihat perbedaan antar perlakuan maka dilakukan uji lanjutan DMRT pada α 5%.
0 20 40 60 80 100 120 2 3 4 5 6 7 8 9 10 11 T in g g i ta n am an ( cm )
Minggu setelah tanam
Kontrol 3 MST 6 MST 9 MST
HASIL DAN PEMBAHASAN
Hasil Pertumbuhan Vegetatif
Rekapitulasi sidik ragam pengaruh periode kekeringan, genotipe dan interaksinya terhadap tinggi tanaman, jumlah daun dan jumlah anakan saat 4-11 MST disajikan pada Lampiran 1. Pengaruh periode kekeringan nyata terhadap pertumbuhan tinggi tanaman dan jumlah anakan setelah 5 MST, sedangkan terhadap jumlah daun setelah 7 MST. Pengaruh genotipe nyata terhadap tinggi tanaman, jumlah daun dan jumlah anakan sejak 4 MST hingga 11 MST. Hal ini menunjukkan bahwa terdapat perbedaan respon tinggi tanaman, jumlah daun dan jumlah anakan pada tiap genotipe. Pengaruh interaksi kekeringan dan genotipe nyata hanya terhadap jumlah daun saat 7 MST, 8 MST dan 9 MST, sedangkan terhadap tinggi tanaman dan jumlah anakan tidak nyata.
Pengaruh perlakuan pengeringan terhadap perkembangan tinggi tanaman, jumlah daun dan jumlah anakan tampak nyata seiring dengan meningkatnya level kekeringan. Pengeringan secara nyata menghambat perkembangan tinggi tanaman terutama pada perlakuan pengeringan mulai 3 MST pada durasi 3 minggu setelah perlakuan kekeringan dilakukan, sementara pengaruh penghambatan terhadap jumlah daun terjadi pada durasi 5 minggu setelah perlakuan kekeringan dilakukan.
Gambar 2. Perkembangan tinggi tanaman padi pada berbagai perlakuan periode kekeringan.
0 20 40 60 80 100 120 140 2 3 4 5 6 7 8 9 10 11 T in g g i ta n am an ( cm )
Minggu Setelah Tanam
Ciherang IPB-97-F-15 IR 64 Jatiluhur Menthik Wangi Rokan Silugonggo Way Apo Buru
Pada Gambar 2 dapat dilihat bahwa respon perkembangan tinggi tanaman saat 2-11 MST pada perlakuan periode pengeringan sejak 6 MST, 9 MST dan kontrol tidak menunjukkan perbedaan namun setelah 5 MST perkembangan tinggi tanaman pada perlakuan periode pengeringan sejak 3 MST yang tampak tertekan. Respon hambatan perkembangan tinggi tanaman pada perlakuan pengeringan sejak 3 MST mulai tampak pada saat umur tanaman mencapai 6 MST yang ditunjukkan dengan melandainya pertumbuhan tinggi yang diduga berkaitan dengan perlakuan pengeringan dan berasosiasi dengan penurunan potensial air tanah. Pada saat 6 MST, tinggi tanaman pada pengeringan 3 MST hanya mencapai 92.01 cm berbeda nyata lebih rendah dibandingkan pengeringan 6 MST (97.13 cm) dan 9 MST (98.94 cm) dan kontrol (98.32 cm).
Dari 8 genotipe padi yang digunakan dalam penelitian, terdapat perbedaan perkembangan tinggi tanaman sejak 3 MST. Perkembangan tinggi tanaman tertinggi ditunjukkan oleh genotipe IPB-97-F-15 dan Jatiluhur sedangkan terendah ditunjukkan oleh IR64 (Gambar 3).
Gambar 3. Perkembangan tinggi tanaman 8 genotipe padi
Perkembangan jumlah daun sampai dengan 5 MST pada berbagai perlakuan periode pengeringan tidak menunjukkan perbedaan, namun pada saat umur tanaman mencapai 6 MST perkembangan daun pada periode pengeringan sejak 3 MST tampak lebih tertekan (Gambar 4). Jumlah daun pada periode pengeringan sejak 3 MST hanya mencapai 29.8 daun berbeda nyata lebih rendah dibandingkan pengeringan saat 6 MST (31.4) , 9 MST (32.2) serta
0 10 20 30 40 2 3 4 5 6 7 8 9 10 11 Ju m la h d au n
Minggu Setelah Tanam
Kontrol 3 MST 6 MST 9 MST 0 10 20 30 40 50 2 3 4 5 6 7 8 9 10 11 Ju m lah d au n
Minggu setelah tanam
Ciherang IPB -97-F-15 IR 64 Jatiluhur Menthik Wangi Rokan Silugonggo Way Apo Buru
kontrol (32.3 daun). Pada Gambar 4 dapat dijelaskan juga bahwa setelah 8 MST jumlah daun pada perlakuan pengeringan sejak 3 MST, 6 MST, 9 MST
ataupun kontrol mengalami penurunan karena mengalami kekeringan. Beberapa daun mengalami kekeringan terutama pada perlakuan periode pengeringan sejak 3 MST yang ditunjukkan dengan jumlah daun yang mengering lebih tinggi dibandingkan perlakuan kekeringan 6 MST, 9 MST ataupun kontrol.
Gambar 4. Perkembangan jumlah daun padi pada berbagai perlakuan periode kekeringan.
Perkembangan jumlah daun pada 8 genotipe padi saat 2-11 MST menunjukkan kisaran antara 1.3-2.2 daun. Perkembangan jumlah daun terendah pada tiap minggu ditunjukkan pada genotipe IPB-97-F-15 dan Jatiluhur berbeda nyata lebih rendah dibandingkan IR64, Rokan, Menthik Wangi dan Silugonggo (Gambar 5).
0 2 4 6 8 10 2 3 4 5 6 7 8 9 10 11 Ju m lah an ak an
Minggu setelah tanam
3 MST 6 MST 9 MST Kontrol
Pada Gambar 6 dapat dilihat bahwa perkembangan jumlah anakan pada umur tanaman 2-5 MST baik pada perlakuan periode pengeringan sejak 3 MST, 6 MST, 9 MST ataupun kontrol tidak menunjukkan perbedaan. Ketika umur tanaman mencapai 6 MST, jumlah anakan pada perlakuan pengeringan sejak 3 MST tampak lebih tertekan yaitu hanya mencapai 8.0 anakan berbeda nyata lebih rendah dibandingkan pengeringan sejak 6 MST (9.1), 9 MST (9.3) ataupun kontrol (9.1).
Gambar 6. Pekembangan jumlah anakan padi pada berbagai periode pengeringan.
Pada Gambar 6 dapat ditunjukkan juga bahwa saat umur tanaman mencapai 6 MST, tanaman yang mendapat perlakuan periode pengeringan sejak 3 MST telah mengalami kondisi kekeringan selama 3 minggu, sedangkan tanaman yang mendapat perlakuan periode pengeringan sejak 6 MST dan 9 MST belum berada dalam kondisi cekaman kekeringan. Dengan demikian perkembangan jumlah anakan tampak lebih tertekan pada periode perlakuan pengeringan sejak 3 MST dan berbeda nyata lebih rendah dibandingkan perlakuan periode pengeringan sejak 6 MST dan 9 MST.
Perkembangan jumlah anakan pada 2-6 MST pada 8 genotipe padi menunjukkan variasi antara 2.4-8.2. Hingga 6 MST, jumlah anakan tertinggi ditunjukkan oleh genotipe Silugongogo (11.2), sedangkan terendah ditunjukkan oleh IPB-97-F-15 (4.8) (Gambar 7).
0 2 4 6 8 10 12 1 2 3 4 5 6 7 8 9 10 11 Ju m lah an ak an
Minggu setelah tanam
Ciherang IPB-97-F-15 IR 64 Jatiluhur Menthik Wangi Rokan Silugonggo Way Apo Buru
Gambar 7. Perkembangan jumlah anakan 8 genotipe padi.
Pada Tabel 3 dapat dilihat bahwa sampai dengan 11 MST perkembangan tinggi tanaman, jumlah daun dan jumlah anakan padi pada perlakuan pengeringan sejak 3 MST dan 6 MST sangat tertekan dibandingkan dengan perlakuan pengeringan saat 9 MST dan kontrol. Tinggi tanaman pada periode kekeringan 3 MST dan 6 MST masing-masing hanya mencapai 98.10 cm dan 103.72 cm, berbeda nyata lebih rendah dibandingkan kontrol (110.80 cm). Tabel 3. Rata-rata tinggi tanaman, jumlah daun dan jumlah anakan 8 genotipe
padi saat 11 MST.
Perlakuan Tinggi tanaman
(cm) Jumlah daun (per rumpun) Jumlah anakan (per rumpun) Periode Kekeringan 3 MST 98.10 c 20.4 b 6.2 c 6 MST 103.72 bc 23.5 ab 7.3 b 9 MST 107.35 ab 26.1 a 7.7 ab Kontrol 110.80 a 26.5 a 8.0 a Genotipe IR64 91.06 e 29.5 a 8.7 ab Ciherang 99.12 cd 20.4 bc 7.5 c IPB-97-F-15 124.30 a 19.6 c 3.2 e
Way Apo Buru 98.93 cd 21.1 bc 8.0 bc
Jatiluhur 122.47 a 18.5 c 5.1 d
Menthik Wangi 94.93 de 23.5 b 8.1 bc
Silugonggo 103.12 bc 31.4 a 8.7 ab
Rokan 106.00 b 29.3 a 9.2 a
Keterangan: Angka yang diikuti oleh huruf yang sama pada kolom yang sama untuk masing-masing perlakuan kekeringan dan genotipe tidak berbeda nyata menurut uji DMRT pada taraf α 5%.
Pada Tabel 3 dapat ditunjukkan juga bahwa walaupun perkembangan tinggi
tanaman, jumlah daun dan jumlah anakan pada perlakuan pengeringan sejak 9 MST tampak lebih rendah dibandingkan kontrol namun secara keseluruhan
respon pertumbuhan vegetatif pada pengeringan sejak 9 MST tidak menunjukkan perberbedaan yang nyata dengan kontrol. Hal ini memberikan petunjuk bahwa ketika terjadi kekeringan saat 9 MST pada sistem padi sawah tadah hujan diduga tidak berpengaruh terhadap perkembangan tinggi tanaman, jumlah daun dan jumlah anakan.
Respon tinggi tanaman, jumlah daun dan jumlah anakan pada 8 genotipe sangat bervariasi dan berkisar antara 94.93-124.30 cm. Tanaman tertinggi ditunjukkan oleh genotipe IPB-97-F-15 dan Jatiluhur masing-masing 124.70 cm dan 122.47 cm dan berbeda nyata dibanding genotipe lainnya. Jumlah daun antar genotipe juga menunjukkan variasi antara 18.5-31.4. Jumlah daun terbanyak ditunjukkan Silugonggo (31.4), IR64 (29.5) dan Rokan (29.3), sedangkan terendah pada Jatiluhur (18.5) dan IPB-97-F-15 (19.55). Jumlah anakan terbanyak ditunjukkan oleh Rokan (9.2), kemudian disusul Silugonggo dan IR64 (8.7), sedangkan terendah ditunjukkan oleh IPB-97-F-15 (3.2).
Hasil dan Komponen Hasil
Pengaruh perlakuan periode kekeringan dan genotipe nyata terhadap jumlah anakan produktif, persen pembungaan, panjang malai, persen gabah hampa, bobot gabah per rumpun, bobot 1000 gabah, bobot basah dan bobot kering tajuk serta indeks panen, sedangkan pengaruh interaksi hanya nyata terhadap jumlah anakan produktif, persen pembungaan dan bobot gabah per rumpun (Lampiran 2).
Pada Tabel 4 dapat dilihat bahwa respon jumlah anakan produktif pada sebagian besar genotipe tidak berbeda nyata baik pada perlakuan pengeringan sejak 6 MST ataupun 9 MST. Walaupun demikian respon yang berbeda ditunjukkan oleh genotipe Rokan dengan jumlah anakan produktif yang berkurang secara nyata baik pada pengeringan sejak 3 MST ataupun 6 MST masing-masing 1.1 dan 2.4 dibandingkan dengan pengeringan saat 9 MST (5.8) dan kontrol (5.7). Genotipe Jatiluhur yang diketahui sebagai tipe padi gogo memiliki jumlah anakan produktif yang tidak berbeda nyata baik pada perlakuan
kekeringan sejak 3 MST, 6 MST dan 9 MST masing-masing 4.0, 5.0 dan 5.1 anakan produktif.
Tabel 4. Rata-rata jumlah anakan produktif. persen pembungaan dan bobot gabah per rumpun 8 genotipe padi pada berbagai perlakuan periode kekeringan.
Genotipe Periode Kekeringan
3 MST 6 MST 9 MST Kontrol
Jumlah anakan produktif per rumpun
IR64 3.7 hijk 6.8 bcde 8.3 ab 8.1 ab
Ciherang 5.3 efg 7.1 bcd 7.6 abc 7.7 abc
IPB-97-F-15 1.3 l 3.1 jk 3.5 ijk 3.5 ijk
Way Apo Buru 4.2 ghij 7.8 abc 8.0 ab 7.8 abc
Jatiluhur 4.0 ghij 5.0 fghi 5.1 fgh 5.9 def
Silugonggo 6.3 cdef 7.8 abc 8.8 a 8.7 a
Menthik Wangi 2.9 jk 6.3 cdef 6.9 bcd 7.0 bcd
Rokan 1.1 l 2.4 kl 5.8 def 5.7 def
Persen pembungaan (%)
IR64 21.2 e 80.8 bc 100.0 a 100.0 a
Ciherang 58.7 d 70.2 cd 100.0 a 100.0 a
IPB-97-F-15 10.6 ef 83.7 abc 100.0 a 100.0 a
Way Apo Buru 18.3 ef 92.3 ab 100.0 a 100.0 a
Jatiluhur 70.4 cd 98.1 a 100.0 a 100.0 a
Silugonggo 78.3 bc 100.0 a 100.0 a 100.0 a
Menthik Wangi 19.2 ef 94.2 ab 100.0 a 100.0 a
Rokan 2.9 f 70.2 cd 100.0 a 100.0 a
Bobot gabah per rumpun (g)
IR64 12.23 h 27.99 efg 39.66 cde 43.48 cd
Ciherang 18.38 gh 39.55 cde 39.25 cde 42.46 cd
IPB-97-F-15 11.88 h 38.64 cde 50.22 bc 49.62 bc
Jatiluhur 39.31 cde 40.11 cde 50.73 bc 50.92 bc
Way Apo Buru 17.47 gh 38.06 cde 51.96 bc 60.56 ab
Silugonggo 18.37 gh 44.38 cd 48.58 bc 68.92 a
Menthik Wangi 13.17 h 40.62 cde 37.99 cde 64.87 a
Rokan 12.87 h 24.34 efg 33.21 def 48.28 bc
Keterangan: MST : Minggu setelah transplanting; Angka yang diikuti oleh huruf yang sama tidak berbeda nyata menurut uji DMRT pada taraf α 5%.
Selain menekan perkembangan anakan produktif, periode pengeringan sejak 3 MST juga nyata menurunkan persen pembungaan 8 genotipe padi dibandingkan dengan kontrol. Secara keseluruhan persen pembungaan terendah ditunjukkan oleh Rokan (2.9%) pada perlakuan periode kekeringan sejak 3 MST. Periode pengeringan sejak 6 MST juga menurunkan persen pembungaan namun penurunan persen pembungaan pada genotipe IPB-97-F-15, Way Apo Buru,