FIRSTA NINDA ROSADI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
I
FIRST
SEKOL INSTITUT
TA NIND
LAH PASC T PERTA
BOGO 2013
A ROSAD
CASARJA ANIAN BO
OR 3
DI
Dengan ini saya menyatakan bahwa tesis Studi Morfologi dan Fisiologi Galur Padi (Oryza sativa L.) Toleran Kekeringan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor
Bogor, April 2013
Firsta Ninda Rosadi
NRP A252090081
SUWARNO dan ISKANDAR LUBIS.
Padi (Oryza sativa L.) merupakan salah satu sumber pangan penting, mengingat beras adalah makanan pokok sebagian besar masyarakat Indonesia. Peningkatan produksi padi setiap tahunnya, tidak mampu memenuhi kebutuhan pokok penduduk Indonesia yang juga semakin bertambah setiap tahun. Selain itu, peningkatan suhu udara atmosfer diduga akan sangat mempengaruhi iklim global dunia, seperti kemungkinan meningkatnya frekuensi kekeringan dibeberapa belahan bumi khususnya Asia dan Afrika. Keadaan ini mempengaruhi pertumbuhan tanaman padi sehingga menyebabkan penurunan produksi tanaman padi. Kekeringan merupakan salah satu faktor lingkungan yang besar pengaruhnya terhadap penurunan produksi tanaman. Seleksi terhadap galur-galur padi merupakan salah satu langkah yang baik untuk mengetahui padi yang toleran terhadap cekaman kekeringan. Tujuan penelitian ini adalah untuk mempelajari mekanisme ketahanan padi terhadap kekeringan secara morfologi dan fisiologi, dan untuk mendapatkan galur padi potensial toleran kekeringan.
Penelitian dilakukan di Rumah kaca, Kebun Percobaan Cikabayan, University Farm, IPB, dari bulan Mei sampai bulan September 2010. Analisis kadar air relatif (KAR) daun dan prolin dilakukan di Laboratorium Molecular marker and Spectrophotometry UV-VIS, Fakultas Pertanian, IPB. Analisis klorofil dan kerapatan stomata dilakukan di Laboratorium Micro technique, Fakultas Pertanian, IPB. Rancangan penelitian yang digunakan adalah rancangan petak terpisah (Split-Plot Design) dalam rancangan acak kelompok (RAK) dengan 3 ulangan. Percobaan terdiri atas dua faktor perlakuan yaitu faktor pertama sebagai petak utama adalah cekaman kekeringan P1 (irigasi sampai tinggi genangan 2.5 cm), P2 (kondisi kekeringan yaitu irigasi dilakukan jika potensial air tanahnya mencapai ≤ – 32 kPa), faktor kedua sebagai anak petak yaitu 100 galur padi. 100 galur padi yang digunakan diperoleh dari Pusat Penelitian Padi Indonesia, 10 benih padi ditaburkan kedalam ember/pot yang berisi media tanam, dimana ukuran permukaan tanah di dalam ember diseragamkan 4 cm dari permukaan ember. 3 benih galur padi yang bagus pertumbuhannya dan seragam dipertahankan di dalam ember, yang dilakukan pada 3 Minggu Setelah Tanam (MST). Pemupukan; Urea, SP18, dan KCl, diberikan masing-masing 2.5 g/ember.
Hasil penelitian ini menunjukkan bahwa karakter morfologi yang merespon cekaman kekeringan adalah lebar daun yang lebar, jumlah anakan yang banyak, tinggi tajuk tanaman yang pendek, bobot kering tajuk tanaman lebih berat pada galur padi toleran. Karakter fisiologi yang merespon cekaman kekeringan adalah kandungan air relatif (KAR) daun yang lebih tinggi, prolin yang lebih tinggi, dan kandungan klorofil b yang lebih banyak pada galur padi toleran. Galur padi toleran kekeringan dari seleksi awal berdasarkan nilai persentase penurunan bobot kering tajuk tanaman ada 18 galur, tiga galur yang paling toleran yaitu B11599D-TB-2-24, B 11629C–MR-1-2-1, B11598C-TB-4-1-1.
FIRSTA NINDA ROSADI. Morphological and Physiological Study for Drought Tolerance in Rice (Oryza sativa L.) Line. Supervised by EKO SULISTYONO, SUWARNO and ISKANDAR LUBIS.
Rice (Oryza sativa L.) is one important source of food, since rice is the staple food of the majority of the people in Indonesia. Increased demand for rice due to the rapid population growth was not met by the slower increase of rice production in Indonesia. Increasing rice production every years, can not compensate for the basic needs population in Indonesia. In addition, an increase in atmospheric temperature is expected to greatly affect of climate global world. Such as the possibility of increasing the frequency and level of drought, this situation can affects the growth and decline in rice production. Drought is a major constrain to increase rice production and it is more prominent with the atmospheric temperature and climate change. Selection for drought tolerant lines and understanding the tolerance mechanism were needed to develop drought tolerant rice variety. A research was conducted in the green house of Cikabayan, University Farm, IPB, during May to September 2010, aimed to study the morphological and physiological aspects related to drought tolerance in rice and to identify drought tolerant lines. A split-Plot Design in a randomized complete block design was applied to a pot experiment with two factor treatments of drought stresses as the main plots and rice lines as the suplots. The drought stress treatment were no drought P1 where the standing water was maintained at 2.5 cm depth and with drought stress P2 where irrigation to 2.5 cm water depth was applied when the soil water potential reaching ≤ - 32 kPa. The line treatments including 100 rice lines obtained from the Indonesian Center for Rice Research (ICRR). Ten rice seeds were sowed on the pots filled with soil media up to 4 cm below the top pot surface. Three healthy and uniform seedlings were maintained in each pots by discarding the others at 3 weeks after sowing. Fertilizers; Urea, TSP, and KCL, were applied rating (2.5 g/pot, respectively). Analysis for prolin and relative water content (RWC) in leaf were done in Molecular marker and Spectrophotometry UV-VIS laboratory, IPB. Analysis chlorophyll content and stomatal density in Micro technique laboratory, IPB.
The results of this study indicated that in drought stress treatment, drought tolerant lines exhibit morphological characteristics of broader leaves, more tillers, shorter canopies, and heavier dry canopy weights; and physiological characteristics of higher relative water content (RWC) of leaves as well as higher content of prolin and chlorophyll b. Eighteen drought tolerant lines were identified based on less dry weight reduction by the drought treatment among which B11599D-TB-2-24, B11629C-MR-1-2-1, B11598C-TB-4-1-1 were the most tolerant.
© Hak Cipta milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau peninjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
FIRSTA NINDA ROSADI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Agronomi dan Hortikultura
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
NRP : A252090081
Disetujui oleh Komisi Pembimbing
Dr Ir Eko Sulistyono, MSi
Ketua
Dr Ir Suwarno, MS Dr Ir Iskandar Lubis, MS
Anggota Anggota
Diketahui oleh
Ketua Program Studi Agronomi dan Dekan Sekolah Pascasarjana Hortikultura
Prof Dr Ir Munif Ghulamahdi, MS Dr Ir Dahrul Syah, MScAgr
Puji syukur penulis panjatkan kehadirat Allah SWT atas anugerahNya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Mei 2010 ini ialah cekaman kekeringan dengan judul Studi Morfologi dan Fisiologi Galur Padi (Oryza sativa L.) Toleran kekeringan.
Terimakasih penulis ucapkan kepada Bapak Dr Ir Eko Sulistyono, MSi, Dr Ir Suwarno, MS, Dr Ir Iskandar Lubis, MS selaku pembimbing yang senantiasa memberikan arahan, perbaikan, saran, semangat, dan motivasi selama penulis melakukan penelitian hingga menyelesaikan tugas akhir. Ucapan terima kasih juga penulis ucapkan kepada keluarga dan teman-teman AGH 2009 yang telah memberikan motivasi dan masukan dan doanya selama penulis melakukan penelitian. Selain itu, penulis ucapkan terima kasih kepada Kerjasama Kemitraan Penelitian Pertanian dengan Perguruan Tinggi Negeri (KKP3T) atas biaya penelitiannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, April 2013
Firsta Ninda Rosadi
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan 2
Hipotesis 3
2 TINJAUAN PUSTAKA 3
Botani dan Morfologi Padi Sawah 3
Peranan Air bagi Tanaman 4
Kelembaban Tanah 5
Respon Tanaman terhadap Kekeringan 6
Mekanisme Tanaman terhadap Cekaman Kekeringan 8
3 BAHAN DAN METODE 10 Tempat dan Waktu 10 Bahan dan Alat 10 Rancangan Penelitan 10 Pelaksanaan Penelitian 11
Variabel Pengamatan 12
4 HASIL DAN PEMBAHASAN 16 Kondisi Umum Penelitian 16 Rekapitulasi Sidik Ragam 17 Pengaruh Cekaman Kekeringan terhadap Peubah-peubah yang Diamati 18 a. Peubah morfologi 18
b. Peubah fisiologi 20
Tingkat Toleransi 22
Karakter Morfologi dari Galur Toleran dan Peka 22
Karakter Fisiologi dari Galur Toleran dan Peka 24
Karakter yang Berkaitan dengan Toleransi Kekeringan 25
a. Peubah morfologi 25
b. Peubah fisiologi 26
5 SIMPULAN DAN SARAN 28
Simpulan 28
Saran 28
DAFTAR PUSTAKA 28
LAMPIRAN 35
x
1. Rekapitulasi sidik ragam respon morfologi dan fisiologi galur-galur
galur padi toleran kekeringan 17
2. Pengaruh cekaman kekeringan terhadap peubah morfologi yang
diamati 18
3. Pengaruh cekaman kekeringan terhadap peubah fisiologi yang di-
amati 21
4. Pengelompokan galur padi berdasarkan toleransinya terhadap keke-
ringan 22
5. Karakter morfologi dari galur toleran dan peka 23
6. Karakter fisiologi dari galur toleran dan peka 25
7. Korelasi persentase penurunan bobot kering tajuk dengan peubah
morfologi yang diamati pada 10 MST 27
8. Korelasi persentase penurunan bobot kering tajuk dengan peubah
fisiologi yang diamati 27
DAFTAR GAMBAR
1. Jumlah curah hujan di Drama Bogor dari bulan Maret sampai
bulan November 2010 16
2. Laju pertumbuhan tanaman padi antara tanaman kontrol dan tana- man yang mengalami stres air, (a) panjang daun; (b) lebar daun; (c) tinggi tanaman; (d) jumlah anakan; (e) lebar tajuk tanaman;
(f) tinggi tajuk tanaman 20
3. Laju pertumbuhan antara galur toleran dan peka, (a) panjang daun tanaman; (b) lebar daun tanaman; (c) tinggi tanaman; (d) jumlah
anakan; (e) lebar tajuk tanaman; (f) tinggi tajuk tanaman 24
DAFTAR LAMPIRAN
1. Galur-galur padi yang ditanam 36
2. Denah penelitian 39
3. Rata-rata suhu rumah kaca perlakuan kekeringan I, II, III, dan IV
dari tanggal 17 Juli 2010 sampai 27 Agustus 2010 40
4. Uji kenormalan data
5. Rata-rata suhu harian dalam rumah kaca dari bulan Juli 2010
sampai bulan Agustus 2010 44
6. Rekapitulasi sidik ragam respon morfologi dan fisiologi galur-galur
padi toleran kekeringan 45
7. Peringkat tingkat toleransi terhadap cekaman kekeringan dari 100
Latar Belakang
Padi (Oryza sativa L.) merupakan salah satu sumber pangan penting, mengingat beras adalah makanan pokok sebagian besar penduduk Indonesia. Padi mampu memenuhi 32 % kebutuhan kalori sebagian besar penduduk di Asia (Bouman et al. 2007; Sarwar dan Kanif 2005). Peningkatan kebutuhan beras akibat peningkatan jumlah penduduk yang pesat, tidak diimbangi dengan peningkatan produksi padi.
Perkembangan produksi padi di Indonesia pada tahun 2011 mengalami sedikit peningkatan dibandingkan dengan tahun 2010. Tahun 2011 Indonesia berhasil memproduksi padi sebesar 67.31 juta ton GKG (Gabah Kering Giling), meningkat sebanyak 895.86 ribu ton (1.35 %) dibandingkan tahun 2010. Tahun 2010, hasil produksi padi Indonesia mencapai 66.41 juta ton, meningkat sebanyak 2.01 juta ton (3.13 %) dibandingkan tahun 2009. Peningkatan produksi yang dicapai Indonesia, bukan tidak mungkin Indonesia menjadi eksportir beras terbesar, namun dengan tingkat konsumsi beras yang sangat tinggi di Indonesia yaitu mencapai 140 kg/orang/tahun hal ini masih belum bisa diwujudkan (BPS 2012).
Populasi penduduk Indonesia pada tahun 2025 diperkirakan berkisar 316 juta sampai 350 juta orang, dan diperkirakan akan membutuhkan beras setara 65.9 juta ton GKG (Gabah Kering Giling). Selama periode 2005-2010 permintaan beras akan mengalami peningkatan dari 52.3 juta ton menjadi 55.8 juta ton setara gabah, dari data tersebut dapat disimpulkan bahwa neraca ketersediaan padi berdasarkan trend 2000-2010 menuju ke arah negatif seiring bertambahnya jumlah penduduk (Badan Litbang Deptan 2005).
Peningkatan suhu udara atmosfir diduga akan sangat mempengaruhi iklim global dunia, seperti kemungkinan meningkatnya frekuensi dan tingkat kekeringan di beberapa belahan bumi khususnya Asia dan Afrika (Pitelka dan Rojas 2001). Keadaan ini mempengaruhi pertumbuhan dan menyebabkan penurunan produksi tanaman padi (Hamim 2005). Apalagi di daerah-daerah pertanian padi tadah hujan, keadaan kekeringan yang berkepanjangan akan menurunkan produksi beras sehingga dapat mengganggu ketahanan pangan nasional.
Menurut Syafi (2008) tipe cekaman kekeringan sangat beragam mulai dari radiasi matahari yang diterima tanaman cukup tinggi sampai pada lahan bermasalah yang mengalami defisit air, dan kelembaban udara sangat rendah di lingkungan yang kering. Cekaman kekeringan pada fase vegetatif dapat menurunkan tinggi tanaman, bobot kering akar, dan tajuk (Sunaryono 2002). Cekaman kekeringan dapat menurunkan hasil panen padi 20-25 % dan produksi 50 % (Bouman et al. 2007).
Respon tanaman terhadap kekeringan berkaitan dengan akumulasi asam amino prolin. Prolin merupakan salah satu metabolit yang terbentuk dan terakumulasi pada daun dalam jumlah banyak apabila tanaman mengalami cekaman kekeringan (Syafi 2008). Kadar prolin meningkat pada tanaman alfalfa untuk merespon penurunan potensial air – 0.1 sampai – 0.2 Mpa. Tingginya kandungan prolin pada umumnya berbanding lurus dengan tingkat cekaman kekeringan yang ditunjukkan oleh penurunan potensial air (Iannucci et al. 2002; Syafi 2008).
Seleksi terhadap galur padi yang telah ada merupakan salah satu langkah yang baik untuk mengetahui padi yang toleran terhadap cekaman kekeringan, akan tetapi padi yang toleran terhadap kekeringan belum tentu memiliki sifat unggul lain yang diharapkan. Diperlukan langkah-langkah untuk perbaikan genetika lebih lanjut sesuai dengan yang diinginkan.
Upaya untuk mendukung program pemuliaan tersebut diperlukan informasi yang mendasar mengenai ciri morfologi dan fisiologi yang berkaitan dengan ketahanan terhadap cekaman kekeringan sehingga proses seleksi berjalan secara efektif dan efisien, pengetahuan tersebut juga sangat bermanfaat dalam membantu menentukan strategi pengembangan padi yang toleran terhadap cekaman kekeringan pada masa yang akan datang.
Banyak penelitian yang telah mempelajari berbagai respons morfologi dan fisiologi tanaman yang berkaitan dengan mekanisme ketahanan terhadap kekeringan. Kirkham (1990) menyatakan bahwa toleransi tanaman terhadap cekaman kekeringan dapat melalui beberapa mekanisme, yaitu : 1) melepaskan diri dari cekaman kekeringan (drought escape), 2) bertahan terhadap kekeringan dengan tetap mempertahankan potensi air yang tinggi dalam jaringan (Levitt 1980) atau yang biasa dikenal sebagai mekanisme menghindar dari kekeringan
(drought avoidance) (Jones dan Turner 1978), dan 3) bertahan terhadap
kekeringan dengan potensial air jaringan yang rendah.
Pengujian dan identifikasi karakter morfologi dan fisiologi galur padi terhadap kekeringan dan relevansinya dengan pertumbuhan dan hasil adalah hal penting yang perlu dilakukan untuk pengembangan padi yang toleran kekeringan. Oleh sebab itu, perlu diketahui studi yang mendasar mengenai ciri morfologi dan fisiologi galur-galur padi terhadap cekaman kekeringan.
Tujuan
Penelitian ini bertujuan untuk
1. Mempelajari mekanisme ketahanan padi terhadap kekeringan secara morfologi dan fisiologi
Hipotesis
1. Terdapat keterkaitan antara morfologi dan fisiologi tertentu dengan toleransi terhadap kekeringan
2. Terdapat perbedaan respon galur - galur padi terhadap cekaman kekeringan
2 TINJAUAN PUSTAKA
Botani dan Morfologi Tanaman Padi
Tanaman padi diklasifikasikan ke dalam Divisio Spermatophyta, dengan Sub divisio Angiospermae, termasuk ke dalam Kelas Monocotyledoneae, Ordo adalah Poales, Famili adalah Graminae, Genus adalah Oryza Linn, dan Speciesnya adalah Oryza sativa L (Vaughan 1989). Di Indonesia pada mulanya tanaman padi diusahakan didaerah tanah kering dengan sistim ladang, akhirnya orang berusaha memantapkan basil usahanya dengan cara mengairi daerah yang curah hujannya kurang. Tanaman padi yang dapat tumbuh dengan baik di daerah tropis ialah Indica, sedangkan Japonica banyak diusakan di daerah sub tropika (Mahyuddin dan Hermanto 2001).
Padi (Oryza sativa L.) termasuk golongan tumbuhan Graminae dengan batang yang tersusun dari beberapa ruas. Ruas-ruas ini merupakan bumbung kosong yang ditutup oleh buku dan panjang ruasnya tidak sama. Ruas yang terpendek berada di pangkal batang, ruas yang kedua dan seterusnya lebih panjang dari ruas-ruas yang lebih bawah. Pada buku bagian bawah dari ruas, tumbuh daun pelepah yang membalut ruas sampai buku bagian atas. Tepat pada buku bagian atas ujung daun pelepah meperlihatkan percabangan dimana cabang yang terpendek menjadi ligule (lidah) daun, dan bagian yang terpanjang dan terbesar menjadi helaian daun. Daun pelepah itu menjadi ligule dan pada helaian daun terdapat dua embel sebelah kiri dan kanan yang disebut auricular. Auricular dan ligule yang kadang-kadang berwarna hijau dan ungu dapat digunakan sebagai alat untuk mendeterminasi dan identifikasi suatu varietas (Siregar 1987). Daun pelepah yang membalut ruas yang paling atas batang umumnya disebut daun bendera. Tepat dimana daun pelepah teratas menjadi ligule dan daun bendera, disitulah timbul ruas yang menjadi bulir padi. (De Datta 1981).
Bunga padi adalah bunga telanjang artinya mempunyai perhiasan bunga. Berkelamin dua jenis dengan bakal buah yang diatas. Jumlah benang sari ada 6 buah, tangkai sarinya pendek dan tipis, kepala sari besar serta mempunyai dua kandung serbuk. Putik mempunyai dua tangkai putik dengan dua buah kepala putik yang berbentuk malai dengan warna pada umumnya putih atau ungu (Hanum 2008). Pada dasar bunga terdapat ladicula (daun bunga yang telah berubah bentuknya). Ladicula berfungsi mengatur dalam pembuahan palea, pada waktu berbunga ia menghisap air dari bakal buah, sehingga mengembang. Pengembangan ini mendorong lemma dan palea terpisah dan terbuka (Grist 1975).
palea serta bagian lain akan membentuk sekam atau kulit gabah, lemma selalu lebih besar dari palea dan menutupi hampir 2/3 permukaan beras, sedangkan sisi palea tepat bertemu pada bagian sisi lemma. Gabah terdiri atas biji yang terbungkus sekam. Sekam terdiri atas gluma rudimenter dan sebagian dari tangkai gabah (pedicel) (Badan Litbang 2009).
Batang padi tersusun dari rangkaian ruas–ruas dan diantara ruas yang satu dengan ruas yang lainnya dipisahkan oleh satu buku. Ruas batang padi di dalamnya berongga dan bentuknya bulat, dari atas ke bawah ruas buku itu semakin pendek. Ruas yang terpendek terdapat di bagian bawah dari batang dan ruas–ruas ini praktis tidak dapat dibedakan sebagai ruas–ruas yang berdiri sendiri. (De Datta 1981; Yoshida 1981).
Tanaman padi membentuk rumpun dengan anakannya, biasanya anakan akan tumbuh pada dasar batang. Pembentukan anakan akan terjadi secara bersusun, yaitu: 1) anakan pertama (primer), anakan primer ini tumbuh di antara dasar batang dan daun sekunder, sedangkan pada pangkal batang anakan primer terbentuk perakaran. Anakan primer ini tetap melekat pada batang utama hingga masa pertumbuhan berikutnya. Namun dalam mendapatkan zat makanan, anakan tersebut tidak tergantung pada batang utama sebab memiliki perakaran sendiri. 2) anakan kedua (sekunder), anakan ini tumbuh pada batang bawah anakan primer, yaitu pada buku pertama dan juga membentuk perakaran sendiri. 3) anakan ketiga (tersier), anakan tersier ini tumbuh pada buku pertama pada batang anakan sekunder dengan bentuk yang serupa dengan anakan primer dan sekunder (Yoshida 1981; Siregar 1987).
Peranan Air Bagi Tanaman
Air adalah salah satu komponen fisik yang sangat vital dan dibutuhkan dalam jumlah besar untuk pertumbuhan dan perkembangan tanaman. Sebanyak 85-90 % dari bobot segar sel-sel dan jaringan tanaman tinggi adalah air (Maynard dan Orcott 1987). Gardner et al. (1991) menjelaskan fungsi air bagi tanaman yaitu : (1) sebagai senyawa utama pembentuk protoplasma, (2) sebagai senyawa pelarut bagi masuknya mineral-mineral dari larutan tanah ke tanaman dan sebagai pelarut mineral nutrisi yang akan diangkut dari satu bagian sel ke bagian sel lain, (3) sebagai media terjadinya reaksi-reaksi metabolik, (4) sebagai reaktan pada sejumlah reaksi metabolisme seperti siklus asam trikarboksilat, (5) sebagai penghasil hidrogen pada proses fotosintesis, (6) menjaga turgiditas sel dan berperan sebagai tenaga mekanik dalam pembesaran sel, (7) mengatur mekanisme gerakan tanaman seperti membuka dan menutupnya stomata, membuka dan menutupnya bunga serta melipatnya daun-daun tanaman tertentu, (8) berperan dalam perpanjangan sel, (9) sebagai bahan metabolisme dan produk akhir respirasi, serta (10) digunakan dalam proses respirasi. Peran air yang sangat penting tersebut menimbulkan konsekuensi bahwa langsung atau tidak langsung, kekurangan air pada tanaman akan mempengaruhi semua proses metaboliknya sehingga dapat menurunkan pertumbuhan tanaman.
ke potensial air rendah yang dipengaruhi oleh proses transpirasi (Taiz dan Zeiger 2002).
Proses transpirasi di daun terutama terjadi pada siang hari dan dipengaruhi oleh cahaya matahari. Ketika terjadi proses transpirasi pada tumbuhan, maka tekanan turgor akan mengalami penurunan. Penurunan ini menyebabkan potensial air di daun lebih rendah daripada di akar, sehingga akan mempermudah aliran air di xilem dari akar sampai daun. Peningkatan aliran air ini dibutuhkan untuk tumbuhan sel tanaman. Aliran air ke sel akan mengakibatkan perbesaran dan pemanjangan sel, sehingga sel dapat tumbuh (Kramer dan Boyer 1995). Setiap tanaman harus dapat menyeimbangkan antara proses kehilangan air dan proses penyerapannya, bila proses kehilangan air tidak diimbangi dengan penyerapan melalui akar maka akan terjadi kekurangan air didalam sel tanaman yang dapat menyebabkan berbagai kerusakan pada banyak proses dalam sel tanaman (Taiz dan Zeiger 2002).
Menurut Adisarwanto (2005) jumlah air yang dibutuhkan tanaman untuk pertumbuhannya berbeda-beda pada tiap jenis tanaman. Menurut Yoshida (1981) tanaman padi membutuhkan air sebanyak 180-300 mm/bulan agar berproduksi dengan baik, bila kebutuhan ini tidak terpenuhi maka proses pertumbuhan dan perkembangan tanaman akan terganggu.
Kelembaban Tanah
Kelembaban tanah merupakan air yang terikat secara adsorbtif pada permukaan butir-butir tanah. Penyerapan air oleh perakaran tanaman tergantung pada persediaan kelembaban air dalam tanah (Dwidjoseputro 1985). Penurunan kelembaban tanah menyebabkan penurunan gerakan air dari sistem tanah ke tanaman dan atmosfir. Fenomena tersebut disebut fungsi reduksi (Van Genuchten 1987; Homaee 1999 dalam Sulistyono dan Yanuar 2007) yaitu semakin besar penurunan kelembaban tanah menyebabkan evapotranspirasi semakin besar. Selama interval irigasi, evapotranspirasi mengurangi potensial osmotik dan matrik dari larutan tanah, akibatnya akan menurunkan absorbsi air oleh akar. Kedua faktor tersebut berubah dengan waktu dan tingkat cekaman tergantung dari ketahanan tanaman (Shalhevert 1994 dalam Sulistyono dan Yanuar 2007). Interval pemberian air sangat berpengaruh terhadap kelembaban tanah, baik untuk setiap jenis tanaman maupun fase pertumbuhannya, apabila air diberikan setiap hari, kelembaban tanah masih diatas 30 % volume, sehingga pemberian air tersebut tidak efisien. (Kurnia et al. 2002).
Kapasitas lapang adalah keadaan tanah yang cukup lembab yang menunjukan air terbanyak yang dapat ditahan oleh tanah terhadap gaya tarik gravitasi. Air yang dapat ditahan oleh tanah tersebut terus menerus diserap oleh akar tanaman atau menguap sehingga tanah makin lama makin mengering, saat akar tanaman tidak mampu lagi menyerap air tersebut sehingga tanaman menjadi layu (titik layu permanen) (Dwidjoseputro 1985).
Kandungan air tanah antara kapasitas lapang dan titik layu permanen disebut total air tanah tersedia TAW (Total Available Water). Titik kritis adalah batas minimum air tersedia yang dipertahankan agar tidak habis mengering diserap tanaman hingga mencapai titik layu permanen. Titik kritis ini berbeda untuk berbagai jenis tanaman, tanah, iklim serta diperoleh berdasarkan penelitian di lapangan (Lakitan 2000). Kandungan air antara kapasitas lapang dan titik kritis disebut RAW (Readily Available Water). Perbandingan antara RAW dengan total air tanah yang tersedia dipengaruhi oleh iklim, evapotranspirasi, tanah, jenis tanaman dan tingkat pertumbuhan tanaman (Allen et al. 1998). Evapotranspirasi adalah kombinasi dari dua proses yaitu proses kehilangan air pada permukaan tanah disebut evaporasi dan proses kehilangan air dari tanaman Selama air tersedia, evapotranspirasi akan berlangsung pada laju maksimum yang mungkin dan hanya tergantung pada jumlah energi yang tersedia (Allen et al. 1998).
Respon Tanaman terhadap Cekaman Kekeringan
Cekaman kekeringan merupakan faktor lingkungan yang menyebabkan air tidak tersedia bagi tanaman, yang dapat disebabkan antara lain oleh tidak tersedianya air di daerah perakaran tanaman dan permintaan air yang besar di daerah daun dimana laju evapotranspirasi melebihi laju absorbsi air oleh akar tanaman (Hamim 2004; Jaleel et al. 2009).
Dalam menghadapi cekaman kekeringan, tanaman umumnya melakukan respon dan modifikasi baik bersifat morfologis maupun fisiologis. Secara morfologis, beberapa respon tanaman dalam menghadapi cekaman kekeringan antara lain : penurunan laju pertumbuhan daun, pembentukan lapisan lilin pada epidermis daun, mempercepat penuaan daun, dan peningkatan kerapatan rambut daun (Banziger et al. 2000; Hamim 2007). Secara fisiologis, yaitu penutupan stomata daun, penyesuaian tekanan osmotik, akumulasi asam amino prolin (Hamim 2007). Respon tanaman padi terhadap cekaman kekeringan tergantung pada tingkat, waktu kekeringan, fase tumbuh, organ tanaman dan genotipe. Tingkat cekaman kekeringan berkaitan dengan nilai potensial air tanah, yang berkaitan erat dengan fase tumbuh tanaman padi (Yu et al. 2008).
Menurut Morgan (1984) tipe cekaman kekeringan sangat beragam mulai dari radiasi matahari yang diterima tanaman cukup tinggi sampai pada lahan bermasalah yang mengalami defisit air, dan kelembaban udara sangat rendah di lingkungan yang kering. Kekurangan air secara internal pada tanaman berakibat langsung pada penurunan pembelahan dan pembesaran sel.
kekeringan pada awal fase vegetatif berpengaruh langsung pada penundaan pembungaan dan pematangan, dan kekeringan pada fase reproduktif menyebabkan penurunan hasil terutama karena penurunan bobot gabah dan peningkatan persentasi gabah hampa.
Cekaman kekeringan dapat menghambat pertumbuhan tanaman. Penghambatan pertumbuhan ini salah satunya dapat dilihat pada perluasan daun. Penurunan luas daun merupakan respon pertama tanaman terhadap kekeringan. Keterbatasan air akan menghambat perpanjangan sel yang secara perlahan akan menghambat pertumbuhan luas daun. Luas daun yang kecil mengakibatkan rendahnya transpirasi, sehingga menurunkan laju suplai air dari akar ke daun. Jika kondisi ini dibiarkan terus menerus, lama kelamaan akan terjadi absisi daun (Taiz and Zeiger 2002).
Tumbuhan dapat tumbuh dan berkembang dengan baik salah satunya ditentukan oleh besarnya potensial air. Potensial air (Ψw) merupakan sistem yang
menggambarkan tingkah laku air dan pergerakan air dalam tanah dan tubuh tumbuhan yang didasarkan atas suatu hubungan energi potensial dengan satuan ukur bar atau pascal/pa. Potensial air tanah berhubungan langsung dengan kapasitas lapangan dan titik layu permanen. Energi potensial air tanah (Ψtanah)
pada kapasitas lapang yaitu -0.1 sampai -0.3 bar, sedangkan (Ψtanah) pada titik layu
permanen berkisar -15 sampai -50 bar, tergantung dari tanaman. Adapun pengertian dari kapasitas lapangan adalah air yang tersimpan dalam tanah yang tidak dapat mengalir ke bawah karena gaya gravitasi, sedangkan titik layu permanen yaitu kondisi air tercekam sehingga tanaman akan layu dan tidak akan segar kembali (Gardner et al. 1991).
Air yang tersedia dalam tanah adalah selisih antara air yang terdapat pada kapasitas lapang dan titik layu permanen. Diatas kapasitas lapang air akan meresap ke bawah atau menggenang, sehingga tidak dapat dimanfaatkan oleh tanaman. Di bawah titik layu permanen tanaman tidak mampu lagi menyerap air karena daya adhesi air dengan butir tanah terlalu kuat dibandingkan dengan daya serap tanaman. Cekaman kekeringan pada tanaman disebabkan oleh kekurangan suplai air di daerah perakaran dan permintaan air yang berlebihan oleh daun dalam kondisi laju evapotranspirasi melebihi laju absorbsi air oleh akar tanaman. Serapan air oleh akar tanaman dipengaruhi oleh laju transpirasi, sistem perakaran, dan ketersediaan air tanah (Lakitan 1996; Shen et al. 2009).
Secara umum tanaman akan menunjukkan respon tertentu bila mengalami cekaman kekeringan. Respon tanaman terhadap stres air sangat ditentukan oleh tingkat stres yang dialami dan fase pertumbuhan tanaman saat mengalami cekaman, bila tanaman dihadapkan pada kondisi kering terdapat dua macam tanggapan yang dapat memperbaiki status air, yaitu (1) tanaman mengubah distribusi asimilat baru untuk mendukung pertumbuhan akar dengan menekan pertumbuhan tajuk, sehingga dapat meningkatkan kapasitas akar menyerap air serta menghambat perluasan daun untuk mengurangi transpirasi; (2) tanaman akan mengatur derajat pembukaan stomata untuk menghambat kehilangan air lewat transpirasi (Mansfield dan Atkinson 1990; Lestari 2005).
menutup pada malam hari saat karbondioksida tidak diperlukan. Ukuran stomata berubah-ubah karena sel-sel penutup tersebut mengembang dan mengempis saat air masuk atau keluar secara osmosis (Brodribb dan Holbrook 2003). Pergerakan pori stomata disebabkan oleh perubahan pada volume sel penjaga yang diatur oleh keluar masuknya ion K+ dan ion-ion lain ke sel penjaga selama proses pembukaan dan penutupan stomata. (Moore et al. 1998, Hopkins 1999 dalam Anggraeni 2010).
Cekaman kekeringan menyebabkan tanaman mengalami penurunan laju fotosintesis akibat penurunan konduktansi stomata (Morison dan Lawlor 1999; Hamim 2004; Hamim 2005). Pada kondisi cekaman kekeringan, maka stomata akan menutup sebagai upaya untuk menahan laju transpirasi (Lestari 2005). Berkaitan dengan faktor tanaman, salah satu aspek fisiologi yang secara tidak langsung mempengaruhi pertumbuhan dan daya hasil tanaman pada kondisi cekaman kekeringan adalah kandungan klorofil tanaman. Klorofil merupakan penyerap (absorber) energi radiasi matahari dan sebagai organel yang dapat mengubah energi radiasi menjadi energi kimia (Daubenmire 1974 dalam
Elfarisma 2000). Klorofil merupakan faktor internal tanaman yang sangat mempengaruhi efisiensi dan laju fotosintesis. Tanaman yang memiliki kandungan klorofil tinggi akan sangat efisien dalam penggunaan energi radiasi matahari untuk melaksanakan proses fotosintesis. Tanaman tersebut juga akan mampu memanfaatkan energi matahari semaksimal mungkin (Lawlor 1987).
Klorofil merupakan molekul organik yang kompleks. Struktur dasar klorofil a dan b adalah seperti porfirin, terdiri dari empat cincin pirol dengan atom Mg di tengahnya. Rumus empiris klorofil a dan b adalah C55H72O5N4Mg dan
C55H70O6N4Mg. Perbedaan kedua rumus tersebut terletak pada cincin ketiga,
dimana pada posisi tersebut klorofil a memiliki satu gugus metil (-CH3) sedangkan klorofil b memiliki gugus aldehid (-CHO) (Moher dan Schopfer 1995).
Respon tanaman terhadap kekeringan berkaitan dengan akumulasi asam amino prolin. prolin merupakan senyawa yang umum diakumulasi berbagai jenis tanaman yang mengalami cekaman kekeringan (Hamim 2004), pada beberapa spesies tanaman seperti Amaranthus edulis, hal tersebut tidak terjadi (Hamim 2003). Kandungan prolin yang tinggi pada umumnya berbanding lurus dengan tingkat cekaman kekeringan yang ditunjukkan oleh penurunan potensial air (Iannucci et al. 2002). Oleh karena hal tersebut, besarnya kandungan prolin diduga menunjukkan bahwa tanaman telah mengalami kekeringan yang sangat berat (Hamim 2003).
Mekanisme Tanaman terhadap Cekaman Kekeringan
pada periode air tersedia antara lain dengan meningkatkan jumlah akar, perkembangan daun menjadi lebih sempit dan mempunyai lapisan kutikula tebal termasuk jumlah stomata pada epidermis bagian bawah, dan kemampuan stomata menutup dengan cepat (Courtois dan Lafitte 1999). Arrandeau (1989) menyatakan bahwadrought avoiders berkaitan dengan kemampuan tanaman mempertahankan status air tanaman terutama potensial air daun tetap tinggi dengan cara menyerap air lebih banyak atau menggunakan air yang lebih lambat sehingga air menjadi lebih tersedia (2) tanaman yang mentoleransi kekeringan (drought tolerators) tanaman yang mentoleransi kekeringan mencakup penundaan dehidrasi atau mentoleransi dehidrasi, penundaan dehidrasi mencakup peningkatan sensitivitas stomata
Salisbury dan Ross (1995) menyatakan bahwa pada kondisi potensial air rendah mengakibatkan pembentukan klorofil terhambat, terjadi penutupan Mekanisme menutup dan membukanya stomata tersebut merupakan bentuk penyesuaian tanaman dalam mempertahankan kadar air (mencegah transpirasi yang berlebih) pada kondisi stres air atau cekaman kekeringan, mekanisme penutupan stomata tersebut tergantung dari tekanan turgor sel tanaman, atau karena perubahan konsentrasi karbondioksida (CO2) (Lakitan 2000). Kadar air
relatif (KAR) daun merupakan parameter ketahanan tanaman menghadapi cekaman kekeringan. Proses fotosintesis pada sebagian besar tanaman akan mulai tertekan bila nilai kadar air relatif daun lebih rendah dari 70 persen, sehingga tanaman memerlukan pengaturan dalam tubuhnya (Quilambo 2004).
Prolin merupakan asam amino bebas yang disintesis tanaman dalam jaringan floem, akar dan biji (Simpson 2001). Kandungan prolin di dalam jaringan juga meningkat seiring terjadinya cekaman. Pada kondisi cekaman kekeringan, beberapa tanaman memiliki mekanisme adaptasi berupa kemampuan untuk mensintesis senyawa osmoprotektan (Ronde et al. 2000). Peranan prolin yaitu sebagai osmoregulator atau sebagai protektor enzim tertentu (Yoshiba et al. 1997) dan memproteksi denaturasi protein, dan menjaga kestabilan membran fosfolipid. Oleh karena itu, kandungan prolin akan meningkat seiring dengan lamanya cekaman (Wijana 2001; Ashri 2006).
Biosintesis prolin diawali dengan asam glutamat dan prosesnya melalui intermediet Δ1-pyroline-5-carboxylic (P5CS). Tahapannya adalah gugus γ-caboxyl
pada asam glutamat direduksi ke dalam sebuah aldehid, tahapan ini diperlukan ATP untuk transfer pospat dalam menghasilkan γ-glutamyl phoshate. Selanjutnya dihasilkan glutamic-γ-semialdehyde dengan bantuan elektron donor seperti NADH atau NADPH. Kedua tahapan reaksi tersebut dikatalis oleh enzim komplek, kemudian melalui reaksi spontan glutamic-γ-semialdehyde diproses lebih lanjut menjadi Δ1-pyroline-5-carboxylic dan tahapan akhir dihasilkan prolin dengan bantuan NADH atau NADPH sebagai elektron donor. Enzim yang bekerja dalam pembentukan prolin adalah Δ1-pyroline-5-carboxylic reduktase (Yoshida
3 BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilakukan di Rumah kaca, Kebun Percobaan Cikabayan, University Farm, IPB, dari bulan Mei sampai bulan September 2010. Analisis kadar air relatif (KAR) daun dan prolin dilakukan di Laboratorium Molecular
marker and Spectrophotometry UV-VIS, Departemen Agronomi dan Hortikultura,
Fakultas Pertanian, IPB. Analisis klorofil dan kerapatan stomata dilakukan di Laboratorium Micro technique, Departemen Agronomi dan Hortikultura,Fakultas Pertanian, IPB.
Bahan dan Alat
Bahan-bahan yang digunakan dalam penelitian ini adalah 100 galur padi (berasal dari Balai Besar Penelitian Tanaman Padi, Muara, Bogor dan Balai Penelitian Tanaman Padi Sukamandi, Jawa Barat, tersaji pada Lampiran 1.), ember plastik (diameter 30 cm, tinggi 20 cm), media tanam yaitu tanah yang diambil dari lahan sawah baru IPB bogor (Latosol). Alat-alat yang digunakan antara lain termometer (alat pengukur suhu), Tensiometer (alat pengukur potensial air tanah), Spektrofotometer (alat untuk pengujian kandungan prolin dan klorofil), label, jaring (untuk melindungi padi dari serangan burung), timbangan analitik, mikroskop, oven, dan alat-alat tulis lainnya.
Rancangan Penelitian
Rancangan penelitian yang digunakan adalah rancangan petak terpisah (Split-Plot Design) dalam rancangan acak kelompok (RAK) dengan 3 ulangan. Percobaan terdiri atas dua faktor perlakuan yaitu faktor pertama sebagai petak utama adalah cekaman kekeringan yaitu P1 (irigasi sampai tinggi genangan 2.5 cm), P2 (kondisi kekeringan yaitu irigasi dilakukan jika potensial air tanahnya mencapai ≤ – 32 kPa, (Suhendar 2010)), faktor kedua sebagai anak petak yaitu 100 galur padi. Sehingga terdapat 600 unit percobaan, setiap unit percobaan terdapat 3 benih galur padi (denah percobaan tersaji pada Lampiran 2).
Model aditif linier yang digunakan untuk penelitian ini adalah : Yijk = µ + Bk + Pi + αik + Vj + (PV)ij + ijk
Keterangan :
Yijk = Nilai pengamatan pengaruh cekaman kekeringan (stres air) taraf ke-i
dan faktor galur padi taraf ke-j pada ulangan ke-k. µ = Nilai tengah umum.
Bk = Pengaruhblok ke-k.
Pi = Pengaruh faktor cekaman kekeringan (stres air) taraf ke-i.
αik = Pengaruh sisa untuk petak utama taraf ke-i pada ulangan ke-k.
Vj = Pengaruh faktor galur padi taraf ke-j.
(PV)ij = Pengaruh interaksi antara cekaman kekeringan pada taraf ke-i dan galur
ijk = Pengaruh sisa karena pengaruh cekaman kekeringan taraf ke-i dan
pengaruh galur padi taraf ke-j pada ulangan ke-k.
Data yang diperoleh selanjutnya dianalisis dengan analisis sidik ragam (Anova), apabila berpengaruh nyata akan dilanjutkan dengan uji beda nyata terkecil (BNT) pada taraf nyata 5 %. Uji t untuk mengetahui perbedaan nilai tengah antara galur toleran, sedang dan peka. Uji korelasi untuk mengetahui karakter yang berkaitan dengan toleran kekeringan dengan menggunakan SAS 9.13.
Pelaksanaan Penelitian
Persiapan media tanam dan penanaman galur – galur padi
Persiapan media tanam dilakukan selama 7 hari. Media tanam diambil dari Kebun Percobaan, Sawah Baru Institut pertanian Bogor (IPB), lalu dimasukkan ke dalam ember dan disusun sesuai rancangan penelitian di dalam rumah kaca Kebun Percobaan Cikabayan, University Farm, IPB. Penjenuhan media tanam dilakukan dengan diirigasi sampai air mencapai permukaan ember, lalu permukaan tanah di dalam ember di seragamkan (4 cm dari permukaan ember), Penanaman galur-galur padi dilakukan pada tanggal 9 Juni 2010, kemudian galur-galur-galur-galur padi ditanam di dalam ember sebanyak 10 benih per ember yang akan dijarangkan sebanyak 2 kali, penjarangan pertama sebanyak 5 tanaman per ember saat 2 minggu setelah tanam (MST), penjarangan kedua menjadi 3 tanaman per ember saat 3 MST. Pemeliharaan tanaman
Untuk pemeliharaan tanaman dilakukan pemupukan dalam 2 tahap menggunakan pupuk dasar Urea, KCl, dan SP18 masing-masing (2.5 g/ember) diberikan 1 minggu setelah tanam (MST) dan pemupukan kedua yaitu Urea dengan dosis yang sama diberikan 7 MST. Pengendalian hama dan penyakit dilakukan 1 kali seminggu. Intensitas serangan hama meningkat maka pengendalian hama dilakukan 2 kali seminggu.
Pengaturan pengairan untuk perlakuan kekeringan
Untuk pengaturan perlakuan kekeringan, pemberian air pada tiap ember disesuaikan dengan kondisi perlakuan. Untuk (P1) pemberian air terus dilakukan hingga 11 MST dan penggenangan tinggi muka air dipertahankan 2.5 cm dari permukaan tanah. Untuk kondisi kekeringan (P2) pengairan dilakukan jika potensial air tanah mencapai ≤ – 32 kPa. Pengukuran potensial air tanah dilakukan dengan Tensiometer, sampel diambil secara acak berdasarkan tampak visual yaitu penggulungan daun dan dengan melihat kondisi tanah tanaman yang mengalami kekeringan (P2). Setiap 1 ulangan diambil 10 sampel/ember, sehingga jumlah sampel untuk pengukuran potensial air tanah 30 sampel. Perlakuan kekeringan diberikan setelah tanaman berumur 4 minggu setelah tanam (MST), sebelum diberikan perlakuan, semua ember diairi sampai 1 cm dari permukaan tanah.
dilakukan irigasi tanggal 24 Juli 2010, lalu diirigasi kembali pada tanggal 6 Agustus 2010 (14 hari kering). Pengairan untuk perlakuan kekeringan III dilakukan irigasi pada tanggal 6 Agustus 2010, lalu dibiarkan kondisi tanahnya kering dan diirigasi kembali pada tanggal 16 Agustus 2010 (11 hari kering). Pengairan untuk perlakuan kekeringan IV dilakukan irigasi kembali pada tanggal 16 agustus, lalu kondisi tanah dibiarkan kering lalu diirigasi kembali tanggal 27 Agustus (12 hari kering). Perbedaan jumlah hari kering pada perlakuan kekeringan dipengaruhi oleh suhu rata-rata bulan Juli dan bulan Agustus 2010 (rata-rata suhu rumah kaca perlakuan kekeringan I, II, III dan IV dari tanggal 17 Juli 2010 – 27 Agustus 2010 tersaji pada Lampiran 3.)
Variabel Pengamatan
A. Morfologi
Peubah morfologi yang diamati mencakup : 1. Panjang daun (cm)
Pengamatan panjang daun, diukur pada daun terpanjang. Pengamatan dilakukan pada 2 minggu setelah perlakuan sampai 10 MST (minggu setelah tanam) dan dilakukan setiap 2 minggu sekali.
2. Lebar daun (cm)
Pengamatan lebar daun dilakukan pada daun terlebar, pengamatan dilakukan pada 2 minggu setelah perlakuan sampai 10 MST dan dilakukan setiap 2 minggu sekali.
3. Tinggi tanaman (cm)
Pengamatan dilakukan 2 minggu setelah perlakuan sampai 10 MST. Dilakukan setiap 2 minggu sekali. Tinggi tanaman diukur dari permukaan tanah sampai ujung daun tertinggi.
4. jumlah anakan per rumpun
Pengamatan dilakukan 2 minggu setelah perlakuan sampai 10 MST, dan dilakukan setiap 2 minggu sekali dengan cara menghitung jumlah anakan yang terbentuk dalam satu rumpun.
5. Tinggi tajuk tanaman (cm)
Pengamatan tinggi tajuk tanaman dilakukan saat 2 minggu setelah perlakuan sampai 10 MST, dan dilakukan setiap 2 minggu sekali. Tinggi tajuk tanaman diukur dari permukaan tanah sampai permukaan tajuk tanaman.
6. Lebar tajuk tanaman (cm)
Pengamatan lebar tajuk dilakukan saat 2 minggu setelah perlakuan sampai 10 MST, dan dilakukan setiap 2 minggu sekali. Lebar tajuk tanaman diukur dari ujung daun yang satu ke ujung daun yang lain berdasarkan diameter tanaman. 7. Bobot kering tajuk (g)
B. Fisiologi
Pemilihan jumlah galur padi yang menjadi sampel untuk peubah fisiologi dipilih berdasarkan pengamatan visual dari 4 MST – 6 MST. Jumlah galur padi yang menjadi sampel peubah fisiologi yaitu 6 galur (galur 71 dan 95 = galur toleran; galur 14 dan galur 78 = galur sedang; galur 82 dan galur 91 = galur peka). 1. Kandungan klorofil (mg/g)
Klorofil a, klorofil b dan klorofil total galur-galur padi pada tanaman kontrol dan tanaman stres air. Analisis klorofil dan pengambilan sampel daun dilakukan pada 8 MST atau pada saat akhir periode perlakuan kekeringan II, sampel daun yang diambil yaitu pada daun kedua dari atas. Cara kerjanya yaitu: daun tanaman padi dipotong kecil lalu ditimbang menggunakan timba- ngan analitik seberat 0.05 g dan dimasukkan dalam mortar poselen, sampel
daun dihaluskan dengan cara menggerus sambil ditetesi aseton 80 % secukupnya, lalu supernatan dipindahkan kedalam labu ukur 100 ml melalui kertas saring. Prosedur diatas diulangi hingga warna hijau hilang (seluruh klorofil telah terekstrasi) kemudian tambahkan aseton 80 % kedalam labu ukur sampai mencapai 100 ml, setelah itu larutan tersebut diambil 5 ml dan dimasukkan kedalam labu ukur 50 ml dan diencerkan sampai volume 50 ml, lalu ekstraks tersebut dimasukkan kedalam micro tube 2 ml, dan di-centrifuse
dengan kecepatan 500 rpm selama 10 menit, pengukuran absorban menggunakan spektrofotometer UV-Visible pada panjang gelombang ( ) 645 dan 663 nm. Untuk menghitung kandungan klorofil, digunakan rumus sebagai berikut (Yoshida 1979)
Klorofil a = 0.0127 x A663-0.00269 x A645
Klorofil b = 0.0229 x A645-0.00468 x A663
Klorofil total = 0.0202 x A645 + 0.00802 x A663
A663 = Absorbansi pada 663 nm
A645 = Absorbansi pada 645 nm
2. Kadar air relatif (KAR) daun (%)
Pengamatan dilakukan saat 11 MST atau pada saat akhir periode perlakuan kekeringan IV, sampel daun yang diambil yaitu pada daun kedua dari atas dengan cara nilai KAR diukur dengan membuat potongan daun berbentuk segiempat dengan ukuran 1x1 cm. Sepuluh potongan daun dari tanaman yang mendapat perlakuan ditimbang dan diberi notasi (BP). Selanjutnya potongan daun tersebut segera direndam dengan air destilata di dalam botol film yang telah dilapisi alumunium foil untuk menghindari cahaya, dan ditempatkan di dalam ruang bersuhu 4oC selama 12 jam agar sel-selnya menyerap air dan memiliki turgor penuh. Setelah 12 jam diangkat dari rendaman, dilap dengan kertas tisu agar air di bagian luarnya kering lalu ditimbang kembali dan diberi notasi (BT). Setelah ditimbang potongan daun di bungkus aluminium foil dan dioven pada suhu 80oC selama 2x24 jam untuk mendapatkan berat kering (BK) (Barr dan Weatherley 1962).
Nilai KAR dihitung dengan menggunakan rumus :
3. Kandungan prolin (mg/g)
Analisis kandungan prolin dilakukan saat 11 MST atau pada saat akhir periode perlakuan kekeringan IV, sampel daun diambil pada daun kedua dari atas, dengan menggunakan metode modifikasi Bates (1973). Tahapan kerjanya sebagai berikut : Daun tanaman diambil dan dimasukkan kedalam cool box, selanjutnya daun dihaluskan dengan nitrogen cair, sehingga terbentuk serbuk daun dengan berat ± 100 mg (0.1 g) kemudian dimasukan kedalam tabung ependaf 1.5 ml (selama pengerjaan analisis sample disimpan pada suhu -20oC). Lalu ditambahkan 1.3 ml asam sulfosalisilat 3 % sambil diaduk sampai homogen dengan vortex lebih kurang 15 menit/sample, dan disentrifuse selama 5 menit. Filtrat dipipet sebanyak 2 ml, kemudian ditera hingga 10 ml lalu dimasukan ke dalam tabung reaksi dan ditambahkan 2 ml asam ninhidrin dan 2 ml asam asetat glasial, kocok sampai semuanya tercampur sempurna. Selanjutnya, direaksikan dalam penangas air selama 60 menit, kemudian reaksi dihentikan dalam ice bath dan ditambahkan 4 ml toulen sambil diaduk dengan menggunakan vortek selama 10-15 detik. Lapisan chomophore yang terbentuk dipipet sebanyak 3 ml kemudian dimasukan ke dalam tabung reaksi. Selanjutnya, diukur absorbansinya pada panjang gelombang 520 nm dengan toluen sebagai blanko. Konsentrasi prolin ditentukan dari kurva standar dan dihitung berdasarkan berat segar dengan rumus sebagai berikut :
Prolin = [( g prolin/ml) x ml toluen]/[ml tera/ml contoh] gram contoh
= g prolin/ g bobot segar 4. Kerapatan stomata
Pengamatan kerapatan stomata dilakukan saat 8 MST atau pada saat akhir periode perlakuan kekeringan II, sampel daun diambil pada daun kedua dari atas.
Adapun prosedurnya adalah sebagai berikut :
1. Kuteks dioleskan pada permukaan atas maupun bawah daun yang akan diamati stomatanya. Kuteks dioleskan sepanjang 2 cm x 2 cm. Kemudian kuteks dibiarkan hingga mengering.
2. Setelah kuteks mengering, isolasi bening ditempelkan pada daun yang telah diolesi kuteks tersebut. Isolasi ditekan agar menempel sempurna. 3. Isolasi kemudian dilepaskan dari daun lalu ditempelkan pada kaca
preparat.
4. Pengamatan stomata dilakukan menggunakan mikroskop, dengan pembesaran 40x dan diameter bidang pandang 0.5 mm (sehingga didapat luas bidang pandang = ¼ x 33.14 x (0.5)2= 0.19625 mm.
C. Lingkungan tumbuh
Pengamatan lingkungan tumbuh yang diamati yaitu suhu harian di dalam rumah kaca mulai bulan Juli (4 MST) sampai bulan Agustus 12 MST.
D. Uji kenormalan data dan analisis data
Sebelum data diolah secara statistika, data mentah semua variabel pengamatan diuji kenormalan datanya. Hasil uji kenormalan data morfologi dan fisiologi disajikan pada Lampiran 4.
Analisis data yang dilakukan yaitu : 1. Analisis ragam
Analisis ragam digunakan untuk mengetahui apakah cekaman kekeringan, galur dan interaksi cekaman kekeringan dengan galur berpengaruh terhadap peubah – peubah yang diamati. Semakin rendah nilai Pr >F (peluang F tabel lebih besar dari F hitung) artinya perlakuan semakin berpengaruh terhadap peubah yang diamati. Jika Pr >F nilainya < 0.05 atau < 0.01 maka perlakuan berpengaruh nyata atau sangat nyata terhadap peubahnya.
2. Uji beda nyata terkecil (BNT)
Jika analisis ragam nyata, maka pembandingan antar nilai tengah dilakukan dengan uji BNT pada taraf 5 %.
3. Uji t
Mekanisme toleransi terhadap cekaman kekeringan dipelajari dengan menggunakan Uji t dengan membandingkan karakter morfologi atau fisiologi masing – masing antara galur toleran, sedang, dan peka.
4. Uji korelasi
Mekanisme toleransi terhadap cekaman kekeringan juga diuji dengan uji korelasi. Uji korelasi dilakukan antara peubah persentase penurunan bobot kering tajuk tanaman dengan peubah-peubah morfologi dan fisiologi.
E. Tingkat toleransi tanaman
Berdasarkan definisi toleransi, tanaman yang toleran yaitu tanaman yang mampu/bertahan hidup pada kondisi tercekam kekeringan dan mampu menghasilkan produksi yang lebih baik daripada tanaman peka atau terjadi sedikit penurunan hasil. Pada penelitian ini, komponen produksi yang didapat yaitu bobot kering tajuk (BKT) tanaman sehingga toleransi tanaman dihitung berdasarkan persentase penurunan BKT tanaman kontrol dengan bobot kering tajuk tanaman stres (Chozin 2006), dengan rumus :
Persentase penurunan bobot kering tanaman = – BKTstres %
Ket : BKTkontrol = Bobot kering tanaman pada kondisi kontrol
BKTstres = Bobot kering tanaman pada kondisi cekaman kekeringan
4 HASIL DAN PEMBAHASAN
Kondisi Umum Penelitian
Penelitian ini dimulai pada bulan Mei sampai September 2010, penanaman dilakukan pada 9 Juni 2010. Lokasi penelitian yaitu di rumah kaca University Farm Cikabayan, Dramaga Bogor. Secara umum, kondisi tanaman padi pada awal pertumbuhan baik, namun pada saat tanaman berumur 10 MST (Minggu Setelah Tanam) ada tanaman yang mati yaitu galur 16, ulangan ke-2 pada tanaman kontrol dan pada 12 MST, ± 90 % dari 600 jumlah tanaman, tanaman diserang hama wereng coklat (Nilaparvata lugens Stal). Data pada pengamatan morfologi dan fisiologi tidak terganggu karena pengamatan morfologi dilakukan pada saat 4 sampai 10 MST, sedangkan pengamatan fisiologi dilakukan pada saat 8 dan 11 MST. Terjadinya serangan hama, karena selama penelitian, terjadi perubahan iklim yang ekstrim yaitu perubahan musim kemarau dan musim hujan yang tidak menentu, dimana faktor iklim merupakan salah satu faktor yang dapat menyebabkan meningkatnya populasi hama wereng coklat pada tanaman padi. Hal ini sesuai dengan penelitian (Hutapea 2011; Ahmadi 2011) yang menyatakan bahwa periodisitas timbulnya suatu hama erat hubungannya dengan periodisitas curah hujan tahunan dan perubahannya. Dalam rumah kaca, suhunya lebih tinggi dan kelembaban rendah dibandingkan suhu dan kelembaban diluar rumah kaca sehingga berpengaruh terhadap tumbuhnya serangan hama. Data jumlah curah hujan di dramaga dari bulan Maret – November 2010 (tersaji pada Gambar 1).
.
Gambar 1 Jumlah curah hujan di Dramaga Bogor dari bulan Maret sampai bulan November 2010.
Gambar 1 dapat dilihat bahwa perubahan jumlah curah hujan dari bulan Maret sampai bulan April menurun drastis dari 414.5 mm menurun menjadi 42.9 mm. Pada bulan Juli jumlah curah hujan mulai terus meningkat sampai bulan September mencapai 601 mm. Akibat perubahan periodisitas curah hujan, diduga menyebabkan meningkatnya serangan hama.
Kondisi lingkungan di dalam rumah kaca yang digunakan untuk penelitian sangat berbeda dengan keadaan di lapang. Rata-rata suhu harian di dalam rumah kaca cukup tinggi terutama pada siang hari. Suhu dalam rumah kaca dari bulan
414.5
42.9
330.9 303.4 270.4 477.6
601
436.2 284.3
0 200 400 600 800
MAR APR MEI JUN JUL AGS SEP OKT NOP
Jumlah curah
hujan/mm
Bulan
Juli sampai Agustus pada siang hari (pukul 12.00 WIB) berkisar antara 31.5 oC – 41.5 oC. Rata- rata suhu harian di dalam rumah kaca dari bulan Juli sampai bulan Agustus 2010 disajikan pada Lampiran 5).
Rekapitulasi Sidik Ragam
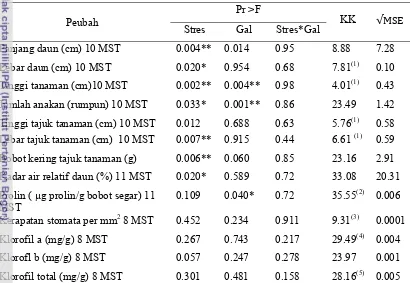
Rekapitulasi hasil sidik ragam respon morfologi dan fisiologi galur-galur padi toleran kekeringan disajikan pada Tabel 1.
Tabel 1 Rekapitulasi sidik ragam respon morfologi dan fisiologi galur-galur padi toleran kekeringan
Peubah
Pr >F
KK √MSE Stres Gal Stres*Gal
Panjang daun (cm) 10 MST 0.004** 0.014 0.95 8.88 7.28 Lebar daun (cm) 10 MST 0.020* 0.954 0.68 7.81(1) 0.10 Tinggi tanaman (cm)10 MST 0.002** 0.004** 0.98 4.01(1) 0.43 Jumlah anakan (rumpun) 10 MST 0.033* 0.001** 0.86 23.49 1.42 Tinggi tajuk tanaman (cm) 10 MST 0.012 0.688 0.63 5.76(1) 0.58 Lebar tajuk tanaman (cm) 10 MST 0.007** 0.915 0.44 6.61 (1) 0.59 Bobot kering tajuk tanaman (g) 0.006** 0.060 0.85 23.16 2.91 Kadar air relatif daun (%) 11 MST 0.020* 0.589 0.72 33.08 20.31 Prolin ( µg prolin/g bobot segar) 11
MST
0.109 0.040* 0.72 35.55(2) 0.006
Kerapatan stomata per mm2 8 MST 0.452 0.234 0.911 9.31(3) 0.0001 Klorofil a (mg/g) 8 MST 0.267 0.743 0.217 29.49(4) 0.004 Klorofl b (mg/g) 8 MST 0.057 0.247 0.278 23.97 0.001 Klorofil total (mg/g) 8 MST 0.301 0.481 0.158 28.16(5) 0.005
Keterangan: (Stres) = Cekaman kekeringan; (Gal) = Galur padi; (Stres*Gal) = Interaksi cekaman kekeringan dengan galur padi; (KK) = Koefisien keragaman; ( √ MSE) = root MSE; (MST) Minggu Setelah Tanam; (P< 0.05) = * nyata; (P< 0.01) = **sangat nyata; (1) = hasil transformasi X^0.5; (2) = pencilan dihilangkan; (3) = hasil transformasi X-0.5; (4) = hasil transformasi X-0.4; (5) = hasil transformasi log natural.
Cekaman kekeringan memberikan pengaruh yang sangat nyata terhadap beberapa peubah-peubah yang diamati antara lain : panjang daun, tinggi tanaman, lebar tajuk tanaman, bobot kering tajuk tanaman. Cekaman kekeringan memberikan pengaruh yang nyata terhadap lebar daun, jumlah anakan dan kadar air relatif daun.
juga memiliki pengaruh. Pengaruh interaksi antara cekaman kekeringan dan galur padi hanya nyata pada peubah tinggi tajuk tanaman 6 MST (Lampiran 6).
Pengaruh Cekaman Kekeringan terhadap Peubah-peubah yang Diamati a. Peubah morfologi
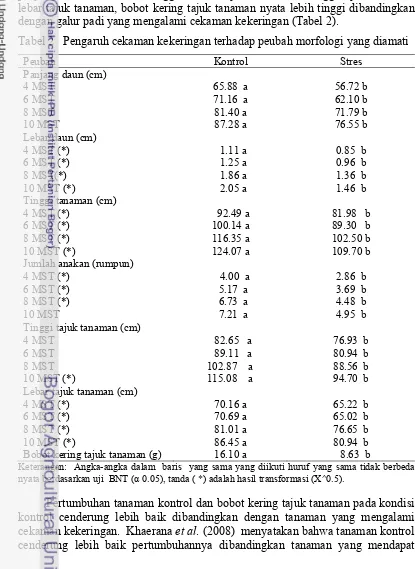
Perlakuan cekaman kekeringan nyata menghambat pertumbuhan dan bobot kering tajuk tanaman. Pada kondisi kontrol, peubah-peubah yang diamati yaitu panjang daun, lebar daun, tinggi tanaman, jumlah anakan, tinggi tajuk tanaman, lebar tajuk tanaman, bobot kering tajuk tanaman nyata lebih tinggi dibandingkan dengan galur padi yang mengalami cekaman kekeringan (Tabel 2).
Tabel 2 Pengaruh cekaman kekeringan terhadap peubah morfologi yang diamati
Peubah Kontrol Stres
Panjang daun (cm)
4 MST 65.88 a 56.72 b
6 MST 71.16 a 62.10 b
8 MST 81.40 a 71.79 b
10 MST 87.28 a 76.55 b
Lebar daun (cm)
4 MST (*) 1.11 a 0.85 b
6 MST (*) 1.25 a 0.96 b
8 MST(*) 1.86 a 1.36 b
10 MST (*) 2.05 a 1.46 b
Tinggi tanaman (cm)
4 MST (*) 92.49 a 81.98 b
6 MST (*) 100.14 a 89.30 b
8 MST (*) 116.35 a 102.50 b
10 MST (*) 124.07 a 109.70 b
Jumlah anakan (rumpun)
4 MST (*) 4.00 a 2.86 b
6 MST (*) 5.17 a 3.69 b
8 MST (*) 6.73 a 4.48 b
10 MST 7.21 a 4.95 b
Tinggi tajuk tanaman (cm)
4 MST 82.65 a 76.93 b
6 MST 89.11 a 80.94 b
8 MST 102.87 a 88.56 b 10 MST (*) 115.08 a 94.70 b Lebar tajuk tanaman (cm)
4 MST (*) 70.16 a 65.22 b
6 MST (*) 70.69 a 65.02 b
8 MST (*) 81.01 a 76.65 b
10 MST (*) 86.45 a 80.94 b
Bobot kering tajuk tanaman (g) 16.10 a 8.63 b
Keterangan: Angka-angka dalam baris yang sama yang diikuti huruf yang sama tidak berbeda nyata berdasarkan uji BNT (α 0.05), tanda ( *) adalah hasil transformasi (X^0.5).
cekaman kekeringan. Arifai (2009) juga menyatakan bahwa perlakuan cekaman kekeringan yang diberikan selama 12 hari setelah perlakuan (HSP) menghambat pertumbuhan tinggi tanaman dan luas daun pada tanaman padi, pada kelapa sawit (Dedywiryanto 2006), hal ini disebabkan ketersediaan air dalam media tumbuh yang rendah dan juga peranan air yang penting bagi pertumbuhan tanaman untuk pembelahan dan menjaga turgor sel agar tidak rusak. Menurut Gardner et al. (1991) dan Parker (2004), menjelaskan fungsi air bagi tanaman yaitu sebagai pelarut, media untuk transport, memberikan turgor pada sel, dan bahan baku untuk fotosintesis. Decoteau (2005) menyatakan bahwa kekeringan dapat menurunkan turgor sel sehingga mengakibatkan penurunan pada ukuran sel, menghambat pembelahan sel dan fotosintesis.
Secara umum dari penelitian ini, perlakuan cekaman kekeringan menyebabkan terhambatnya laju pertumbuhan tanaman. Hambatan pertumbuhan disebabkan oleh berkurangnya tekanan turgor sel akibat menurunnya potensial air sehingga proses pembesaran dan pemanjangan sel akan terhambat (Levitt 1980; Tezara et al. 2002; Hamim 2003). Hal ini berkaitan dengan perlakuan cekaman kekeringan yang diberikan, yang menyebabkan keterbatasan air sebagai pelarut unsur hara, sehingga kemampuan akar untuk menyerap unsur hara tersebut menjadi turun. Penurunan penyerapan hara dan air oleh akar, mengakibatkan suplai zat-zat yang dibutuhkan dalam pertumbuhan tanaman tidak terpenuhi, sehingga pertumbuhan tanaman menjadi terhambat (Sharma dan Flotcher 2002).
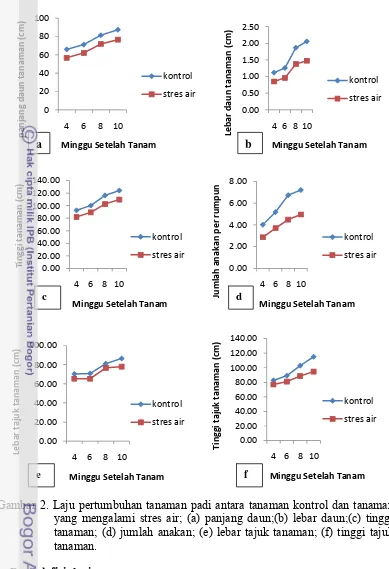
Laju pertumbuhan tanaman kontrol pada 8 MST sampai 10 MST lebih cepat jika dibandingkan tanaman yang mengalami stres air (Gambar 2). Tanaman kontrol, peningkatan laju pertumbuhan pada lebar daun, jumlah anakan, dan tinggi tajuk tanaman meningkat secara tajam pada 8 MST dan 10 MST. Tanaman yang mengalami stres air laju pertumbuhan tanamannya lebih terhambat, dibandingkan tanaman kontrol, hal ini disebabkan oleh peranan air yang penting bagi pertumbuhan dan perkembangan tanaman, dimana pada tahap pertumbuhan vegetatif, air digunakan oleh tanaman untuk melangsungkan proses pembelahan dan pembesaran sel.
Gardner et al. (1991) menjelaskan fungsi air bagi tanaman yaitu : (1) sebagai senyawa utama pembentuk protoplasma, (2) sebagai senyawa pelarut bagi masuknya mineral-mineral dari larutan tanah ke tanaman dan sebagai pelarut mineral nutrisi yang akan diangkut dari satu bagian sel ke bagian sel lain, (3) menjaga turgiditas sel dan berperan sebagai tenaga mekanik dalam pembesaran sel, (4) berperan dalam perpanjangan sel.
Gambar 2. Laju pertumbuhan tanaman padi antara tanaman kontrol dan tanaman yang mengalami stres air; (a) panjang daun;(b) lebar daun;(c) tinggi tanaman; (d) jumlah anakan; (e) lebar tajuk tanaman; (f) tinggi tajuk tanaman.
b. Peubah fisiologi
Hasil penelitian menunjukkan bahwa, tanaman kontrol memiliki nilai kadar air relatif (KAR) daun nyata lebih tinggi daripada tanaman stres yaitu 70.25 % (kontrol), 52.53 % (stres), kandungan prolin daun pada tanaman stres (0.055 µg prolin) nyata lebih tinggi daripada tanaman kontrol (0.021 µg prolin), namun untuk kerapatan stomata, klorofil b dan klorofil total respon tanaman kontrol sama dengan tanaman yang mengalami cekaman kekeringan (Tabel 3).
0
Minggu Setelah Tanam
kontrol
Minggu Setelah Tanam
kontrol
Minggu Setelah Tanam
kontrol
Minggu Setelah Tanam
kontrol
Minggu Setelah Tanam
kontrol
Minggu Setelah Tanam
Cekaman kekeringan salah satu faktor lingkungan terbesar yang menghambat pertumbuhan dan perkembangan tanaman, dengan menurunnya potensial air tanah maka pada tanaman akan terjadi berbagai perubahan proses fisiologi. Proses fisiologi yang menunjukkan toleransi tanaman terhadap cekaman kekeringan antara lain, mengurangi dehidrasi sel dengan merendahkan konduktansi stomata terhadap penguapan air, dan tekanan osmotik (Sinaga 2008). Tabel 3 Pengaruh Cekaman kekeringan terhadap peubah fisiologi yang diamati
Peubah Kontrol Stres KAR (%) daun 70.25 a 52.53 b Prolin ( µg prolin/g bobot segar)(1) 0.021 a 0.055 b Kerapatan stomata per mm2(2) 442.18 a 467.94 a Klorofil a (mg/g)(3) 0.328 a 0.247 b Klorofil b (mg/g) 0.110 a 0.107 a Klorofil total (mg/g)(4) 0.366 a 0.353 a
Keterangan: Angka-angka dalam baris yang sama yang diikuti huruf yang sama tidak berbeda nyata berdasarkan uji BNT (α 0.05), angka dalam tanda kurung adalah hasil transformasi (1) = pencilan dihilangkan; (2) = hasil transformasi X-0.5; (3) = hasil transformasi X-0.4; (4) = hasil transformasi log natural
Hasil analisis kadar air relatif (KAR) daun menunjukkan bahwa, nilai KAR pada tanaman kontrol nyata lebih tinggi daripada tanaman stres. Rendahnya nilai KAR pada tanaman stres menunjukkan bahwa tanaman menurunkan turgor serendah mungkin agar dapat bertahan, sehingga sel menjadi berukuran lebih kecil, luasan daun menjadi lebih sempit, jumlah anakan menjadi lebih sedikit, dan tinggi tanaman menjadi lebih pendek daripada tanaman kontrol (Gambar 2).
KAR (Kadar air relatif) daun merupakan peubah ketahanan tanaman menghadapi cekaman kekeringan (Quilambo 2004; Hamim 2007), karena dapat menggambarkan status air dan tekanan turgor dari sel-sel daun tanaman tersebut, khususnya pada saat tanaman mengalami penurunan potensial air. cekaman kekeringan menyebabkan turunnya tekanan turgor, sehingga stomata menutup. Penurunan KAR daun akan menurunkan konduktansi stomata daun dan dengan perlahan akan menurunkan konsentrasi CO2 didalam daun, sehingga dapat
menurunkan laju fotosintesis (Hamim 2007). Ashri (2006) menyatakan bahwa rendahnya nilai KAR tanaman yang tercekam kekeringan menunjukkan bahwa, tanaman menurunkan tekanan turgor serendah mungkin agar tetap bertahan, sehingga sel menjadi berukuran lebih kecil dari tanaman kontrol. Hanya tanaman yang memiliki kemampuan untuk melakukan pengaturan potensial osmotik biasanya dapat mempertahankan nilai KAR tetap tinggi.
2001; Ashri 2006; Riduan 2007). Menurut Claussen (2005) prolin merupakan indikator tanaman yang mengalami stres air.
Tingkat Toleransi
Berdasarkan definisi toleransi, tanaman yang toleran yaitu tanaman yang mampu/bertahan hidup pada kondisi tercekam kekeringan dan mampu menghasilkan produksi yang lebih baik daripada tanaman peka atau terjadi sedikit penurunan hasil. Pada penelitian ini, komponen produksi yang didapat yaitu bobot kering tajuk (BKT) tanaman sehingga toleransi tanaman dihitung berdasarkan persentase penurunan BKT tanaman kontrol dengan bobot kering tajuk tanaman stres (Chozin 2006).
Persentase penurunan bobot kering tanaman = – BKTstres % Persentase penurunan BKT tanaman digunakan untuk pengelompokkan galur berdasarkan toleransinya terhadap kekeringan dengan kategori sebagai berikut : sangat toleran = y < X - 1.5 SD; toleran = X -1.5 SD < y< X - 0.5 SD; sedang = X - 0.5 SD < y< X + 0.5 SD; peka = X + 0.5 SD < y< X + 1.5 SD; sangat peka = y > X + 1.5 SD, dimana y adalah persentase penurunan bobot kering, X adalah nilai tengah dan SD adalah standar deviasi (Sulistyono 1998).
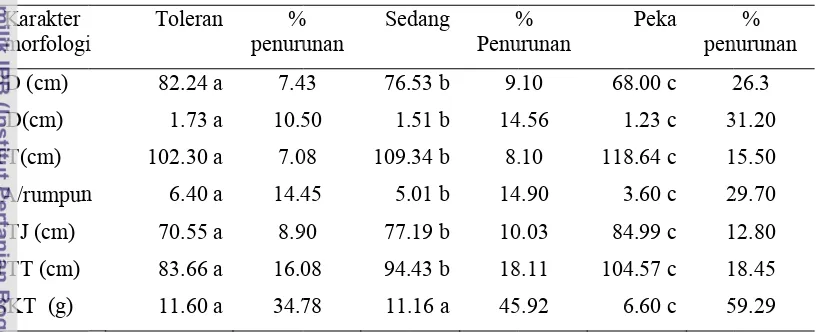
Berdasarkan perhitungan di atas, dari 100 galur padi yang diuji didapatkan hasil pengelompokkan toleransi tanaman yaitu sangat toleran = persentase penurunan BKT < 21.84 (ada 2 galur); Toleran = persentase penurunan BKT dari 21.84 – 37.78 (ada 16 galur); Sedang = persentase penurunan BKT dari 37.78 – 53.72 (ada 66 galur); Peka = persentase penurunan BKT dari 53.72 – 69.66 (ada 16 galur) (Tabel 4). Peringkat tingkat toleransi terhadap kekeringan dari 100 galur padi disajikan pada Lampiran 7.
Tabel 4 Pengelompokan galur padi berdasarkan toleransinya terhadap kekeringan Toleransi
terhadap kekeringan
Jumlah galur
Penurunan BKT (%) rata –rata kisaran
... (%) ...
Sangat toleran 2 19.85 19.85 – 19.86
Toleran 16 34.78 25.89 – 37.65
Sedang 66 45.92 38.02 – 53.70
Peka 16 59.29 54.02 – 67.94
Karakter Morfologi dari Galur Toleran dan Peka
cekaman kekeringan tanaman kacang tanah memiliki nilai KAR lebih tinggi dibandingkan varietas peka. Penelitian Bosch dan Penuelas (2004) pada tanaman strawberry juga menunjukkan bahwa penurunan nilai KAR sampai dengan 50 %, tanaman mengalami cekaman berat sehingga mengakibatkan terjadinya kerusakan pigmen fotosintesis. Penelitian Afa (2012) juga mengumukakan bahwa pada kondisi kadar air tanah 60 %, kadar air relatif daun nyata lebih tinggi pada galur toleran dibandingkan galur peka IR77298-14-1-2-B.
Tabel 6 Karakter fisiologi dari galur toleran dan peka
Pada penelitian ini, prolin terakumulasi lebih banyak pada galur yang mengalami cekaman kekeringan dan juga lebih banyak terakumulasi pada galur toleran, dibandingkan galur yang peka. Rendahnya kandungan prolin daun pada galur peka, disebabkan terbatasnya kemampuan galur peka kekeringan untuk mengakumulasi kandungan prolin daun pada potensial air tanah ≤ – 32 kPa. Menurut Yang dan Kao (1999) prolin yang terakumulasi didalam sitoplasma dapat digunakan tanaman sebagai energi bagi pemulihan proses fisiologi tanaman pasca cekaman kekeringan.
Penurunan potensial osmotik tanaman sebagai respon terhadap kekeringan merupakan mekanisme yang telah banyak diketahui ketika tanaman mengalami cekaman kekeringan (Levitt 1980), hal ini dapat dihasilkan melalui peningkatan konsentrasi solut, yang dapat mempertahankan turgor sel (Patakas et al. 2002). Kandungan klorofil berpengaruh terhadap besarnya hasil fotosintesis yang terjadi. Penurunan kandungan klorofil pada galur peka pada kondisi cekaman kekeringan disebabkan oleh kerusakan kloroplas akibat aktifitas Reactive Oxygen Species
(ROS) dan terjadi penurunan enzim Rubisco tanaman, sehingga menurunkan laju fotosintesis.
Karakter yang Berkaitan dengan Toleransi Kekeringan a. Peubah morfologi
Dari hasil penelitian, toleransi kekeringan dihitung berdasarkan peubah bobot kering tajuk tanaman. Karakter yang berkaitan dengan toleransi kekeringan diperoleh berdasarkan uji korelasi antara persentase penurunan bobot kering tajuk tanaman dengan peubah-peubah yang diamati (Tabel 7).
Tabel 7 dapat dilihat bahwa, persentase penurunan bobot kering tajuk berkorelasi negatif dengan lebar daun, jumlah anakan dan bobot kering tajuk tanaman, artinya semakin kecil nilai persentase penurunan bobot kering tanaman, tanaman akan semakin toleran. Semakin toleran tanaman, lebar daunnya akan
Karakter fisiologi Toleran Sedang Peka
Kadar air relatif daun (%) 11 MST 51.19 50.42 30.25 Prolin ( µg prolin/g bobot segar)
11 MST
0.093 0.042 0.009
Kerapatan stomata per mm2 8 MST 412.74 419.16 463.69
Klorofil a (mg/g) 8 MST 0.250 0.252 0.220
Klorofl b (mg/g) 8 MST 0.102 0.110 0.101
semakin lebar, jumlah anakannya akan semakin banyak, dan bobot kering tajuk tanaman akan semakin besar.
Setiap jenis tanaman memiliki mekanisme yang berbeda dalam menanggapi kondisi lingkungan yang tidak optimal. Telah banyak dipelajari berbagai respon morfologi dan fisiologi tanaman yang berkaitan dengan mekanisme ketahanan terhadap kekeringan. Cekaman kekeringan dapat menghambat pertumbuhan tanaman. Penghambatan pertumbuhan ini salah satunya dapat dilihat pada perluasan daun. Penurunan luas daun merupakan respon pertama tanaman terhadap kekeringan. Keterbatasan air akan menghambat perpanjangan sel yang secara perlahan akan menghambat pertumbuhan luas daun. Sempitnya luas daun mengakibatkan rendahnya transpirasi, sehingga menurunkan laju suplai air dari akar ke daun (Taiz and Zeiger 2002). Hasil penelitian menunjukkan bahwa cekaman air berpengaruh terhadap respon morfologi jarak pagar pada peubah tinggi tanaman dan luas daun (Courtois dan Lafitte 1999; Syafi 2008; Lapanjang
et al. 2008), jumlah anakan (Tubur 2011).
Secara umum mekanisme ketahanan terhadap cekaman kekeringan sudah banyak diketahui yaitu (1) drought escape, tanaman mampu menyelesaikan siklus hidup sebelum terjadi cekaman, (2) drought avoidance terdiri dari (2a). Toleran kekeringan pada potensial air jaringan tinggi misalnya perakaran dalam, stomata sedikit, adanya bulu daun, dan (2b). Toleran kekeringan pada potensial air jaringan rendah yaitu dengan cara mempertahankan turgor melalui akumulasi senyawa terlarut dalam sitoplasma (prolin barley 7-10 x kontrol, prolin kedelai 5-7 x kontrol), dan meningkatkan elastisitas jaringan. Karakter morfologi yang berkaitan dengan adaptasi terhadap stress antara lain ukuran tajuk seperti jumlah anakan sedikit, menunda pembungaan, pengurangan anakan produktif (Oosterom
et al. 2003), total bahan kering rendah (Pantuwan et al. 2002), daun lebih pendek
(Nguyen et al. 2009).
b. Peubah fisiologi
Karakter fisiologi yang berkorelasi dengan persentase penurunan berat kering tajuk tanaman (BKT) adalah KAR dan klorofil b (Tabel 8). Persentase penurunan bobot kering tajuk (BKT) tanaman berkorelasi negatif dengan KAR daun, artinya semakin kecil nilai persentase penurunan bobot kering tanaman, tanaman akan semakin toleran. Semakin toleran tanaman, nilai KAR daun semakin besar.