HASIL DAN PEMBAHASAN
Sebelum melakukan analisis QTL untuk studi pemetaan, telah dilakukan estimasi keberadaan gen mayor dengan analisis mixture-models. Dengan demikian dapat diperkiraan ada tidaknya segregasi sifat penting yang mempunyai pengaruh besar tersebut atau disebut gen mayor yang diwariskan dari tetua kepada keturunannya. Pemetaan QTL dengan analisis QTL, pada dasarnya juga melihat ada atau tidaknya segregasi sifat kuantitatif, namun estimasi pada analisis QTL ini akan lebih mendekati ketepatan karena telah melibatkan penciri molekuler sebagai
landmark yang mengarahkan terhadap keberadaan gen kandidat yang dicari.
Penapisan Penciri
Sesuai dengan anjuran Grosz & MacNeil (2001), dinyatakan bahwa guna memaksimalkan hasil dan mengefisienkan proses allele scoring dari hasil genotyping, perlu dilakukan screening atau penapisan penciri mikrosatelit terhadap panel mikrosatelit praseleksi. Penapisan dimaksudkan agar penciri mikrosatelit yang digunakan akan menghasilkan genotipe heterosigot pada turunan domba dari pustaka keluarga yang terbentuk pada genotyping progeny nantinya.
Lebih lanjut diterangkan oleh Grosz & MacNeil (2001), penapisan penciri (screening markers) mikrosatelit dilakukan terhadap tiga generasi, yaitu Bapak atau pejantan F1 (Sire), Kakek (Grandsire = GS) dan Nenek (Grandam = GD). Studi ini menggunakan empat pejantan F1, oleh karena dalam penapisan penciri mikrosatelit juga digunakan ke empat pejantan F1 dengan tetuanya (GS dan DD). Ke empat pejantan F1 tersebut dengan ID 1261 (3050/GS dan 6/GD), 1262 (25/GS dan 1/GD), 1263 (2153/GS dan 13/GD) dan 1273 (102/GS dan 12/GD).
Pada penapisan penciri tersebut dapat dihasilkan penciri mikrosatelit informatif heterosigot, informatif homosigot atau penciri yang tidak informatif. Namun, pada studi ini digunakan dua kriteria pertama untuk dijadikan sebagai penciri mikrosatelit yang informatif. Penciri informatif yang digunakan dalam studi genotyping progeny yaitu apabila anak mewarisi ke dua alel tetuanya. Namun, dapat terjadi diperoleh
mikrosatelit yang tidak informatif, karena tidak mewarisi kedua alel dari ke orang tuanya (Sire= Bapak ataupun Dam= Ibu).
Contoh hasil pengerjaan penapisan penciri mikrosatelit, ditampilkan seperti pada Gambar 1. Pada Gambar 1, ditunjukkan bahwa penciri BMS528 adalah penciri informatif dan heterosigot untuk F1 sires 1263 dan 1273, karena penciri tersebut mewarisi 2 alel, masing-masing dari GrandSire dan GrandDam. BMS528 juga informatif untuk F1 Sire 1261 tetapi homosigot (alel menumpuk tebal jadi satu). Sementara itu, penciri BMS528 tidak informatif terhadap F1 Sire 1262, karena tidak mewarisi alel dari kedua orang tuanya (GrandSire dan GrandDam). Hasil penapisan penciri mikrosatelit yang berjumlah 250, diperoleh 136 penciri mikrosatelit yang informative setelah diadakan seleksi.
Genotyping Progeny
Genotyping dilakukan pada seluruh populasi progeny domba silang balik dan
sebagian F2 serta tetuanya yang berjumlah seluruhnya 381 domba. Contoh hasil pengerjaan progeny ditampilkan pada Gambar 2.
Gambar 1. Hasil penapisan penciri BMS528. BMS528 adalah penciri informatif heterosigot untuk pejantan F1 1263 dan 1273, dan informatif homosigot untuk 1261. Namun BMS528 tidak informatif untuk pejantan F1 1262.
Notasi 1, 2 dan 3 menunjukkan susunan individu Kakek, Nenek, Pejantan F1 secara berturut-turut
Gambar2. Hasil genotyping keluarga 1261 dengan penciri BMS2213 Pada Gambar 2 ditunjukkan salah satu hasil genotyping pada acuan keluarga 1261 dengan penciri mikrosatelit BMS2213. Contoh keluarga acuan 1261 terdiri dari susunan Kakek (GS) 3050, Nenek (GD) 6, Anak F1 (Sire) 1261 masing-masing menempati sumur (well) nomor urut 1, 2 dan 3. Kemudian disusul progenynya sebanyak 45 dengan urutan dari no. 4 sampai dengan no. 48. Dengan demikian jumlah keseluruhan ada 48 individu domba untuk setiap pengerjaan genotyping pada gel PAGE (Polyacrylamide Gel Electrophoresis). Namun dalam satu gel tersebut dapat dimaksimalkan sampai 6 kali pengerjaan genotyping. Dengan demikian akan diperoleh 288 (6x48) genotyped individu dalam satu gel untuk setiap kali running
genotyping. Sementara itu, jumlah penciri mikrosatelit yang digunakan sebanyak 136
dan jumlah individu progeny 381, dengan demikian studi ini telah menghasilkan 51.816 individu genotipe. Memang belum ada standard yang menyebutkan banyaknya penciri genetik dan jumlah populasi yang digunakan dalam studi pemetaan. Namun Raadsma et al. (2002a; 2002b), menyarankan diperlukan populasi besar dan jumlah penciri genetik besar apabila belum diketahui lokasi gen yang dicari. Lebih banyak penciri genetik dan populasi akan mempersempit jarak rentang estimasi lokasi QTL atau gen. Lynch & Walsh (1998) menyarankan setidaknya antara
50 sampai 200 penciri genetik atau lebih dapat digunakan dalam analisis QTL untuk mempersempit jarak rentang daerah estimasi lokasi QTL.
Scoring allele, setelah diperoleh sebaran alel pada genotyping, perlu dilakukan
evaluasi atau menilai sebaran alel. Disarankan oleh Crawford et al. (1995), penilaian alel pada genotyping setidaknya dilakukan oleh dua (2) peneliti. Namun dalam studi ini selain dilakukan penilaian oleh 2 peneliti juga dibantu dengan perangkat lunak (software: Gene ImagIR) yang menjadi satu paket dengan mesin LI-COR DNA Analyzer yang digunakan untuk genotyping. Terdapat kesepakatan pada penilaian penyebaran alel, seperti disebutkan sebelumnya bahwa alel dari Pejantan (Sire) dinilai dengan 1, alel dari Ibu (Dam) dinilai 2, sedangkan tidak termasuk keduanya diberikan nilai 6 dan tidak menunjukan alel apapun di notasikan dengan nol (0). Penilaian alel tersebut diperlukan dalam penyusunan input data terutama untuk Genotype file guna keperluan analisis QTL dengan program QTL Express, contoh penyusunan input data untuk genotype file dapat dilihat pada Lampiran 4.
Pada Lampiran 4 (genotype file), disusun genotipe alel dari semua individu yang berasal dari ke empat keluarga acuan yang di genotipe dengan 136 penciri mikrosatelit.
Identifikasi Gen Mayor – Analisis Segregasi
Pada awal pembahasan telah disinggung bahwa ada baiknya untuk melakukan analisis segregasi sebelum melakukan pemetaan QTL. Pada analisis segregasi ini tidak melibatkan penciri genetik, namun hanya menggunakan data fenotipik. Seperti diterangkan oleh Crawford et al. (2000) bahwa analisis segregasi dirancang untuk mendeteksi segregasi genetik pada sifat bernilai ekonomi tinggi tanpa melibatkan keberadaan informasi penciri genotipe.
Hasil analisis segregasi dengan mixture-models (Lynch & Walsh 1998) dan setelah diuji dengan Likelihood Ratio Test (LRT) untuk parameter yang diamati dapat dilihat pada Tabel 7. Keberadaan gen mayor terdeteksi untuk sifat berat lahir (BL) dari populasi silang balik yang berasal dari keluarga acuan 1263 (3.47±1.09 kg). Segregasi gen mayor juga nampak pada berat umur 90 (BB90) dari populasi domba
Gambar 3. Kurva bi-modal (terjadi segregasi) terindikasi pada populasi domba silang balik untuk sifat pra-sapih: a) indikasi keberadaan gen mayor pada populasi keluarga acuan 1263 untuk berat lahir (BL); segregasi gen mayor untuk BB90 pada keluarga acuan 1261 (b), pada keluarga
acuan 1262 (c), pada keluarga acuan 1273 (d); segregasi gen mayor untuk pertambahan bobot badan B1-90 pada keluarga acuan 1261 (e)
silang balik yang berasal dari keluarga acuan 1261 (13.86±3.46 kg), dari keluarga acuan 1262 (14.66±4.02 kg) dandari keluarga acuan 1273 (13.38±3.42kg). Selain itu gen mayor juga teridentifikasi pada pertambahan bobot badan pbb1-90 pada populasi
keluarga acuan 1261 (112.87±35.80 gram) dan 1263 (110.85±42.98 gram). Ke enam indikasi keberadaan segregasi gen (ditunjukkan dengan kurva bi-modal) untuk sifat pertumbuhan pra-sapih yang diamati dari keluarga acuan yang berbeda, dapat dilihat pada Gambar 3.
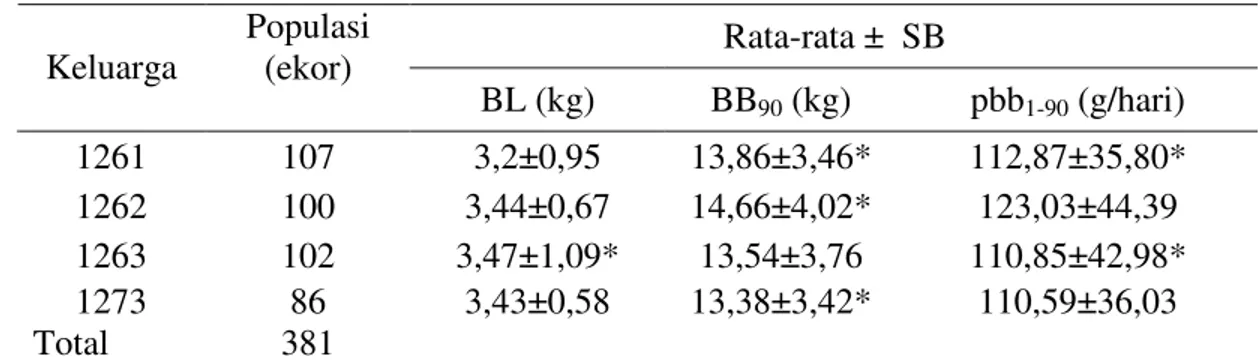
Tabel 7. Rataan sifat pertumbuhan pra-sapih (BL dan BB90) pada populasi
domba silang balik dari empat keluarga acuan (saudara tiri) Rata-rata ± SB
Keluarga Populasi (ekor)
BL (kg) BB90 (kg) pbb1-90 (g/hari) 1261 107 3,2±0,95 13,86±3,46* 112,87±35,80* 1262 100 3,44±0,67 14,66±4,02* 123,03±44,39 1263 102 3,47±1,09* 13,54±3,76 110,85±42,98* 1273 86 3,43±0,58 13,38±3,42* 110,59±36,03 Total 381
* Terjadi Segregasi (lihat Gambar 3) SB= simpangan baku
Dinyatakan oleh Lynch & Wals (1998), bahwa kurva bi-modal adalah sebagai campuran distribusi nilai fenotipe normal. Namun bilamana tidak terjadi campuran distribusi normal yang menyusun kurva bi-modal, hal ini mungkin gen mayor tidak berada pada keseimbangan Hardy – Weinberg yang mendasari penyebaran fenotipe untuk setiap genotipe marker-locus.
Analisis segregasi telah menunjukkan indikasi keberadaan gen mayor untuk beberapa sifat pertumbuhan pra-sapih yang diamati pada populasi silang balik. Analisis segregasi tersebut dimaksudkan untuk mengetahui penyebaran fenotip hewan dan diuji untuk ketepatan terhadap konsistensi penyebaran pada populasi dimana gen mayor bersegregasi (ACIAR report 2001). Kelemahan analisis segregasi dengan hanya menggunakan data fenotipik yaitu menunjukkan indikasi keberadaan gen mayor. Kemajuan teknologi penciri molekuler, segregasi gen mayor dapat dikonfirmasi melalui analisis keterpautan dengan menggunakan polymorphic markers
yang terletak mendekati gen mayor (Van der Werf 2000d). Hal ini hanya dapat dibuktikan melalui analisis QTL yang melibatkan sejumlah besar jumlah penciri genetik mikrosatelit. Keberadaan lokasi (QTL) untuk sifat penting yang ditandai dengan penciri genetik yang terletak dekat dengan gen atau terdapat asosiasi antara penciri genetik dengan gen mayor (Raadsma et al. 2002b; Dominik 2005b).
Identifikasi QTL
Keberadaan QTL - Significance Threshold
Uji statistik dimaksudkan untuk memastikan secara statistik dapat diambil kesimpulan yang wewakili. Pada analisis QTL ini, pengujian dilakukan dengan uji permutasi (de Koning et al. 2001) yang dimaksudkan untuk memperoleh nilai ambang empiris (empiric threshold) atas pengaruh QTL terhadap sifat yang diteliti. Pada studi analisis QTL ini, nilai ambang nyata (significance threshold) diuji pada taraf permutate-chromosome wide (Seaton et al. 2002) dan kemudian untuk melihat apakah pengaruh nilai ambang nyata QTL tersebut masih dapat tetap dipertahankan kuat, maka dilakukan uji permutate-experiment wide (Seaton et al. 2002).
Hasil dari ke dua uji tersebut terhadap sifat pertumbuhan untuk berat lahir (BL), berat umur 90 hari (BB90), umur 180 hari (BB180), umur 270 hari (BB270) dan umur
360 hari (BB360), masing-masing disajikan pada Tabel 8, 9, 10, 11 dan Tabel 12. Dari
ke lima Tabel tersebut, dapat dilihat pengaruh nyata kandidat QTL untuk ke lima sifat berat badan yang diamati setelah diuji dengan permutate-chromosome wide, terdeteksi sebanyak 15 lokasi QTL pada kromosom domba. Ke lima belas kromosom yaitu kromosom 5 untuk BL, kromosom 2, 7, 18 untuk sifat BB90, kromosom 18 dan
23 untuk BB180, kromosom 6, 7, 8, 18 dan 23 untuk BB270, kromosom 7, 8, 18 dan 23
untuk BB360.
Sifat pertumbuhan pada berat lahir (BL) menunjukkan pengaruh QTL yang nyata untuk berat lahir (BL) anak domba pada kromosom 5 (p<0.01). Penemuan ini berbeda dari analisis gen mayor sebelumnya yang tidak menunjukkan adanya pewarisan segregasi sifat berat lahir yang ditemukan pada populasi kecuali hanya terjadi pada keluarga acuan 1263 (Margawati & Subandriyo 2004; Margawati et al.
2004). Hal ini dapat terjadi seperti diketahui bahwa identifikasi gen mayor dengan metode mixture-models untuk analisis segregasi dinyatakan kurang kuat atau less
powerfull (Crawford et al. 2000). Dengan demikian setelah dilakukan analisis QTL
dengan melibatkan sejumlah besar penciri genetik mikrosatelit (136), pewarisan segregasi gen untuk sifat berat lahir (BL) nampak kuat pada kromosom 5.
Tabel 8. Significance threshold berat lahir (BL)
Chromosome-Wide Experiment-Wide
No
Krom Nilai-F 5% 1% 5% 1%
5 4.42 3.2373 4.3577** 5.4262 6.4222
Chromosome wide: uji permutasi untuk menetapkan signifikansi pada setiap kromosom
Experiment wide: uji permutasi untuk menetapkan signifikasni pada setiap kromosom yang
dibandingkan dengan seluruh 26 kromosom
* Berbeda nyata pada 5% (p<0.05), ** Berbeda nyata pada 1% (p<0.01), berlaku baik pada uji permutasi chromosome wide maupun experiment wide
Tabel 9. Significance threshold berat badan 90 (BB90)
Chromosome-Wide Experiment-Wide No Krom Nilai-F 5% 1% 5% 1% 2 4.43 4.0092* 5.3145 5.3728 6.5258 7 4.33 3.2942 4.0787** 5.3728 6.5258 18 4.13 3.2773* 4.7927 5.3728 6.5258
* Berbeda nyata pada 5% (p<0.05), ** Berbeda nyata pada 1% (p<0.01), berlaku baik pada uji permutasi chromosome wide maupun experiment wide
Seperti diketahui bahwa persaingan pertumbuhan anak domba baru kelihatan setelah lepas sapih (Subandriyo et al. 1995). Nampaknya hal ini dibuktikan dengan diperolehnya lebih banyak lokasi QTL untuk sifat pertumbuhan mulai berat umur 90 (BB90) terdapat 3 lokasi QTL pada kromosom berbeda (Tabel 9), BB180 terdapat 2
lokasi QTL pada kromosom berbeda (Tabel 10), BB270 terdapat 5 lokasi QTL (Tabel
Tabel 10. Significance threshold berat badan 180 (BB180)
Chromosome-Wide Experiment-Wide
No Krom Nilai-F 5% 1% 5% 1%
18 4.3 3.1801 4.0585** 5.1151 6.3466
23 3.21 2.9808* 3.6954 5.1151 6.3466
* Berbeda nyata pada 5% (p<0.05), ** Berbeda nyata pada 1% (p<0.01), berlaku baik pada uji permutasi chromosome wide maupun experiment wide
Tabel 11. Significance threshold berat badan 270 (BB270)
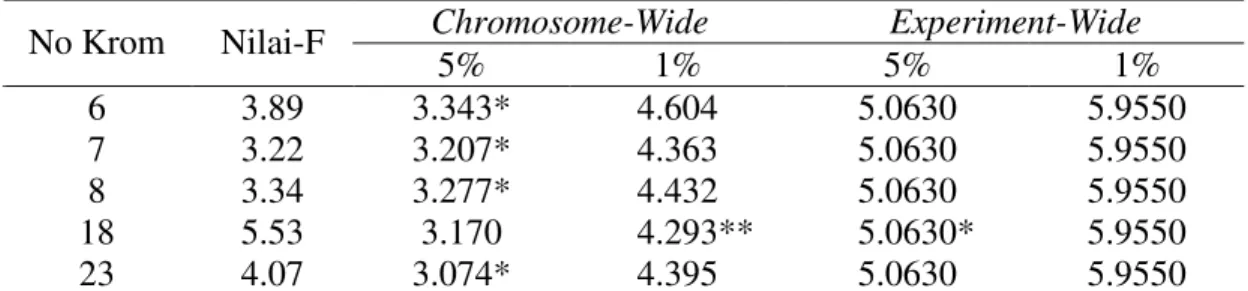
Chromosome-Wide Experiment-Wide No Krom Nilai-F 5% 1% 5% 1% 6 3.89 3.343* 4.604 5.0630 5.9550 7 3.22 3.207* 4.363 5.0630 5.9550 8 3.34 3.277* 4.432 5.0630 5.9550 18 5.53 3.170 4.293** 5.0630* 5.9550 23 4.07 3.074* 4.395 5.0630 5.9550
* Berbeda nyata pada 5% (p<0.05), ** Berbeda nyata pada 1% (p<0.01), berlaku baik pada uji permutasi chromosome wide maupun experiment wide
Tabel 12. Significance threshold berat badan 360 (BB360)
Chromosome-Wide Experiment-Wide No Krom Nilai-F 5% 1% 5% 1% 7 3.77 3.2435* 4.2703 5.2299 6.0719 8 3.87 3.2086* 4.4071 5.2299 6.0719 18 6.27 3.2908 4.3544** 5.2299 6.0719** 23 3.88 3.1133* 4.0084 5.2299 6.0719
* Berbeda nyata pada 5% (p<0.05), ** Berbeda nyata pada 1% (p<0.01), berlaku baik pada uji permutasi chromosome wide maupun experiment wide
Tabel 12 di atas menunjukkan bahwa pengaruh QTL untuk sifat pertumbuhan pada berat hidup 360 hari (BB360) menunjukkan tetap bertahan pada signifikansi
nyata (p<0,01) setelah uji permutasi eksperimen (permutate-experiment wide). Gambaran ini menunjukkan betapa kemungkinan keberadaan QTL sifat pertumbuhan domba pada kromosom 18. Sementara itu pada uji yang sama, pengaruh QTL untuk BB270 hanya menunjukkan signifikansi tingkat 5% (p<0,05), Tabel 11 dan untuk
BB180 tidak menunjukkan tingkat signifikansi (Tabel 10). Meskipun demikian dapat dinyatakan bahwa lokasi QTL sifat pertumbuhan pasca sapih (BB• 180 hari) terletak pada kromosom 18. Hasil perolehan pada kromosom 18 domba ini ternyata
memperkuat pendapat sebelumnya untuk sifat pertumbuhan dan karkas pada populasi komersial domba Suffolk dan Texel Walling et al. (2004).
Kurva significance threshold untuk beberapa sifat berat badan (BL, BB90,
BB180, BB270, BB360) yang menunjukkan pengaruh nyata QTL pada taraf 1% (p<0.01)
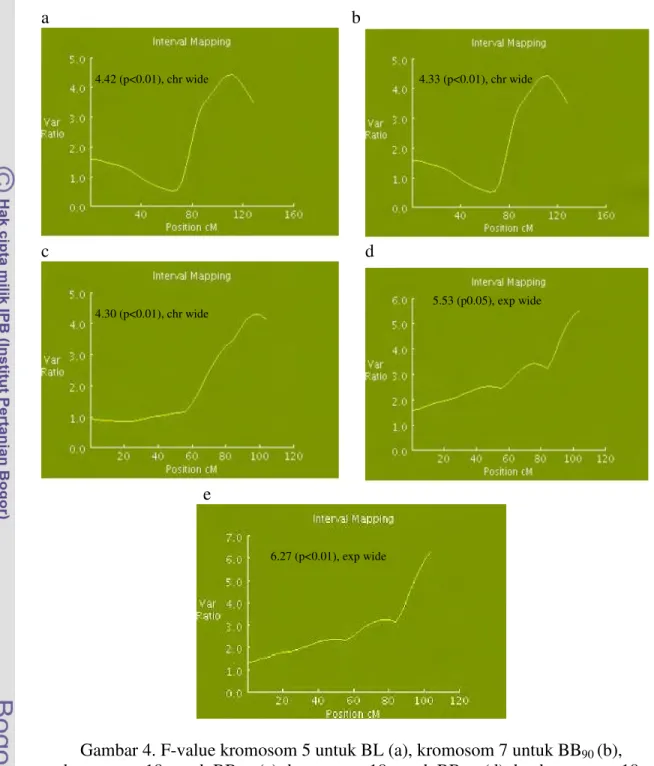
dan 5% (p<0.05), disajikan pada Gambar 4. Gambar 4a, menunjukkan tingkat signifikansi keberadaan QTL untuk BL adalah 1% (p<0.01) setelah uji permutasi
chromosome wide. Gambar 4b dan 4c, masing-masing menunjukkan tingkat
keberadaan QTL pada 1% (p<0.01) setelah uji permutasi chromosome wide, masing-masing BB90 dan BB180. Sementara Gambar 4d dan 4e, masing-masing menunjukan
keberadaan QTL tetap dipertahankan pada 5% (p<0.05) dan 1% (p<0.01) setelah uji permutasi experiment wide untuk BB270 dan BB360.
Dari keseluruhan hasil uji permutasi tersebut diatas, nampak secara jelas terdapat pola pengaruh QTL secara nyata terhadap pertumbuhan anak domba terdeteksi pada kromosom 18 mulai dari BB90 sampai BB360 (Tabel 9, 10, 11, 12).
Hasil uji permutasi (untuk melihat significance threshold) pada semua karakter berat hidup disajikan pada Lampiran 6.
a b
c d
e
Gambar 4. F-value kromosom 5 untuk BL (a), kromosom 7 untuk BB90 (b),
kromosom 18 untuk BB180 (c); kromosom 18 untuk BB270 (d) dan kromosom 18
untuk BB360 (e). chr wide = chromosome wide; exp wide = experiment wide
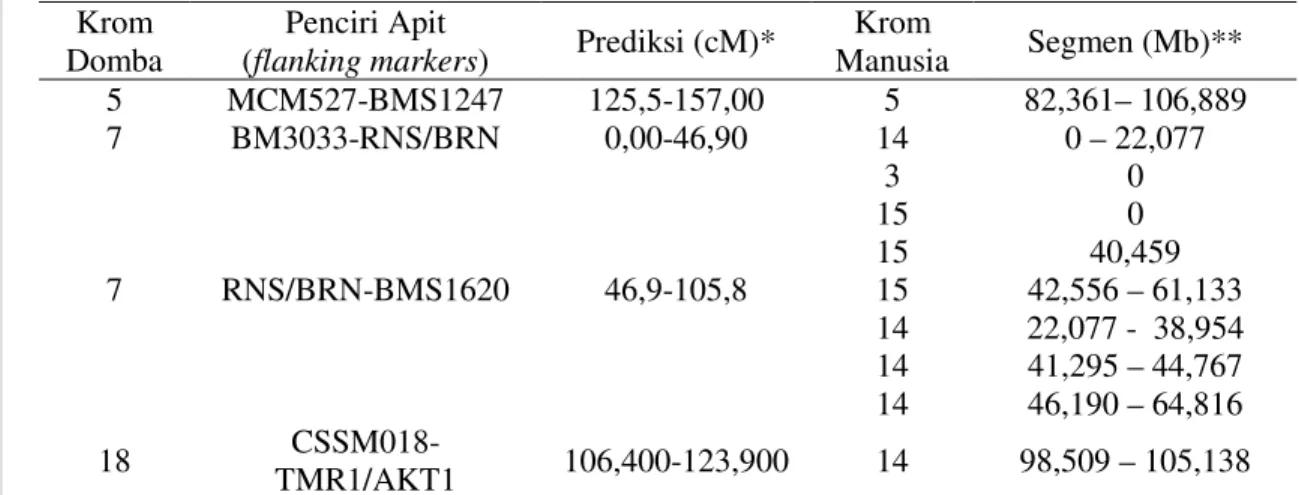
Lokasi QTL dan Penciri Apit (Flanking Markers)
Pengaruh nyata QTL sifat pertumbuhan pada masing-masing berat badan yang diamati telah dipetakan dengan menunjukkan lokasi pada kromosom nomor tertentu,
4.42 (p<0.01), chr wide
6.27 (p<0.01), exp wide
4.30 (p<0.01), chr wide 5.53 (p0.05), exp wide 4.33 (p<0.01), chr wide
jarak (interval), penciri DNA yang mengapit lokasi QTL, homologi dengan kromosom manusia serta posisi QTL pada bagian (segment) kromosom manusia (Mb), lihat paparan pada Tabel 13.
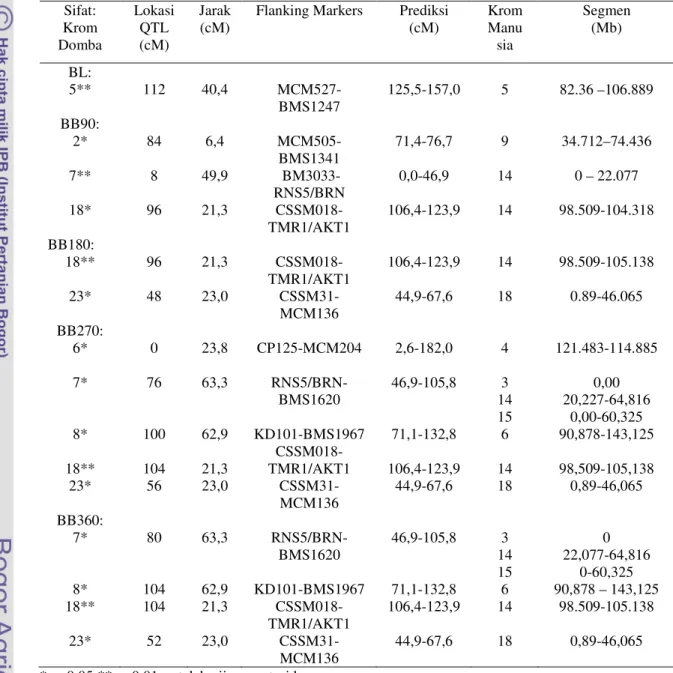
Tabel 13. Lokasi QTL dan penciri apit (flanking Markers) sifat pertumbuhan Sifat: Krom Domba Lokasi QTL (cM) Jarak
(cM) Flanking Markers Prediksi (cM) Krom Manu sia Segmen (Mb) BL: 5** 112 40,4 MCM527-BMS1247 125,5-157,0 5 82.36 –106.889 BB90: 2* 7** 18* 84 8 96 6,4 49,9 21,3 MCM505-BMS1341 BM3033-RNS5/BRN CSSM018-TMR1/AKT1 71,4-76,7 0,0-46,9 106,4-123,9 9 14 14 34.712–74.436 0 – 22.077 98.509-104.318 BB180: 18** 23* 96 48 21,3 23,0 CSSM018-TMR1/AKT1 CSSM31-MCM136 106,4-123,9 44,9-67,6 14 18 98.509-105.138 0.89-46.065 BB270: 6* 7* 8* 18** 23* 0 76 100 104 56 23,8 63,3 62,9 21,3 23,0 CP125-MCM204 RNS5/BRN-BMS1620 KD101-BMS1967 CSSM018-TMR1/AKT1 CSSM31-MCM136 2,6-182,0 46,9-105,8 71,1-132,8 106,4-123,9 44,9-67,6 4 3 14 15 6 14 18 121.483-114.885 0,00 20,227-64,816 0,00-60,325 90,878-143,125 98,509-105,138 0,89-46,065 BB360: 7* 8* 18** 23* 80 104 104 52 63,3 62,9 21,3 23,0 RNS5/BRN-BMS1620 KD101-BMS1967 CSSM018-TMR1/AKT1 CSSM31-MCM136 46,9-105,8 71,1-132,8 106,4-123,9 44,9-67,6 3 14 15 6 14 18 0 22,077-64,816 0-60,325 90,878 – 143,125 98.509-105.138 0,89-46,065 * p<0.05 ** p<0.01, setelah uji permutasi kromosom
Setelah uji permutasi eksperimen, membuktikan tetap nyata untuk BB270 (p<0.05) dan BB360 (p<0.01)
Prediksi (cM)= prediksi lokasi gen pada kromosom domba
Segmen (Mb), lokasi gen pada rentangan kromosom manusia, setelah dilakukan analisis melalui ASGMWS (2004)
Penciri apit lokasi QTL dimaksudkan untuk mendeterminasi gen kandidat yang terletak diantara ke dua titik penciri DNA pengapit tersebut dan ditelusuri melalui situs ncbi: http://www.ncbi.nih.gov. Dari ke lima sifat kuantitatif berat badan (BL, BB90, BB180, BB270 dan BB360) diperoleh bukti nyata pengaruh QTL untuk sifat
pertumbuhan pada 7 kromosom yang berbeda yaitu kromosom 2, 5, 6, 7, 8, 18 dan 23, lihat Tabel 8, 9, 10, 11 dan 12 dengan tingkat signifikansi antara 5% sampai 1%.
Dukungan terhadap pendugaan keberadaan QTL yang mempengaruhi perototan pada kromosom 2 diperoleh dari peta perbandingan genom sapi (Grosz & MacNeil 2001). Pada studi ini, teridentifikasi pengaruh QTL untuk pertumbuhan domba pada berat umur 90 hari (BB90) terletak pada kromosom 2 (p<0.05) setelah uji permutasi
eksperimen. Penemuan ini kemungkinan pada genotyping digunakan 3 dari 12 penciri mikrosatelit berasal dari sapi (Bovine).
Diperkirakan lokasi QTL untuk BB90 yang terletak pada kromosom 2 berada
disekitar penciri BMS1341 terletak pada 74,8cM atau sekitar penciri BM81124 terletak pada 146,3cM atau sekitar BMS1126 terletak pada 215,3cM. Lokasi QTL untuk BB90 pada kromosom 2 domba ini terletak menyebar di tiga daerah dari kromosom 2 dengan 3 penciri terpaut pada estimasi lokasi gen (linked to genes), yaitu disekitar penciri BMS1341, BM81124 dan disekitar BMS1126 (Gambar 5).
Gambar 5. Lokasi penciri mikrosatelit digunakan dalam analisis alel (genotyping) pada kromosom 2, QTL teridentifikasi diperkirakan berlokasi disekitar
Hasil penelitian ini mendukung penelitian Walling et al. (2002) yang melaporkan penemuan lokasi QTL pada kromosom 2 dan 18 domba untuk sifat berat badan (kg) dan ketebalam lemak (mm) pada domba Texel. Penelitian tersebut tidak melaporkan penciri genetik yang menunjukkan lokasi QTL pada ke dua kromosom, namun menyebutkan lokasinya. Dilaporkan bahwa lokasi QTL yang mempengaruhi pertumbuhan otot terletak sekitar 60cM, dimana pada penelitian domba ITT dan Merino ini menunjukkan penemuan lokasi QTL untuk sifat pertumbuhan terletak pada74,8cM dengan penciri BMS1341 (Gambar 5). Sementara lokasi lemak menurut laporan Walling et al (2002) terletak sekitar 170cM, lokasi tersebut juga terdeteksi pada penelitian ini yaitu disekitar penciri apit BM81124 dan BMS1126 yang berjarak antara 146,3 dan 215,3cM (Gambar 5). Terlihat pada penelitian ini, masih perlu menggunakan penciri mikrosatelit yang lebih rapat atau jarak penciri mikrosatelit yang lebih pendek (Gambar 5), untuk mempersempit identifikasi lokasi QTL berasosiasi dengan sifat pertumbuhan.
Hasil analisis keberadaan QTL sifat pertumbuhan pada kromosom 2 (p<0.05) dan 18 (p<0.01) pada penelitian ini juga mendukung studi sebelumnya pada lima keluarga kelompok domba Texel di United Kingdom (UK) (Walling et al. 2001). Dari hasil analisis mereka, dikonfirmasi pembuktian untuk pengaruh QTL tersebut pada ketebalan otot dan lemak yaitu terletak pada kromosom 2 dan 18 domba Texel di UK. Sedangkan pada studi lainnya telah mengindikasikan keberadaan gen mayor untuk peningkatan perototan pada domba Texel Belgia, Australia dan New Zealand (Marcq et al. 1998; Marshall et al. 1999; Broad et al. 2000). Domba Texel dari Belgia dan New Zealand telah menunjukkan gen myostatin atau locus yang dekat dengan gen myostatin yang berperan pada peningkatan perototan. Sementara itu pendapat Walling et al. (2001) menyatakan bahwa lokasi pada kromosom 2 dan 18 adalah mirip dengan gen Carwell. Studi Walling et al. (2001) menggunakan data pertumbuhan setelah 8 minggu sementara pada studi ini pengamatan dilakukan mulai dari berat lahir (BL) sampai dengan berat badan umur 360 (BB360). Perkembangan
perototan ini dinyatakan oleh Cockett et al. (2001) dimulai setelah anak domba berumur 3 minggu.
Sebanyak 5 penciri mikrosatelit yang digunakan untuk genotipe alel (genotyping) pada kromosom 18, penciri mikrosatelit dan lokasinyan pada kromosom 18 dipaparkan (Gambar 6). Hasil analisis menunjukkan bahwa QTL teridentifikasi pada kromosom 18 untuk beberapa karakter berat badan (Tabel 13) terletak diantara penciri apit CSSM018 dan TMR1 pada jarak antara 107,1 dan 124,8 cM (Gambar 6).
Gambar 6. Lokasi penciri mikrosatelit digunakan dalam analisis alel (genotyping) pada kromosom 18, QTL teridentifikasi diperkirakan berlokasi diantara penciri apit
berhuruf tebal (CSSM018 dan TMR1)
Pengaruh QTL terhadap pertumbuhan juga dibuktikan secara nyata (p<0.05) setelah uji permutasi eksperimen (permutate-experiment wide) pada kromosom 6 untuk berat badan umur 270 (BB270). Namun pada penelitian sebelumnya pengaruh
QTL pada kromosom 6 dilaporkan berasosiasi dengan beragam sifat. Seperti dilaporkan oleh Montgomery et al. (1994) bahwa gen Booroola dipetakan pada kromosom domba 6 (OOV6). Sebelumnya, Montgomery et al. (1993) melaporkan bahwa gen fekunditas Booroola domba berasosiasi dengan penciri genetik (linked to
markers) pada daerah kromosom 9 manusia. Sampai saat ini identifikasi gen,
dimungkinkan untuk menggunakan peta perbandingan dengan jenis hewan budidaya lain (Archibald 1998; Cockett et al. 2001), dengan membandingkan pada sederetan gen yang telah dipetakan pada manusia dan mencit (Cockett et al. 2001). Penelitian pada persilangan ITT dan Merino ini juga menggunakan peta genetik manusia untuk melihat homologi lokasi kromosom (ASGMWS 2004) pada identifikasi gen kandidat. QTL teridentifikasi pada kromosom 6 domba ini terletak homolog dengan kromosom 4 manusia (Tabel 13).
Sementara itu Walling et al. (2000) melaporkan bahwa hewan pembawa satu alel Booroola menjadi lebih ringan berat badannya dari pada hewan bukan pembawa gen Booroola. Dikatakan lebih lanjut bahwa gen Booroola tidak mempunyai efek
pleitropic atau bertanggung jawab pada sejumlah tampilan fenotipik yang berbeda,
tetapi mungkin berasosiasi dekat dengan QTL berpengaruh pada pertumbuhan dari lahir sampai berat sapih. Diistilahkan bahwa QTL sifat pertumbuhan dapat membonceng atau hitchhike dengan alel Booroola selama program introgresi (Walling et al. 2000). Introgresi adalah penyusupan gen sedikit demi sedikit dalam kurun waktu yang lama. Walling et al. (2000) mengindikasikan bahwa sifat pertumbuhan tidak dipengaruhi QTL setelah sapih.
Hasil studi pada populasi backcross (Merino dengan ITT) ini manambah informasi yang menunjukkan adanya kontribusi pada kedekatannya dengan gen
Booroola terletak kromosom 6 domba (p<0.05). Hal yang mendukung pernyataan
tersebut karena pada populasi silang balik (backcross) yang digunakan dalam studi analisis QTL ini berasal dari dua sub-jenis Merino dan domba Indonesia ekor tipis, dimana keduanya Merino (Davis et al. 1982) dan Ekor tipis Indonesia termasuk domba priangan (Tiesnamurti 2001) adalah galur domba mewarisi sifat fekunditas tinggi atau termasuk domba peridi (jumlah anak per satu kelahiran lebh dari 1). Dengan demikian diindikasikan penemuan pengaruh QTL pertumbuhan pada kromosom 6 pada studi ini juga berhubungan dengan sifat fekunditas tinggi yang pewarisannya berasal dari domba Merino dan domba lokal ekor tipis. Penemuan pada studi ini juga mendukung penemuan Walling et al. (2000) yang kemungkinan gen pertumbuhan membonceng dengan gen Boorrola. Gen Booroola pertama kali diidentifikasi pada domba Merino Booroola yang menunjukkan peningkatan laju oulasi yang kuat (Davis et al. 1982).
Studi lain dilaporkan oleh Diez-Tascon et al. (2001) bahwa pengaruh QTL pada kromosom 6 domba berpengaruh untuk sifat produksi susu yang teridentifikasi pada domba Spanish Churra. Mereka menggunakan analisis interval mapping dengan 8 keluarga tiri (half-sibs), significance threshold diestimasikan melalui uji permutasi diikuti koreksi untuk multiple testing. Lebih lanjut disebutkan bahwa daerah (region)
pada kromosom 6 domba berdekatan dengan kelompok Casein dengan pengaruh utama pada susu yaitu persentase protein. Selain itu, perolehan pada domba Churra ini berdekatan dengan data yang tersedia untuk sapi sebagai konsekuensi pemetaan komparatif untuk jenis yang berkaitan dalam ruminansia. Diperolehnya banyak sifat yang bervriasi pada kromosom 6, kemungkinan disebabkan oleh pengaruh sifat
polygene yang kebanyakan tedapat pada pewarisan sifat kuantitatif (Nicholas 1996).
Pada studi ini, pengaruh QTL juga teridentifikasi nyata (p<0.05) setelah uji
permutate-chromosome wide pada kromosom domba 23 untuk sifat pertumbuhan
berat hidup 270 (BB270) dan 360 hari (BB360). Sebanyak 4 penciri mikrosatelit
digunakan untuk menggenotipe alel (genotyping) pada kromosom 23, lokasi ke empat penciri mikrosatelit dipaparka pada Gambar 7.
Gambar 7. Lokasi penciri mikrosatelit digunakan dalam analisis alel (genotyping) pada kromosom 23, QTL teridentifikasi diperkirakan berlokasi diantara penciri apit
berhuruf tebal (CSSM031 dan MCM136)
Hasil analisis menunjukkan bahwa QTL teridentifikasi untuk karakter berat badan pasca sapih (BB180, BB270 dan BB360, Tabel 13) pada penelitian ini terletak diantara penciri apit CSSM031 dan MCM136 pada jarak antara 42,8 dan 65,8 cM (Gambar 7).
Pengaruh QTL pada studi ini juga diidentifikasi pada kromosom 5 (p<0.01) untuk berat lahir (BL), pada kromosom 7 (p<0.01) untuk berat badan umur 90 (BB90)
dan pada kromosom 8 (p<0.05) untuk berat badan umur 270 (BB270) dan umur 360
(BB360) setelah uji chromosome-wide (p<0.05). Teridentifikasinya QTL yang lebih
beragam pada penelitian ini, kemungkinan berhubungan dengan rancangan hewan percobaan yang digunakan menggunakan persilangan dari dua sub-jenis domba yang
sangat berbeda baik dari faktor klimat (sub tropis dan tropis) maupun sifat karakteristik berat badan yang berbeda secara genetis. Persilangan ke dua sub-jenis yang sangat berbeda ini diperkirakan mewariskan pola penyebaran alel yang lebih beragam atau lebih polimorfik pada variasi genetik (de Koning et al. 2001). Warisan berat hidup yang lebih beragam ini diperkirakan berasal dari ayah (paternal). Lebih variatif ini, kemungkinan juga didukung dari populasi domba progeny yang digunakan dalam studi ini diperoleh dari turunan pustaka keluarga tiri (half-sib) dimana disusun dari satu garis ayah (F1 sire) dengan banyak ibu (dam) pada setiap keluarga acuannya. Pengaruh pembentukan populasi atau rancangan hewan percobaan dalam studi QTL ini juga ditunjukkan oleh peneliti sebelumnya. de Koning
et al. (2001) melaporkan bahwa analisis regresi dengan model paternal half-sib
(Knott et al. 1996) menunjukkan pengaruh nyata QTL untuk kualitas warna daging pada babi terletak pada kromosom 3, 4 dan 13. Sementara dengan pendekatan
line-cross (Haley et al. 1994) antara Meishan dan Dutch large white dan galur Landrace
untuk mendapatkan populasi F2 sebagai animal experimental design ternyata gagal dalam pendeteksian QTL pada kualitas warna daging babi. Disimpulkan oleh de Koning et al. (2001) bahwa kelemahan rancangan line-cross mempunyai frekuensi alel berasal dari tetuanya (founder) adalah serupa pada pengaruh QTL sifat warna daging babi. Dengan demikian dapat disimpulkan bahwa pemilihan rancangan hewan percobaan untuk penelusuran lokasi QTL sifat kuantitatif penting.
Gen Kandidat pada Kromosom 5, 7 dan 18
Keseluruhan analisis QTL pada penelitian ini, terdapat 3 lokasi kromosom berbeda yang menunjukan pengaruh kuat QTL pada sifat pertumbuhan, yaitu pada kromosom 5, 7 dan 18. Terlebih pengaruh nyata QTL masih tetap kuat dipertahankan setelah uji permutate-experiment wide terutama pada kromosom 18 (p<0.01) dan kromosom 7 (p<0.05). Oleh karena itu pada investigasi gen difokuskan hanya pada ke tiga kromosom tersebut.
Akses GenBank
Kemajuan teknologi informasi telah memberi kemudahan para peneliti bidang molekuler untuk menyelidiki lebih luas dan lebih dalam sampai tingkat molekuler. Seperti diketahui bahwa genom manusia telah diidentifikasi lebih lengkap dari kelompok hewan. Berdasarkan rekomendasi the International Society of Animal
Genetics, loci yang terpetakan pada domba atau genom hewan budidaya (farmed animal genomes) lainnya diberi nama dengan petunjuk nomenklatur manusia (Broad et al. 1998). Petunjuk pada manusia ini dirancang untuk mempermudah dalam
identifikasi keragaman pada domba, termasuk identifikasi gen dan data runutan nukleotida pada peta genomnya. Nama dan simbol gen manusia dapat diperoleh melalui database dari situs: http://www.gene.ucl.ac.uk/nomenclature/. Petunjuk singkat peraturan kesamaan (homologues) dengan jenis lain akhir – akhir ini telah dipublikasikan oleh White et al. (1997).
Pada studi QTL ini, guna mengetahui lebih lanjut gen yang berperan pada sifat pertumbuhan perlu mengacu pada kemajuan genom manusia. Pada awal studi ini setelah diketahui posisi kromosom untuk melihat pengaruh QTL (melalui QTL Express: http://qtl.cap.ed.ac.uk) dan setelah dikonversi homologinya dengan kromosom manusia serta didapatkan satuan ukuran dalam Mega base (Mb) untuk mempermudah investigasi gen melalui situs Australian Sheep Gene Mapping: http://rubens.its.unimelb.edu.au/~jillm/jill.htm. Selanjutnya untuk mengidentifikasi gen kandidat yang berperan pada sifat pertumbuhan seperti diterangkan sebelumnya, maka perlu mengacu pada human genom dengan akses GenBank melalui situs: http://www.ncbi.nih.gov.
Segmen dalam Mb pada kromosom manusia dimaksudkan sebagai petunjuk secara keseluruhan lokasi gen, sementara itu penciri apit (flanking markers) yang mengapit keberadaan lokasi gen atau penciri berdekatan dengan loci dapat mempersempit investigasi gen berhubungan dengan QTL sifat pertumbuhan. Oleh karena itu, penelusuran gen pada studi ini mengacu pada perolehan penciri apit (flanking markers) yang secara rinci ditampilkan pada Tabel 13. Penciri apit pada Tabel 13, memaparkan semua QTL pertumbuhan yang terdeteksi pada 15 lokasi yang
terletak pada 7 kromosom (No. 2, 5, 6, 7, 8, 18 dan 23). Namun pada investigasi gen ini hanya dikonsentrasikan pada 3 kromosom yang secara statistik menunjukkan nilai ambang nyata (significance threshold) pada taraf 1% (p<0.01). Ke tiga kromosom tersebut yaitu kromosom nomor 5, 7 dan 18. Pasangan penciri apit dari ke tiga kromosom tersebut termasuk kesamaan (homologies) dengan kromosom manusia dan lokasi gen kandidat pada segmen kromosom manusia (Mb), dipaparkan pada Tabel 14.
Tabel 14. Sheep mapping website
Krom
Domba (flanking markers) Penciri Apit Prediksi (cM)* Manusia Krom Segmen (Mb)**
5 MCM527-BMS1247 125,5-157,00 5 82,361– 106,889 7 BM3033-RNS/BRN 0,00-46,90 14 0 – 22,077 7 RNS/BRN-BMS1620 46,9-105,8 3 15 15 15 14 14 14 0 0 40,459 42,556 – 61,133 22,077 - 38,954 41,295 – 44,767 46,190 – 64,816 18 TMR1/AKT1 CSSM018- 106,400-123,900 14 98,509 – 105,138
* Prediksi (cM)= prediksi lokasi gen pada kromosom domba
**Segmen (Mb), lokasi gen pada rentangan kromosom manusia, setelah dilakukan analisis melalui ASGMWS
Deskripsi Gen
Deskripsi gen dilakukan dengan akses GenBank dengan melihat segmen atau daerah yang diapit oleh ke dua titik penciri loci (flanking markers) dan hanya pada ke tiga kromosom (Tabel 14). Dua hal yang harus diketahui sebelum mendeteksi gen, yaitu pengertian segmen dan penciri apit loci berasosiasi dengan gen (linked to
genes). Segmen hanya menunjukkan lokasi putative genes secara keseluruhan,
sementara daerah yang diapit oleh ke dua titik penciri loci merupakan daerah putative
genes yang lebih mendekati ketepatan. Posisi homologi untuk kromosom 5, 7 dan 18
domba dengan menunjukkan segmen atau lokasi yang diapit oleh 2 titik penciri apit pada kromosom manusia untuk ke tiga kromosom domba (Gambar 8, 9 dan 10).
Gambar 8. Lokasi gen kandidat pada kromosom 5 domba terletak diantara penciri apit MCM527 – BMS1247 dan homolog dengan kromosom 5 manusia
Gambar 9. Lokasi gen kandidat pada kromosom 7 domba terletak diantara penciri apit BM3033 – RNS5/BRN dan antara RNS5- BMS1620, dan homolog
dengan kromosom 14 dan 15 manusia (http://rubens.its.unimelb.edu.au/~jillm/jill.htm)
Gambar 10. Lokasi gen kandidat pada kromosom 18 domba terletak diantara penciri apit CSSM018 –TMR1/AKT1, homolog dengan kromosom 14 manusia
(http://rubens.its.unimelb.edu.au/~jillm/jill.htm)
Terdapat kerumitan dalam investigasi gen tersebut karena kromosom domba sebelumnya harus dihomologkan dengan kromosom manusia. Sebagai contoh, pengaruh QTL untuk sifat pertumbuhan terlihat pada kromosom 7 baik untuk BB90
(p<0.01) dan BB270 dan BB360 (p<0.05). Kromosom 7 domba mempunyai homologi
dengan kromosom manusia 3, 14 dan 15. Sementara itu pada kromosom 7 domba yang pertama untuk BB90 (p<0.01) yaitu hanya terletak homologi dengan kromosom
manusia 14 dan tidak menempati segmen kromosom manusia manapun. Pada kromosom 14 tersebut, penciri loci yang mengapit adalah BM3033-RNS5/BRN terletak pada segmen 0 sampai 22.077 Mb. Sementara itu pada kromosom 7 lainnya (BB270 dan BB360; p<0.05), keduanya diapit oleh dua titik penciri loci RNS/BRN dan
BMS1620 dan homolog dengan kromosom manusia nomor 3, 14 dan 15 dengan segmen yang diapit (setelah dirapikan) berlokasi mulai 0 Mb (tidak ada atau belum diketemukan gennya) untuk segmen pada kromosom 3, terletak antara 20,227-64,816
Mb untuk segmen pada kromosom 14 dan terletak antara 0,00-60,325 Mb untuk segmen pada kromosom 15 (Tabel 14). Segmen dengan satuan ukuran Mb pada kromosom manusia sebelum dirapikan terletak sangat tidak teratur atau campur aduk (jumble up), lihat Lampiran 8. Segmen (Mb) pada kromosom homologi lainnya, yaitu kromosom 5 (p<0.01) domba homolog dengan kromosom 5 manusia (Lampiran 7) dan kromosom 18 (p<0.01) domba homolog dengan kromosom 14 manusia, juga terletak sangat tidak teratur (Lampiran 9).
Hal lain yang perlu diperhatikan pada investigasi gen kandidat yaitu pembacaan deskripsi gen pada Online Mendelian Inheritance of Man (OMIM) dari situs NCBI (http://www.ncbi.nih.gov). Investigasi gen hanya mengacu pada prediksi gen yang disertai OMIM, dan hanya yang memuat keterangan yang berasosiasi dengan pertumbuhan (sesuai dengan tujuan penelitian) yang dipertimbangkan sebagai
putative genes untuk pertumbuhan. OMIM memuat hasil penelitian sebelumnya yang
menyertakan keterangan fungsi gen, dan lebih jauh tentang ukuran gen, runutan basa gen (sequence) deskripsi molekuler lainnya. Hal ini diperlukan untuk penelitian lebih lanjut bilamana ingin melakukan kloning dan sebagainya.
Pada akses GenBank, yang diperhatikan hanya lokasi segmen yang diapit oleh 2 titik penciri apit. Pada akses tersebut tidak selalu mendapatkan lokasi persis sesuai dengan panjang rentangan diantara 2 penciri apit. Tampilan pada akses GenBank untuk masing-masing kromosom disertai simbol loci yang menandakan linked to
genes.
Hasil investigasi gen untuk masing-masing lokasi QTL pada kromosom 5, 7 dan 18 yang telah dihomologkan dengan kromosom manusia, secara rinci informasi gen kandidat pada masing-masing segmen kromosom manusia dapat dilihat pada Lampiran 10, 11a, 11b dan 12. Pada lampiran tersebut dipaparkan runutan segmen secara rinci beserta keberadaan gen kandidat yang tidak secara penuh menutup sepanjang rentang segmen namun ada beberapa jarak (interval) yang belum ditemukan gennya. Sebagai contoh pada kromosom 7, estimasi keberadaan gen kandidat terletak sangat tidak teratur, terutama pada homologinya dengan kromosom 14 manusia (Tabel 14).
Gen Kandidat pada Kromosom 5. Gen kandidat diperkirakan terletak diantara
dua titik penciri apit MCM527-BMS1247 pada segmen kromosom 5 manusia antara 82,361– 106,889 Mb (Tabel 14). Dari 14 referensi Online Mendelian Inheritance of
Man (OMIM) pada kromosom 5 (Lampiran 10), setelah dianalisis teridentifikasi satu
gen kandidat yang berasosiasi dengan sifat pertumbuhan. Gen kandidat tersebut yaitu dengan simbol CAST pada locus 5q15-q21 pada segmen antara 96,057-96,136Mb dengan deskripsi Calpastatin. Pada referensi OMIM diterangkan bahwa Calpastatin adalah penghambat alami Calpain, aktivitas Calpain ditunjukkan pada distropi otot atau necrosis otot (OMIM 2005). Ideogram gen CAST yang dipetakan pada lokasi 5q15-q21 dapat dilihat pada Gambar 11. Menurut Nonneman et al. (1999) bahwa
Calpastatin adalah spesifik, penghambat endogenous dari calpain (kerjanya
tergantung keberadaan calcium secara alami) yang berperan pada pengaturan proteolisis otot. Diterangkan oleh Palmer et al. (1998) bahwa fungsi Calpastatin yaitu berperan mengatur pertumbuhan otot dan pengaturan dalam keempukan daging setelah pemotongan (postmortem). Keempukan daging postmortem dihubungkan dengan degradasi rataan calpastatin pada otot postmortem (Delgado et al. 2001).
Gambar 11. Ideogram lokasi gen CAST pada 5q15-q21 (96,057-96,136Mb) Menurut Spencer & Mellgren (2002) Ekspresi Calpastatin yang berlebihan ditemukan pada mencit transgenik (mdx mice). Daerah exon dan intron gen
menggunakan primers yang didasarkan pada sekuens sapi (Killefer & Koohmaraie 1994, GenBank accession no. L14450) dan gen Calpastatin domba (Collingwood et
al. 1992). Perbedaan atau polimorfisme tersebut dapat digunakan untuk marked-aasissted selection (MAS) domba dengan kualitas daging yang berbeda. Penggunaan
MAS dalam aplikasi seleksi genetik untuk keempukan daging ini dapat digunakan dengan uji DNA yang dapat mengungkapkan apakah hewan yang diuji membawa spesifik gen yang berasosiasi dengan keempukan daging (Smith 2003). Lebih lanjut Smith (2003) mengemukakan bahwa telah tersedia di Amerika uji keempukan dengan GeneSTAR (R) yang dikembangkan di Australia dan TenderGENE (R) yang dikembangkan di Amerika untuk memberi jaminan kepada konsumen atas keempukan daging.
Pada studi lainnya, telah dideskripsikan fungsi gen Calpastatin. Dilaporkan bahwa penyebab terjadinya kekerasan pada perototan domba yang lebih tua karena adanya tingkat Calpastatin yang tinggi (Freking et al. 1999; Duckett et al. 2000). Dilaporkan oleh Smith (2003) bahwa uji di atas berkenaan dengan perbedaan gen yang dihubungkan dengan keberadaan enzim secara normal atau alami (Calpastatin dan Calpain) yang berpengaruh pada keempukan daging normal yang terjadi selama pelayuan postmortem (post-harvest aging). Lebih lanjut Smith (2003) menyatakan bahwa uji keempukan dengan GeneSTAR dapat mengidentifikasi keberadaan 2 macam gen Calpastatin, satu dihubungkan dengan sifat keempukan sementara yang lainnya dihubungkan dengan kekerasan daging.
Teridentifikasinya QTL sifat pertumbuhan berat badan pada kromosom 5 dari studi ini yaitu dengan ditemukannya gen Calpastatin, maka memperjelas laporan Palmer et al. (1998) yang menyatakan tidak dikenalnya gen Calpastatin pada domba pada masa atau waktu penelitian tersebut. Sementara Hediger et al. (1991) dan Crawford et al. (1995) hanya menyatakan didapatkannya ovine gene terletak pada kromosom 5. Sementara pada sapi, gen Calpastatin teridentifikasi pada kromosom 7 dengan posisi 117,8cM (Bishop et al. 1993; Kappes et al. 1997). Studi terakhir, gen
Calpastatin juga ditemukan pada kromosom 2 babi yang mempengaruhi sifat kualitas
Peningkatan aktivitas Calpastatin dan pada penurunan degradasi protein kelihatannya dihubungkan dengan pertumbuhan otot pada sapi (Morgan et al. (1993) dan anak domba Callipyge (Lorenzen et al. 2000). Lebih lanjut Lorenzen et al. (2000) melaporkan bahwa peningkatan pada calpastatin otot telah diketahui selama pertumbuhan anak domba Callipyge dan pada anak domba yang diberi makan growth
promotant beta-agonist, (Speck et al. 1993). Ditemukannya gen Calpastatin dengan demikian dapat diasosiasikan dengan kemungkinan adanya ekspresi gen Callypige pada kromosom 5 yang diperoleh dari studi ini.
Gen Kandidate pada Kromosom 7. Pengaruh QTL terhadap sifat pertumbuhan
yang terdeteksi pada kromosom 7 ini terletak pada 2 segmen yang masing-masing diapait oleh sepasang penciri apit berbeda. Kedua segmen tersebut masing-masing pada sifat pertumbuhan untuk berat badan 90 atau BB90 (p<0.01) yang diapit oleh
BM3033-RNS/BRN dan untuk BB270 dan BB360 (p<0.05) yang diapit oleh
RNS/BRN-BMS1620. Hasil analisis gen dengan akses GenBank-NCBI pada kedua segmen tersebut dijelaskan seperti pada paparan berikut di bawah ini.
Gen Kandidat diantara BM3033 - RNS/BRN. Gen kandidat pada segmen pertama dari kromosom 7 yang diapit oleh dua titik penciri apit BM3033 - RNS/BRN terletak homolog dengan kromosom 14 manusia pada segmen antara 0 – 22,077Mb (Tabel 14). Hasil analisis dari GenBank-ncbi menunjukkan bahwa gen kandidat yang telah ditemukan terletak diantara 18,241 - 22,077 Mb (Lampiran 11a). Pada lampiran 11a dipaparkan secara rinci jumlah gen yang sudah ditemukan sebanyak 208 terletak pada rentangan 18,241 - 22,077 Mb dan 17 gen diantaranya sudah terdapat referensi OMIM. Dari ke 17 referensi OMIM tersebut tidak ditemukan gen kandidat yang berasosiasi dengan sifat pertumbuhan yang diteliti. Dengan demikian pada segmen pertama dari kromosom 7 domba (antara penciri apit BM3033 - RNS/BRN) tidak diketemukan gen pertumbuhan. Hal ini diperkirakan tidak cukup jumlah penciri mikrosatelit yang digunakan untuk menutup kromosom 7 dalam genotyping. Pada penelitian ini hanya menggunakan 5 penciri mikrosatelit yang digunakan untuk menutup kromosom 7 domba (Lampiran 1). Kemungkinan lain diperkirakan dengan
menggunakan comparative mapping, belum semua gen yang ditemukan pada domba telah teridentifikasi pada kromosom manusia. Kemungkinan perlu dilakukan analisis dengan mengacu pada pemetaan hewan domestik ruminansia apabila peta genetik hewan ruminansia (missal sapi) telah lebih lengkap dipetakan.
Gen Kandidat diantara RNS/BRN-BMS1620. Gen kandidat pada segmen kedua dari kromosom 7 yang diapit oleh dua titik penciri apit RNS/BRN-BMS1620 terletak homolog dengan kromosom 3, 14 dan 15 manusia pada segmen (Mb) yang secara rinci ditampilkan pada Tabel 14. Hasil analisis dari GenBank-NCBI menunjukkan bahwa belum ditemukan gen kandidat pada kromosom 3 manusia (=0Mb). Sementara gen kandidat yang telah ditemukan pada kromosom 14 dan 15, dipaparkan pada Lampiran 11b.
Kromosom 14 Manusia. Keseluruhan paparan gen pada kromosom 14 manusia
dapat dilihat pada Lampiran 11b. Dari keseluruhan gen yang ditemukan pada kromosom 14 manusia tersebut, terdeteksi satu (1) gen kandidat yang berasosiasi dengan sifat pertumbuhan. Gen kandidat yang terdeteksi tersebut dinotasikan dengan simbol SSTR1 dan dideskripsikan sebagai gen Somatostatin Receptor1 yang persisnya terentang pada segmen 37,746-37,752Mb dengan lokasi locus pada 14q13. Ideogram lokasi gen SSTR1 dapat dilihat pada Gambar 12.
Deskripsi pada OMIM 182450 (2005) diterangkan bahwa somatostatin atau SST menggunakan kekuatan pengaruh biologinya dengan mengikat pada penerima khusus yang mempunyai afinitas tinggi (specific high-affinity receptors), dalam beberapa kasus diikat dengan GTP-binding proteins. Somatostatin atau SST adalah
tetradecapeptide pertama kali diisolasi dari ekstrak hipotalamik dan menunjukkan
penghambat kuat pada sekresi hormon pertumbuhan dari anterior pituitary (OMIM 182450 2005). Somatostatin tersebar luas pada sistem syaraf pusat dan jaringan permukaan (peripheral tissues) seperti perut, usus dan pancreas. Reaksi atau aksinya pada banyak tempat untuk menghambat pelepasan banyak hormon dan sekresi protein lainnya. Dua somatostatin receptors yang berbeda adalah SSTR1 dan SSTR2, masing-masing terdiri dari 391 dan 369 asam amino (Yamada et al. 1992). Menggunakan analisis segregasi pada satu panel reduced human-hamster somatic cell hybrids, Yamada et al. (1992) menetapkan lokasi gen SSTR1 pada kromosom 14 dengan lokasi 14q13. DNA polimorfisme short tandem repeat (STR) yang sangat informatif telah diidentifikasi terdapat pada SSTR1 (OMIM 182451. 2005).
Pada kamus medis (Anonymous 2005), diterangkan bahwa somatostatin sebagai suatu hormon polypeptide yang dihasilkan pada hypothalamus atau jaringan lain dan organ. Somatostatin menghambat pelepasan hormon pertumbuhan Somatotropin dan juga mengatur fungsi fisiologi penting dari ginjal, pancreas dan saluran pencernaan (gastrointestinal). Somatostatin receptor diekspresikan secara luas diseluruh tubuh, bertindak sebagai pemancar syaraf (neurotransmitter) pada sistem saraf pusat dan permukaan. Dengan demikian teridentifikasinya gen somatostatin receptor1 pada studi ini diperkirakan dapat mengatur pelepasan hormon pertumbuhan. Seperti diketahui bahwa pada mekanisme pelepasan hormon pertumbuhan dikontrol oleh factor hipotalamus yang diketahui sebagai growth hormone releasing factor. Hormon pertumbuhan disekresikan oleh pituitary, mempunyai pengaruh pada otot yang diatur melalui IGF-I (Insulin-like Growth Factor-I) dan binding sites untuk IGF-I. Hormon pertumbuhan saling berpengaruh dengan receptor pada sel membran hati, ginjal dan otot yang menyebabkan pelepasan IGF-I pada peredaran atau sirkulasi darah. IGF-I dapat memicu proliferasi dan diferensiasi myoblast. Myoblast pada perkembangan
embrio hewan merespon terhadap perbedaan sinyal yang mengontrol proliferasi dan migrasi sel.
Kromosom 15 Manusia. Keseluruhan paparan gen pada kromosom 15 manusia
dapat dilihat pada Lampiran 11b. Dari keseluruhan gen yang ditemukan pada kromosom 15 manusia tersebut, terdeteksi satu (1) gen kandidat yang berasosiasi dengan sifat pertumbuhan. Gen kandidat yang terdeteksi tersebut dengan simbol
CAPN3 dan dideskripsikan sebagai gen Calpain3 yang persisnya terentang pada
segmen 40,427-40,491Mb dengan lokasi locus pada 15q15.1-q21.1. Ideogram lokasi gen CAPN3 dapat dilihat pada Gambar 13.
Gambar 13. Ideogram lokasi gen CAPN3 pada 15q15.1-q21.1 (40.427-40.491Mb) Pada OMIM 114240 (2005) diterangkan bahwa Calpain atau enzim protease netral yang diaktifkan oleh ion calcium (Ca) atau enzim yang memerlukan calcium untuk aktivitas katalisnya (OMIM 11420. 2005). Diterangkan lebih lanjut enzim tersebut adalah enzim cystein intraseluler nonlysosomal. Enzim proteolitik
nonlisosomal bekerja aktif pada pH rata-rata lebih tinggi (• 6.5) dari enzim proteolitik lisosomal (<6.5) pada awal pelayuan (Etherington 1984).
Termasuk dalam Calpain hewan mamalia yaitu CAPN1 (OMIM 114220. 2005)
otot (muscle-specific). Dilaporkan oleh Richard et al. (1995) bahwa gen CAPN3 terdiri dari 24 exons dengan ukuran 40 kb. Ohno et al. (1989) memetakan gen
CAPN3 pada kromosom 15 manusia. Dalam peta fisik dan genetik dari daerah
kromosom 15 mengandung gen untuk limb-girdle muscular dystrophy type 2A atau LGMD2A (OMIM 253600. 2005) dan gen kandidat LGMD2A tersebut telah diisolasi. Richard et al (1997) menerangkan bahwa Limb-girdle muscular dystrophies
(LGMDs) adalah satu grup dari penyakit otot saraf (neuromuscular) menunjukkan
heterogen klinis sangat besar (great clinical heterogeneity). Penyakit ini jarang terjadi, merupakan penyakit cacat molekuler (molecular defect) yang mempunyai keragaman luas.
Dengan penapisan mutasi pada grup LGMD2A, Richard et al. (1995) telah mengidentifikasi 15 mutasi (nonsense, splice site, frameshift atau missense) CAPN3 yang mengadakan segregasi secara bersamaan (co-segregating) dengan sifat penyakit (OMIM 114240. 2005). Gen CAPN3 adalah gen kandidat yang menarik sebab fungsinya pada otot (OMIM 114240. 2005) atau gen yang menyandi calpain khusus pada otot (Richard et al. 1997). Aktivitas m-calpain (proenzyme calpain2) lebih besar pada otot biceps femoris dan longissimus dari domba Callipyge dari pada domba normal (Delgado et al. 2001). Ditemukannya gen CAPN3 pada penelitian ini, dengan demikian ada kemungkinan secara tidak langsung lokasi QTL pada kromosom 7 yang diapit oleh penciri RNS/BRN-BMS1620 menunjukkan ada hubungannya dengan kualitas daging.
Gen Kandidat pada Kromosom 18. Estimasi keberadaan gen kandidat dari pengaruh QTL yang berlokasi pada kromosom 18 terdeteksi kuat (p<0.01) setelah uji
permutate-chromosome wide dan masih tetap dipertahankan (p<0.01) setelah uji permutate-experiment wide. Gen kandidat diperkirakan pada rentangan 98,508 –
105,008 Mb dari kromosom 14 manusia yang diapit oleh penciri loci CSSM018-TMR1/AKT1 (Tabel 14). Dari rentangan segmen tersebut ditemukan 102 simbol gen dengan 32 diantaranya dengan referensi OMIM. Secara rinci deskripsi untuk semua simbol gen tersebut dapat dilihat pada Lampiran 12. Dari ke 32 gen kandidat yang
terdapat pada rentangan segmen 98,508 – 105,008 Mb kromosom 14 manusia belum diketemukan gen yang berasosiasi dengan sifat pertumbuhan. Hal ini kemungkinan disebabkan karena pada analisis gen kromosom 18 domba hanya diketahui mempunyai satu daerah (region) homolog pada kromosom manusia, yaitu kromosom 14 manusia. Pada kromosom 14 manusia tersebut hanya diwakili pada rentangan segmen 98,508 – 105,008 Mb (Tabel 14). Oleh karenanya, diperkirakan masih banyak yang belum terdeteksi gen kandidat yang ditemukan pada peta komparatif manusia, termasuk di dalamnya gen kandidat untuk sifat produksi domba yang belum diketemukan.
Pada studi QTL ini, sebagian besar sifat pertumbuhan berat badan (BB90, BB180,
BB270, dan BB360) menunjukkan hasil yang konsisten dengan pendapat Cockett et al.
(2001) dan Walling et al. (2001; 2004) yaitu lokasi QTL untuk sifat pertumbuhan terletak pada kromosom 18. Lokasi QTL yang terdeteksi pada kromosom 18 diapait oleh penciri mikrosatelit CSSM018 dan TMR1/AKT1 (Gambar 6). Penemuan ini didukung oleh studi sebelumnya yang melaporkan bahwa locus pada kromosom 18 yang berdekatan dengan penciri CSSM018 dikatakan menyebabkan peningkatan perototan baik pada daerah longissimus dorsi maupun peningkatan berat pada rusuk ke sebelas domba (Nicoll et al. 1998). Lebih lanjut disebutkan bahwa locus tersebut dinamakan locus Carwell.
Teridentifikasinya QTL pada penelitian ini diantara ke dua penciri tersebut (CSSM018-TMR1/AKT1) memperkuat perolehan Cockett et al. (1994) yang menyatakan bahwa penciri CSSM018 ternyata sesuai dengan keterpautan penciri yang ditempati oleh gen Callipyge. Locus gen tersebut terletak dibagian akhir kromosom 18 domba. Lebih lanjut dilaporkan oleh Cockett et al. (1994) bahwa lokasi
Callipyge tersebut berpaut dengan penciri TGLA122, CSSM18 dan GMBT16
sedangkan lebih tepat gen tersebut dipetakan pada jarak 4,6 cM diantara penciri IDVGA30 dan OY3. Seperti dinyatakan oleh Dominik (2005a) bahwa terdapat keterpautan antara landmark penciri genetik pada kromosom terhadap gen sifat kuantitatif. Dilaporkan oleh Walling et al. (2004) bahwa QTL yang berasosiasi dengan kedalaman otot terletak pada kromosom 18 dikatakan dekat dengan locus gen
Callipyge dan Carwell. Gen Callipyge pertama kali dideteksi karena pengaruhnya
yang sangat nyata yaitu mempunyai pembesaran perototan atau muscular hypertrophy (Haley 2001). Dilaporkan bahwa ekspresi gen Callipyge mengakibatkan beberapa individu otot (individual muscle) dapat mencapai lebih dari 40% lebih berat pada anak domba Callipyge dibandingkan kontrol (Duckett et al. 2000). Studi akhir-akhir ini telah melakukan penelitian potitional candidates dan diketahui adanya kejadian mutasi secara alami yang menghasilkan tampilan fenotipik yang unik pada domba (Cockett et al. 2001). Keberadaan gen Callipyge ini sebetulnya lebih nyata di kromosom 18 domba seperti yang dilaporkan beberapa peneliti sebelumnya (Cockett
et al. 1996; Freking et al. 1998, Haley 2001).
Melihat kekuatan pengaruh QTL untuk sifat pertumbuhan pada kromosom 18 domba yang sangat kuat dan penemuan sebelumnya pada bangsa domba berasal dari daerah dingin, maka diperkirakan bahwa gen pertumbuhan jelas berada pada kromosom 18 domba yang diteliti (persilangan ITT dan Merino).
Gen Callipyge. Suatu sifat yang diwariskan pada ternak (contoh: pada domba) yang menghasilkan terhadap ketebalan daging, kaki belakang lebih berdaging dan oleh karenanya menghasilkan produksi daging per ekor lebih tinggi (Zaid et al. 2001). Asal-usul tentang gen Callipyge yaitu diawali kelahiran dari keturunan domba jantan Dorset Solid Gold pada tahun 1983 dari kawanan domba Moffat dekat Piedmondt, Oklahoma, Amerika. Beberapa hasil keturunannya menunjukkan perkembangan pasca lahir yang luar biasa dan fenotipe dengan otot yang berat atau padat. Sebutan Callipyge berasal dari bahasa Yunani yang berarti pantat yang montok atau nice buttock (Dominik 2005a). Hal ini dikarenakan domba pembawa gen
Callipyge menunjukkan perototan yang berlimpah pada bagian pantat dengan
peningkatan sampai 46% dibandingkan dengan domba bukan Callipyge (Cockett et
al. 1996). Dinyatakan oleh Jackson et al. (1997) bahwa perbandingan fenotipik dari
genotipe yang diturunkan dari perkawinan terancang telah konsisten mengandung alel mutan baru dengan aksi gen non-resesif pada locus autosom dan tunggal. Selanjutnya Cockett et al. (1994) menyebutkan bahwa locus tersebut dinyatakan sebagai
Callipyge (CLPG), dipetakan pada daerah telomer dari kromosom 18 domba. Lebih
lanjut Cockett et al. (1996) menerangkan bahwa aksi gen CLPG didasarkan pada sejumlah terbatas anaknya berasal dari lima kombinasi perkawinan berbeda dimana alel berasal dari Ibu (maternal) dan dari Bapak (paternal) diduga berasal dari 5 penciri DNA pada kromosom 18. Sehubungan dengan CLPG tersebut, terdapat istilah polar
overdominance (Cockett et al. 1996; Freking et al. 1998). Polar overdominance yaitu
menggambarkan karakterisasi genetik dari satu locus yang menunjukkan suatu model pewarisan unik dimana hanya individu anak heterosigot yang mewarisi mutasi dari ekspresi fenotipe bapaknya (sire). Pengaruh fenotipe Callipyge sebelumnya telah diestimasikan dari data yang dikoleksi pada heterosigot atau alel CLPG mutan diwariskan dari Bapak/sire dan anak domba yang bukan pembawa sifat (non-carrier), dengan masing-masing anak domba dikelompokkan sebagai yang mengekspresikan
Callipyge atau yang mempunyai fenotipe normal.
Gen Callipyge adalah suatu mutasi genom domba yang bertanggung jawab pada pembesaran (hyperthrophy) otot yang sangat besar dari serat otot fast-twitch, terutama pada otot pinggul (Koohmaraie et al. 1995; Carpenter et al. 1996). Pembesaran otot ini tidak terekspresi pada anak yang baru lahir dan baru berkembang setelah berumur 3 minggu (Cockett et al. 2001) dengan demikian tidak menimbulkan masalah dalam kelahiran (Jackson et al. 1997). Ternak Callipyge menghasilkan daging tanpa lemak dan persentase karkas yang lebih tinggi (Jackson et al. 1997; Koohmaraie et al. 1995) namun kelenturan atau kelembutannya pada daging pinggang (loin) berkurang (Kerth
et al. 1995; Koohmaraie et al. 1995). Dari studi fisiologi telah dipelajari domba yang
mewarisi gen Callipyge dan dibandingkan dengan domba normal, gambaran jaringan otot longisimus dan domba Callipyge dapat dilihat pada Gambar 14.
Gambar 14. Foto mikro otot longisimus normal (a), Callipyge (b) dan domba
Callipyge (c) (Sumber: Pringle 2005)
Gen Carwell. Gen lain yang dihubungkan dengan peningkatan kualitas karkas
yaitu gen Carwell. Gen Carwell adalah gen yang mengkode pewarisan sifat pada peningkatan pada otot lingkar atau otot mata rusuk (rib-eye muscle= REM) ditemukan pada jenis domba Poll Dorset Australia (Banks 1997). Dibandingkan dengan gen
Callipyge, pengaruh gen REM lebih rendah yaitu hanya 11% peningkatan pada masa
otot dan terbatas pada longissimus dorsi (Marcq et al. 1998; 1999; Jopson et al. 2001). Selain itu, gen Carwell tidak berpengaruh pada penumpukan lemak dengan demikian tidak mempunyai pengaruh negatif pada keempukan daging (Dominik 2005a). Lebih lanjut diterangkan bahwa gen Carwell terletak dekat tetapi bukan gen
Callipyge. Pengaruh serupa pada perototan dan tidak ada perlemakan juga ditemukan
pada kawanan domba Texel Inggris (Walling et al. 2001) dan penciri gen telah diidentifikasi (Dominik 2005a). Kemungkinan gen tersebut adalah variasi dari gen
REM atau Callipyge.
c