IDENTIFIKASI KERAGAMAN GEN FOLLICLE STIMULATING
HORMONE RECEPTOR (FSHR|AluI) PADA SPESIES SAPI
Bos javanicus, Bos taurus, DAN Bos indicus
DENGAN METODE PCR-RFLP
SKRIPSI
SEPTYANINGTYAS ANGGIA SARI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2012
RINGKASAN
SEPTYANINGTYAS ANGGIA SARI. D14080028. 2012. Identifikasi Keragaman Gen Follicle Stimulating Hormone Receptor (FSHR|AluI) pada Spesies Sapi Bos javanicus, Bos taurus, dan Bos indicus dengan Metode PCR-RFLP. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan. Fakultas Peternakan. Institut Pertanian Bogor.
Pembimbing Utama : Prof. Dr. Ir.Cece Sumantri, M.Agr.Sc. Pembimbing Anggota : Dr. Jakaria, S.Pt,M.Si.
Sapi dari spesies Bos javanicus, Bos taurus,dan Bos indicus merupakan sapi-sapi yang tersebar diseluruh wilayah Indonesia. Sumber daya genetik ternak yang harus dipertahankan agar mencegah terjadinya kepunahan menjadi alasan untuk mengkaji keragaman gen FSHR yang berperan dalam fungsi reproduksi. Gen FSHR (Follicle Stimulating Hormone Receptor) akan menghasilkan produk yaitu FSHR yang diperlukan oleh hormone FSH untuk mencapai sel target sehingga dapat melaksanakan fungsi reproduksi (menghasilkan sperma dan ovum). Penelitian ini bertujuan untuk mengetahui keragaman gen Follicle Stimulating Hormone Receptor (FSHR|AluI) dengan menggunakan metode PCR-RFLP pada spesies sapi Bos javanicus, Bos taurus, dan Bos indicus.
Sampel yang digunakan berjumlah 207 sampel yaitu terdiri dari sampel spesies sapi Bos javanicus 59 sampel, spesies Bos taurus 137 sampel, dan spesies Bos indicus 11 sampel. Sampel spesies sapi Bos javanicus berasal dari sapi Bali, sampel sapi Bos taurus terdiri dari sapi Limousin (57 sampel), Simmental (50 sampel), Angus (6 sampel), dan FH (24 sampel), serta sampel spesies sapi Bos indicus terdiri dari sampel Brahman (9 sampel) dan sapi Ongole (2 sampel). Sampel sapi yang digunakan berasal dari BIB Lembang, BIB Singosari, BPTU Bali, BIBD Bali, dan BET Cipelang. Penelitian ini menggunakan metode PCR (Polymerase Chain Reaction) dan RFLP (Restriction Fragment Length Polymorphism) untuk mendeteksi adanya keragaman genetic pada gen FSHR menggunakan enzim rektriksi AluI yang memotong basa AG|CT. Penelitian ini dilakukan di Laboratorium Genetika Molekuler Ternak sejak bulan Oktober 2011 hingga Februari 2012. Keragaman gen FSHR ditentukan dengan analisis frekuensi genotipe, frekuensi alel, nilai heterozigositas pengamatan (Ho), dan
nilai heterozigositas harapan (He).
Amplifikasi gen FSHR menghasilkan fragmen dengan panjang 306 bp yang terletak pada ekson 10 dengan suhu annealing 60oC selama 45 detik. Genotyping gen FSHR|AluI menghasilkan dua jenis alel yaitu alel C dan alel G serta menghasilkan tiga jenis genotipe yaitu CC (243 dan 63 bp), CG (243, 293, 50, 63 bp), dan GG (193, 50, dan 63 bp). Frekuensi genotipe GG tertinggi pada spesies sapi Bos javanicus yaitu 1,00, Bos indicus sebesar 0,46 dan terendah pada spesies sapi Bos taurus yaitu 0,11. Frekuensi genotype CC paling tinggi pada spesies sapi Bos taurus yaitu 0,57, Bos indicus sebesar 0,18, dan terendah pada spesies sapi Bos javanicus. Nilai heterozigositas Ho dan He paling tinggi pada spesies sapi Bos indicus sedangkan nilai
terendah pada spesies sapi Bos javanicus.
ABSTRACT
Identification Polymorphism of The Follicle Stimulating Hormone Receptor (FSHR|AluI) Gene in Species Bos javanicus, Bos taurus, and Bos indicus
Using PCR-RFLP Method Sari, S.A., C. Sumantri, and Jakaria
Follicle Stimulating Hormone Receptor (FSHR) gene is a very important in reproduction trait. FSHR is required by Follicle Stimulating Hormone (FSH) to carry out on target tissues. The purpose of this study was to identify polymorphism of the Follicle Stimulating Hormone Receptor gene in Bos javanicus, Bos taurus, and Bos indicus Cattle. Blood samples used 207 samples consisting of Bos javanicus (Bali Cattle) 59 heads, Bos taurus 137 heads (Limousin 57 heads, Simmental 50 heads, Angus 6 heads, and Holstein Friesian (FH) 24 heads), and Bos indicus 11 heads (Brahman 9 heads and Ongole 2 heads). The polymorphism site analysis from digestion with AluI restriction enzyme allowed the genotype identification. AluI restriction enzyme cut the base position at AG|CT in 193 and 243 bp. Genotyping FSHR gene produced two alleles, namely C and G alleles. These allele produced three genotypes respectively, there are CC (243 and 63 bp), GG (193, 63, and 50 bp), and CG (243, 193, 63, and 50 bp). The highest G allele frequency was found in Bos javanicus (Bali cattle), but the lowest in Bos taurus. The highest C allele frecuency was found in Bos taurus. The value of heterozigosity was the highest in Bos indicus and the lowest in Bos javanicus. The result of this study showed that the allele of FSHR gene in Bos taurus and Bos indicus were polymorphic, while in Bos javanicus was monomorphic.
IDENTIFIKASI KERAGAMAN GEN FOLLICLE STIMULATING
HORMONE RECEPTOR (FSHR|AluI) PADA SPESIES SAPI
Bos javanicus, Bos taurus, DAN Bos indicus
DENGAN METODE PCR-RFLP
SEPTYANINGTYAS ANGGIA SARI D14080028
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2012
KATA PENGANTAR
Puji dan syukur senantiasa penulis dipanjakan kepada Allah SWT atas segala rahmat dan karunia-Nya sehingga penelitian dan penulisan skripsi ini dapat terselesaikan. Skripsi yang berjudul Identifikasi Keragaman Gen Follicle Stimulating Hormone Receptor (FSHR|AluI) pada Spesies Sapi Bos javanicus, Bos taurus, dan Bos indicus dengan Metode PCR-RFLP ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada Fakultas Peternakan, Institut Pertanian Bogor.
Upaya perbaikan produktivitas ternak khususnya dalam reproduksi dapat dilakukan melalui seleksi menggunakan penciri genetik pada level DNA. Sifat reproduksi ternak dikontrol oleh banyak gen salah satunya adalah gen FSHR. Gen FSHR memiliki produk bernama FSHR yang berfungsi membawa hormon FSH ke sel target. Interaksi antara keduanya berfungsi dalam proses spermatogenesis dan oogenesis pada sapi yang berhubungan dengan reproduksi ternak. Teknik PCR-RFLP dapat digunakan untuk mendeteksi adanya keragaman gen FSHR tersebut.
Manfaat dari penelitian ini untuk memberikan informasi mengenai identifikasi keragaman gen FSHR menggunakan metode PCR-RFLP dengan enzim rektriksi AluI pada spesies sapi Bos javanicus, Bos taurus, dan Bos indicus. Informasi mengenai keragaman gen FSHR diharapkan menjadi dasar seleksi pada level DNA. Penulis sangat menyadari penulisan skripsi ini masih jauh dari sempurna, namun penulis berharap semoga skripsi ini dapat memberikan manfaat bagi dunia peternakan. Amin.
Bogor, Juli 2012
RIWAYAT HIDUP
Penulis dilahirkan di kota Depok pada tanggal 25 September 1990. Penulis adalah anak pertama dari dua bersaudara pasangan Bapak Imam Sudarmo dan Ibu Sri Indrayati Faidah. Pendidikan taman kanak-kanak penulis diselesaikan di TK Panca Daya Depok pada tahun 1996. Sekolah dasar diselesaikan pada tahun 2002 di SD Negeri Mekarjaya 8 Depok. Pendidikan menengah pertama diselesaikan pada tahun 2005 di SMP Negeri 4 Depok dan pendidikan menengah atas diselesaikan pada tahun 2008 di SMA Al-Ma’soem Bandung.
Penulis diterima sebagai mahasiswa Fakultas Peternakan, Departemen Ilmu Produksi dan Teknologi Peternakan, Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI) pada tahun 2008. Selama mengikuti pendidikan, penulis aktif di berbagai kepanitiaan dan organisasi. Penulis aktif dalam Tim Manajerial Olahraga Fakultas Peternakan sebagai Manajer Badminton Fapet IPB periode 2010-2012, penulis juga menjadi pengurus Badan Eksekutif Mahasiswa (BEM) Fakultas Peternakan sebagai sekertaris departemen Informasi dan Komunikasi (Infokom) periode 2010-2011, penulis menjadi anggota Himaproter, anggota Animal Breeding and Genetic Student Community (ABGSCi) periode 2011-2012, serta menjadi anggota paguyuban mahasiswa Bandung (Pamaung).
Penulis pernah mengikuti Program Kreativitas Mahasiswa (PKM) di bidang penelitian yang dibiayai Dikti dengan judul “Penggunaan Gen Cyt-b sebagai Pendeteksi Adanya Kandungan Gelatin Babi pada Produk Permen Jelly”. Penulis juga menjadi asisten praktikum mata kuliah Teknologi Pengolahan Telur dan Daging Unggas (TPTDU) tahun ajaran 2011-2012. Penulis berkesempatan menjadi penerima beasiswa PPA (Peningkatan Prestasi Akademik) pada tahun 2010-2011 dan 2011-2012.
DAFTAR ISI Halaman RINGKASAN………. ii ABSTRAK ……… iii LEMBAR PERNYATAAN ……….... iv LEMBAR PENGESAHAN ………. v RIWAYAT HIDUP ………. vi
KATA PENGANTAR ……… vii
DAFTAR ISI ……… viii
DAFTAR TABEL ……….... x
DAFTAR GAMBAR ……… xi
DAFTAR LAMPIRAN ………. xii
PENDAHULUAN ……… 1 Latar Belakang ………... Tujuan ……….... 1 2 TINJAUAN PUSTAKA ……… 3
Asal Usul Sapi di Indonesia ……… Keragaman Genetik ……….. Gen Follicle Stimulating Hormone Receptor (FSHR) ……… Polymerase Chain Reaction – Restrictiom Fragment Length Polymorphism (PCR-RFLP) ……….... 3 6 6 7 MATERI DAN METODE ……… 9
Lokasi dan Waktu ……… Materi ………....
Ternak ……..……… Pengambilan Sampel ………. Ekstraksi DNA ……… Primer ……… Amplifikaasi Gen FSHR|AluI ……… Elektroforesis ……… Genotyping ……… Prosedur ……… Pengambilan Sampel ……… Ekstraksi DNA ……… Amplifikasi DNA ………..………… Elektroforesis ……… Genotyping ……… Rancangan dan Analisis Data………
Frekuensi Alel dan Genotipe ………
9 9 9 9 10 10 10 10 10 11 11 11 12 12 13 13 13
ix Heterozigositas ……… 14
HASIL DAN PEMBAHASAN ……… 15
Amplifikasi Gen FSHR ………. Keragaman Gen FSHR|AluI ………... Frekuensi Genotipe dan Frekuensi Alel Gen FSHR|AluI ………… Heterozigositas ………
15 16 18 22 KESIMPULAN DAN SARAN ………. 24
Kesimpulan ……… Saran ………... 24 24 UCAPAN TERIMAKASIH ………. 25 DAFTAR PUSTAKA ……… 26 LAMPIRAN ……… 29
DAFTAR TABEL
Nomor Halaman
1. Karakteristik Bangsa Sapi pada Beberapa Spesies……… 4 2. Jumlah Sampel yang Digunakan dalam Penelitian ……….. 9 3. Frekuensi Genotipe dan Frekuensi Alel Gen FSHR|AluI ………….... 18 4. Frekuensi Alel Gen FSHR|AluI dari Beberapa Sumber ………. 21 5. Nilai Heterozigositas Pengamatan (Ho) dan Harapan (He)
xi DAFTAR GAMBAR
Nomor Halaman
1. Fenotipik Berbagai Bangsa Sapi (a) sapi Bali, (b) sapi Limousin, (c) sapi Simmental (d) sapi Angus, (e) sapi FH, (f) sapi Brahman
dan (g) sapi PO ………... 5 2. Struktur Gen FSHR ……….………. 7 3. Fragmen Gen FSHR|AluI berdasarkan GenBank ……… 13 4. Visualisasi Amplifikasi PCR Fragmen Gen FSHR menggunakan
Gel Agarose 1,5% ………..
15 5. Hasil Visualisasi PCR-RFLP Gen FSHR|AluI dalam Gel
Agarose 2% ……… 17
DAFTAR LAMPIRAN
Nomor Halaman
xiii LAMPIRAN
PENDAHULUAN
Latar Belakang
Sapi-sapi yang tersebar luas di Indonesia terbagi menjadi tiga kriteria yaitu sapi asli Indonesia, sapi impor, dan sapi yang sudah beradaptasi (Sarbaini, 2004). Secara garis besar, spesies sapi yang tersebar di Indonesia terdiri dari spesies sapi Bos taurus yang merupakan sapi asal Eropa dan Afrika Barat, spesies Bos indicus yang berasal dari wilayah Asia dan Afrika Selatan, serta Bos javanicus yang merupakan spesies asli Indonesia yang berasal dari domestikasi Banteng (Rahmatullah, 2011). Spesies Bos taurus dan Bos indicus merupakan spesies sapi yang di impor ke Indonesia dan beberapa diantaranya telah dapat beradaptasi dengan lingkungan di Indonesia seperti sapi FH. Sapi impor tersebut kemudian disebar keseluruh wilayah Indonesia untuk disilangkan dengan sapi lokal Indonesia dan membentuk bangsa sapi baru seperti sapi Peranakan Ongole.
Kebutuhan akan pangan dan produksi dari ternak sapi di Indonesia menjadi perlu untuk mempertahankan secara optimal sumber daya genetik ternak terutama ternak asli Indonesia. Departemen Pertanian (2006) menyatakan jika sumber daya genetik ternak merupakan substansi ternak yang secara genetik unik, terbentuk dari proses domestikasi yang memiliki nilai potensial serta dapat dimanfaatkan dan dikembangkan dengan baik untuk menciptakan rumpun atau galur unggul. Sapi Bali merupakan salah satu sumberdaya genetik ternak asli Indonesia yang menjadi plasma nutfah nasional. Keberadaan sumberdaya genetik ternak perlu dipertahankan dan dimanfaatkan. Oleh karena itu, perlu adanya upaya peningkatan mutu genetik ternak
Upaya yang dapat dilakukan untuk meningkatkan mutu genetik ternak sapi adalah melalui seleksi dan persilangan. Seleksi dapat dilakukan berdasarkan sifat reproduksi dari masing-masing ternak. Pemanfaatan penerapan teknologi molekuler dapat digunakan sebagai metode seleksi pada tingkat DNA, dalam hal ini DNA yang berperan dalam sifat reproduksi. Salah satu gen yang memiliki peranan penting dalam reproduksi adalah gen FSHR (Follicle Stimulating Hormone Receptor). Gen ini sangat penting dalam proses pembentukan sperma pada sapi jantan dan ovum pada sapi betina. Gen ini terletak pada kromosom ke 11 yang terdiri dari 10 ekson dan 9 intron (Aguirre dan Timossi, 1998). Teknik yang dapat digunakan untuk mengidentifikasi keragaman gen FSHR adalah dengan kombinasi PCR (Polymerase
2 Chain Reaction) dengan RFLP (Restriction Fragment Length Polymorphism). Metode RFLP menggunakan enzim pemotong AluI untuk mengidentifikasi polymorfisme gen FSHR pada ekson 10 dengan menggunakan primer forward dan reverse berdasarkan Houde et al. (1994). Apabila polymorfisme yang terjadi pada gen FSHR berhubungan dengan sifat produksi dan reproduksi sapi, maka dapat dijadikan sebagai Marker Assisted Selection (MAS). Untuk dapat memenuhi kriteria seleksi MAS maka perlu terlebih dahulu dilakukan identifikasi keragaman gen. Melalui seleksi MAS diharapkan ketepatan seleksi dapat lebih baik sejak periode anakan.
Tujuan
Penelitian ini bertujuan untuk mengetahui keragaman gen Follicle Stimulating Hormone Receptor (FSHR|Alu I) dengan menggunakan metode PCR-RFLP pada spesies sapi Bos javanicus, Bos taurus, dan Bos indicus.
TINJAUAN PUSTAKA
Asal Usul Sapi di Indonesia
MacHugh (1996) menyatakan jika terdapat dua spesies sapi yang tersebar diseluruh dunia yaitu spesies tidak berpunuk dari Eropa, Afrika Barat, dan Asia Utara yang disebut dengan spesies Bos taurus, serta spesies sapi berpunuk dari Asia Selatan dan Afrika yang disebut dengan Bos indicus. Perbedaan antara keduanya terletak dari ada/tidaknya punuk, cara beradaptasi (Bos indicus lebih mendiami daerah kering dan panas, sedangkan Bos taurus di daerah beriklim sedang dengan curah hujan cukup).
Sapi yang terdapat di Indonesia terbagi menjadi : 1) sapi asli Indonesia, 2) sapi impor, dan 3) sapi yang telah beradaptasi (Sarbaini, 2004). Kusdiantoro et al. (2007) menyatakan pejantan sapi Ongole di-impor ke Indonesia pada akhir abad ke-19 ke pulau Jawa dan beberapa pulau lainnya di Indonesia kecuali pulau Bali dan Madura. Utoyo (2002) menjelaskan penyebaran sapi Ongole pada abad ke-20 dimaksudkan agar dapat disilangkan dengan sapi asli jawa yang akhirnya membentuk bangsa sapi peranakan ongole dan sapi Madura. Sapi domestikasi yang ada di Indonesia merupakan sapi spesies bibos dan sapi silangan yang berbeda dengan sapi domestikasi di Afrika dan Eropa (Jakaria, 2008).
Menurut Rollinson (1984) sapi Bali (Bibos sondaicus) berasal dari hasil domestikasi banteng liar (Bibos banteng). Proses domestikasi sapi Bali itu terjadi sebelum 3.500 SM di Indonesia atau Indochina. Banteng liar saat ini bisa ditemukan di Jawa bagian Barat dan bagian Timur, di Pulau Kalimantan, serta ditemukan juga di Malaysia (Payne dan Rollinson 1973). Penyebaran sapi Bali di Indonesia dimulai pada tahun 1890 dengan adanya pengiriman ke Sulawesi, pengiriman selanjutnya dilakukan pada tahun 1920 dan 1927. Pada tahun 1964 di Bali terjadi musibah penyakit jembrana secara besar-besaran yang menyebabkan sapi Bali tidak boleh dikeluarkan lagi dari pulau Bali sebagai ternak bibit. Mulai periode inilah sumber bibit sapi Bali bagi daerah lain di Indonesia digantikan oleh NTT, Sulawesi Selatan dan NTB (Talib, 2002).
Bangsa sapi dari spesies Bos taurus yang banyak dimanfaatkan di Indonesia adalah sapi Limousin, Simmental, Angus, dan Brahman. Bangsa sapi dari spesies
4 Bos indicus yang banyak dimanfaatkan adalah sapi Peranakan Ongole dan Brahman. Karakteristik bangsa sapi dari berbagai spesies tercantum pada Tabel 1.
Tabel 1. Karakteristik Berbagai Bangsa Sapi pada Tiga Spesies Spesies Bangsa Sapi Karakteristik Bos
javanicus
Sapi Bali Ciri sapi ini yaitu memiliki warna bulu merah bata kecoklatan hingga berumur 6 bulan dan akan berubah warna (bagi jantan) menjadi coklat tua hingga hitam mencapai dewasa. Sapi ini memiliki warna putih pada bagian pantat, bagian kaki (white stocking) hingga di atas kuku (Williamson dan Payne, 1993). Presentase karkas tinggi (55,85-60,80%), tingkat fertilitas tinggi (83-86%), serta angka kelahiran tinggi (72,6-92,6%) (Martodjo, 1990).
Bos taurus Limousin Sapi ini memiliki karakteristik warna bulu merah keemasan, kaki dan lutut kebawah memiliki warna merah agak muda, dan umumnya terdapat lingkaran berwarna merah muda disekeliling mata (Pane, 1986). Simmental Karakteristik sapi ini memiliki warna bulu kecoklatan
hingga sedikit merah, bulu muka berwarna putih dan bagian ekor serta lutut kebawah berwarna putih pula. Ukuran tanduk tidak begitu besar (Pane, 1986).
Angus Sapi Angus memiliki karakteristik kulit berwarna hitam, tidak bertanduk, tubuh rata, lebar dan dalam, seperti balok, padat dengan urat daging yang baik. Berat badan betina dewasa mencapai 1600 pounds sedang jantan dewasa 2000 pounds (Pane, 1986).
FH Sapi perah ini memiliki warna putih dengan belang hitam, hitam belang putih, sampai warna hitam. Sapi ini merupakan bangsa sapi perah dengan produksi susu paling tinggi (Syarif dan Sumopastowo, 1984).
Bos indicus Brahman Sap ini memiliki karakteristik ponok besar dan kulit yang longgar dengan lipatan kulit dibawah leher dan perut yang lebar, telinga menggantung. Warna kulit merah kecoklatan dengan tanduk melengkung keatas (Balai Pembibitan Ternak Potong Nusa Tenggara Timur, 2009).
PO Sapi ongole memiliki punuk besar dan berglambir (lipatan-lipatan kulit yang terdapat dibagian bawah leher dan perut). Telinganya panjang dan menggantung. Kepala relatif pendek dengan profil melengkung, mata besar dan tenang. Kulit disekitar lobang mata berwarna hitam, tanduk pendek, kadang-kadang hanya bungkul kecil saja (Rachmat, 2001).
5 Perbedaan karakteristik fenotipik dari berbagai bangsa sapi dapat terlihat jelas dengan melihat gambar. Gambar 1 merupakan gambar fenotipik berbagai bangsa sapi dari spesies sapi Bos javanicus, Bos taurus, dan Bos indicus.
(a) (b) (c) (d) (e) (f) (g)
Gambar 1. Fenotipik Berbagai Bangsa Sapi. (a) sapi Bali (Saputra, 2008); (b) sapi Limousin (Khairdin, 2012); (c) sapi Simmental (Khairdin, 2012); (d) sapi Angus (Khairdin, 2012); (e) sapi FH (Khairdin, 2012); (f) sapi Brahman (Khairdin, 2012); dan (g) sapi PO (Khairdin, 2012).
6 Keragaman Genetik
Sumber daya genetik yang semakin beragam menurut Frankham et al.(2002) akan semakin tahan populasi tersebut untuk hidup dalam jangka waktu yang lama, serta semakin tinggi daya adaptasi populasi terhadap perubahan lingkungan. Estimasi perhitungan keragaman genetik dalam populasi secara kualitatif dapat diperoleh melalui dua ukuran keragaman variasi populasi yaitu proporsi lokus polimorfisme dalam populasi dan rata-rata proporsi individu heterozigot dalam lokus (Nei, 1987). Keragaman genetik dalam antara subpopulasi dapat diketahui dengan melihat persamaan dan perbedaan frekuensi alel di antara subpopulasi (Li et al., 2000). Falconer dan Mackay (1996) menyatakan jika suatu alel dinyatakan polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0,99.
Noor (2000) menyatakan jika frekuensi genotipe suatu populasi yang cukup besar akan selalu dalam keadaan seimbang bila tidak ada seleksi, migrasi, mutasi dan genetic drift, selain itu silang dalam dan silang luar juga dapat mempengaruhi frekuensi genotipe. Derajat heterozigositas merupakan rataan presentase lokus heterozigositas tipe individu atau rataan presentase individu heterozigot dalam populasi (Nei, 1987). Avise (1994) menyatakan jika semakin tinggi derajat heterozigositas dalam suatu populasi maka daya tahan populasi tersebut akan semakin tinggi. Javanmard et al. (2005) menyatakan jika nilai heterozigositas dibawah 0,5 (50%) mengindikasikan rendahnya variasi suatu gen dalam populasi. Machado et al. (2003) menyatakan jika nilai Ho lebih rendah dari He mengindikasikan adanya proses seleksi yang intensif.
Gen Follicle Stimulating Hormone Receptor (FSHR)
Follicle stimulating hormone (FSH) adalah hormon yang disintesa dan disekresikan oleh gonadotropes di kelenjar anterior pituitary. Fungsi utama FSH adalah menstimulasi pertumbuhan dan pematangan folikel de graaf di dalam ovarium dan spermatogenesis di dalam tubuli seminiferi testis. Proses spermatogenesis meliputi proses spermatocytogenesis (pembentukan spermatosit primer dan sekunder) dan spermiogenesis (proses pembentukan spermatozoa) (Toelihere, 1979).
Keberadaan hormon FSH sangat dipengaruhi oleh adanya gen FSH dan gen FSHR. Gen FSH terbagi atas dua subunit yaitu α-subunit dan β-subunit dengan struktur yang berbeda (Aguirre dan Timossi, 1998). Gen FSHR diperlukan untuk
7 memproduksi FSH reseptor yang membawa FSH ke target jaringan sehingga dapat berfungsi untuk spermatogenesis pada jantan dan oogenesis pada betina. FSH reseptor terekspreksi pada sel-sel granulose di ovarium dan sel-sel sertoli di testis. Gen FSHR terdapat pada kromosom 11 dan terdiri dari 10 ekson dan 9 intron dengan panjang keseluruhan ekson 2375 bp (Houde et al., 1994). Struktur pada sembilan ekson pertama mengkodekan bagian terminal-amino ekstra-selular yang besar sedangkan pada ekson kesepuluh mengkodekan bagian transmembran dan intraselular dari protein. Ekson kedua hingga kedelapan memiliki panjang yang hampir sama (68-77 bp) (Heckert and Griswold, 2002). Marson (2008) menyebutkan jika penentuan genotipe gen FSHR adalah GG (193 dan 63/50 bp), CG (243/193 dan 63/50 bp), dan CC (243 dan 63 bp). Huhtaniemi dan Kristiina (1998) menyatakan FSHR merupakan produk dari gen FSHR sangat menentukan ukuran testis, jumlah sperma, dan motilitas sperma bagi pejantan, serta keberhasilan fungsi ovarium pada betina.
Gambar 2. Struktur gen FSHR (Aguirre dan Timossi, 1998)
Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP)
Poymerase Chain Reaction (PCR) menurut Mullis (1986) merupakan suatu teknologi untuk mengamplifikasi (memperbanyak) fragmen DNA spesifik secara in-vitro. Williams (2005) menambahkan jika PCR merupakan suatu teknik yang dapat menggandakan jumlah molekul DNA pada ruas-ruas tertentu dan monomer-monomer nukleotida yang berjalan dengan bantuan primer dan enzim polymerase. Proses PCR terdiri dari tiga tahapan yaitu : (1) Denaturasi, yaitu perubahan struktur DNA utas ganda menjadi utas tunggal, (2) Anealing, yaitu penempelan primer pada sekuens DNA komplementer yang akan diperbanyak, dan (3) Ekstensi, yaitu pemanjangan primer oleh DNA polymerase (Muladno, 2002). Viljoen et al. (2005)
8 menyatakan jika proses PCR berlangsung dalam mesin thermocycler dimana pada tahap denaturasi, annealing, dan ekstension berlangsung dalam 25-30 siklus. Tahap DNA berlangsung dalam suhu 94oC sehingga DNA utai ganda dapat terpisah menjadi utai tunggal. Tahap yang paling menentukan adalah tahap penempelan primer, karena setiap pasang primer memiliki suhu penempelan primer yang spesifik. Tahap pemanjangan primer berlangsung pada suhu 27oC. Pada tahap ini enzim taq polymerase, buffer, dNTP, dan Mg2+ memulai aktifitasnya memperpanjang primer.
PCR-RFLP merupakan salah satu metode dalam PCR yang dikembangkan untuk memvisualisasikan perbedaan level DNA yang didasarkan oleh penggunaan enzim pemotong (restriction enzyme). Enzim restriksi ini dapat memotong DNA pada sekuens nukleotida spesifik (Montaldo dan Herrera, 1998). Metode PCR memanfaatkan runutan nukleotida yang dikenali oleh enzim restriksi dan disebut dengan situs restriksi. Jika situs retriksi mengalami mutasi maka enzim restriksi tidak mampu mengenalinya. Analisi RFLP biasa digunakan untuk mendeteksi ada atau tidaknya keragaman pada gen yang berhubungan dengan sifat ekonomis (Sumantri et al., 2007).
MATERI DAN METODE
Lokasi dan Waktu
Penelitian ini dilakukan di Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetik Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian dilaksanakan pada bulan Oktober 2011 hingga bulan Februari 2012.
Materi
Ternak
Sampel darah yang digunakan berasal dari tiga spesies sapi yaitu Bos javanicus (sapi Bali), Bos taurus (sapi Limousin, Simmental, Angus, dan FH), dan Bos indicus (sapi Brahman dan PO) dari berbagai lokasi. Sampel tersebut merupakan koleksi Laboratorium Genetika Molekuler Ternak dengan jumlah seperti yang tertera pada Tabel 2.
Tabel 2. Jumlah Sampel yang Digunakan dalam Penelitian
No Spesies Bangsa Asal Jumlah Sampel
1 Bos javanicus Bali (59) BPTU Bali BIBD Bali
9 50 2 Bos taurus Limousin (57)
Simmental (50) Angus (6) FH (24) BET Cipelang BIB Singosari BIB Lembang BET Cipelang BIB Singosari BIB Lembang BET Cipelang BIB Lembang BET Cipelang BIB Lembang 13 21 23 13 18 19 5 1 7 17 3 Bos indicus Brahman (9)
PO (2) BET Cipelang BIB Lembang BIB Lembang 5 4 2
Total Keseluruhan Sampel 207
Keterangan : Angka di dalam kurung menunjukkan jumlah sampel setiap bangsa sapi
Pengambilan Sampel
Bahan yang digunakan untuk pengambilan sampel adalah alkohol 70%, es, dan kapas. Alat yang digunakan adalah jarum venoject, tabung vacutainer 10 ml, dan termos.
10 Ekstraksi DNA
Bahan yang digunakan untuk ekstraksi DNA adalah sampel darah, Destilation Water (DW), NaCl, Proteinase-K, 1 x STE (5M NaCl, 2M tris HCl, 0,2M EDTA), SDS 10% (sodium dodesil sulfat), CIAA, etanol absolute, etanol 70%, phenol, buffer TE 80% (tris EDTA). Peralatan yang digunakan adalah tabung eppendorf 1,5 ml, satu set mikro pipet, tip, vortexmixer, mikrosentrifuge, inkubator, dan freezer.
Primer
Primer yang digunakan dalam penelitian untuk mengamplifikasi gen FSHR|Alu1 berdasarkan Houde et al. (1994) adalah forward : 5’-CTG CCT CCC TCA AGG TGC CCC TC-3’; dan reverse : 5’-CCC CCT AAG ACA TTT AGC CAA GAA CT -3’.
Amplifikasi Gen FSHR|Alu1
Bahan yang digunakan dalam amplifikasi metode analisa PCR-RFLP (Polymerase Chain Reaction-Restriction Fragment Lenght Polymorphism) adalah sampel DNA, destilated water (DW), buffer, MgCl2, pasangan primer fragmen Gen
FSHR|AluI, enzim Taq polymerase, dan dNTP (deoxy Nukleotida Triposfat). Alat yang digunakan adalah satu set pipet mikro, sentrifuge, mesin thermocycler, rak dan tabung eppendorf, tip pipet dan vortex.
Elektroforesis
Bahan yang digunakan adalah produk PCR, agarose, loading dye, marker 100 pb, 0,5 x TBE, dan Ethidium Bromide. Alat yang digunakan adalah tip pipet, mikropipet, gelas ukur, stirrer, microwave, gel tray, pencetak untuk sumur (comb), power supply electrophoresis 100 volt, alat foto UV trans-iluminator, dan sarung tangan.
Genotyping (Penentuan Genotipe)
Bahan yang digunakan untuk genotyping adalah produk PCR, agarose, loading dye, marker 100pb, enzim restriksi AluI, DW, buffer tango, 0,5 x TBE, dan Ethidium Bromide (EtBr). Alat yang digunakan adalah tip pipet, mikropipet 10 P Gilson, gelas kimia, gelas ukur, stirrer, microwave, gel tray, pencetak untuk sumur
11 (comb), power supply electrophoresis 100 volt, alat foto UV trans iluminator, dan sarung tangan.
Prosedur
Pengambilan Sampel
Sampel darah diambil melalui vena jugularis dengan menggunakan jarum venoject. Sampel darah ditaruh pada tabung vaccutainer yang ditambahkan etanol absolute dengan perbandingan 1:2 dan disimpan dalam lemari es hingga akan digunakan lebih lanjut.
Ekstraksi DNA
Ekstraksi DNA dilakukan dengan menggunakan metode yang dikemukakan oleh Sambrook et al. (1989). Ekstraksi DNA dimulai dengan preparasi sampel. Sampel darah sebanyak 200 µl dimasukkan ke dalam tabung 1,5 ml dan ditambahkan 1000 µl DW (destilation water). Campuran sampel darah dan DW tersebut selanjutnya dikocok atau di vortex dan didiamkan selama lima menit, setelah itu disentrifugasi dengan kecepatan 8000 rpm selama lima menit. Bagian supernatannya dibuang selanjutnya diulangi seperti proses sebelumnya. Tahap selanjutnya dari ekstraksi DNA adalah degradasi protein. Proteinase-K sebanyak 10 µl, 350 µl 1xSTE (sodium tris-EDTA) serta 40 µl 10% SDS (sodium dodesil sulfat) ditambahkan. Campuran tersebut kemudian diinkubasi pada suhu 55oC selama 2 jam sambil dikocok perlahan menggunakan alat pemutar (nutator).
Degradasi bahan organik dilakukan dengan menambahkan 40 µl 5M NaCl, 400 µl larutan fenol, dan 400 µl CIAA (chloroform iso amil alcohol), lalu dikocok pelan pada suhu ruang selama satu jam. Tahap terakhir dari ekstraksi DNA adalah presipitasi DNA yakni dengan larutan hasil degradasi bahan organik disentrifugasi dengan kecepatan 12.000 selama lima menit sehingga terbentuk fase DNA. Fase DNA tersebut selanjutnya diambil sebanyak 40 µl dan dipindahkan ke tabung baru 1,5 ml. NaCl 5M sebanyak 40 µl dan etanol absolut sebanyak 800 µl ditambahkan, kemudian dihomogenkan, dan didiamkan over night pada suhu -20oC. Molekul DNA kemudian dipisahkan daru etanol absolute dengan cara disentrifugasi pada kecepatan 12.000 rpm selama lima menit.Supernatan yang terbentuk dibuang sehingga didapatkan endapan molekul DNA. Endapan tersebut didiamkan sampai kering dan
12 disuspensikan dalam 100 µl 80% buffer TE (tris EDTA) lalu di spin down. Sampel selanjutnya dielektroforesis untuk mengetahui ada atau tidaknya DNA menggunakan agar 1,5%.
Amplifikasi DNA
Amplifikasi gen FSHR dilakukan dengan menggunakan metode PCR-RFLP. Pereaksi yang dicampurkan untuk melakukan amplifikasi terdiri dari 1 µl sampel DNA, 10,85 µl DW, 0,3 µl primer, 0,05 µl taq polymerase, 1,5 µl buffer, 0,3 µl dNTP, dan 1 µl MgCl2. Amplifikasi invitro ini menggunakan mesin thermocycler.
Kondisi suhu yang terjadi pada setiap tahapan PCR yaitu pra-denaturasi 95oC selama lima menit dalam satu siklus, 35 siklus untuk tahapan denaturasi 95oC selama 45 detik, annealing pada suhu 60oC selama 45 detik, dan elongasi pada suhu 72oC selama satu menit. Tahap selanjutnya adalah elongasi akhir 72oC selama lima menit dalam satu siklus.
Elektroforesis
Elektroforesis produk PCR dilakukan dengan terlebih dahulu membuat gel agarose 1,5% dengan cara agarose 0,45 g dilarutkan dalam larutan 0,5 x TBE sebanyak 30 ml lalu dipanaskan dalam microwave selama lima menit dan distirer dengan menambahkan 2,5 µl EtBr. Larutan yang masih cair dituangkan ke dalam pencetak gel serta sisir ditempatkan di dekat tepian gel dan dibiarkan mengeras. Sisir dicabut setelah gel mengeras sehingga terbentuk sumur-sumur. Gel selanjutnya ditempatkan ke dalam gel tray elektroforesis yang sudah berisi larutan buffer. Produk PCR sebanyak 5 µl dicampur dengan loading dye (bromothymol blue 0,01%, xylene cyanol 0,01%, dan gliserol 50%) sebanyak 1 µl dengan menggunakan mikropipet dimasukkan dalam sumur-sumur gel. Marker sebanyak 2 µl ditaruh dalam sumur paling kiri sebagai penanda. Gel tray selanjutnya dialiri listrik 100 volt selama 30 menit, molekul DNA yang bermuatan negatif pada pH netral akan bergerak (bermigrasi) ke arah positif. Setelah elektroforesis selesai, gel agarose diambil untuk dilihat panjang pita DNA dengan menggunakan sinar ultraviolet dalam mesin UV-transiluminator. Pembacaan fragmen DNA dilakukan dengan menarik garis lurus antara posisi pita dari masing-masing sampel DNA dengan posisi pita DNA marker.
13 Genotyping
Tahapan RFLP dilakukan dengan memindahkan 5 µl produk PCR kedalam tabung 0,5 ml dan ditambahkan 0,3 µl enzim restriksi AluI, buffer tango 0,7 µl, dan DW 1 µl. Campuran tersebut kemudian diinkubator pada suhu 37oC selama 16 jam.
Sampel DNA yang telah dipotong dengan enzim restriksi ditambahkan loading dye sebanyak 1 µl, dielektroforesis pada gel agarose 2% (0,6 agarose dalam 30 ml 0,5 x TBE) dengan tegangan 100 volt selama 45 menit. Setelah dielektroforesis, gel agarose diambil untuk dilihat panjang pita DNA dengan menggunakan sinar ultraviolet dan dibandingkan dengan marker untuk mengetahui panjangnya. Gambar 3 merupakan posisi penempelan primer gen FSHR dan posisi pemotongan oleh enzim AluI.
forward
1 ctgcctccct caaggtgccc ctcatcactg tgtccaagtc aaagatcctc ctggtcctgt 61 tctaccccat caactcctgt gccaacccct tcctctatgc catcttcacc aagaacttcc 121 gcagggattt cttcattctg ctgagcaagt ttggctgcta tgaagtgcaa gcccagacct 181 ataggtcaga aACCTcatcc actgcccaca actttcatcc aaggaatggc cactgccccc 241 cAG|CTcccag ggttactaat ggttccaatt acacccttat ccccctaaga catttagcca
301 agaact
reverse
Alel C : 5’---tcagaaACCTcatcc---3’ Alel G : 5’---tcagaaAGCTcatcc---3’ Keterangan : Alel C mempunyai basa C pada posisi basa ke 193
Alel G mempunyai basa G pada posisi basa ke 193
Gambar 3. Fragmen gen Follicle Stimulating Hormone Reseptor (FSHR|AluI) berdasakan sekuens gen FSHR di genbank (kode akses: L22319.1)
Analisis Data
Frekuensi Alel dan Genotipe
Frekuensi genotipe adalah rasio dari jumlah suatu genotipe terhadap suatu populasi dengan menghitung perbandingan antara jumlah genotipe tertentu pada setiap populasi. Rumus menghitung frekuensi genotipe menurut Nei dan Kumar (2000) adalah sebagai berikut :
𝒙ii = 𝒏𝑵𝒊𝒊
Keterangan :
𝑥ii = frekuensi genotipe ke-ii
nii = jumlah individu bergenotipe ii
14 Frekuensi alel adalah rasio suatu alel terhadap keseluruhan alel pada satu lokus dalam populasi. Frekuensi alel (𝑥i) gen FSHR dapat dihitung berdasarkan
rumus Nei dan Kumar (2000) seperti di bawah ini :
𝒙i =
𝟐𝒏𝒊𝒊+∑𝒏𝒊𝒋
𝟐𝑵
Keterangan :
𝑥i = frekuensi alel ke-i
nii = jumlah individu bergenotipe ii
nij = jumlah individu bergenotipe ij
N = jumlah individu sampel Heterozigositas
Keragaman genetik dapat diketahui melalui estimasi frekuensi heterozigositas pengamatan yang diperoleh dari masing-masing populasi dengan menggunakan rumus Weir (1996) sebagai berikut :
𝑯𝒐= �𝒏𝑵𝒊𝒋
𝒊≠𝒋 Keterangan :
Ho = heterozigositas pengamatan (populasi) nij = jumlah individu heterozigot
N = jumlah individu yang diamati
Heterozigositas harapan (He) berdasarkan frekuensi alel dihitung dengan menggunakan rumus Nei dan Kumar (2000) :
𝑯𝒆=𝟏 − � 𝒙𝒊𝟐 𝒒 𝒊=𝟏 Keterangan :
He = nilai heterozigositas harapan
𝑥i = frekuensi alel
HASIL DAN PEMBAHASAN
Amplifikasi Gen FSHR
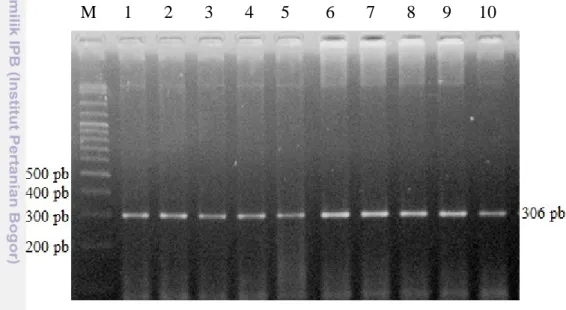
Ruas gen Follicle Stimulating Hormone Receptor (FSHR) diamplifikasi dengan metode Polymerase Chain Reaction (PCR) menggunakan primer yang dirancang oleh Houde et al. (1994). Suhu annealing yang digunakan dalam penelitian ini adalah 60oC selama 45 detik. Produk amplifikasi ruas gen FSHR adalah sebesar 306 pb dengan presentase keberhasilan amplifikasi mencapai 100% (207/207). Hasil amplifikasi gen FSHR yang divisualisasikan pada gel agarose 1,5% ditampilkan dalam Gambar 4.
M 1 2 3 4 5 6 7 8 9 10
Gambar 4. Visualisasi Amplifikasi PCR Fragmen Gen FSHR Menggunakan Gel Agarose 1,5 %. Ket : M=marker (100 pb); pb= pasang basa; 1-10= beberapa sampel sapi yang digunakan
Suhu annealing merupakan suhu yang memungkinkan terjadinya penempelan primer pada DNA yang merupakan proses paling penting dalam amplifikasi. Viljoen et al. (2005) menyatakan jika keberhasilan amplifikasi gen ditentukan oleh keberhasilan penempelan primer pada gen target. Loss et al. (2008); Marson et al. (2008); dan Hernandez et al. (2009) menggunakan suhu annealing 58 oC selama 30 detik dalam mengamplifikasi ruas gen FSHR dan menghasilkan produk PCR yang baik. Perbedaan suhu annealing dengan hasil penelitian ini dapat disebabkan adanya perbedaan komposisi mix sebagai komponen PCR. Faktor-faktor yang menentukan keberhasilan penempelan primer pada gen target yakni kondisi suhu PCR (denaturasi, annealing, dan ekstensi) serta interaksi antar komponen pereaksi PCR.
16 Rychilk (1995) menambahkan jika konsentrasi sampel DNA, konsentrasi primer, garam, dan konsentrasi pelarut dalam Premix yang digunakan juga mempengaruhi keberhasilan proses amplifikasi.
. Gen FSHR merupakan gen yang berperan dalam proses reproduksi. Produk dari gen FSHR yaitu FSHR berperan untuk membawa hormon FSH menuju sel target sehingga dapat melakukan fungsi reproduksinya (pembentukan ovum pada betina dan sperma pada jantan). Gen FSHR pada sapi terletak di kromosom 11 dan terdiri atas 10 ekson dan 9 intron (Aguirre dan Timossi, 1998). Panjang produk amplifikasi ruas gen FSHR pada sapi-sapi Bos Javanicus, Bos Taurus, dan Bos Indicus dihitung dari awal primer forward menempel hingga akhir penempelan primer reverse. Penempelan primer dalam ruas gen FSHR ini terletak pada ekson 10 dengan panjang 306 pb. Loss et al. (2008), Marson et al. (2008), dan Hernandez et al. (2009) juga menemukan panjang produk amplifikasi gen FSHR sepanjang 306 pb.
Keragaman Gen FSHR|AluI
Pendeteksian keragaman genetik gen FSHR dilakukan dengan metode RFLP. Metode ini menggunakan enzim pemotong AluI (FSHR|AluI) yang mengenali situs pemotongan AG|CT. Pemotongan ruas gen hasil amplifikasi ini menentukan genotipe dan alel masing-masing individu sapi yang berperan dalam pendeteksian keragaman genetiknya. Hasil pemotongan sekuens DNA gen FSHR pada sapi-sapi Bos taurus dan Bos indicus ditemukan dua alel (alel C dan G) dalam ruas gen FSHR, sedangkan pada Bos javanicus hanya ditemukan satu alel yaitu alel G. Alel C dalam ruas gen FSHR diakibatkan oleh adanya satu titik potong yang dikenali enzim AluI pada posisi basa ke 243 pb. Pemotongan tersebut menghasilkan dua fragmen yang panjangnya 243 pb dan 63 pb. Sedangkan alel G diakibatkan oleh adanya dua titik potong yaitu pada posisi basa ke 193 pb dan 243 pb. Pemotongan tersebut menghasilkan tiga fragmen yang panjangnya 193, 63, dan 50 pb.
Hasil PCR-RFLP ruas gen FSHR|AluI menunjukkan adanya pola pita beragam dengan tiga macam genotipe. Genotipe CC terdiri dari dua pita (243 pb dan 63 pb), genotipe CG terdiri dari empat pita (243, 193, 63, dan 50 pb), dan genotipe GG terdiri dari tiga pita (193, 63, dan 50 pb). Individu CC dan GG disebut sebagai individu homozigot karena kedua tetuanya menyumbangkan alel yang sama, sedangkan individu CG disebut sebagai individu heterozigot karena kedua tetuanya
17 menyumbangan alel yang berbeda. Hasil visualisasi produk PCR dengan pada gel agarose 2% ditampilkan pada Gambar 4. Hasil visualisasi dalam Gambar 5, pita dengan panjang 50 bp dan 63 bp tidak terlihat dalam agarose 2% karena panjang pita tersebut terlalu rendah (terjadi running out) sehingga butuh konsentrasi agarose yang lebih tinggi sehingga dapat memunculkannya. Muladno (2002) menyatakan jika dalam konsentrasi agarose 2% kisaran ukuran DNA yang dapat dianalisis berada diantara 100 hingga 2000 pb.
M 1 2 3 4 5 6 7 10 11 12 13 14
GG GG CG CG CC CC GG GG CG CG CC CC
Gambar 5. Hasil Visualisasi PCR-RFLP Gen FSHR|AluI dalam Gel Agarose 2% Perbedaan alel yang ditemukan dalam ruas gen FSHR dari individu sapi yang satu dengan lainnya disebabkan adanya perubahan basa sehingga dapat dikenali oleh enzim AluI. Mutasi yang ditemukan dalam ruas gen FSHR termasuk dalam tipe mutasi subtitusi transversi karena basa Cytosine berubah menjadi basa Guanin. Sebagaimana yang dinyatakan Li dan Graur (1991) mutasi tipe transversi terjadi akibat adanya pertukaran basa antara basa purin (A=Adenin, G=Guanin) dengan basa pirimidin (C=Cytosine, T=Timin). Mutasi tersebut menyebabkan terjadinya perubahan susunan asam amino yaitu asam amino Asparagin berubah menjadi asam amino lysin (AAC – Asparagin; AAG – Lysin). Akan tetapi dalam penelitian ini, mutasi hanya terjadi pada sapi-sapi Bos taurus dan Bos indicus saja. Pada sapi Bali (Bos javanicus) meskipun alel yang ditemukan dalam ruas gen FSHR ini hanya alel G saja namun hal tersebut bukan disebabkan oleh mutasi melainkan karena adanya perbedaan nukleotida pada sapi Bali dengan sapi Bos taurus dan Bos indicus. Hasil pemotongan tersebut dapat digunakan sebagai penciri genetik gen FSHR|AluI untuk sapi Bali (Bos javanicus).
18 Genotipe yang ditemukan pada sapi Bos taurus dan sapi Bos indicus adalah CC, CG, dan GG, sedangkan pada sapi Bos javanicus (sapi Bali) hanya ditemukan satu genotipe saja yaitu genotipe GG. Sapi limousin, simmental, FH, yang merupakan sapi Bos taurus muncul ketiga genotipe tersebut, akan tetapi pada sapi Angus hanya muncul dua genotipe saja yaitu CC dan CG. Jumlah sampel sapi angus yang digunakan dalam penelitian ini hanya enam sampel. Jumlah tersebut harus diperbanyak agar mendapatkan hasil yang lebih akurat. Sapi Bos indicus yaitu sapi Brahman yang digunakan dalam penelitian ini muncul ketiga genotipe CC, CG, dan GG, akan tetapi pada sapi PO hanya menunjukkan dua genotipe yakni CC dan GG.
Frekuensi Genotipe dan Frekuensi Alel Gen FSHR
Frekuensi genotipe adalah rasio dari jumlah suatu genotipe dengan menghitung perbandingan antara jumlah genotipe tertentu. Sedangkan frekuensi alel adalah rasio suatu alel terhadap keseluruhan alel pada satu lokus dalam populasi. Frekuensi genotipe dan frekuensi alel dari penelitian yang dilakukan ditampilkan pada Tabel 3.
Tabel 3. Frekuensi Genotipe dan Frekuensi Alel dari Gen FSHR|AluI
Spesies Bangsa n Genotipe Alel
CC CG GG C G
Bos javanicus Sapi Bali 59 0,00 (0) 0,00 (0) 1,00 (59) 0,00 1,00 Bos taurus Limousin
Simmental Angus FH 57 50 6 24 0,63 (36) 0,54 (27) 0,83 (5) 0,42 (10) 0,32 (18) 0,28 (14) 0,17 (1) 0,46 (11) 0,05 (3) 0,18 (9) 0,00 (0) 0,12 (3) 0,79 0,68 0,92 0,65 0,21 0,32 0,08 0,35 Total 137 0,57 0,32 0,11 0,73 0,27 Bos indicus Brahman
PO 9 2 0,12 (1) 0,50 (1) 0,44 (4) 0,00 (0) 0,44 (4) 0,50 (1) 0,33 0,5 0,67 0,5 Total 11 0,18 0,36 0,46 0,36 0,64 Keterangan : angka di dalam kurung menunjukkan jumlah sampel
Hasil analisis frekuensi genotipe dan frekuensi alel dalam Tabel 3 menunjukkan jika pada sapi Limousin, Simmental, dan Angus memiliki frekuensi genotipe CC yang lebih besar dibandingkan dua genotipe lainya dengan nilai frekuensi genotipe CC masing-masing sebesar 0,63, 0,54, dan 0.83. Sapi Limousin
19 dan sapi Simmental memiliki nilai frekuensi genotipe GG yang paling kecil yaitu 0,05 dan 0,18. Pada sapi Angus hanya ditemukan genotipe CC dan CG saja dengan nilai frekuensi genotipe CG sebesar 0,17. Sapi FH memiliki frekuensi genotipe CG yang lebih besar dibandingkan kedua genotipe lainnya dengan nilai 0,46. Sapi Brahman memiliki frekuensi genotipe CC yang paling rendah yaitu 0,12 sedangkan genotipe CG dan GG memiliki nilai frekuensi genotipe yang sama yaitu 0,44. Sapi PO yang diamati dalam penelitian hanya menunjukkan dua genotipe saja yaitu CC dan GG dengan nilai yang sama masing-masing yaitu 0,50. Sapi Bali yang merupakan sapi lokal Indonesia hanya menunjukkan satu buah genotipe saja dari keseluruhan sampel yaitu genotipe GG. Hal ini menyebabkan frekuensi genotipe GG pada sapi Bali sangat tinggi yaitu 1,00. Gambar 6 menunjukkan grafik frekuensi genotipe pada spesies sapi Bos javanicus, Bos taurus, dan Bos indicus.
Gambar 6. Frekuensi Genotipe FSHR|AluI di Berbagai Spesies Sapi (dalam Persen) Gambar 6 menunjukkan dengan jelas perbedaan antara spesies sapi Bos taurus dengan spesies sapi Bos indicus. Spesies sapi Bos taurus memiliki genotipe CC yang terbesar, sedangkan pada spesies sapi Bos indicus genotipe yang terbesar adalah genotipe GG. Genotipe GG yang merupakan genotipe satu-satunya yang
0 10 20 30 40 50 60 70 80 90 100
Bos javanicus Bos taurus Bos indicus
0 57 18 0 32 36 100 11 46 F re kue ns i G eno ti pe ( p ers en ) Spesies Sapi CC CG GG
20 muncul pada sapi Bali (Bos javanicus) menjadikan frekuensi genotipe GG pada spesies sapi Bos javanicus adalah sebesar 1,00, sedangkan frekuensi genotipe CC dan CG adalah 0,00. Sapi Bos taurus memiliki frekuensi genotipe CC yang lebih besar dari dua genotipe lainnya yaitu sebesar 0,57. Genotipe GG merupakan genotipe dengan frekuensi yang paling rendah dari spesies sapi Bos taurus yaitu sebesar 0,11. Hasil ini berbanding terbalik dengan spesies sapi Bos indicus dengan frekuensi genotipe GG paling besar yaitu 0,46 dan frekuensi genotipe CC paling rendah yaitu 0,18. Frekuensi genotipe CG pada sapi Bos taurus dan Bos indicus secara berurutan adalah 0,32 dan 0,36.
Tipe alel yang ditemukan dalam penelitian ini adalah alel C dan alel G. Frekuensi alel C pada sapi Simmental, Limousin, Angus, dan FH lebih tinggi dibandingkan frekuensi alel G. Nilai frekuensi alel C dan G pada sapi Simmental, Limousin, Angus, dan FH berturut-turut adalah 0,680 dan 0,320, 0,789 dan 0,211, 0,917 dan 0,083, serta 0,646 dan 0,354. Pada sapi Bali dan Brahman nilai frekuensi alel G lebih besar dibandingkan frekuensi alel C yaitu sebesar 1,000 dan 0,000 pada sapi Bali serta 0,667 dan 0,333 pada sapi Brahman. Sapi PO memiliki nilai frekuensi alel C dan G yang sama yaitu 0,500. Hal ini disebabkan hanya ditemukan 2 genotipe saja pada sapi PO yaitu CC dan GG.
Frekuensi alel C pada spesies Bos taurus lebih besar dibandingkan dengan frekuensi alel G. Frekuensi alel C Bos taurus 0,73 sedangkan frekuensi alel G sebesar 0,27. Hasil tersebut sesuai dengan penelitian Ishak (2012) yang menunjukkan frekuensi alel C lebih besar dibandingkan frekuensi alel G pada sapi Bos taurus. Hasil tersebut berbanding terbalik dengan spesies Bos javanicus dan Bos indicus. Kedua spesies ini menujukkan jika frekuensi alel G lebih besar dibandingkan frekuensi alel C. Frekuensi alel G pada spesies Bos javanicus adalah 1,00 dan pada spesies Bos indicus adalah 0,64. Penelitian yang dilakukan Ishak (2012) menunjukkan jika sapi dari spesies Bos javanicus (sapi Bali) memiliki nilai frekuensi alel G 0,951 dan alel C 0,049. Terdapatnya alel G dalam hasil tersebut dikarenakan sapi Bali yang diteliti oleh Ishak (2012) berasal dari NTT, NTB dan Sulawesi yang diduga telah mengalami pencemaran. Penelitian yang dilakukan oleh Loss et al. (2008) pada sapi spesies Bos indicus (sapi Nellore) memiliki hasil yang sama
21 dengan penelitian yaitu memiliki frekuensi alel G yang besar dibandingkan dengan alel C.
Nilai frekuensi alel dalam penelitian ini pada spesies sapi Bos taurus dan Bos indicus bersifat polimorfik. Nilai frekuensi alel pada spesies sapi Bos javanicus hanya ditemukan genotipe GG dan hanya memiliki satu alel yaitu alel G dengan nilai 1,000. Oleh karena itu Gen FSHR yang ditemukan pada sapi Bos javanicus bersifat monomorfik atau seragam. Hal ini sesuai dengan pendapat Nei (1987) yang menyatakan bahwa suatu alel dikatakan polimorfik atau beragam jika memiliki frekuensi alel sama dengan atau kurang dari 0,99, akan tetapi jika terjadi lebih dari 0,99 dikatakan monomorfik atau seragam. Keragaman yang bersifat monomorfik pada sapi Bos javanicus menunjukkan jika adanya manajemen perkawinan yang tidak acak terlebih sampel sapi Bali yang digunakan bersasal dari daera Bali saja, selain itu adanya seleksi dalam sifat tertentu, serta tingkat silang dalam yang tinggi.
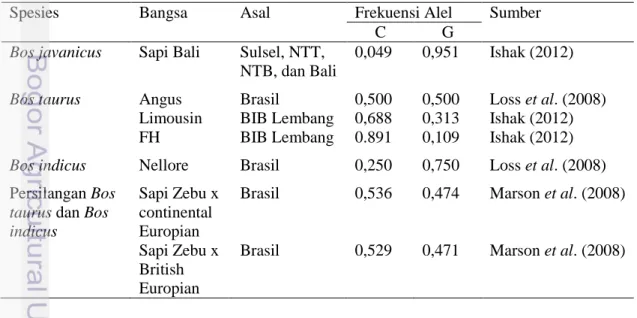
Tabel 4 menunjukkan beberapa pembanding hasil frekuensi alel gen FSHR|AluI dari beberapa literatur yang juga menggunakan metode PCR-RFLP. Pada tabel tersebut terlihat jika Marson et al (2008) mengidentifikasi keragaman gen FSHR menggunakan sapi persilangan spesies Bos taurus dan Bos indicus. Hasil dari penelitian tersebut adalah nilai frekuensi alel C lebih tinggi dibandingkan alel G. Antara nilai frekuensi alel C dan alel G juga tidak berbeda jauh. Hasil penentuan genotipe sapi persilangan ini memiliki frekuensi genotipe CG yang lebih tinggi dibandingkan genotipe CC dan GG.
Tabel 4.Frekuensi alel gen FSHR|AluI pada Beberapa sumber
Spesies Bangsa Asal Frekuensi Alel Sumber
C G
Bos javanicus Sapi Bali Sulsel, NTT, NTB, dan Bali 0,049 0,951 Ishak (2012) Bos taurus Bos indicus Angus Limousin FH Nellore Brasil BIB Lembang BIB Lembang Brasil 0,500 0,688 0.891 0,250 0,500 0,313 0,109 0,750 Loss et al. (2008) Ishak (2012) Ishak (2012) Loss et al. (2008) Persilangan Bos
taurus dan Bos indicus Sapi Zebu x continental Europian Sapi Zebu x British Europian Brasil Brasil 0,536 0,529 0,474 0,471 Marson etal. (2008) Marson etal. (2008)
22 Gen FSHR berperan dalam fungsi reproduksi sangat menentukan kualitas sperma sapi pada sapi jantan. Huhtaniemi dan Kristiina (1998) menyatakan jika FSHR yang merupakan produk dari gen FSHR sangat menentukan ukuran testis, jumlah sperma, dan motilitas sperma bagi pejantan, serta keberhasilan fungsi ovarium pada betina. Ishak (2012) dalam penelitianya menunjukkan jika sapi Bos taurus dan Bos indicus dengan genotipe GG memiliki tingkat keabnormalitas yang lebih tinggi dibandingkan dua genotipe lainnya. Ishak (2012) juga menyatakan bangsa sapi Bali (Bos javanicus) memiliki kualitas sperma yang paling baik dibandingkan dengan bangsa sapi dari dua spesies lainnya. Arifianti et al. (2010) menyatakan bahwa presentase abnormanilitas sapi Bali (Bos javanicus) sebesar 6% dengan 2% kecatatan pada kepala. Kualitas sperma sapi tidak hanya dipengaruhi oleh faktor genetik saja. Faktor lingkungan juga memiliki pengaruh besar pada kualitas sperma sapi seperti kandungan nutrisi pakan, manajemen pemeliharaan, dan cuaca lingkungan.
Heterozigositas
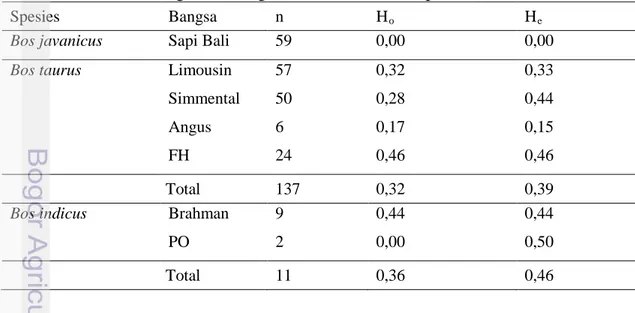
Nilai heterozigositas dihitung untuk mengetahui keragaman genetik gen FSHR|AluI pada suatu populasi. Analsisis hetorozigositas pada berbagai sapi ditampilkan pada Tabel 5.
Tabel 5. Nilai Heterozigositas Pengamatan (Ho) dan Harapan (He) Gen FSHR
Spesies Bangsa n Ho He
Bos javanicus Sapi Bali 59 0,00 0,00 Bos taurus Limousin
Simmental Angus FH 57 50 6 24 0,32 0,28 0,17 0,46 0,33 0,44 0,15 0,46 Total 137 0,32 0,39
Bos indicus Brahman PO 9 2 0,44 0,00 0,44 0,50 Total 11 0,36 0,46
Pendugaan nilai heterozigositas memiliki arti penting untuk diketahui karena dapat memberikan informasi mengenai tingkat polimorfisme suatu alel, serta prospek populasi di masa yang akan datang (Falconer dan Mackay, 1996). Marson et al.
23 (2005) menambahkan jika pendugaan nilai heterozigositas diperoleh untuk mendapatkan kergaman genetik dalam populasi sehingga dapat dimanfaatkan untuk program seleksi dan menjadi sumber genetik pada generasi berikutnya. Pendugaan nilai heterozigositas pada penelitian ini berdasarkan nilai heterozigositas pengamatan (Ho) dan nilai heterozigositas harapan (He). Heterosigositas harapan (He) merupakan
penduga keragaman genetik yang tepat pada populasi hewan ternak karena perhitungannya langsung berdasarkan pada frekuensi alel (Moioli et al., 2004).
Heterozigositas pengamatan (Ho) dalam penelitian ini paling besar adalah
pada spesies sapi Bos indicus yaitu 0,36 dan yang paling terendah adalah spesies Bos javanicus yaitu 0,00. Pada spesies sapi Bos taurus, sapi FH memiliki nilai Heterozigositas pengamatan (Ho) yang paling tinggi yaitu 0,46 dan pada spesies sapi
Bos indicus sapi Brahman memiliki nilai yang paling tinggi yaitu 0,44.
Javanmard et al. (2005) menyatakan jika nilai heterozigositas dibawah 0,5 (50%) mengindikasikan rendahnya variasi suatu gen dalam populasi.Hal demikian menunjukkan jika rendahnya keragaman gen FSHR|AluI pada sapi-sapi spesies Bos javanicus, Bos taurus, dan Bos indicus. Nilai heterozigositas dipengaruhi oleh banyaknya sampel, jumlah alel, dan frekuensi alel dalam suatu populasi (Nei, 1987). Jumlah sampel pada spesies Bos indicus yang sangat rendah (11 sampel) mengurangi keakuratan nilai heterozigositas.
Nilai heterozigositas harapan (He) pada spesies sapi Bos javanicus dan Bos
taurus yang diamati tidak menunjukkan perbedaan yang besar dengan nilai heterozigositas pengamatannya (Ho). Akan tetapi, pada spesies sapi Bos indicus
memiliki perbedaan yang cukup tinggi. Tambasco et al. (2003) menyatakan jika terjadi pebedaan yang besar antara heterozigositas harapan (He) dan heterozigositas
pengamatannya (Ho) maka terjadi ketidakseimbangan genotipe dalam populasi yang
diamati. Hal yang serupa juga terjadi pada nilai heterozigositas sapi Simmental dari spesies Bos tarus. Nilai Ho pada keseluruhan spesies sapi memiliki nilai yang lebih
rendah dari nilai He. Machado et al. (2003) menyatakan jika nilai Ho lebih rendah
dari He mengindikasikan adanya proses seleksi yang intensif. Berdasarkan hal
tersebut maka deiperkirakan telah terjadi proses seleksi intensif berdasarkan gen FSHR|AluI.
KESIMPULAN DAN SARAN
Kesimpulan
Gen FSHR|AluI pada spesies sapi Bos taurus dan Bos indicus bersifat polimorfik dengan ditemukannya dua alel (alel C dan G) dan tiga genotipe (genotipe CC, CG, dan GG), sedangkan pada spesies Bos javanicus (sapi Bali) bersifat monomorfik dengan ditemukan satu alel (G) dan satu genotipe (GG). Derajat heterozigositas pada spesies Bos taurus (Limousin, Simmental, Angus, dan FH) dan Bos indicus (Brahman dan Peranakan Ongole) memiliki nilai heterozigositas tinggi dibandingkan dengan sapi yang termasuk dalam spesies Bos javanicus (sapi Bali).
Saran
Penelitian terhadap gen FSHR|AluI perlu dilakukan dengan jumlah sampel yang banyak terutama pada sampel sapi yang termasuk spesies Bos indicus. Selain itu, perlu dicari situs lain yang dianggap polimorfik dalam gen FSHR.
UCAPAN TERIMA KASIH
Bismillahirrahmanirrahim,
Puji dan syukur penulis panjatkan kehadirat Allah SWT yang telah memberikan Taufik dan Rahmat-Nya sehingga penulis dapat menyelesaikan penulisan skrispi ini. Shalawat dan salam senantiasa dicurahkan kepada suri tauladan seluruh umat manusia, Baginda Rasulullah Muhammad SAW beserta sanak saudara, keluarga, serta para sahabat dan seluruh umat pengikutnya. Penulis menyampaikan terimakasih yang sebesar-besarnya kepada kedua orang tua tercinta, Imam Sudarmo dan Sri Indrayati Faidah atas segala kasih sayang, doa, motivasi serta bimbingannya untuk penulis. Terima kasih kepada adik Dimas Yukafie Hersurastopo serta keluarga besar dari Mama dan Bapak yang senantiasa menyemangati dan memberikan doa kepada penulis.
Penulis mengucapkan banyak terima kasih kepada Bapak Prof. Dr. Ir. Cece Sumantri, M. Agr. Sc. selaku dosen pembimbing akademik dan dosen pembimbing utama serta Bapak Dr. Jakaria, S.Pt.,M.Si. selaku pembimbing anggota atas arahan, motivasi, serta ilmu yang diberikan kepada penulis. Penulis juga mengucapkan terima kasih kepada Bapak Prof. Dr. Ir. Muladno, MSA dan Bapak Ir. Kukuh Budi Satoto, MS atas masukan dan saran pada saat ujian akhir penulis.
Ucapan terima kasih penulis sampaikan kepada Kakak Eryk, Kakak Ferdy, Kakak Irene, Bapak Andi, dan David yang telah banyak membantu penulis dalam menyelesaikan penelitian. Ucapan terimakasih juga penulis sampaikan kepada sahabat-sahabat Laboratorium Genetika Molekuler Ternak (Cicha, Furqon, Icha, Eka, Rey, Tessa, Adil, Sri, dan Menix), Ariandanu Catur Biandana, sahabat-sahabat tersayang (Tia, Rissa, Ii, Irma, Feby, Rina, dan Vita), dan teman-teman IPTP 45 atas doa, semangat, canda, dan kebersamaannya. Terakhir penulis ucapkan terima kasih kepada semua pihak yang telah membantu penulis yang tidak bisa disebutkan satu persatu. Terima kasih atas segala doa dan motivasi yang telah diberikan sehingga penulis dapat menyelesaikan studi dan tugas akhir ini dengan sebaik-baiknya.
Bogor, Juli 2012
DAFTAR PUSTAKA
Aguirre, U.A., & C. Timossi. 1998. Structure-function relationship of follicle stimulating hormone and its receptor. J. Hum. Rep. 4:260-283.
Arifiantini, R.I, T. Wresdiyati, & E.F. Retnani. 2006. Pengujian morfologi spermatozoa sapi Bali (Bos sondaicus) menggunakan pewarnaan William. J. Indones. Trop. Anim. Agric. 2:105-110.
Avise, J.C. 1994. Molecular Markers. Natural History Evolution. Chapman and Hall, Inc., Washington.
Balai Pembibitan Ternak Potong Nusa Tenggara Timur. 2009. Sapi Potong. http://ntt.litbang.deptan.go.id. [14 Oktober 2011].
Departemen Pertanian. 2006. Peraturan Menteri Pertanian Nomor 35/Permentan/ OT.140/8/2006. Pedoman Pelestarian dan Pemanfaatna Sumberdaya Genetik Ternak. Deptan, Jakarta.
Falconer, D.S., & T.F.C. Mackay. 1996. Introduction to Quantitative Genetics. 4th Ed. Longman, New York.
Frankham, R., J. D. Ballou, & D.A. Briscoe. 2002. Introduction to Conversation Genetics, Cambridge University Press.
Heckert, L.L.,& M.D. Griswold. 2002. The expression of the follicle stimulting hormone receptor in spermatogenesis. J. Endoc. Soc. 129-148.
Hernandez, B.C., P. Cevrantes., F. Montiel., R. Canseco., & A. Carrasco. 2009. Allelic variants of fshr gene in cow of different genotypes in mexico. J. Anim. Vet. Adv. 8:2489-2494.
Houde, A., Lambert A, Saumade J, & Silversides D.W. 1994. Structure of the bovine follicle stimulating hormone receptor complementary dna and expression in bovine tissues. Mol. Rep. Dev. 39: 127-135.
Huhtaniemi, I.T & A. Kristiina. 1998. Mutations of follicle-stimulating hormone and its receptor;effects on gonadal function. J. Europ. Endoc. 138: 473–481. Ishak, A.B.L.2012 Identifikasi keragaman gen FSH sub unit beta, gen FSH reseptor
dan gen GH pada sapi Bali jantan sebagai penanda kualitas sperma. Disertasi, Sekolah Pascasarjana. Institut Pertanian Bogor. Bogor.
Jakaria. 2008. Keragaman genetik gen hormon pertumbuhan pada sapi Pesisir Sumatra barat. Disertasi. Sekolah Pascasarjana. Institut Pertanian Bogor. Bogor.
Javanmard, A., N. Asadazadeh., M. H. Banahazi., & J. Tavakolian. 2005. The alleleand genotype frequencies of bovine pituitary specific transcription factor and leptin genes in Iranian cattle and buffalo populations using PCR-RFLP. J. Iranian. Biotech. 3: 104-108.
Khairdin, S. 2012. Jenis-jenis sapi potong di Indonesia. http://epetani.deptan.go.id. [9 Juni 2012]
Kusdiandoro, M., M. Olsson, G. Andersson, B. Purwantara, T.H.T.A. Van Tol, S. Mikko, H.R. Martinez, B. Colenbrader & J.A. Lenstra. 2007. Genetic
27 diversity and conservation of south-east asian cattle:from Indian Zebu to Indonesian Banteng, and then to the Cambodian Kouprey. Symposium. Kasetsart University. Bangkok, Thailand. 120-124.
Li, W.& D. Graur. 1991. Fundamentals of molecular evolution. Sinauer Associates Inc. Publisher. Sunderland, Massachusetts.
Li, X., K.Li, B.Fan, Y. Gong, S. Zhao, Z. Peng & B. Liu. 2000. The genetic diversity of seven pigs breeds in china, estimated by means of microsatellites. J. Anim. Sci 9:1193-1195.
Loss, P.R.A., J.C. Silveira, W.G. Glanzer, J.C.F. Moraes, & T.A. Weimer. 2008. Diversidade genetic de doadoras de embrioes das racas Nelore e Aberdeen angus.Veterinaria em foco. 5: 85-92.
Machado, M. A., I. Schuster, M. L. Martinez, & A. L. Campos. 2003. Genetic diversity of four breed using microsatellite markers. Rev. Bras. De Zool. 32: 93-98.
Machugh, D.E.1996. Molecular biogeoghrapy and genetic structure of domestic cattle. Thesis. University of Dubin. Irlandia.
Marson, E.P., J.B.S. Ferraz, F.V. Meirelles, J.C.C. Balieireo, & J.P. Eler. 2008. Effects of polymorphism of LHR and FSHR genes on sexual precocity in a Bos Taurus X Bos Indicus beef composite population. Genet. Mol. Res. 7: 243-25.
Martodjo, H. 1990. Perkembangan Sapi Bali. Proc. Seminar Nasional Sapi Bali; Denpasar 20-22 September Fakultas Peternakan Universitas Udayana, Bali. Moioli, B., F. Napolitano, & G. Catillo. 2004. Genetic diversity between
Piedmontese, Maremmana, and Podolica cattle breeds. J. Hered. 95:250-256. Montaldo, H.H., & C.A.M. Herrera. 1998. use of molecular markers and major genes
in genetic improvement of livestock. J.Biotech. 1:2
Muladno. 2002. Seputar Teknologi Rekayasa Genetika. Pustaka Wirausaha Muda dan USESE Foundation, Bogor.
Mullis, K. 1986. Specific enzymatic amplification of DNA in-vitro: the polymerase chain reaction. Cold Spring Harb Symp Quant Biol 51:263-273.
Nei, M., & S. Kumar. 2000. Molecular Evaluation and Phylogenetics. Oxford University Press, New York.
Nei, M. 1987. Molecular Evoluntionery Genetics. Columbia University Press, New York.
Noor, R.R. 2000. Genetika Ternak. Penebar Swadaya. Depok. Pane, I. 1986. Pemuliabiakan Ternak Sapi. Gramedia, Jakarta.
Payne W.J. & D.H.L Rollinson. 1973. Bali cattle. World. Anim. Rev. 7:13-21.
Rachmad, B. 2001. Penampilan produksi sapi peranakan Ongole (PO) yang dipelihara dalam feedlot dengan pemanfaatan pucuk tebu pada lama pemeliharaan yang berbeda. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.
28 Rahmatullah, S.N. 2011. Identifikasi keragaman gen hormon pertumbuhan di daerah
exon pada sapi lokal Indonesia berdasarkan metode PCR-SSCP. Thesis. Sekolah Pascasarjana. Institut Pertanian Bogor. Bogor.
Rollinson, D.H.L. 1984. Bali cattle. in Mason, IL. (Ed.): Evolution of Domesticated Animals. NewYork. Longman.
Rychilk, W.1995. Priming efficiency in PCR. J.Biotech. 18:84-86.
Sambrook, J., E.F. Fritsch, & J.F. Medrano. 1989. Molecular Cloning : A Laboratory Manual. Second Edition. Cold Spring Harbor Laboratory Press, New York. Saputra, E.D. 2008. Sapi bali sebagai plasma nutfah Indonesia dan peranannya bagi
petani. http://balivetman.wordpress.com/. [14 Oktober 2011].
Sarbaini. 2004. Kajian keragaman karakteristik eksternal dan DNA mikrosatelit sapi Pesisir Sumatera Barat. Disertasi. Sekolah Pascasarjana IPB. Bogor.
Sumantri, C., A. Anggraeni, A. Farajallah, & D. Perwitasari. 2007. Keragaman mikrosatelit DNA sapi perah FH di Balai Pembibitan Ternak Unggul Baturaden. JITV 12:124-133.
Talib, C. 2002. Sapi bali di daerah sumber bibit dan peluang pengembangannya. Wartazoa 12:3-
Tambasco, D.D. et al. 2003. Candidate genes for growth traits in beef cattle crosses Bos taurus x Bos indicus. J. Anim. Breed. Genet. 120:51.
Toelihere, M.R. 1979. Fisiologi Reproduksi pada Ternak. Angkasa, Bandung.
Utoyo, D.P. 2002. Management of the Farm Domestic Animal Genetic Resource in Indonesia. Directorate Generale of Livestock Services. Ministry of Agriculture Indonesia. Jakarta.
Viljoen, G.J., L.H. Nel, & J.R. Crowther. 2005. Molecular Diagnostic PCR Handbook. Springer, Dordrecht, Netherland.
Weir, B.S. 1996. Genetic Data Analysis: Method for Discrete Population Genetic Data. Second ed. Sinauer Associates Sunderland, MA USA.
Williams, J.L. 2005 the use of marker-assisted selection in animal breeding and biotechnology. Rev. Sci.tech. Off. Int. Epiz., 24:379-391.
Williamson, G. Dan W.J. Payne. 1993. Pengantar Peternakan di Daerah Tropis. Edisi Ketiga (Terjemahan). Gajah Mada University Press, Yogyakarta.
30 Lampiran 1. Sekuens Gen FSHR (diakses dari GenBank No L22319)
LOCUS BOVFSHR 2375 bp mRNA linear MAM 13-OCT-1994 DEFINITION Bovine follicle stimulating hormone receptor (FSHR) mRNA, complete cds.
ACCESSION L22319
VERSION L22319.1 GI:404671
KEYWORDS follicle stimulating hormone (FSH) receptor. SOURCE Bos taurus (cattle)
ORGANISM Bos taurus
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi;
Mammalia; Eutheria; Laurasiatheria; Cetartiodactyla; Ruminantia; Pecora; Bovidae; Bovinae; Bos.
REFERENCE 1 (bases 1 to 2375)
AUTHORS Houde,A., Lambert,A., Saumande,J., Silversides,D.W. and Lussier,J.G.
TITLE Structure of the bovine follicle-stimulating hormone receptor complementary DNA and expression in bovine tissues
JOURNAL Mol. Reprod. Dev. 39 (2), 127-135 (1994) PUBMED 7826612
REFERENCE 2 (bases 1 to 2375) AUTHORS Lussier,J.G.
TITLE Direct Submission
JOURNAL Submitted (30-SEP-1993) Jacques G. Lussier, CRRA, Faculty of Veterinary Medicine, University of Montreal, P.O. Box 5,000, 3,200 Sicotte, St-Hyacinthe, Quebec, J2S 7C6, Canada
COMMENT Original source text: Bos taurus cDNA to mRNA. FEATURES Location/Qualifiers source 1..2375 /organism="Bos taurus" /mol_type="mRNA" /strain="Holstein" /db_xref="taxon:9913" /tissue_type="ovaries/testes" gene 1..2375 /gene="FSHR" 5'UTR 1..64 /gene="FSHR" CDS 65..2152 /gene="FSHR" /codon_start=1
/product="follicle stimulating hormone receptor" /protein_id="AAC37324.1"
31 /db_xref="GI:404672" /translation="MALLLVALLAFLSLGSGCHHRLCHCSNGVFLCQESKVTEIPSDL PRDAVELRFVLTKLRVIPKGAFSGFGDLEKIEISQNDVLEVIEANVFSNLPKLHEIRI EKANNLLYIDPDAFQNLPNLRYLLISNTGIKHLPAVHKIQSLQKVLLDIQDNINIHTV ERNSFMGLSFESMTVWLSKNGIQEIHNCAFNGTQLDELNLSDNSNLEELPNDVFQGAS GPVILDISRTRIRSLPSYGLENLKKLRAKSTYRLKKLPSLEKFVTLVEASLTYPSHCC AFANWRRQTSDLHPICNKSILRQEVDDMTQARGQRVSLAEDDEPSYAKGFDVMYSEFD YDLCNEVVDVTCSPEPDAFNPCEDIMGDDILRVLIWFISILAITGNILVLVILITSQY KLTVPRFLMCNLAFADLCIGIYLLLIASVDVHTKTEYHNYAIDWQTGAGCDAAGFFTV FASELSVYTLTAITLERWHTITHAMQLECKVQLRHAASIMLVGWIFAFAVALFPIFGI SSYMKVSICLPMDIDSPLSQLYVMSLLVLNVLAFVVICGCYTHIYLTVRNPNITSSSS DTKIAKRMAMLIFTDFLCMAPISFFAISASLKVPLITVSKSKILLVLFYPINSCANPF LYAIFTKNFRRDFFILLSKFGCYEVQAQTYRSETSSTAHNFHPRNGHCPPAPRVTNGS NYTLIPLRHLAKN" sig_peptide 65..118 /gene="FSHR" mat_peptide 119..2149 /gene="FSHR"
/product="follicle stimulating hormone receptor" 3'UTR 2150..2375
/gene="FSHR" ORIGIN
1 tctgaaatct gtgcaggatt ttctctgcag aggcagaaga aagcaggtgg atggataagt 61 aaacatggcc ttgctcctgg tggccttgct cgcattcctg agcctgggct caggatgcca 121 tcatcgactc tgtcactgct ctaacggagt tttcctctgc caggagagca aggtgacaga 181 gattccctcc gacctcccca gggatgcagt cgaactgagg tttgttctca ccaagcttcg 241 agtcatccca aaaggagcct tttcaggatt tggagacctg gagaaaatag agatctcaca 301 gaatgatgtc ttggaagtga tagaggcaaa cgtgttctcc aacctgccca aactacatga 361 aattagaatt gaaaaggcca acaacctgct atacatcgac cctgatgcct tccagaacct 421 tcccaacctc cgatatctgt taatatccaa cacaggtatt aagcacttgc cagctgttca 481 caagattcag tctctccaaa aggttttact agatattcaa gataatataa acatccacac 541 agttgaaaga aattctttca tggggctaag ttttgaaagt atgactgtat ggctgagtaa 601 gaatgggatt caagaaatac acaactgtgc attcaatgga acccaactcg atgagctgaa 661 tctaagtgat aacagtaatt tggaagaact gcctaatgat gttttccagg gagcctctgg 721 accggtcatt ctagatattt caagaaccag gatccgttcc ctgcctagct atggcttaga 781 aaatcttaag aagctgcggg ccaagtcaac ttaccgctta aaaaagcttc ccagtctgga 841 aaaatttgtc acactcgtgg aggccagcct cacctacccc agccactgct gtgcctttgc 901 aaactggagg cggcaaacct ctgaccttca tccaatttgc aacaaatcta ttttaaggca 961 agaagttgat gacatgactc aggctagggg tcagagagtc tctttggcag aagatgatga 1021 gcccagctat gccaaaggat ttgacgtgat gtacagtgaa tttgactatg acttatgcaa 1081 tgaagtggtt gatgtgactt gctcccctga gccagacgca tttaatccat gtgaagatat 1141 catgggggat gatattctca gagtcttgat atggtttatt agcatcctgg ccatcactgg
32 1201 gaacatccta gtgctggtga tcctgatcac cagccagtat aaactcacag tcccccggtt 1261 cctcatgtgc aatctggcct ttgccgatct ctgcattgga atctacctgc tgctcatagc 1321 ctcagttgat gtccacacca aaacggagta ccacaactat gccatcgact ggcaaactgg 1381 agcaggctgt gatgctgccg gctttttcac tgtctttgcc agtgagctct cagtctacac 1441 tctgaccgcc atcacgctgg aaagatggca taccatcacc catgccatgc agctcgaatg 1501 caaagtgcag ctccgccatg ctgccagcat catgctggtg ggctggatct ttgcttttgc 1561 agttgccctt tttcccatct ttggcatcag cagctacatg aaggtgagca tctgcctgcc 1621 catggacatt gacagcccct tgtcacaact ctatgtcatg tccctccttg tgctcaatgt 1681 cctggccttt gtggtcatct gtggctgcta cactcacatc tacctcacgg tgaggaaccc 1741 caacatcaca tcctcttcta gtgacaccaa gatagccaag cgcatggcca tgctcatctt 1801 caccgacttc ctctgcatgg cacccatctc cttctttgcc atctctgcct ccctcaaggt 1861 gcccctcatc actgtgtcca agtcaaagat cctcctggtc ctgttctacc ccatcaactc 1921 ctgtgccaac cccttcctct atgccatctt caccaagaac ttccgcaggg atttcttcat 1981 tctgctgagc aagtttggct gctatgaagt gcaagcccag acctataggt cagaaacctc 2041 atccactgcc cacaactttc atccaaggaa cggccactgc cccccagctc ccagggttac 2101 taatggttcc aattacacac ttatccccct aagacattta gccaagaact aaaacacaat 2161 gtacaaatgt ttctgagtgt tgaatgacag ataagtcttg cctttgaaga atatgccatg 2221 gaagagctga cagagcactt ccacttactt catctaattt aatctccctg gcacatccat 2281 aaggtaaatt ggtcagaaac tattaacttc acatgattca ttaggaaact gaagacttca 2341 taacaacatt aataattaaa ataatgtagt actac