ABSTRACT
One alternative to increase national productivity of rice is using marginal soil (acid soil). However, the soil has high acidity, low mineral solubility (N, P, K, Ca, Mg, and Mo), and high Al and Mn toxicity. Finding the variety tolerant to envi-ronmental stresses could solve the problem. The experiment was conducted to investigate callus induction methods of rice and their regeneration and appropriate in vitro selection of rice for Al on pH resistance. The experiment was arranged in completely randomized design with two treatments, that are two varieties (T 309 and Rojolele) and six levels of Al concentration (0, 100, 200, 300, 400, and 500 ppm). Selec-tion was conducted on regeneraSelec-tion, embryo, and callus stages. Result of the experiment showed that the two varieties had the same response in all media used. Treatment of MS + 2,4-D 2 mg/l + casein hydrolisat 3 g/l produced more nodules or bud than others. After in vitro selection, explant could regenerate in all Al concentrations, except in Al 500 ppm. Selection on callus stage produced the same result. Increasing Al concentration decreased regeneration ability. In the regen-eration and embryo stages, regenregen-eration of T 309 was not different with Rojolele. However, on the callus stage regener-ation of T 309 (47.76%) was higher than that of Rojolele (15.38%).
[Keywords: Oryza sativa, in vitro selection, aluminium resistance]
ABSTRAK
Salah satu upaya untuk meningkatkan produksi nasional padi adalah dengan memanfaatkan lahan masam. Namun, pe-manfaatan lahan masam menghadapi kendala kemasaman tanah tinggi, ketersediaan hara N, P, K, Ca, Mg, dan Mo rendah, serta adanya toksisitas Al dan Mn. Penanaman varietas toleran terhadap lahan masam dapat memecahkan masalah tersebut. Penelitian ini bertujuan untuk mendapatkan metode induksi kalus padi dan regenerasinya secara in vitro
serta memperoleh metode seleksi in vitro yang tepat untuk toleransi tanaman padi terhadap Al dan pH. Penelitian disusun secara faktorial dalam rancangan acak lengkap dengan dua perlakuan, yaitu varietas (T 309 dan Rojolele) dan konsentrasi Al (0, 100, 200, 300, 400, dan 500 ppm). Seleksi dilakukan pada tiga tahap yaitu tahap regenerasi, embrio, dan kalus. Hasil penelitian menunjukkan bahwa kedua varietas mem-punyai respons yang sama pada semua jenis media yang digunakan. Media MS + 2,4-D 2 mg/l + casein hidrolisat 3 mg/l
lebih banyak membentuk nodul-nodul bakal mata tunas dari-pada media lainnya. Regenerasi eksplan setelah perlakuan seleksi menunjukkan bahwa umumnya kedua varietas dapat beregenerasi pada semua perlakuan seleksi kecuali pada per-lakuan Al 500 ppm. Seleksi pada tahap kalus, regenerasi, dan tahap embrio menunjukkan hasil yang sama, yaitu makin meningkat konsentrasi Al, makin menurun daya regenerasi eksplan. Pada tahap embrio dan kalus, regenerasi varietas T 309 tidak berbeda dengan Rojolele, sedangkan pada tahap kalus, daya regenerasi T 309 (47,76%) lebih tinggi daripada Rojolele (15,38%).
[Kata kunci: Oryza sativa, seleksi in vitro, ketahanan terhadap aluminium]
PENDAHULUAN
Padi (Oryza sativa L.) merupakan tanaman pangan yang sangat penting karena hingga kini beras me-rupakan makanan pokok bagi sebagian penduduk dunia terutama Asia. Di Indonesia, beras juga me-rupakan komoditas strategis karena mempunyai pe-ngaruh yang besar terhadap kestabilan ekonomi dan politik.
Upaya untuk meningkatkan produksi beras terus dilakukan seiring dengan bertambahnya jumlah pen-duduk serta berkurangnya sawah beririgasi di Jawa akibat alih fungsi menjadi kawasan industri dan pemukiman. Selain itu, kemarau panjang serta banjir yang terjadi hampir setiap tahun ikut mempengaruhi kapasitas penyediaan beras sehingga impor tidak dapat dihindari. Kondisi ini diperburuk oleh krisis moneter yang berdampak pada melemahnya daya beli petani terhadap sarana produksi terutama pupuk dan pestisida.
Salah satu upaya untuk meningkatkan produksi padi adalah dengan memanfaatkan lahan kering yang ter-sedia cukup luas di luar Jawa. Dari luas total daratan Indonesia, sekitar 47,6 juta ha (32,4%) merupakan lahan kering yang umumnya didominasi oleh tanah masam Podsolik Merah Kuning (Karama dan Abdur-rachman 1993). Menurut Adhi (1993), tanah masam di Indonesia seperti Podsolik, Aluvial Hidromorf berpirit
Seleksi
in vitro
tanaman padi untuk sifat ketahanan
terhadap aluminium
In vitro selection of rice for aluminum resistance
Ragapadmi Purnamaningsih dan Ika Mariska
1Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian
(tanah sulfat masam) dan Organosol memiliki luas berturut-turut 27,5; 7,7; dan 20 juta ha.
Penelitian pada lahan Podsolik Merah Kuning menunjukkan bahwa pertumbuhan tanaman pada umumnya terhambat dan produktivitasnya rendah, karena adanya cekaman lingkungan berupa kemasam-an tinggi (pH rendah), ketersediakemasam-an hara N, P, K, Ca, Mg, dan Mo rendah, serta konsentrasi Al dan Mn yang mencapai tingkat beracun (Notohadiprawiro 1993). Keracunan Al dapat mengurangi pertumbuhan akar dan tajuk, akar menebal dan pendek karena proses pemanjangan sel terhambat sehingga penyerapan air dan hara berkurang (Marschner 1995).
Pengapuran dapat memperbaiki lahan masam, tetapi biayanya mahal. Pendekatan yang lebih efisien dan ramah lingkungan untuk menanggulangi hambatan tersebut adalah melalui pemuliaan untuk mendapatkan kultivar yang tahan terhadap cekaman lingkungan di samping produktivitas dan mutu hasilnya tinggi. Setiap kultivar memiliki ketahanan yang berbeda-beda terhadap Al (Van Sint Jan et al. 1997).
Keragaman genetik yang tinggi sangat diperlukan dalam perbaikan sifat-sifat tanaman. Secara konven-sional, peningkatan keragaman genetik dilakukan dengan memanfaatkan berbagai bahan genetik yang tersedia di alam dan selanjutnya dilakukan persilang-an secara konvensional. Namun demikipersilang-an, sifat-sifat tertentu sering tidak ditemukan pada sumber gen yang ada.
Dengan berkembangnya teknik kultur in vitro, ke-ragaman genetik dapat ditingkatkan antara lain melalui keragaman somaklonal. Salah satu metode variasi somaklonal adalah seleksi in vitro. Metode ini lebih efektif dan efisien karena penyaringan sifat yang diinginkan dapat dilakukan lebih terarah. Selain itu menurut Wenzel dan Fouroughi-Wehr (1993), seleksi
in vitro mempunyai beberapa keunggulan, yaitu tidak terlalu dipengaruhi lingkungan serta memungkinkan untuk melakukan seleksi pada tingkat sel dan untuk satu faktor tunggal. Selanjutnya Ramulu (1986) me-nyatakan bahwa keragaman somaklonal dan seleksi in vitro dapat menyebabkan terjadinya mutasi pada tingkat sel. Metode tersebut telah banyak dilakukan untuk memperoleh varian-varian tanaman yang resis-ten terhadap herbisida dan stres lingkungan.
Seleksi in vitro untuk mendapatkan varietas yang tahan lahan masam dilakukan dengan menggunakan komponen seleksi AlCl3.6H2O dan pH rendah (sekitar 4) (Short et al. 1987). Metode tersebut telah diaplika-sikan pada tanaman tomat dan kentang (Starvarek dan Rains 1984), sorgum (Smith et al. 1983), wortel (Ojima dan Ohira 1986), dan tembakau (Yamamoto et al. 1994)
serta dapat menghasilkan varietas baru yang tahan terhadap cekaman lingkungan. Penelitian ini bertuju-an untuk mendapatkbertuju-an metode induksi kalus padi dbertuju-an regenerasinya secara in vitro serta memperoleh meto-de seleksi in vitro yang tepat untuk toleransi tanaman padi terhadap Al dan pH.
BAHAN DAN METODE
Penelitian dilaksanakan di laboratorium kultur jaringan Balai Besar Penelitian dan Pengembangan Bioteknolo-gi dan Sumbedaya Genetik Pertanian pada bulan September 2001-Januari 2003. Percobaan disusun secara faktorial dengan rancangan lingkungan acak lengkap. Perlakuan yang diuji yaitu varietas dan media tumbuh serta varietas dan konsentrasi Al.
Eksplan yang digunakan adalah benih padi varietas Rojolele (Javanica) dan Taipei 309/T 309 (Japonica) yang merupakan varietas peka terhadap Al. Media dasar yang digunakan adalah media Murashige-Skoog (MS). Penelitian terdiri atas dua kegiatan yaitu (1) induksi kalus dan regenerasi sebelum seleksi, serta (2) seleksi kalus secara in vitro dan regenerasi setelah seleksi.
Pada kegiatan pertama, penelitian dilakukan untuk mendapatkan formulasi media untuk produksi kalus embriogenik serta regenerasinya. Kalus embriogenik adalah kalus yang mudah diregenerasikan menjadi planlet. Kalus diperoleh dengan cara menanam benih padi steril pada media MS0 (kontrol) dalam keadaan gelap selama 2-3 hari. Setelah itu embrio diisolasi dan ditumbuhkan pada media induksi kalus. Formulasi media untuk induksi kalus adalah MS + 2,4-D 0,5 mg/l + NAA 1 mg/l + BA 1 mg/l; MS + 2,4-D 2 mg/l + casein hidrolisat (CH) 3 g/l, serta MS + 2,4-D 20 mg/l, sedang-kan untuk regenerasi kalus adalah MS + BA 5 mg/l + IAA 0,8 mg/l. Formulasi media terbaik digunakan untuk seleksi in vitro serta regenerasi setelah seleksi.
Pada kegiatan kedua, seleksi secara in vitro dilaku-kan dengan menggunadilaku-kan komponen seleksi AlCl3.6H2O dan pH 4,0. Konsentrasi Al yang diuji yaitu 0, 100, 200, 300, 400, dan 500 ppm dengan 20 ulangan. Untuk me-munculkan sifat toksisitas dari Al pada media seleksi, garam-garam makro dari media MS dimodifikasi, yaitu kandungan NH4NO3 ditingkatkan dari 1.650 mg/l menjadi 2.400 mg/l, CaCl2.2H2O diturunkan dari 440 mg/l menjadi 15 mg/l, dan KH2PO4 diturunkan dari 170 mg/l menjadi 13 mg/l. Sebagai sumber Fe digunakan FeSO4 28 mg/l. Seleksi dilakukan pada tahap embrio, kalus, dan re-generasi. Seleksi pada tahap embrio dilakukan dengan cara meletakkan embrio yang diisolasi dari benih steril
pada media untuk induksi kalus yang diberi komponen seleksi, sedangkan seleksi pada tahap kalus dilakukan dengan cara meletakkan kalus pada media induksi kalus yang diberi komponen seleksi. Media yang digunakan untuk seleksi tahap embrio dan kalus ada-lah MS + 2,4-D 2 mg/l + CH 3 g/l + komponen seleksi. Seleksi pada tahap regenerasi dilakukan dengan cara menanam kalus pada media MS + 2,4-D 0,5 mg/l + BA 5 mg/l + IAA 0,8 mg/l ditambah komponen seleksi. Eksplan yang hidup pada media seleksi dipindahkan pada media regenerasi agar dapat tumbuh menjadi planlet.
Planlet hasil seleksi selanjutnya diaklimatisasi di rumah kaca. Pengamatan dilakukan terhadap persen-tase pembentukan kalus, diameter kalus, struktur kalus, persentase kalus yang dapat beregenerasi setelah seleksi, serta daya multiplikasi biakan setelah seleksi (jumlah anakan, panjang akar, dan jumlah akar).
HASIL DAN PEMBAHASAN Induksi kalus
Hasil analisis statistik menunjukkan bahwa tidak terdapat perbedaan yang nyata untuk jumlah kalus yang terbentuk dari kedua varietas, formulasi media yang digunakan serta interaksinya (Tabel 1). Hal tersebut mengindikasikan bahwa kedua varietas mempunyai respons yang sama pada semua jenis media, atau ketiga media yang digunakan mempunyai kemampuan yang sama dalam menginduksi pemben-tukan kalus. Karena itu, ketiga media tersebut dapat digunakan untuk menginduksi pembentukan kalus.
Kalus terbanyak diperoleh pada media MS + 2,4-D 20 mg/l. Menurut Wattimena (1992), zat pengatur tumbuh dari golongan auksin berperan antara lain dalam pembentukan kalus, morfogenesis akar dan tunas, serta embriogenesis. Pemilihan konsentrasi dan jenis auksin ditentukan antara lain oleh tipe per-tumbuhan dan perkembangan eksplan yang dike-hendaki. Menurut Sellars et al. (1990), auksin dengan daya aktivitas kuat (antara lain 2,4-D, NAA atau dikombinasikan dengan sitokinin konsentrasi rendah) umumnya digunakan untuk induksi kalus embriogenik. Hasil penelitian Hutami et al. (1999) menunjukkan bahwa penggunaan auksin dengan konsentrasi tinggi (10-40 mg/l) memberikan hasil yang lebih baik untuk perkembangan kalus embriogenik tanaman kedelai.
Pada penelitian ini terlihat adanya respons yang berbeda dari kedua varietas pada masing-masing formulasi media (Tabel 2). Diameter kalus T 309 lebih besar dan berbeda nyata dengan Rojolele, yang berarti
sel-sel kalus T 309 lebih cepat berdediferensiasi dari-pada Rojolele. Pertumbuhan kalus dari-pada media MS + 2,4-D 0,5 mg/l + NAA 1 mg/l + BA 1 mg/l lebih cepat dibandingkan pada media lainnya dengan diameter kalus 0,18 cm2.
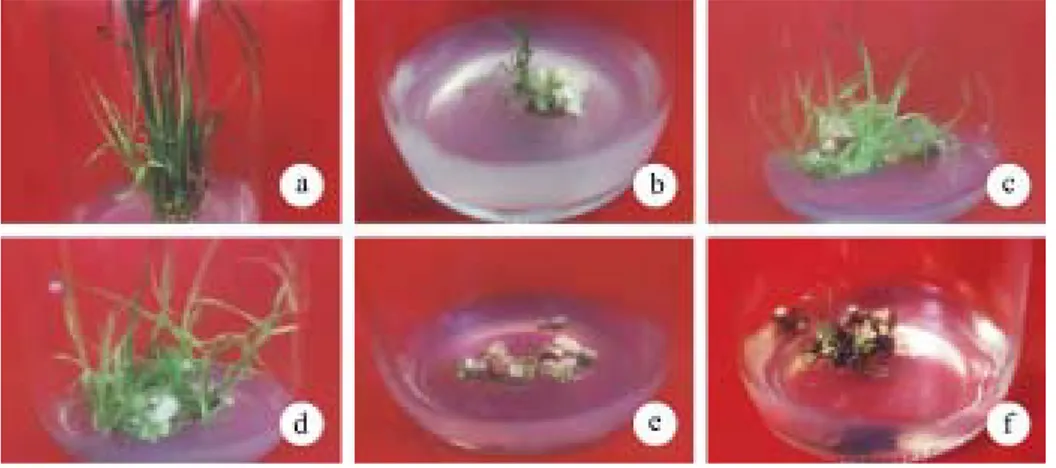
Berdasarkan penampakan kalus secara visual, ketiga formulasi media menghasilkan kalus yang remah (friabel), berwarna bening dan terbentuk nodul-nodul (Gambar 1). Hal ini menunjukkan bahwa kalus ke-mungkinan dapat beregenerasi menjadi planlet. Kalus pada media MS + 2,4-D 2 mg/l + CH 3 g/l lebih banyak membentuk nodul-nodul dibandingkan pada media lainnya sehingga diharapkan dapat diperoleh planlet lebih banyak. Pada media MS + 2,4-D 0,5 mg/l + NAA 1 mg/l + BA 1 mg/l, walaupun diperoleh kalus dengan ukuran lebih besar, kalus tersebut merupakan kalus rhizogenik, yaitu kalus yang lebih cepat membentuk akar daripada tunas. Hal ini kemungkinan disebabkan oleh ketidakseimbangan auksin dan sitokinin (2,4-D, NAA dan BA) dalam media sehingga eksplan lebih dahulu membentuk akar daripada tunas, padahal tunas diperlukan agar tanaman dapat melakukan fotosintesis. Wattimena (1992) menyatakan bahwa morfogenesis tunas dan akar dipengaruhi oleh nisbah auksin dan sitokinin. Nisbah auksin-sitokinin yang tinggi akan mendorong morfogenesis akar, sebaliknya nisbah sitokinin-auksin yang rendah akan mendorong pem-bentukan tunas.
Tabel 1. Persentase pembentukan kalus padi varietas T 3 0 9 d a n R o j o l e l e p a d a b e r b a g a i f o r m u l a s i m e d i a , umur 4 minggu.
Table1. Percentage of callus formation of T 309 and Rojolele rice varietas on some medium formulation, 4 weeks after treatment. Formulasi media Pembentukan kalus Media formulation Callus formation ( % ) T 309 Rojolele Rata-rata Average MS + 2,4-D 0.5mg/l + NAA 1 mg/l + BA 1 mg/l 9 2 , 4 5 9 2 , 0 0 9 2 , 3 1 MS + 2,4-D 2 mg/l + CH 3 g/l 9 0 , 0 0 8 7 , 5 0 8 8 , 8 9 MS + 2,4-D 20 mg/l 9 6 , 6 7 9 3 , 3 3 9 4 , 6 7 Rata-rata 9 2 , 4 8 9 0 , 9 1
Keterangan: Angka yang diikuti oleh huruf yang sama pada baris dan kolom yang sama, tidak berbeda nyata berdasarkan uji Duncan 5%.
Note: For each trait, means followed by one common letter are not significantly different according to Duncan’s Multiple Range Test (α = 0.05).
Pembentukan kalus embriogenik ditentukan oleh sumber N pada media. Thompson et al. (1981) dalam
Gunawan (1988) menyatakan bahwa asam amino me-rupakan sumber N-organik yang lebih cepat dapat diserap tanaman daripada N-anorganik. Penambahan asam amino seperti glutamin, casein hidrolisat, dan arginin pada media yang sudah mengandung auksin dapat meningkatkan keberhasilan pembentukan kalus embriogenik, karena dalam kloroplas asam amino dapat berperan sebagai prekursor untuk pembentukan asam nukleat dan proses seluler lainnya. Berdasarkan kemampuan dalam menginduksi pembentukan dan pertumbuhan kalus serta penampakan kalus yang dihasilkan, media MS + 2,4D 2 mg/l + CH 3 g/l selanjut-nya digunakan untuk induksi kalus.
Seleksi in vitro dan regenerasi tanaman
Penampakan eksplan pada saat diseleksi dengan Al dan pH rendah berbeda-beda, bergantung pada ke-tahanan masing-masing sel terhadap komponen
seleksi serta taraf konsentrasi Al (Gambar 2). Sel-sel yang tidak mempunyai sifat ketahanan berwarna hitam serta tidak mampu tumbuh, sedangkan sel-sel yang tahan tetap tumbuh dan berwarna hijau. Pada umum-nya makin tinggi konsentrasi Al, kalus yang berwarna hitam makin banyak. Kalus yang tetap berwarna hijau diharapkan mempunyai sifat ketahanan terhadap Al.
Regenerasi eksplan setelah perlakuan seleksi
Eksplan T 309 dan Rojolele dapat beregenerasi se-telah perlakuan seleksi in vitro, tetapi waktu yang diperlukan untuk beregenerasi berbeda bergantung pada varietas dan tahapan seleksi (Tabel 3). Penampak-an biakPenampak-an pada media regenerasi setelah seleksi disajikan pada Gambar 3.
Pada umumnya T 309 lebih cepat beregenerasi pada semua tahapan seleksi dibandingkan Rojolele. Seleksi saat eksplan berada pada tahap regenerasi menyebab-kan eksplan lebih cepat beregenerasi, baik pada varietas T 309 maupun Rojolele.
Gambar 1. Pembentukan kalus padi pada beberapa formulasi media; a = MS + 2,4-D 0,5 mg/l + NAA 1 mg/l + BA 1 mg/l, b = MS + 2,4-D 2 mg/l + casein hidrolisat 3 g/l, c = MS + 2,4-D 20 mg/l.
Fig. 1. Callus formation of rice on some medium formulation; a = MS + 2,4-D 0.5 mg/l + NAA 1 mg/l + BA 1 mg/l, b = MS + 2,4-D 2 mg/l + casein hidrolisat 3 g/l, c = MS + 2,4-D 20 mg/l.
Tabel 2. Diameter kalus dari eksplan padi T 309 dan Rojolele pada perlakuan formulasi media umur 4 minggu.
Table 2. Callus diameter of T 309 and Rojolele rice varieties on some medium formulatio at, 4 weeks after treatment.
Formulasi media
Diameter kalus/Callus diameter
Visual/Visual Media formulation (cm2) T 309 Rojolele Rata-rata Average MS + 2,4D 0,5 mg/l + NAA1mg/l
+ BA1mg/l 0 , 1 9 0 , 1 2 0 , 1 8 a Bening, globuler/Transparant, globuler
MS + 2,4-D 2 mg/l + CH 3 g/l 0 , 1 4 0 , 0 7 0 , 1 2 b Bening, globuler/Transparant, globuler
MS + 2,4-D 20 mg/l 0 , 1 4 0 , 0 7 0 , 1 1 b Bening, globuler/Transparant, globuler
Rata-rata/Average 0 , 1 9 a 0 , 0 8 b
Keterangan: Angka pada baris dan kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan DMRT 5%.
Seleksi langsung pada embrio tampaknya tidak dapat menghentikan proses pembelahan dan pertum-buhan sel, sehingga kalus tetap terbentuk dan ber-kembang walaupun ukurannya lebih kecil. Hasil penelitian Mariska et al. (2000) pada tanaman kedelai menunjukkan bahwa seleksi pada tahap embrio menyebabkan sebagian besar eksplan tidak dapat membentuk kalus.
Seleksi pada saat eksplan pada fase kalus memberi-kan respons regenerasi yang paling lambat, yaitu 21 hari untuk T 309 dan 40 hari untuk Rojolele (Tabel 3). Diduga hal ini disebabkan eksplan terlalu lama berada pada tahap kalus, yaitu pada saat induksi kalus (6 minggu) dan selama seleksi (8 minggu). Selama fase tersebut sel terus membelah sehingga dapat menurun-kan daya regenerasi jaringan. Sel somatik yang terlalu lama diinkubasi pada media yang mengandung auksin kuat dapat menurun daya regenerasinya. Namun makin lama eksplan berada pada fase kalus maka kemungkin-an terjadinya mutasi makin besar, sehingga harapkemungkin-an untuk memperoleh nomor-nomor tanaman padi yang tahan terhadap Al dan pH rendah makin besar. Selain
Gambar 2. Seleksi in vitro padi pada beberapa perlakuan aluminium; a = kontrol pH 5,8, b = kontrol pH 4,0, c = Al 100 ppm, d = Al 200 ppm, e = Al 300 ppm, f = Al 400 ppm, g = Al 500 ppm.
Fig. 2.In vitro selection of rice on some medium formulation; a = control pH 5.8, b = control pH 4.0, c = Al 100 ppm, d = Al 200 ppm, e = Al 300 ppm, f = Al 400 ppm, g = Al 500 ppm.
Gambar 3. Regenersi eksplan setelah perlakuan seleksi in vitro; a = kontrol pH 5.8, b = kontrol pH 4.0, c = Al 100 ppm, d = Al 200 ppm, e = Al 300 ppm, f = Al 400 ppm.
Fig. 3. Regeneration of the explant after in vitro selection; a = control pH 5.8, b = control pH 4.0, c = Al 100 ppm, d = Al 200 ppm, e = Al 300 ppm, f = Al 400 ppm.
Tabel 3. Waktu regenerasi eksplan padi T 309 dan Rojolele pada beberapa tahapan seleksi.
Table 3. Time of explant regeneration on different selection stage of T309 and Rojolele rice varieties.
Tahapan seleksi Waktu regenerasi (hari)
Selection stage Regeneration time (day)
T 309 Rojolele
Tahap regenerasi/Regeneration 7 1 5
Tahap embrio/Embryo 1 0 1 6
itu penggunaan auksin seperti 2,4-D dapat menyebab-kan terjadinya perubahan sifat genetik. Keragaman yang ditimbulkan disebabkan oleh daya aktivitasnya yang kuat dalam memacu proses dediferensiasi, se-hingga kromosom menjadi tidak stabil dan meng-ganggu replikasi DNA (Ahloowalia 1986).
Seleksi in vitro
Seleksi in vitro pada tahap regenerasi
Pada tahap seleksi ini, embrio terlebih dahulu diinduksi membentuk kalus kemudian kalus dipindahkan pada media regenerasi dengan menambahkan Al sebagai komponen seleksi. Umumnya eksplan dari kedua varietas dapat beregenerasi membentuk planlet pada semua taraf konsentrasi Al kecuali pada Al 500 ppm (Tabel 4; Gambar 4). Persentase regenerasi tertinggi diperoleh pada kontrol/tanpa seleksi (pH 5,8) yaitu 69,6%. Penurunan pH menjadi 4,0 mengurangi daya regenerasi eksplan menjadi 42,9%.
Varietas T 309 dan Rojolele memberikan respons yang sama dan tidak berbeda nyata berdasarkan uji statistik. Namun, jumlah eksplan Rojolele yang be-regenerasi lebih banyak daripada T 309, tetapi waktu yang diperlukan T 309 untuk beregenerasi lebih singkat. Diduga hal ini karena proses dediferensiasi kalus T 309 yang sangat cepat dapat menurunkan kemampuan regenerasi, sehingga organ yang ter-bentuk tertutup oleh kalus.
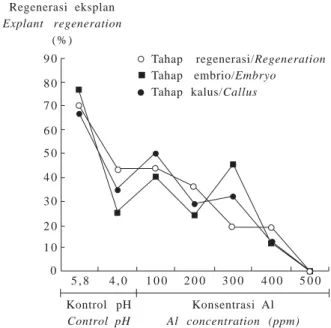
Gambar 4. Persentase regenerasi eksplan padi pada beberapa tahap seleksi in vitro.
Fig. 4. Percentage of rice explant regeneration on some in vitro selection stage.
Kontrol pH Control pH 5 , 8 4 , 0 1 0 0 2 0 0 3 0 0 4 0 0 5 0 0 Konsentrasi Al Al concentration (ppm) Regenerasi eksplan Explant regeneration ( % ) 9 0 8 0 7 0 6 0 5 0 4 0 3 0 2 0 1 0 0
Tahap regenerasi/Regeneration
Tahap embrio/Embryo
Tahap kalus/Callus
Penambahan Al menurunkan daya regenerasi jaringan. Makin tinggi konsentrasi Al, daya regenerasi eksplan makin rendah meskipun secara statistik tidak berbeda nyata antara perlakuan kontrol pH 5,8 dengan kontrol pH 4,0 dan Al 100 ppm. Perbedaan nyata baru terlihat pada penambahan Al 200, 300, 400, dan 500 ppm dengan kontrol pH 5,8.
Seleksi in vitro pada tahap embrio
Seperti halnya seleksi pada tahap regenerasi, seleksi langsung pada embrio memberikan hasil yang sama untuk kedua varietas. Eksplan varietas T 309 yang dapat beregenerasi berkisar 33,33% dan tidak berbeda nyata dengan Rojolele (31,25%) (Tabel 5; Gambar 4). Penurunan pH dan penambahan Al pada semua taraf konsentrasi mengurangi daya regenerasi eksplan dan berbeda nyata dengan kontrol pH 5,8. Persentase regenerasi tertinggi diperoleh dari perlakuan kontrol pH 5,8 yaitu 76,7%. Penurunan pH mengurangi daya regenerasi eksplan menjadi 25%. Daya regenerasi eksplan meningkat dengan penambahan Al 100 ppm dan 300 ppm walaupun tidak berbeda nyata dengan kontrol pH 4,0. Diduga pada perlakuan Al 100 dan 300 ppm terjadi mutasi, yaitu eksplan mempunyai mekanisme untuk bertahan dari toksisitas Al sehingga dapat memenuhi kebutuhan akan unsur hara, atau eksplan dapat melindungi diri dari kerusakan karena toksisitas Al. Dengan kemasaman yang rendah, beberapa komponen organik dan anorganik seperti
Tabel 4. Persentase kalus T 309 dan Rojolele yang dapat m e m b e n t u k t u n a s p a d a s e l e k s i t a h a p r e g e n e r a s i .
Table 4. Percentage of callus forming shoots on regenera-tion selecregenera-tion stage of T 309 and Rojolele rice varieties.
Perlakuan seleksi
Kalus membentuk tunas
Selection treatment
Callus forming shoots
( % ) T 309 Rojolele Rata-rata Average Kontrol pH 5,8 7 2 , 7 3 6 0 , 0 0 6 9 , 6 0 a Kontrol pH 4,0 4 4 , 4 4 4 0 , 0 0 42,90ab Al 100 3 0 , 0 0 6 2 , 5 0 44,40ab Al 200 2 3 , 0 8 3 3 , 3 3 36,40b Al 300 2 3 , 0 8 1 5 , 3 0 19,20bc Al 400 2 0 , 0 0 1 6 , 6 7 18,80bc Al 500 0 , 0 0 0 , 0 0 00 , 0 0 c Rata-rata/Average 3 2 , 0 5 3 4 , 4 3
Keterangan: Angka pada baris dan kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan DMRT 5%.
Notes: For each trait, means followed by one common letter are not significantly different according to DMRT 5%.
PO4-, NH 4
+ dan vitamin B1 yang terdapat dalam media
tidak dapat larut secara sempurna sehingga tidak tersedia secara maksimal.
Eksplan Rojolele mempunyai daya regenerasi yang lebih kecil dibandingkan T 309, bahkan pada perlakuan Al 400 dan 500 ppm eksplan tidak dapat beregenerasi. Kemungkinan hal ini disebabkan Rojolele lebih sensitif terhadap Al. Hasil penelitian Mariska et al. (2000) pada tanaman kedelai menunjukkan bahwa seleksi pada tahap embrio umumnya tidak dapat menghasilkan kalus dan tidak dapat beregenerasi. Menurut Taylor (1991), eksplan yang dapat bertahan hidup dan be-regenerasi menunjukkan adanya sifat ketahanan pada tingkat sel. Selanjutnya Bertin dan Bahaurmont (1997) menyatakan bahwa seleksi pada tingkat sel sangat potensial untuk menghasilkan genotipe baru yang adaptif terhadap cekaman lingkungan.
Pada penelitian ini, seleksi langsung pada eksplan embrio lebih banyak memunculkan sel-sel yang mempunyai sifat ketahanan terhadap Al. Bahkan pada penambahan Al 300 ppm, jumlah tanaman yang beregenerasi lebih tinggi daripada seleksi pada tahap kalus dan regenerasi (Gambar 4).
Variasi somaklonal akan menghasilkan individu-individu baru sebagai akibat adanya ketidakstabilan selama periode kultur in vitro. Ketidakstabilan ter-sebut dapat berupa perubahan jumlah atau struktur kromosom, penggunaan zat pengatur tumbuh dengan aktivitas kuat seperti 2,4-D atau lamanya periode kultur (Larkin dan Scowcroft 1981).
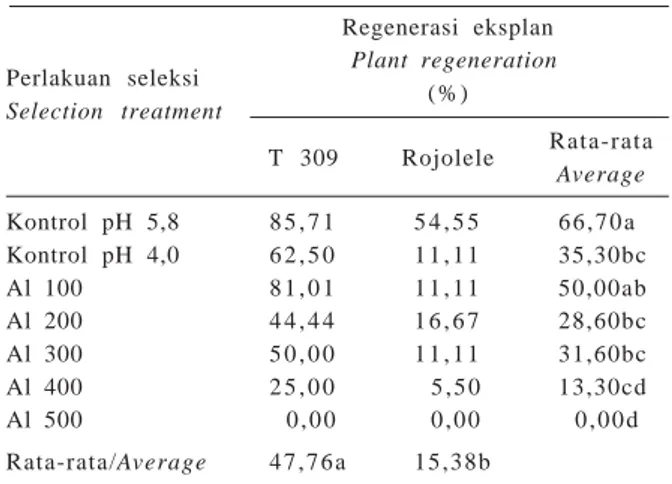
Seleksi in vitro pada tahap kalus
Kedua varietas memberikan respons yang berbeda setelah perlakuan seleksi pada tahap kalus. Umumnya kedua varietas dapat beregenerasi pada semua perlakuan konsentrasi Al kecuali pada konsentrasi Al 500 ppm (Tabel 6; Gambar 4). Eksplan T 309 yang beregenerasi lebih banyak (47,76%) dan berbeda nyata dengan Rojolele (15,38%). Penambahan Al pada beberapa taraf konsentrasi juga memperlihatkan respons yang berbeda pada kedua varietas serta berbeda nyata secara statistik. Persentase regenerasi tertinggi diperoleh dari perlakuan kontrol pH 5,8 yaitu 66,7% dan berbeda nyata dengan semua perlakuan Al dan penurunan pH kecuali perlakuan Al 100 ppm. Makin tinggi konsentrasi Al, makin rendah persentase regenerasi bahkan pada Al 500 ppm eksplan tidak dapat beregenerasi. Diduga hal ini disebabkan oleh makin lamanya eksplan berada pada fase kalus di mana proses dediferensiasi terus berlangsung tanpa terjadi proses diferensiasi. Selain itu penambahan Al ke dalam media menyebabkan terjadinya ketidakseimbangan ketersediaan unsur hara sehingga proses diferensiasi terhambat dan akhirnya tanaman tidak mampu bere-generasi. Hal ini sejalan dengan hasil penelitian Van Sint Jan et al. (1997) bahwa peningkatan konsentrasi Al menurunkan kemampuan regenerasi tanaman padi. Memberikan kondisi stres dengan konsentrasi Al yang tinggi menguntungkan karena stres dapat me-nurunkan jumlah tanaman yang akan diseleksi secara
in vitro sehingga merupakan seleksi bertahap.
Me-Ta b e l 5 . P e r s e n t a s e re g e n e r a s i e k s p l a n p a d i v a r i e t a s T 309 dan Rojolele pada seleksi tahap embrio.
Ta b l e 5 . P e rc e n t a g e o f e x p l a n t re g e n e r a t i o n o n e m b r y o selection of T 309 and Rojolele rice varieties.
Perlakuan seleksi Regenerasi eksplan Selection treatment Explant regeneration ( % ) T 309 Rojolele Rata-rata Average Kontrol pH 5,8 8 0 , 0 0 7 5 , 0 0 7 6 , 7 0 a Kontrol pH 4,0 2 5 , 0 0 2 5 , 0 0 25,00bc Al 100 6 2 , 5 0 3 3 , 3 3 40,00b Al 200 1 8 , 1 8 2 8 , 5 7 24,00bcd Al 300 6 0 , 0 0 3 0 , 0 0 45,00b Al 400 1 8 , 7 5 00 , 0 0 11,50cd Al 500 00 , 0 0 00 , 0 0 00,00d Rata-rata/Average 3 3 , 3 3 3 1 , 2 5
Keterangan: Angka pada baris dan kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan DMRT 1%.
Notes: For each trait, means followed by one common letter are not significantly different according to DMRT 5%.
Tabel 6. Persentase regenerasi eksplan padi varietas T 309 dan Rojolele pada seleksi tahap kalus.
Table 6. Percentage of explant regeneration on callus selection stage of T 309 and Rojolele rice varieties.
Perlakuan seleksi Regenerasi eksplan Selection treatment Plant regeneration ( % ) T 309 Rojolele Rata-rata Average Kontrol pH 5,8 8 5 , 7 1 5 4 , 5 5 6 6 , 7 0 a Kontrol pH 4,0 6 2 , 5 0 1 1 , 1 1 35,30bc Al 100 8 1 , 0 1 1 1 , 1 1 50,00ab Al 200 4 4 , 4 4 1 6 , 6 7 28,60bc Al 300 5 0 , 0 0 1 1 , 1 1 31,60bc Al 400 2 5 , 0 0 5 , 5 0 13,30cd Al 500 0 , 0 0 0 , 0 0 0 , 0 0 d Rata-rata/Average 4 7 , 7 6 a 15,38b
Keterangan: Angka pada baris dan kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan DMRT 1%.
Notes: For each trait, means followed by one common letter are not significantly different according to DMRT 5%.
nurut Ojima dan Ohira (1986), masalah regenerasi akan meningkat bila massa sel dikulturkan pada media yang mempunyai komponen seleksi yang bersifat toksik. Diperlukan waktu 4 tahun untuk meregenerasikan sel wortel yang tahan Al. Demikian pula sel tembakau yang telah diseleksi dengan Al sulit beregenerasi (Yamamoto et al. 1994).
Lebih tingginya eksplan T 309 yang beregenerasi kemungkinan disebabkan kemampuan regenerasi T 309 lebih baik daripada Rojolele. Selain itu makin lama eksplan berada pada fase kalus makin menurun daya regenerasi jaringan. Eksplan yang dapat be-regenerasi kemungkinan karena telah terjadi mutasi sehingga eksplan dapat bertahap hidup. Zat pengatur tumbuh merupakan salah satu faktor utama yang menyebabkan mutasi pada kultur in vitro (Singh et al. 1987). Wattimena (1992) menyatakan bahwa pe-nyimpanan kalus dalam waktu yang lama dalam media yang mengandung 2,4-D dapat meningkatkan ke-ragaman genetik.
Daya regenerasi Rojolele lebih rendah pada semua perlakuan Al daripada T 309 dan penurunan kemam-puan regenerasinya sangat besar. Diduga Rojolele sangat sensitif terhadap Al dan pH rendah.
Daya regenerasi biakan setelah seleksi
Eksplan yang bertahan hidup setelah perlakuan se-leksi dengan penambahan Al dan pH rendah (pH 4,0) dipindahkan ke media regenerasi untuk memulihkan kondisi eksplan serta menumbuhkannya menjadi planlet. Untuk mengetahui kondisi biakan setelah perlakuan seleksi perlu diketahui daya multiplikasi biakan. Planlet yang dapat bertahan dari stres lingkungan, apabila ditumbuhkan pada lingkungan tanpa stres diharapkan tetap tumbuh dengan baik.
Jumlah anakan T 309 dan Rojolele tidak berbeda nyata secara statistik, tetapi perlakuan penurunan pH dan penambahan Al memperlihatkan respons yang berbeda-beda (Tabel 7). Jumlah anakan terbanyak diperoleh dari perlakuan Al 200 ppm yang berbeda nyata dengan perlakuan lainnya kecuali Al 300 ppm. Jumlah anakan paling sedikit berasal dari perlakuan kontrol pH 4,0.
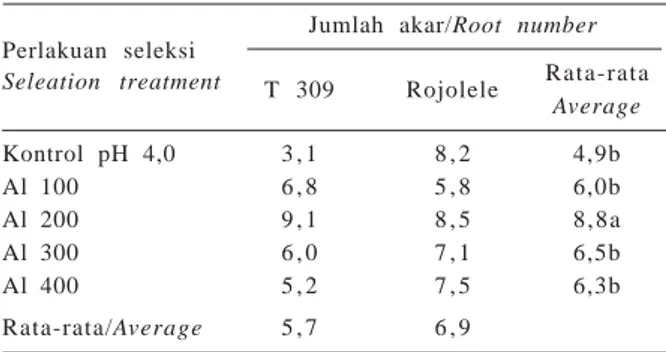
Seperti halnya jumlah anakan, jumlah akar T 309 tidak berbeda nyata dengan Rojolele, namun penu-runan pH dan penambahan Al menghasilkan jumlah akar yang berbeda. Jumlah akar terbanyak diperoleh dari perlakuan Al 200 ppm yaitu 8,8 dan berbeda nyata dengan perlakuan lainnya (Tabel 8).
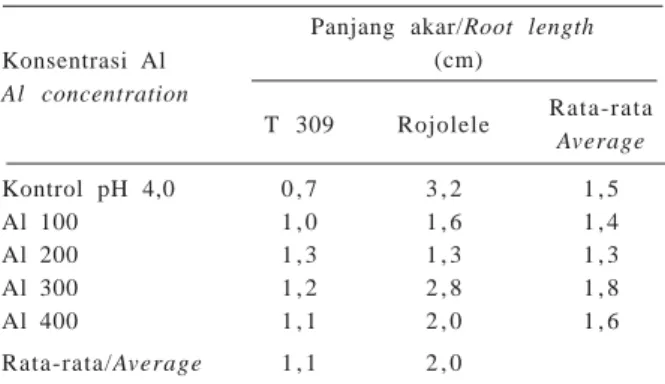
Panjang akar T 309 dan Rojolele tidak berbeda nyata secara statistik pada semua perlakuan penurunan pH
dan penambahan Al (Tabel 9). Semua planlet dapat tumbuh dengan baik pada media regenerasi. Biakan dapat membentuk anakan baru dan berakar.
KESIMPULAN
Ketiga formulasi media yang digunakan menghasilkan kalus yang remah, globuler dan berwarna bening, tetapi media MS + 2,4-D 2 mg/l + casein hidrolisat 3 g/l lebih banyak membentuk nodul-nodul bakal mata tunas. Seleksi pada tahap embrio dan tahap kalus lebih banyak memunculkan tanaman-tanaman baru yang
Tabel 8. Jumlah akar planlet pada varietas T 309 dan Rojolele hasil seleksi in vitro, umur 1 bulan.
Table 8. Root number of T 309 and Rojolele plantlet from in vitro selection treatment at 1 month.
Perlakuan seleksi
Jumlah akar/Root number Seleation treatment T 309 Rojolele Rata-rata
Average Kontrol pH 4,0 3 , 1 8 , 2 4,9b Al 100 6 , 8 5 , 8 6,0b Al 200 9 , 1 8 , 5 8 , 8 a Al 300 6 , 0 7 , 1 6,5b Al 400 5 , 2 7 , 5 6,3b Rata-rata/Average 5 , 7 6 , 9
Keterangan: Angka yang diikuti oleh huruf yang sama pada kolom atau baris yang sama tidak berbeda nyata berdasarkan uji Duncan 5%.
Notes: For each trait, means followed by one common letter are not significantly different according to Duncan’s Multiple R a n g e .
Tabel 7. Jumlah anakan planlet padi varietas T 309 dan Rojolele hasil seleksi in vitro umur 1 bulan.
Table 7. Shoot number of rice planlet of T 309 and Rojolele rice varieties from in vitro selection at 1 month.
Perlakuan seleksi Jumlah anakan/Tiller number
Selection treatment T 309 Rojolele Rata-rata Average Kontrol pH 4,0 3 , 8 4 , 0 2 , 7 c Al 100 3 , 4 3 , 6 3,5bc Al 200 6 , 8 3 , 6 4 , 7 a Al 300 3 , 4 4 , 5 3,8ab Al 400 3 , 0 3 , 0 3,0bc Rata-rata/Average 3 , 3 3 , 7
Keterangan: Angka yang diikuti oleh huruf yang sama pada baris dan kolom yang sama tidak berbeda nyata berdasarkan DMRT 5%.
Notes: For each trait, means followed by one common letter are not significantly different according to DMRT at 5%.
Bogor 23-25 Agustus 1993. Pusat Penelitian dan Pe-ngembangan Tanaman Pangan, Bogor.
Larkin, P.J. and W.R. Scowcroft. 1981. Somaclonal variation-a novel source of variability from cell cultures for plant improvement. Theor. App. Genet. 60: 197-214. Mariska, I., Hobir., M. Tombe, D. Manohara, S. Hutami. W.H.
Adil, Y. Rusyadi, E. Gati, R. Purnamaningsih, D. Sukma-djaja, M. Kosmiatin, A. Husni., dan S. Rahayu. 2000. Peningkatan keragaman genetik melalui seleksi in vitro dan keragaman somaklonal untuk ketahanan terhadap faktor biotik dan abiotik. Laporan Hasil Penelitian Balai Penelitian Bioteknologi Tanaman Pangan, Bogor. Marschner, H. 1995. Mineral Nutrition of Higher Plants. 2nd
Ed. Academic Press Harcourt Brace & Company, London. 889 pp.
Notohadiprawiro, T. 1983. Persoalan tanah masam dalam pembangunan pertanian Indonesia. Buletin Fakultas Per-tanian, Universitas Gadjah Mada 18: 44-47.
Ojima, K. and K. Ohira. 1986. Characterization and regen-eration of aluminium tolerant variant from carrot cell culture. Japan Annual Plant Tissue Culture, Tokyo. Ramulu, S R. 1986. Origin and nature of somaclonal variation
in potato. p. 189-197. In J. Serial (Ed.). Somaclonal Variation and Crop Improvement. Martinus Nijhoff Publisher, USA. Sellars, R.M., G.M. Southward, and G.C. Philips. 1990. Adventitious somatic embryogenesis from culture immature zygotic embryos of peanut and soybean. Crop Sci. 30: 408-413. Short, K.C., I. Warburton, and A.V. Roberts. 1987. In vitro
hardening of cultured cauliflower and chrysanthemum plantlets to humidity. Acra Hort. (212): 329-334. Singh, K J., K.P. Kollipara, and T. Haymowithz. 1987. Inter
subgeneric hybridization of soybean with perennial species
Glycine clandestina Wendl. Theor. App. Genet. 74: 391-396. Smith, R.H., S. Bhaskaran, and K. Scherts. 1983. Sorghum plant regeneration from aluminium selection media. Plant Cell Rep. 2: 129-132.
Starvarek, S.Y. and D.W. Rains. 1984. The development of tolerance cell to mineral stress. Hort. Sci. 19: 377-382. Taylor, G.J. 1991. Current views of the Al stress response: the
physiological basis of tolerance. Curr. Top. Plant Biochem Physiol. 10: 57-93.
Van Sint Jan,V., C.C. de Macedo, J.M. Kinet, and J. Bouharmont. 1997. Selection of Al resistant plants from a sensitive rice cultivar using somaclonal variation, in vitro
selection, and hydroponic cultures. Euphytica 97: 303-310. Wattimena, G.A. 1992. Bioteknologi Tanaman. Pusat Antar Universitas, Institut Pertanian Bogor, Bogor. 308 hlm. Wenzel, G. and B. Foroughi-Wehr. 1993. In vitro selection. p.
45-59 In M.D. Hayward, N.O. Bosemark, and Romagosa (Eds.). Plant Breeding. Principle and Prospects. Chapman & Hall, London.
Yamamoto, Y., R. Sanae, Yi-Chieh, K. Ono, K. Monibu, and H. Matsumoto. 1994. Quantitative estimation of alumi-nium toxicity in cultured tobacco cells. Correlation between aluminium uptake and growth inhibitor. Plant Cell Physiol. 35(4): 575-583.
mempunyai sifat ketahanan terhadap Al dan pH rendah. Makin meningkat konsentrasi Al, daya regenerasi eksplan membentuk tunas makin menurun. Pada konsentrasi Al 500 ppm semua eksplan Rojolele maupun T 309 tidak mampu beregenerasi.
DAFTAR PUSTAKA
Adhi, I P.G.W. 1993. Pengaruh tanah masam untuk kedelai. hlm. 171-188. Dalam Somaatmadja, M. Ismunaji, Sumarno, M. Syam, S.O. Manurung, Yuswadi (Ed.). Kedelai. Pusat Penelitian dan Pengembangan Tanaman Pangan, Bogor. Ahlowalia, B.S. 1986. Limitations to the use of somaclonal
variation in crop improvement. p. 14-27. In. J. Semal (Ed.). Somaclonal Variation and Crop Improvement. Martinus Nijhoff Publisher, USA.
Bertin P. and J. Bouharmont. 1997. Use of somaclonal variation and in vitro selection for chilling tolerance improvement in rice. Euphytica 96: 135-142.
Gunawan, L.V. 1988. Teknik Kultur Jaringan Tumbuhan. PAU Bioteknologi, Institut Pertanian Bogor, Bogor. 304 hlm. Hutami, S., I. Mariska, A. Husni, dan M. Kosmiatin. 1999. Regenerasi dan seleksi in vitro untuk mendapatkan sifat ketahanan terhadap aluminium pada tanaman kedelai. Laporan Hasil Penelitian Balai Penelitian Bioteknologi Tanaman Pangan, Bogor. 20 hlm.
Karama, A.S. dan A. Abdurrachman. 1993. Optimasi pemanfaatan sumberdaya lahan berwawasan lingkungan. hlm. 98-112. Prosiding Simposium Penelitian Tanaman Pangan III,
Tabel 9. Panjang akar planlet padi T 309 dan Rojolele hasil seleksi in vitro, umur 1 bulan.
Table 9. Root length of T 309 and Rojolele plantlet from in vitro selection treatment at 1 month.
Konsentrasi Al
Panjang akar/Root length Al concentration (cm) T 309 Rojolele Rata-rata Average Kontrol pH 4,0 0 , 7 3 , 2 1 , 5 Al 100 1 , 0 1 , 6 1 , 4 Al 200 1 , 3 1 , 3 1 , 3 Al 300 1 , 2 2 , 8 1 , 8 Al 400 1 , 1 2 , 0 1 , 6 Rata-rata/Average 1 , 1 2 , 0
Keterangan: Angka yang diikuti oleh huruf yang sama pada baris dan kolom yang sama tidak berbeda nyata berdasarkan uji Duncan 5%.
Notes: For each trait, means followed by one common letter are not significantly different according to Duncan’s Multiple R a n g e .