L
Journal of Experimental Marine Biology and Ecology, 241 (1999) 159–177
Predation: a causal mechanism for variability in intertidal
bivalve populations
a ,* a b
Michael G. Richards , Mark Huxham , Andy Bryant a
Department of Biological Sciences, Napier University, 10 Colinton Road, Edinburgh EH10 5DT, Scotland,
UK b
Marine Laboratory, P.O. Box 101, Victoria Road, Aberdeen AB11 9DB, Scotland, UK Received 1 December 1998; received in revised form 8 April 1999; accepted 1 May 1999
Abstract
Two caging experiments were conducted on an intertidal mudflat to assess the impact of predation on two species of bivalves; Macoma balthica and Cerastoderma edule. The first caging
2
experiment enclosed the shore crab, Carcinus maenas, carapace width 15–25 mm, in 1-m cages. The three treatments were zero Carcinus (control), five Carcinus (the highest natural densities found at this site) and 20 Carcinus (a raised density). The experiment ran for approximately 1 month in August 1996. An exclosure experiment was conducted at the same site the following year and ran for approximately 2 months during September and October. The three treatments were full cages, partial cages and an un-caged control. Results from the enclosure indicate that crab predation was responsible for a reduction of both species of bivalve even at high naturally occurring crab densities. Significant reductions were observed between all treatments for C. edule and between the control and 20 Carcinus treatment for M. balthica. Significant differences occurred in the exclosure experiment between all treatments for C. edule. Almost all of the C. edule were removed in the un-caged areas. Significant differences only occurred between the full cages and un-caged areas for M. balthica. In both experiments there was a preference for C. edule over M. balthica and for the larger size classes of both species. Previous work at the present study site showed Carcinus predation to be site-specific, with predation rates dependent on the sediment type. The density of Carcinus at this site is extremely variable; this together with sediment-specific consumption of bivalves may be one of the causal mechanisms for both the spatial and temporal variability shown in populations of M. balthica and C. edule at this site. 1999 Elsevier Science B.V. All rights reserved.
Keywords: Caging experiments; Carcinus maenas; Cerastoderma edule; Macoma balthica; Predation; Variability
*Corresponding author. Tel.: 144-131-455-2372; fax: 144-131-455-2291.
E-mail address: [email protected] (M.G. Richards)
1. Introduction
Many authors have demonstrated temporal and spatial variability of bivalves in ¨
intertidal areas (e.g. Gunther, 1991; Skilleter, 1994; Thrush et al., 1997). The causal mechanisms of this variability may include: hydrodynamics (e.g. Butman, 1987; Armonies, 1996; Lindegarth, 1996), adult / juvenile interactions (e.g. Thrush et al., 1996;
¨
Thrush et al., 1997), larval choice (e.g. Moller, 1986; Butman, 1987), and predation (e.g. ´
Wilson, 1991; Olafsson et al., 1994 and references therein; Hines et al., 1997). This paper looks at the last of these by utilising caging experiments.
There is much debate over the usefulness of caging experiments in marine soft ´
sediments (e.g. Virnstein, 1978; Hall et al., 1990a,b; Wilson, 1991; Olafsson et al., 1994), however they are still recognised as the most reliable way of assessing the impact of predation within these systems (Hall et al., 1990a). Most caging studies have looked at the effects of predation at the community level. However, it is suggested that they are better used to look at the effect of predation on one, or a few species (Hall et al., 1990a). If used to assess predation on a few species, they can be timed to avoid potential artefacts such as settlement into cages of juvenile prey or predators during the course of the experiment. As any movement into and out of cages may confound the results of caging experiments (Thrush, 1986; Frid, 1989; Hall et al., 1990a,b; Hewitt et al., 1997) it is best to avoid the main settling and redistribution events. This becomes more difficult to achieve when trying to assess the impact of predation on a community level as prey species may settle, and or redistribute, over larger temporal scales.
The main large predators occurring at the present study site (Blackness, S.E. Scotland) are: epibenthic invertebrates, the shore crab, Carcinus maenas (L.) (henceforth Carcinus) and the common shrimp, Crangon crangon (L.); fish, the flounder, Platichthys flesus (L.); avian, shelduck, Tadorna tadorna (L.) and redshank, Tringa totanus (L.). The impact of predation on macrobenthos is equivocal, several experiments have failed to show direct significant effects of predation by fish and shorebirds on intertidal prey densities (Kent and Day, 1983; Raffaelli and Milne, 1987; Jaquet and Raffaelli, 1989) whereas others have shown significant direct or indirect effects (Kent and Day, 1983; Quammen, 1984; Thrush et al., 1994; Hines et al., 1997). Even when considering the effect of one predator such as Carcinus the results have varied. Scherer and Reise (1981) and Gee et al. (1985) have reported significant effects of Carcinus predation on the macrofauna. These contrast with studies by Frid and James (1988) and Raffaelli et al. (1989) who found little or no impact. The differences may be due to site specific interactions or, as seems more likely, the use of unrealistically high predator densities in the studies which have shown significant results (Raffaelli et al., 1989).
successful recruitment after severe winters (Beukema, 1991 and references therein). This has been attributed to reduced survival and later migration onto the tidal flats by
¨
Carcinus (Pihl and Rosenberg, 1982; Moller, 1986; Beukema, 1991).
There are two standard approaches to caging experiments, enclosure and exclosure, and there is some debate as to which is preferable (Hall et al., 1990a; Wilson, 1991;
´
Olafsson et al., 1994). Given the limitations of both enclosures and exclosures, the present study has utilised both designs to enable a clearer picture to be drawn of the effects of predation on the two species of bivalves. As significant results from enclosing Carcinus have only been achieved in the past by using raised levels of the predator, this study has used high but naturally occurring densities. As recently settled bivalves represent by far the greatest proportion of the population a mesh size was employed which would retain the entire population of the two dominant species of bivalves at the present site: Macoma balthica (L.) an infaunal surface deposit-feeding and facultative suspension feeder (Eggleston et al., 1992) and Cerastoderma edule (L.) a suspension-feeder (Reise, 1985). Other enclosure studies which have failed to find a significant reduction of the macrobenthos due to crab predation have used larger mesh sizes which would not have retained all of the juveniles (Thrush, 1986; Frid and James, 1988; Raffaelli et al., 1989). The enclosure experiment, conducted in 1996, enclosed Carcinus,
2
at three different densities viz: zero Carcinus, five Carcinus and 20 Carcinus, in 1-m cages. The exclosure experiment was conducted in the same area in 1997, the three treatments were full cages, partial cages and un-caged areas. The null hypothesis being tested in the enclosure experiment was that Carcinus predation will have no effect on either M. balthica or C. edule densities. The null hypothesis for the exclosure experiment is that predation, by all predators, will have no effect on either species of bivalve.
Routine sampling of the present study site has confirmed temporal and spatial
1 2
variability, both over small (m ) and large (m ) scales, for the two dominant bivalve species (Richards, unpublished data). Carcinus has also been shown to exhibit heterogenous distribution at the study site. The impact crabs have on prey populations has been shown to be influenced by microhabitat characteristics, at other sites (Lipcuis and Hines, 1986; Eggleston et al., 1992; Schratzberger and Warwick, 1998) and at the present site (Fernandes et al., 1999). The Fernandes et al. (1999) study has shown that Carcinus predation at this site is dependent on sediment type. The non-uniform distribution of Carcinus found at the present site, together with this sediment-dependent predation, could be a cause of the variability in the populations of its molluscan prey. The main aim of the present study was to investigate whether naturally occurring densities of Carcinus could significantly reduce bivalve densities. We discuss our results in the context of the spatial and temporal variability of the bivalves found at this site.
2. Materials and methods
2.1. Study site
Fig. 1. The Forth Estuary, showing the location of the study site.
The area is relatively unpolluted, being over 8 km downstream of industrial effluents (McLusky et al., 1983). The mudflat is approximately 650 m from MHWS to MLWS. Midtidal level was chosen for both caging experiments, as regular sampling of the site had shown this to be the area where the highest density of juvenile bivalves occurred. This area is classed as a muddy site with a carbon level of 6.4% and a silt fraction, ,63
mm, of 29% (Fernandes et al., 1999).
2.2. Bivalve densities
Routine sampling of the study site was conducted from March 1996 until October 1997 (Richards, unpublished data). This consisted of either three or five cores (10310 cm) being taken from within each of five stations on a transect from low to high water. All individual bivalves found were identified, counted and measured on each sampling occasion.
2.3. Crab densities
2
Routine sampling at the study site had revealed a maximum crab density of 5 m (Richards, unpublished data). This sampling was supplemented at the end of August
2
2.4. Enclosure
2
The 1-m cages were constructed by stapling 3-mm mesh plastic netting to four wooden corner posts. Similar size mesh was used for the tops. Plywood boards, 10 cm wide, were nailed around the bottom of the cages, the netting was then attached to these boards. When the cages were placed in position they were sunk into the mud up to the top of the boards, preventing any crabs from escaping under the cages (Raffaelli et al., 1989). The height of the cages above the sediment was 20 cm. Each cage within a block was randomly assigned to one of three initial crab densities: 0 (control), five or 20 Carcinus. All Carcinus were caught in the immediate area. Cages were placed in a randomised block design, three cages to a block with five replicates. Blocks were separated by 6 m and within blocks cages were laid out in a staggered line approximately 1 m apart (Fig. 2).
Only Carcinus with a carapace width between 15 and 25 mm were used in the enclosures as crabs of this size show little tidal migration and stay in the intertidal area (Warman et al., 1993; Mathieson and Berry, 1997), therefore reducing the effect of behavioural changes which occur when enclosing a predator (Gee et al., 1985; Hall et al., 1990a). Care was taken not to include any Carcinus with a soft carapace or damaged chaela. However, they were not sexed as crabs of this size show no evidence of sex-related dietary differences (Raffaelli et al., 1989). Any Carcinus seen in the cages at the start of the experiment were removed, but no attempt was made to physically search the sediment as this would have disturbed the bivalves. Three 2.5 cm diameter cores were taken from within the cages for sediment analysis. Carcinus were added and netting was stapled over the tops. The experiment ran from 29 / 07 / 96 until 29 / 08 / 96. This period was chosen as regular sampling had shown that settlement of both bivalves and large epibenthic predators had ceased before the experiment commenced. Settlement for M. balthica and C. edule had begun in June and had peaked by August, Carcinus had settled onto the mudflat at the end of May (Richards, unpublished data). The 3-mm mesh size was sufficiently small to exclude any previously settled young of these predators.
2.5. Exclosure
The exclosure experiment consisted of three treatments, viz. complete (full) cages, the same design as for the enclosure experiment; partial cages using a similar design but with a 10-cm strip of netting removed around the bottom of the sides; and an un-caged area of the same size. The full cages acted as a total exclosure for large mobile epibenthic predators. The partial cages, with the bottom half of the sides removed, were used to mimic the hydrodynamic effects of the complete cages but to allow access for the epibenthic predators. The un-caged area acted as a control for the enclosed areas. There were three treatments per block, the experimental layout was the same as for the enclosure experiment (Fig. 2) except each block was replicated eight times. Three 2.5-cm diameter cores were taken for sediment analysis. This experiment ran from 26 / 08 / 97 until 24 / 10 / 97. This experiment started slightly later than the enclosure to reduce any possible confounding artefacts due to relocation of juvenile bivalves (Hewitt et al., 1997).
During the course of both experiments, regular visits were made to the cages, any algae or other material which was likely to foul the cages was removed and any damage to the cages repaired. At the completion of the experiment, four 10310 cm cores were taken to a depth of 7 cm, from randomly selected points within each of the cage replicates of the enclosure experiment, and three from each cage replicate of the exclosure experiment, for faunal analysis. To avoid edge effects no cores were taken from within 20 cm of the edge of cages (Raffaelli and Milne, 1987). Three 2.5-cm diameter cores were taken from each cage to a depth of 4 cm for sediment analysis. These were pooled for particle size analysis. Each enclosure cage was systematically searched for surviving predators, which were preserved for later counting, measuring and gut contents analysis to look for the presence or absence of bivalves. Each of the faunal cores was split into two depths. The top 1 cm was sieved through a 212-mm sieve and the next 6 cm sieved through a 500-mm sieve. Earlier work had shown that these sieve sizes would retain all of the bivalves at these two depths and also that none of the dominant bivalve species occurred deeper than 7 cm at the study site (Richards, unpublished data). All of the bivalves retained were preserved in 70% alcohol. Bivalves were identified and measured to the nearest 0.1 mm to look for any effect on prey size (Jaquet and Raffaelli, 1989).
2.6. Data analyses
species. Paired t-tests (or non-parametric equivalent) were used to test for differences in percentage silt content between the start and end of the experiment. One-way ANOVA was used to test for differences in percentage silt between treatments at the end of each experiment. Significance levels of statistical tests were set at p50.05, except for the chi-square tests which were modified by the Bonferroni procedure (Underwood, 1997, pp. 138–139) giving an a level of 0.017.
3. Results
3.1. Bivalve densities and variability
Routine sampling adjacent to the experimental work showed a peak abundance was
2
reached for both species in August. The mean per 100 cm core (6S.E.) in August for M. balthica was 101.4 (614.5), n59. The maximum recorded per core was 175, the minimum 48. For C. edule the mean was 15.7 (61.66), n59, maximum 26, minimum 11. Mean densities decreased afterwards and in November the mean for M. balthica was 22.2 (63.83) n510, maximum 43, minimum 3. C. edule mean was 1.6 (60.65), n510, maximum 7, minimum 0.
3.2. Carcinus density and variability
Carcinus, in the size range 15–25 mm, were present on the mudflat at the study site
22
with a mean (6S.E.) density of 2 per m (60.47), n510. The maximum was five and the minimum 0.
3.3. Enclosure
3.3.1. Survivorship of crabs
Table 1
Gut contents analysis of Carcinus recovered from enclosure experiment Five Carcinus 20 Carcinus
(a) Food present 77 59
(b) C. edule present 77 58
M. balthica present 54 37
a
Percentage of Carcinus with food present (n572). b
Percentage of Carcinus, from all those with food present (n551), in which the two species of bivalves were found.
3.3.2. Sediment analysis
Paired t-tests showed significant increases for percentage silt during the course of the experiment in the control and five Carcinus treatments ( p ,0.05). One-way ANOVA showed no significant differences between treatments at the end of the experiment.
3.3.3. Bivalve totals
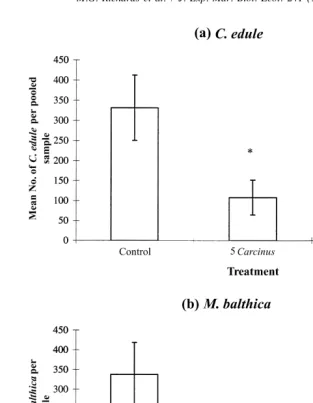
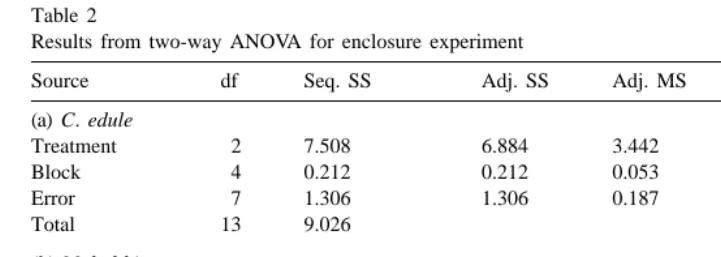
The only bivalve species present in large enough numbers for statistical analysis were M. balthica and C. edule. Treatment means (695% CI) show that the trend for both C. edule (Fig. 3a) and M. balthica (Fig. 3b) was lower densities recovered as Carcinus densities increased. Treatment had a significant effect on both C. edule and M. balthica (Table 2). Multiple comparison Tukey tests showed significant effects between all of the pairs of treatments for C. edule, and between the control and 20 Carcinus treatments for M. balthica.
3.3.4. Size frequency
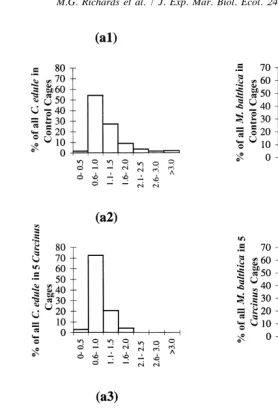
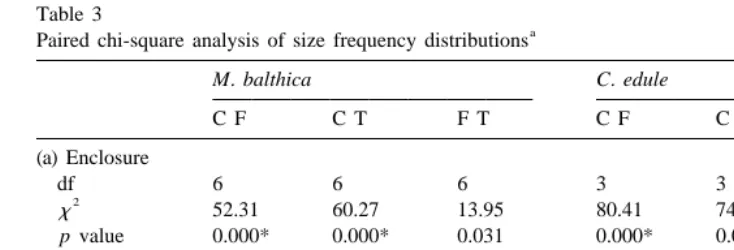
Fig. 4 shows size frequency distributions for C. edule (a1–a3) and M. balthica (b1–b3) expressed as percentages of the treatment total occurring within seven separate size class. Only a few percent of C. edule above 1.5 mm remained in the five and 20 Carcinus treatments. Because of these low numbers all C. edule above 1.5 mm had to be grouped into one size class for chi-square analysis. Chi-square tests, between treatments, on the size frequency distributions showed significant differences between the control and five Carcinus treatments, and between control and 20 Carcinus treatments for both species (Table 3a). The contingency tables indicated that the greatest differences between treatments occurred in the largest size class, i.e. less than expected in the greater than 3 mm size class for M. balthica; and less than expected in the greater than 1.5 mm for C. edule.
3.4. Exclosure
3.4.1. Sediment analysis
Fig. 3. Mean number, per pooled sample, of individuals recovered at end of enclosure experiment(695% CI). Pooled sample is total of four cores. (a) C. edule, (b) M. balthica. Significant Tukey test results are indicated by: * Significantly different from control treatments, ** Significantly different from five Carcinus treatments.
3.4.2. Bivalve totals
Table 2
Results from two-way ANOVA for enclosure experiment
Source df Seq. SS Adj. SS Adj. MS F p
(a) C. edule
Treatment 2 7.508 6.884 3.442 18.45 ,0.01
Block 4 0.212 0.212 0.053
all treatments for C. edule, and between the full cages and un-caged control treatments for M. balthica.
3.4.3. Size frequency
Percentage size frequency distributions (Fig. 6) are shown for C. edule (a1–a3) and M. balthica (b1–b3). No C. edule larger than 1 mm length were recorded from the partial or un-caged controls. No chi-square test was performed for C. edule due to too few size classes and a low number of observations in the control areas. Chi-square tests for M. balthica showed significant differences between full cages and partial cages, and between full cages and un-caged control (Table 3b). The greater than 3-mm size class accounted for the greatest variability between these distributions. There was no significant difference between partial cages and un-caged control.
4. Discussion
The overall trend in both experiments was for lower densities of C. edule to be recovered as predation pressure increased. All pairwise comparisons were significantly
22
Fig. 4. Size frequency distribution of C. edule (a1–a3) and M. balthica (b1–b3) at end of enclosure experiment. Expressed as percentages of the treatment total occurring within seven separate size classes for either C. edule (a1–a3) or M. balthica (b1–b3).
Table 3
a Paired chi-square analysis of size frequency distributions
M. balthica C. edule
C F C T F T C F C T F T
(a) Enclosure
df 6 6 6 3 3 3
2
x 52.31 60.27 13.95 80.41 74.54 7.3
p value 0.000* 0.000* 0.031 0.000* 0.000* 0.064
(b) Exclosure C CC C UCC CC UCC
Treatment abbreviations: C, control; F, five Carcinus; T, 20 Carcinus; C, cage; CC, cage control; UCC, un-caged control.
*
Significant p values (a 50.017).
tidal flats during autumn (Beukema, 1992), therefore reducing its impact on the benthos. No protection from predation in the un-caged areas led to almost no survival for C. edule. This concurs with the laboratory study by Sanchez-Salazar et al. (1987), who concluded that size-selective predation exerted by Carcinus resulted in almost 100% mortality of C. edule below 15 mm length. The data from the transect work also supports these results. C. edule were reduced from a mean of 15.7 per core in August to less than 2 in November, with several cores containing no C. edule.
The overall trend for M. balthica was similar to that of C. edule in both experiments. However, significant differences were found only between the full cages and un-caged controls in the exclosure study and between the control and 20 Carcinus treatments in the enclosure study. One explanation for why the other comparisons were not significant is that cages which contained no predators may have acted as refuges for predatory infauna (Ambrose, 1984), which have been shown to have a negative effect on juvenile
¨
bivalve survivorship (Ratcliffe et al., 1981; Reise, 1985; Ronn et al., 1988). These effects can reduce the bivalve densities in control treatments and mask any differences between the treatments. As all comparisons were significant for C. edule this indicated that any effect of predation by infaunal predators in the control cages was smaller than the effect of predation by Carcinus. Comparisons for M. balthica between the control and five Carcinus treatments approached significance (Fig. 3), and it is possible that epibenthic predators finding refuge from their own predators had influenced this result. Of course the inclusion of one cage containing two Carcinus in the control treatment will have affected this result. The inclusion of this replicate means that our interpretation of Carcinus predation is conservative. Although M. balthica numbers were reduced in partial cages when compared to full cages in the exclosure experiment, this difference was non-significant. It is only when the full suite of predators (including large fish and birds) were allowed to prey on an un-caged area that a significant difference was observed between treatments.
Fig. 5. Mean number, per pooled sample, of individuals recovered at end of enclosure experiment(695% CI). Pooled sample is total of three cores. (a) C. edule, (b) M. balthica. Significant Tukey test results are indicated by: * Significantly different from full cage treatments, ** Significantly different from partial cage treatments.
predator can account for most of the temporal variability recorded in the routine sampling, especially for C. edule. Areas with high densities of Carcinus may account for the temporal variability shown for M. balthica but it may require all predators to account for all of the reduction.
Most crab enclosure studies that have shown significant differences have only achieved these at raised predator densities and only one study (Hoffman et al., 1984) has
´ ´
Table 4
Results from two-way ANOVA for exclosure experiment
Source df SS MS F ratio p
(a) C. edule
Treatment 2 5.5498 2.7749 62.5 ,0.001
Block 7 0.3793 0.0542
Error 14 0.6291 0.0444
Total 23 6.5509
(b) M. balthica
Treatment 2 12 803 6402 6.408 ,0.05
Block 7 14 108 2015
Error 14 13 987 999
Total 23 40 899
(1994) definition of a ‘strong effect’ is a two fold difference of a prey item(s) in treatment cages compared to controls. Although the control treatment cannot strictly be called a control, as one cage had two crabs inadvertently enclosed, our conservative interpretation suggest that Carcinus predation at high but realistic densities exhibits a ‘strong effect’ on C. edule densities. The Hoffman et al. (1984) study of the fiddler crab (Uca pugnax) used a small sieve size and showed large reductions in some meiofaunal species. The present study used a mesh size of 212 mm, suitable to retain even the smallest newly settled C. edule (Guillou and Tartu, 1994) and M. balthica (Caddy, 1969). As most other studies enclosing ambient densities of crabs have used a sieve size of 500 mm (e.g. Thrush, 1986; Raffaelli et al., 1989; Hall et al., 1990a) and have reported mostly non-significant effects, it is possible that the effect of predation has been underestimated. Size selective predation was evident from both of the caging experi-ments for both species of bivalve. Fewer large C. edule were recovered from cages with five or 20 Carcinus. The exclosure experiment showed similar changes with lower percentages of larger C. edule found when the partial cages and un-caged areas are compared to the full cages. This indicates that predators are preferentially feeding on the largest C. edule. Chi-square tests on the M. balthica populations also indicate that the larger individuals are being taken by Carcinus in the enclosure cages and by Carcinus and other predators in the partial cages and un-caged areas. The results from this study show that, although all size classes have been affected, the main impact was on the largest bivalves. Therefore the studies using the larger size sieves may have correctly estimated the effect of crab predation, but only by using smaller mesh sizes could this be confirmed.
Fig. 6. Size frequency distribution of C. edule (a1–a3) and M. balthica (b1–b3) at end of exclosure experiment. Expressed as percentages of the treatment total occurring within seven separate size classes for either C. edule (a1–a3) or M. balthica (b1–b3).
to distinguish between predation and disturbance caused by the predators (Schratzberger and Warwick, 1998). If hydrodynamic and disturbance artefacts had been more important than predation in the current work then the size frequency distributions would show the largest effects on the smaller bivalves, which are closer to the surface and more
¨
Although the main settling period of both species of bivalves was avoided, movement of prey into and out of cages may affect the results of caging experiments (Thrush, 1986; Raffaelli and Milne, 1987; Jaquet and Raffaelli, 1989; Hall et al., 1990a,b; Englund, 1997). The mobility of prey items may occur at the scale of the cage (Hall et al., 1990a,b or over larger scales (Hewitt et al., 1997). Although prey movement may have occurred at the scale of the cage it is more likely to have occurred over a broader
¨
scale due to hydrodynamic effects. Gunther (1991) and Armonies and Hellwig-Armonies (1992) have shown a net transport of bivalves from initially settled areas (midtidal level at our study site) to higher intertidal sites. If this movement occurred during the course of the experiments it would have confounded the impacts of predation, leading to a type II error (Englund, 1997; Fernandes et al., 1999). Hence, if prey movement on a broader scale than that of the cage, was a factor in the present study, our interpretation of the impacts of crab predation would be conservative. One of the weak points in the present study is that the two experiments were conducted over different time scales in different years. Due to the interannual variation shown in bivalve populations (Bachelet, 1986), no direct comparison can be made between the studies. However, the use of both enclosure and exclosures together with gut contents analysis, has given strong evidence that both species of bivalves are reduced even at naturally occurring densities Carcinus.
Holland et al. (1980) and Arnold (1984) have shown that crab predation on bivalves varies with habitat, and differences in the foraging efficiency of the blue crab (Callinectes sapidus) between muddy and sandy sediments has been invoked to explain spatial variability in Mya arenaria and M. balthica (Eggleston et al., 1992). The bivalve routine sampling incorporated a sandy station approximately 100 m away from the experimental area. Numbers of C. edule were on average three times higher in the sandy area when compared to the muddy site. This may be due to C. edule being a filter feeding, which may clog up in muddier areas, passive or active distribution (Armonies and Hellwig-Armonies, 1992; Armonies, 1996; Hewitt et al., 1997), or, at least in part, to different foraging efficiency in sandier areas (Fernandes et al., 1999). The differential predation between habitats together with the spatial variability exhibited by Carcinus may explain the spatial variability shown by C. edule, and possibly M. balthica at this site. Therefore we conclude that predation, especially by Carcinus, can significantly reduce bivalve densities and may act as a causal mechanism for both the spatial and temporal variability in intertidal bivalves.
Acknowledgements
We thank Michaela Schratzberger, Teresa Fernandes and two anonymous referees for suggestions and corrections of the draft manuscript.
References
Armonies, W., 1996. Changes in distribution patterns of 0-group bivalves in the Wadden Sea: Byssus-drifting releases juveniles from the constraints of hydrography. J. Sea Res. 35, 323–334.
Armonies, W., Hellwig-Armonies, M., 1992. Passive settlement of Macoma balthica spat on tidal flats of the Wadden Sea and subsequent migration of juveniles. Neth. J. Sea Res. 29, 371–378.
Arnold, W.S., 1984. The effects of prey size, predator size, and sediment composition on the rate of predation of the blue crab, Callinectes sapidus (Rathbun), on the hard clam, Mercineria mercineria (Linne). J. Exp. Mar. Biol. Ecol. 80, 207–219.
Bachelet, G., 1986. Recruitment and year-to-year variability in a population of Macoma balthica (L.). Hydrobiologia 142, 233–248.
Beukema, J.J., 1991. The abundance of shore crabs Carcinus maenas (L.) on a tidal flat in the Wadden Sea after cold and mild winters. J. Exp. Mar. Biol. Ecol. 153, 97–113.
Beukema, J.J., 1992. Dynamics of juvenile shrimp Crangon crangon in a tidal-flat nursery of the Wadden Sea after mild and cold winters. Mar. Ecol. Prog. Ser. 83, 157–165.
Butman, C.A., 1987. Larval settlement of soft-sediment invertebrates: the spatial scales of pattern explained by ˆ
active habitat selection and the emerging role of hydrodynamical processes. Oceanogr. Mar. Biol. Annu. Rev. 25, 113–165.
Caddy, J.F., 1969. Development of mantle organs, feeding, and locomotion in postlarval Macoma balthica (L.) (Lamellibranchiata). Can. J. Zool. 47, 609–617.
Eggleston, D.B., Lipcius, R.N., Hines, A.H., 1992. Density-dependent predation by blue crabs upon infaunal clam species and abundance patterns. Mar. Ecol. Prog. Ser. 85, 55–68.
Englund, G., 1997. Importance of spatial scale and prey movements in predator caging experiments. Ecology 78, 2316–2325.
Fernandes, T., Huxham, M., Piper, S., 1999. Predator caging experiments: A test of the importance of scale. J. Exp. Mar. Biol. Ecol. (in press).
Frid, C.L.J., 1989. The role of recolonization processes in benthic communities, with special reference to the interpretation of predator-induced effects. J. Exp. Mar. Biol. Ecol. 126, 163–171.
Frid, C.L.J., James, R., 1988. The role of epibenthic predators in structuring the marine invertebrate community of a British coastal salt marsh. Neth. J. Sea Res. 22, 307–314.
Gee, J.M., Warwick, R.M., Davey, J.T., George, C.L., 1985. Field experiments on the role of epibenthic predators in determining prey densities in an estuarine mudflat. Estuar. Coast. Shelf Sci. 21, 439–448. Guillou, J., Tartu, C., 1994. Post-larval and juvenile mortality in a population of the edible cockle
Cerastoderma edule (L.) from northern Brittany. Neth. J. Sea Res. 33, 103–111.
¨
Gunther, C.-P., 1991. Settlement of Macoma balthica on an intertidal sandflat in the Wadden Sea. Mar. Ecol. Prog. Ser. 76, 73–79.
Hall, S.J., Raffaelli, D., Turrell, W.R., 1990a. Predator-caging experiments in marine systems: A reexamination of their value. Am. Nat. 136, 657–672.
Hall, S.J., Raffaelli, D., Robertson, M.R., Basford, D.J., 1990b. The role of the predatory crab, LioCarcinus
depurator, in a marine food web. J. Anim. Ecol. 59, 421–438.
Hewitt, J.E., Pridmore, R.D., Thrush, S.F., Cummings, V.J., 1997. Assessing the short-term stability of spatial patterns of macrobenthos in a dynamic estuarine system. Limnol. Oceanogr. 42, 282–288.
Hines, A.H., Whitlatch, R.B., Thrush, S.F., Hewitt, J.E., Cummings, V.J., Dayton, P.K., Legendre, P., 1997. Non-linear foraging response of a large marine predator to benthic prey: Eagle ray pits and bivalves in a New Zealand sandflat. J. Exp. Mar. Biol. Ecol. 216, 191–210.
Hoffman, J.A., Katz, J., Bertness, M.D., 1984. Fiddler crab deposit-feeding and meiofaunal abundance in salt marsh habitats. J. Exp. Mar. Biol. Ecol. 82, 161–174.
Holland, A.F., Mounford, N.K., Hiegel, M.H., Kaumeyer, K.R., Mihursky, J.A., 1980. Influence of predation on infaunal abundance in Upper Chesapeake Bay, USA. Mar. Biol. 57, 221–235.
Hurlbert, S.H., 1984. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 54, 117–229.
Jaquet, N., Raffaelli, D., 1989. The ecological importance of the sand goby Pomatoschistus minutus (Pallas). J. Exp. Mar. Biol. Ecol. 128, 147–156.
Jensen, K.T., Jensen, J.N., 1985. The importance of some epibenthic predators on the density of juvenile benthic macrofauna in the Danish Wadden Sea. J. Exp. Mar. Biol. Ecol. 89, 157–174.
Lipcuis, R.N., Hines, A.H., 1986. Variable functional responses for a marine predator in dissimilar homogeneous microhabitats. Ecology 67, 1361–1371.
Lindegarth, M., 1996. Spatial population structure of bivalves in shallow sediments: Hydrodynamic effects on ¨
recruitment processes. PhD thesis, Goteborg University, Sweden.
Mathieson, S., Berry, A.J., 1997. Spatial temporal and tidal variation in crab populations in the Forth Estuary, Scotland. J. Mar. Biol. Assoc. UK 77, 167–183.
McLusky, D.S., Anderson, F.E., Wolfe-Murphy, S., 1983. Distribution and population recovery of Arenicola
marina and other benthic fauna after bait digging. Mar. Ecol. Progr. Ser. 11, 173–179.
¨
Moller, P., 1986. Physical factors and biological interactions regulating infauna in shallow boreal areas. Mar. Ecol. Prog. Ser. 30, 33–47.
´
Olafsson, E.B., Peterson, C.H., Ambrose, W.G., 1994. Does recruitment limitation structure populations and communities of macro-invertebrates in marine soft sediments: The relative significance of pre- and post-settlement processes. Oceanogr. Mar. Biol. Annu. Rev. 32, 65–109.
Pihl, L., Rosenberg, R., 1982. Production, abundance, and biomass of mobile epibenthic marine fauna in shallow waters, western Sweden. J. Exp. Mar. Biol. Ecol. 57, 273–301.
Quammen, M.L., 1984. Predation by shorebirds, fish and crabs on invertebrates in intertidal mudflats: an experimental test. Ecology 65, 529–537.
Raffaelli, D., Milne, H., 1987. An experimental investigation of the effects of shorebird and flatfish predation on estuarine invertebrates. Estuar. Coast. Shelf Sci. 24, 1–13.
Raffaelli, D., Conacher, A., McLachlan, H., Emes, C., 1989. The role of epibenthic crustacean predators in an estuarine food web. Estuar. Coast. Shelf Sci. 28, 149–160.
Ratcliffe, P.J., Jones, N.V., Walters, N.J., 1981. The survival of Macoma balthica (L.) in mobile sediments. In: Jones, N.V., Wolff, W.J. (Eds.), Feeding and Survival Strategies of Estuarine Organisms, Plenum Press, New York, pp. 91–107.
Reise, K. (Ed.), 1985. Tidal Flat Ecology. an Experimental Approach To Species Interactions, Springer, Berlin, p. 191.
¨
Ronn, C., Bonsdorff, E., Nelson, W.G., 1988. Predation as a mechanism of interference within infauna in shallow brackish water soft bottoms; experiments with an infaunal predator, Nereis diversicolor O.F.
¨
Muller. J. Exp. Mar. Biol. Ecol. 116, 143–157.
Sanchez-Salazar, M.E., Griffiths, C.L., Seed, R., 1987. The effect of size and temperature on the predation of cockles Cerastoderma edule (L.) by the shore crab Carcinus maenas (L.). J. Exp. Mar. Biol. Ecol. 111, 181–193.
Scherer, B., Reise, K., 1981. Significant predation on micro- and macrobenthos by the crab Carcinus maenas (L.) in the Wadden Sea. Kierler Meeresforsch. Sonerh. 5, 490–500.
Schratzberger, M., Warwick, R.M., 1998. Impact of predation and sediment disturbance by Carcinus maenas (L.) on free-living nematode community structure. J. Exp. Mar. Biol. Ecol. (in press).
Skilleter, G.A., 1994. Refuges from predation and the persistence of estuarine clam populations. Mar. Ecol. Prog. Ser. 109, 29–42.
Thrush, S.F., 1986. Community structure on the floor of a sea-lough: are large epibenthic predators important? J. Exp. Mar. Biol. Ecol. 104, 171–183.
Thrush, S.F., Pridmore, R.D., Hewitt, J.E., Cummings, V., 1994. The importance of predators on a sandflat: interplay between seasonal changes in prey densities and predator effects. Mar. Ecol. Prog. Ser. 107, 211–222.
Thrush, S.F., Hewitt, J.E., Pridmore, R.D., Cummings, V., 1996. Adult / juvenile interactions of infaunal bivalves: Contrasting outcomes in different habitats. Mar. Ecol. Prog. Ser. 132, 83–92.
Thrush, S.F., Cummings, V.J., Dayton, P.K., Ford, R., Grant, J., Hewitt, J.E., Hines, A.H., Lawrie, S.M., Pridmore, R.D., Legendre, P., McArdle, B.H., Schneider, S.J., Whitlatch, R.B., Wilkinson, M.R., 1997. Matching the outcome of small-scale density manipulation experiments with larger scale patterns an example of bivalve adult / juvenile interactions. J. Exp. Mar. Biol. Ecol. 216, 153–169.
Underwood, A.J. (Ed.), 1997. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance, Cambridge University Press, p. 504.
Warman, C.G., Reid, D.G., Naylor, E., 1993. Variation in the tidal migratory behaviour and rhythmic light-responsiveness in the shore crab, Carcinus maenas. J. Exp. Mar. Biol. Ecol. 73, 355–364. Wilson, W.H., 1991. Competition and predation in marine soft-sediment communities. Annu. Rev. Ecol. Syst.
21, 221–241.