BAB I PENDAHULUAN 1.1. Latar Belakang
Sebagian wilayah di Provinsi Bali terutama di bagian Utara dan Timur (Kabupaten Buleleng Barat dan sebagian Karangasem) tergolong lahan kering (Antara, 2004 ; Daryana, 2010). Para petani di daerah tersebut menggunakan sistem pertanian yang sangat bergantung pada musim (Aditya, 2011). Pada musim penghujan para petani menanam tanaman jenis kacang-kacangan (Kacang Tanah, Kacang Hijau, Kacang Panjang, Kacang Undis, Kacang Koma), Cabai dan Jagung. Pada musim kemarau petani mengandalkan tanaman Singkong atau bahkan untuk lahan yang sangat kering tidak ditanami tanaman pertanian sama sekali (Arsana et al., 2010). Kondisi tanah pertanian yang tersebut diatas merupakan faktor utama yang menyebabkan hasil pertaniannya kurang produktif (Suprapto et al., 2001).
Salah satu jenis tanaman yang dikembangkan di lahan kering adalah mente. (A. occidentale L). Tanaman mente merupakan salah satu komoditas ekspor yang ekonomis dan mampu menghasilkan produksi biji (gelondong) mente yang relatif stabil harganya sehingga status tanaman mente yang sebelumnya merupakan tanaman penghijauan beralih menjadi tanaman komoditas unggulan (Zaubin dan Daras, 2002). Salah satu usaha untuk menstabilkan produktifitas tanaman mente diperlukan suatu pengembangan berorientasi agribisnis agar dapat memberikan dampak positif kepada para petani dan industri (Hadad et al., 2000). Kendala yang dihadapi para petani mente di Bali dan Nusa Tenggara adalah hasil produksi buah mente tidak stabil karena petani cenderung
mempertahankan pohon mente yang sudah berumur tua tanpa melakukan peremajaan pohon induk (Suharto, 2011). Dinas Perkebunan Provinsi Bali selama ini melakukan pembibitan mente dengan para petani plasma. Berdasarkan hasil survey di lapangan terdapat beberapa kendala yang dihadapi para petani mente dalam melakukan pembibitan mente antara lain: akar bibit mente mudah patah pada waktu pemindahan dari kebun bibit ke areal perkebunan, bibit mudah mati bila sudah dipindahkan ke areal perkebunan, pertumbuhan bibit yang tidak seragam, jumlah perakaran (serabut) yang dihasilkan bibit mente sangat sedikit sehingga menyebabkan kemampuan absorbsi hara dan mineral dari tanah rendah (Sukawidana, 2010 ; Darmais, 2011 pers.com). Kendala-kendala tersebut pada akhirnya menyebabkan produktivitas mente di Bali dan Nusatenggara, khususnya di lahan kering perkebunan mente Desa Sendang Kecamatan Gerokgak Kabupaten Buleleng dan Desa Sukadana Kecamatan Kubu Kabupaten Karangasem khususnya menjadi tidak/kurang maksimum.
Salah satu upaya untuk mengatasi permasalahan tanaman pertanian khususnya mente di lahan kering tersebut adalah penggunaan cendawan endomikoriza. Spora-spora cendawan endomikoriza yang diinokulasikan pada akar tanaman inang pada fase pembibitan diharapkan dapat mengatasi salah satu kendala pertumbuhan mente di lahan kering dengan asumsi bibit yang telah diinokulasi dengan cendawan endomikoriza pada fase pembibitan, bibit mente tersebut akan terkolonisasi oleh cendawan endomikoriza sehingga terbantu dalam absorbsi air dari tanah dan terlindungi dari serangan penyakit sejak dipembibitan sampai di areal perkebunan. Menurut Smith et al. (2010),
simbiosis cendawan endomikoriza dengan akar tanaman dapat meningkatkan absorbsi air dan mineral seperti P, N, K, Cu, Mo, Zn, menstimulasi pertumbuhan, meningkatkan resistensi tanaman terhadap kekeringan dan serangan / infeksi mikroba patogen di tanah.
Penelitian tentang peran penting endomikoriza telah banyak dilakukan antara lain; inokulasi cendawan endomikoriza untuk meningkatkan laju pertumbuhan dan produktivitas tanaman jagung (Zea mays) di lahan kritis (Idwar dan Ali. 2000); Inokulasi spora dan propagul endomikoriza Acaulospora tuberculata dan Gigaspora margarita untuk meningkatkan serapan hara pada bibit kelapa sawit yang dibudidayakan di tanah masam (Widiastuti et al., 2002); Inokulasi spora endomikoriza untuk memacu proses reproduksi tanaman Barley (Hordeum vulgare L.) (Therrien, 2008); Penggunaan endomikoriza untuk meningkatkan produksi kedelai di lahan kering (Hapsoh, 2008); penambahan endomikoriza Acaulospora sp. untuk meningkatkan produksi umbi pada tanaman ubi kayu (Jauhari dan Sumarno, 1995); penambahan spora Glomus sp. untuk meningkatkan toleransi tanaman hias yang ditanam pada tanah pertanian pasca bioremediasi (Al-Zalzaleh et al., 2009). Hasil-hasil penelitian oleh Cruzz et al. (2000) ; Hameeda et al. (2007) dan Douds et al. (2010) mampu membuktikan bahwa simbiosis cendawan mikoriza dengan tanaman dapat mengurangi ketergantungan tanaman tersebut pada pupuk dan pestisida sintetik sehingga penggunaan mikoriza merupakan salah satu upaya untuk mengurangi ketergantungan terhadap pupuk ataupun pestisida kimia.
Penggunaan endomikoriza komersial sudah diaplikasikan pada tanaman pertanian di Bali meskipun masih dalam taraf skala uji coba (pilot project),
misalnya: pemberiaan pupuk mikoriza komersial “Biosfer” pada pembibitan tanaman Vanili (Tirta, 2006); kombinasi mikoriza komersial dan cendawan Trichoderma sp untuk mengurangi infeksi Jamur Akar Putih (JAP) pada tanaman Mente (Priono, 2008); Aplikasi pupuk mikoriza “Technofert” untuk meningkatkan pertumbuhan tanaman pakan ternak (Sukasta et al., 2010). Penelitian aplikasi mikoriza komersial pada tanaman pertanian dan perkebunan di Bali telah banyak dilakukan, namun penggunaan mikoriza komersial tersebut belum terlihat hasilsecara nyata dalam meningkatkan pertumbuhan dan produktifitas tanaman (pertanian dan perkebunan) ataupun menurunkan intensitas infeksi tanaman yang terinfeksi oleh mikroba patogen tersebut. Kurang berhasilnya kemampuan endomikoriza komersial tersebut untuk aplikasi tanaman pertanian atau perkebunan diduga bahwa endomikoriza komersial yang berasal dari “pabrik” atau “ Balai penelitian” dengan sumber inokulum bukan berasal dari daerah setempat sehingga diasumsikan akan memerlukan waktu adaptasi lebih lama terhadap kondisi lingkungan yang baru.
Menurut Bohrer dan Amon (2004); Thamsurakul dan Charoensook (2006) dan Suryatmana et al. (2009), mikoriza indigenus memiliki potensi yang tinggi untuk dijadikan sebagai “pupuk hayati” atau “Biofertilizer” tanaman pertanian di lahan kering daerah setempat karena jenis-jenis endomikoriza indigenus lebih adaptif sehingga hifa dan spora cendawan tersebut dapat dengan cepat bergerminasi dan mengkolonisasi sistem perakaran tanaman disekitarnya khususnya tanaman yang merupakan inang dari endomikoriza tersebut.
Penelitian ekplorasi endomikoriza di alam diperlukan faktor lingkungan yang sehingga dapat diketahui jenis-jenis dan kepadatan populasi organisme
tersebut pada musim dan tempat yang berbeda (Begon et al., 1990 ; Brundrett et al., 1996). Penelitian Bohrer et.al. (2001) yang dilakukan di Boswana mengindikasikan bahwa kepadatan atau kerapatan jumlah spora endomikoriza di alam sangat dipengaruhi oleh perbedaan tempat, musim dan curah hujan. Hasil penelitian Danesh et al. (2007) di Iran mengindikasikan bahwa keanekaragaman endomikoriza pada suatu kawasan sangat dipengaruhi porositas dan tingkat kejenuhan tanah. Hal ini menjadi suatu alur pemikiran bahwa untuk mengetahui kerapatan jumlah spora, kemampuan kolonisasi endomikoriza pada tanaman inangnya dan jenis-jenis endomikoriza di Bali, penelitian eksplorasi perlu dilakukan dalam satu siklus musim dan tempat yang berbeda sehingga akan dapat diketahui apakah perbedaan tempat dan musim akan berpengaruh terhadap keberadaan jumlah spora, persen kolonisasi pada tanaman inang dan jenis endomikoriza tersebut di alam.
Kendala pada aplikasi endomikoriza di lapangan adalah keterbatasan akan ketersediaan isolat mikoriza dalam bentuk pupuk hayati dan merupakan masalah yang belum dapat terselesaikan sehingga kalangan petani sulit mendapatkan pupuk hayati mikoriza tersebut. Pupuk hayati dapat dibeli atau disediakan oleh produsen tetapi harganya mahal sehingga aplikasi di kalangan petani sulit dilakukan (Aryantha et al., 2002; Sukasta et al., 2010).
Berdasarkan uraian tersebut diatas, diperlukan suatu penelitian bagaimana caranya memperbanyak endomikoriza indigenus Bali (spora dan atau propagul) sehingga endomikoriza hasil eksplorasi tersebut dapat diaplikasikan atau siap digunakan sebagai pupuk hayati yang dapat digunakan langsung oleh kalangan petani, khususnya pada tahap pembibitan.
Perbanyakan spora dan propagul dipengaruhi oleh banyak faktor baik faktor internal (kemampuan germinasi dari spesies endomikoriza) dan faktor eksternal (bahan organik tanah dan ketersediaan hara). Penambahan pupuk sebagai sumber hara dan mineral sangat diperlukan dalam perbanyakan spora endomikoriza terutama untuk pertumbuhan akar tanaman inang (Novriani dan Madjid, 2009Chalimah, 2007; Lukiwati, 2007). Salah satu sumber hara yang standar digunakan sebagai pupuk untuk perbanyakan atau propagsi spora endomikoriza adalah larutan Hara Johnson. Hara Johnson mempunyai kandungan unsur hara makro dan mikro yang sudah standard namun larutan ini dapat dimodifikasi konsentrasi hara makro dan mikro didalamnya sehingga hara ini sering digunakan untuk memperbanyak spora dan hifa endomikoriza pada tanaman inang (Widiastuti, 2002; Simanungkalit, 2003). Jenis tanaman inang yang umum digunakan untuk memperbanyak spora adalah tanaman semusim karena cepat tumbuh dan menghasilkan banyak akar serabut dibanding tanaman perenial sehingga perbanyakan endomikoriza tidak membutuhkan waktu lama (Widiastuti, 2004). Tanaman semusim seperti Jagung dan Shorgum merupakan inang sangat kompatibel dengan endomikoriza (Simanungkalit, 2003; Hapsoh, 2008,) sehingga tanaman Jagung dan Shorgum merupakan inang yang digunakan untuk perbanyakan spora endomikoriza (Widiastuti, 2004).
Berdasarkan paparan latar belakang tersebut di atas perlu dilakukan penelitian tentang “Pemanfaatan Endomikoriza Indigenus pada Lahan Kering di Bali untuk Memacu Pertumbuhan Mente (Anacardium occidentale L.) “
1.2. Rumusan Masalah
1. Apa saja spesies endomikoriza dan bagaimana kerapatan spora serta kolonisasi pada tanaman di lahan kering Buleleng dan Karangasem Bali. 2. Berapakah konsentrasi P dalam hara Johnson yang optimum untuk
perbanyakan spora endomikoriza indigenus Bali pada tanaman Jagung? 3. Bagaimanakah respon pertumbuhan bibit mente terhadap inokulasi
endomikoriza dalam bentuk spora tunggal dan campuran spora-propagul? 4. Bagaimanakah respon pertumbuhan bibit mente terhadap spora dan
propagul endomikoriza pada media pembawa yang berbeda?
1.3.Tujuan Penelitian
Tujuan penelitian ini adalah untuk:
1. Mengetahui spesies, kerapatan spora endomikoriza dan persentase kolonisasi endomikoriza indigenus pada tanaman di lahan kering Buleleng dan Karangasem Bali
2. Mengetahui konsentrasi P optimum dalam hara Johnson untuk perbanyakan spora endomikoriza indigenus Bali pada tanaman Jagung
3. Mengetahui respon pertumbuhan bibit mente (A. occidentale L.) terhadap inokulasi endomikoriza indigenus Bali dalam bentuk spora, propagul saja dan campuran spora-propagul
4. Mengetahui pengaruh spora dan propagul endomikoriza dalam media pembawa berbeda terhadap pertumbuhan bibit mente.
1.4. Manfaat Penelitian
Hasil penelitian ini diharapkan dapat menghasilkan temuan yang baru tentang spesies cendawan endomikoriza yang terdapat di lahan kering Bali dan
inokulasi cendawan endomikoriza terhadap respon pertumbuhan bibit mente serta memberikan kontribusi secara akademik (untuk perkembangan ilmu pengetahuan dan teknologi) dan manfaat praktis (dapat digunakan untuk memberikan solusi atau memecahkan masalah pertanian yang terdapat di masyarakat), yaitu:
1.4.1. Manfaat akademik
1. Memberikan informasi spesies apa saja, kerapatan jumlah spora dan kolonisasi endomikoriza indigenus pada tanaman-tanaman di lahan kering Buleleng dan Karangasem Bali
2. Perbanyakan spora endomikoriza pada tanaman Jagung dengan variasi P hara Johnson dapat memberikan informasi tentang propagasi spora.
3. Hasil uji coba endomikoriza pada tanaman mente memberikan informasi tentang penggunaan endomikoriza indigenus Bali untuk memacu pertumbuhan bibit mente.
1.4.2. Manfaat praktis
Hasil penelitian ini diharapkan dapat digunakan sebagai salah satu acuan dalam pembuatan dan pengembangan “pupuk hayati “ berbasis spora dan propagul endomikoriza Indigenus Bali untuk diaplikasikan pada pembibitan mente sehingga dapat meningkatkan kualitas pertumbuhan dan ketahanan hidup bibit tersebut pada lahan kering di Bali.
BAB II
KAJIAN PUSTAKA
2.1. Biologi Cendawan Endomikoriza
Mikoriza berasal dari bahasa Yunani yang secara harfiah berarti “cendawan akar” (mykos = miko= cendawan dan rhiza = akar ) atau “cendawan tanah” karena hifa dan sporanya selalu berada di tanah terutama areal rhizosfer tanaman (Mikola, 1980; Smith and Read, 1997). Asosiasi antara cendawan mikoriza dengan tanaman inang merupakan hubungan simbiosis mutualisme (Simanungkalit, 2003; Brundrett et al., 2008). Simbiosis tersebut bermanfaat bagi keduanya, yaitu cendawan mikoriza memperoleh karbohidrat dalam bentuk gula sederhana (glukosa) dan Karbon (C) dari tumbuhan, sebaliknya cendawan melalui hifa eksternal yang terdistribusi di dalam tanah dapat menyalurkan air, mineral dan hara tanah untuk membantu aktifitas metabolisme tumbuhan inangnya (Brundrett et al., 2008; Smith et al., 2010).
Cendawan mikoriza terutama kelompok endomikoriza mendapat banyak perhatian para peneliti karena beberapa spesies dari cendawan endomikoriza di alam mampu bersimbiosis secara mutualistik dengan lebih dari 80% spesies tanaman berpembuluh (Smith dan Read, 1997 ; Hapsoh, 2008; Brundrett et al., 2008). Simbiosis cendawan mikoriza dapat terjadi secara alami atau dengan cara diinokulasikan pada tanaman inangnya dan proses kolonisasi secara intensif terjadi pada fase pembibitan tanaman tersebut (Alizadeh, 2011). Tanaman yang telah terkolonisasi atau terinfeksi oleh cendawan mikoriza pada fase pembibitan, tanaman tersebut akan membawa hifa ataupun spora cendawan
mikoriza tersebut selama tanaman tersebut tumbuh (Brundrett et al., 2008). Peran simbiosis endomikoriza yang terutama adalah membantu tanaman-tanaman yang terdapat pada tanah atau lahan yang miskin air dan hara (Simanungkalit, 2003 ; Delvian, 2006a), khususnya tanah yang miskin akan kandungan Fosfat, Zink, Nitrogen (Smith et.al., 2010).
Kondisi tanah pertanian atau perkebunan di Indonesia umumnya memiliki kandungan Fosfat yang cukup tinggi (jenuh) (Delvian, 2006b). Kandungan Fosfat yang terdapat di alam seringkali terikat dalam bentuk senyawa pada mineral liat tanah sehingga menyebabkan P yang melimpah ditanah namun tidak tersedia bagi tanaman (Brundrett et al., 2008; Smith et.al., 2010). Cendawan mikoriza adalah salah satu mikroba yang dapat menghasilkan enzim fosfatase sehingga dengan enzim tersebut hifa-hifa cendawan mampu melepaskan ikatan P dari mineral liat pada tanah dan merombak P bentuk ion fosfor sehingga dapat dimanfaatkan bagi tanaman (Novriani dan Madjid, 2009).
Pertumbuhan dan aktivitas endomikoriza di tanah atau rhizosfer tanaman sangat tergantung oleh keberadaan jenis-jenis atau spesies endomikoriza yang terdapat pada areal tersebut, lingkungan yang mendukung pertumbuhan spesies endomikoriza dan tanaman inang yang kompatibel (Widiastuti et al., 2002 ; Smith dan Read, 2008). Menurut Abbott dan Robson (1984) dan Danesh et al., (2007), setiap spesies endomikoriza mempunyai “innate effectiveness” atau kemampuan yang spesifik dari spesies tersebut untuk meningkatkan pertumbuhan tanaman pada kondisi tanah yang kurang menguntungkan dengan cara membentuk hifa ekstensif di dalam tanah dan pada seluruh sistem perakaran tanaman untuk menyerap fosfor dari larutan tanah.
Cendawan mikoriza dapat digolongkan menjadi tiga kelompok yaitu ektomikoriza, ektendomikoriza dan endomikoriza. Pengelompokan ini berdasarkan pada bentuk morfologi hifa mikoriza yang mengkolonisasi akar (Smith and read, 1997 ; Brundrett, 2008). Cendawan Ektomikoriza merupakan kolonisasi hifa mikoriza di luar kortek akar dengan cara membentuk jaringan hifa (mantel hifa) yang menyelimuti bagian luar epidermis akar tanaman sebaliknya endomikoriza merupakan cendawan yang mampu membentuk eksternal hifa dan internal hifa. Kolonisasi internal hifa berkembang di dalam kortek akar dengan membentuk vesikel dan atau arbuskula tergantung pada tiap genera (Brundrett et al., 2008; Smith dan Read, 2008). Ektendomikoriza adalah cendawan mikoriza yang mampu mengkolonisasi perakaran baik secara internal hifa di kortek maupun eksternal hifa yang menyelimuti epidermis akar. Hifa yang terbentuk baik ekternal maupun internal sangat sedikit atau tipis sehingga simbiosis ektendomikoriza tidak jelas peran dan fungisinya bagi tanaman (Mikola, 1980 ; Smith dan Read, 1997).
Beberapa ahli dan peneliti yang fokus dalam cendawan endomikoriza mengelompokkan cendawan endomikoriza dengan beberapa istilah yaitu, Fungi Mikoriza Arbuskula (FMA) atau cendawan endomikoriza (Simanungkalit, 2003; Kramadibrata, 2008), Cendawan Mikoriza Arbuskula (CMA) ( Delvian, 2006; Novriani dan Madjid, 2009, Hapsoh, 2008) atau Vesicular Arbuscular Mycorrhiza (VAM)-fungi (Smith and Read, 1997; Schϋβler et al., 2001; INVAM, 2005; Brundrett et al., 2008).
Menurut Smith dan Read (1997) dan Brundrett et al., (2008), Arbuskula/Arbuscular adalah struktur hifa yang berasal dari percabangan hifa di
dalam korteks akar inang. Bentuk arbuskula menyerupai pohon kecil dan berfungsi sebagai tempat pertukaran zat-zat metabolit primer (terutama Glukosa dan Fosfor) antara cendawan endomikoriza dan akar tanaman. Brundrett et al (2008) dan Hapsoh (2008) menyatakan bahwa arbuskula mempunyai peran yang sangat vital dari cendawan endomikoriza karena berfungsi sebagai tempat masuknya hara mineral dari tanah yang diabsorbsi oleh akar dan hifa ke dalam sel inang. Proses tersebut menyebabkan terjadinya peningkatan sitoplasma, respirasi dan aktivitas enzim pada kedua organisme tersebut sehingga tanaman/inang akan dapat memanfaatkan fosfor dari cendawan dan sebaliknya cendawan endomikoriza mengabsorbsi glukosa dan karbon dari inangnya.
Vesikula atau vesikel (vesicular/vesicle) adalah merupakan hifa cendawan endomikoriza yang mengalami penggembungan (melebar). Penggembungan hifa bisa terjadi secara internal di dalam sel atau keluar dari sel akar inang yang terbentuk pada hifa terminal dan interkalar. Bentuk vesikel bulat atau oval/lonjong, berisi senyawa lemak sehingga vesikel merupakan organ penyimpanan cadangan makanan bagi cendawan endomikoriza (Brundrett et. al., 2008). Pada kondisi tertentu terutama vesikel yang telah dewasa dapat berperan sebagai spora atau alat pertahanan cendawan tersebut jika berada pada lingkungan yang tidak menguntungkan (Pattimahu, 2004; Mosse, 1991).
Vesikular Arbuskular Mikoriza (VAM-fungi) atau cendawan endomikoriza yang dapat membentuk arbuskel dan atau vesikel merupakan kelompok cendawan yang awal pengklasifikasiannya termasuk ke dalam kelas Zygomycetes, ordo dan Glomales. Ordo Glomales terdiri dari dua (2) sub ordo, yaitu Gigasporinea dan Glomineae. Sub-ordo Gigasporineae hanya
mempunyai satu famili Gigasporacaeae dan hanya terdiri 2 genus, yaitu Gigaspora dan Scutellospora. Pada sub-ordo Glominea mempunyai 4 familia, yaitu famili Glomaceae dengan genus Glomus, famili Acaulosporaceae dengan genus Acaulospora dan Entrophospora, famili Paraglomaceae dengan genus Paraglomus, dan famili Archaeosporaceae dengan genus Archaespora. (INVAM, 2009).
Menurut Schϋβler et al (2012) perkembangan klasifikasi dan identifikasi terbaru, genus Sclerocystis mempunyai kemiripan atau homolog dengan genus Glomus, sehingga Sclerocystis pada beberapa acuan klasifikasi tidak lagi sebagai genus tersendiri tetapi termasuk dalam genus Glomus
Menurut Schϋβler et al. (2001) dan INVAM (2005), perkembangan klasifikasi dari cendawan endomikoriza dikelompokkan sebagai berikut: Kingdom : Mycota
Phyllum : Glomeromycota Kelas : Glomeromycetes Ordo : Glomales Familia 1 : Glomaceae
Genus : Entrophospora, Glomus, Paraglomus, Sclerocystis. Famili 2 : Gigasporineae
Genus : Gigaspora, Scutellospora. Famili 3 : Acaulosporaceae
Genus : Acaulospora
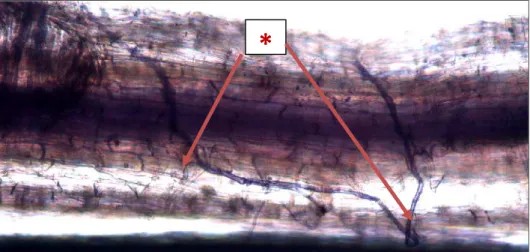
Pada cendawan endomikoriza, terbentuknya vesikel memiliki fungsi dan karakter yang paling menonjol dibandingkan tipe cendawan mikoriza lainnya
(ektomikoriza) karena salah satu spesies cendawan endomikoriza yaitu Glomus sp., merupakan cendawan endomikoriza yang selalu membentuk vesikel struktur tersebut merupakan salah satu struktur inti untuk identifikasi sampai pada tingkat genera pada Glomus (Mosse, 1991; Brundrett et al., 1996)). Glomus sp. mempunyai spora yang tumbuh pada ujung hifa (hifa terminal) dapat berupa spora tunggal, berkelompok (cluster) atau spora-sporanya tersimpan dalam sporokarp (Smith dan Read, 1997). Gigaspora sp. merupakan endomikoriza yang tidak menghasilkan vesikel dalam kortek akar inangnya (Walker, 1983).
Salah satu karakter yang menarik dari endomikoriza adalah pada sporanya karena spora-spora endomikoriza mampu bertahan di dalam tanah tanpa inang sampai 6 bulan bahkan beberapa spesies seperti Scutelospora sp, Gigaspora sp. dapat bertahan sampai satu–dua tahun (Brundrett et al., 2008). Spora-spora yang dihasilkan secara aseksual maupun seksual pada prinsipnya merupakan salah satu bentuk atau alat pertahanan diri di alam yang dapat berfungsi untuk proses adaptasi terutama apabila mikoriza tersebut belum menemukan tanaman inang yang kompatibel (Smith dan Read, 2008). Spora-spora endomikoriza yang telah menemukan inang yang kompatibel akan segera bergerminasi dengan cara membentuk appresorium pada permukaan dinding sel akar inangya dan selanjutnya akan membentuk juluran-juluran hifa. Hifa-hifa tersebut selanjutnya akan menginfeksi/mengkolonisasi ke dalam akar tanaman inang dengan cara menembus atau melalui celah antar sel epidermis dan akhirnya membentuk hifa yang dapat tersebar secara inter-intraseluler dalam korteks akar tanaman (Mosse, 1991; Smith and Read, 1997 ; Brundrett et al., 2008).
2.2. Perkembangan dan Penyebaran Endomikoriza
Faktor biotik dan abiotik dapat mempengaruhi perkembangan dan penyebaran endomikoriza di alam (Smith dan Read, 1997). Faktor lingkungan abiotik mencakup faktor fisika-kimia antara lain: periode musim, perbedaan tempat, suhu, tekstur tanah, intensitas cahaya, kadar air tanah, bahan organik, ketersediaan hara-mineral tanah, logam berat dan fungisida (Sieverding, 1991). Faktor lingkungan biotik adalah mikroorganisme tanah dan tanaman inang (Smith dan Read, 2008).
2.2.1. Perbedaan musim dan lokasi
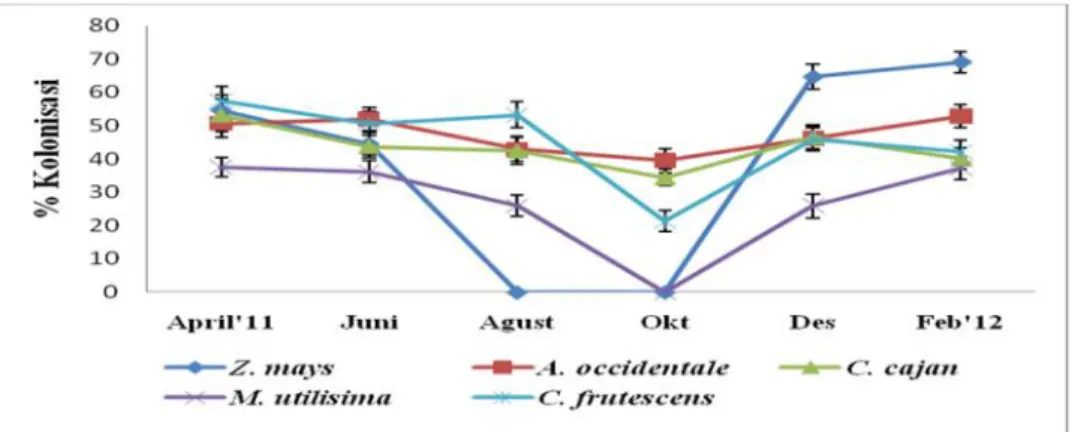
Perbedaan musim atau waktu (temporal) dan tempat (spatial) dapat mempengaruhi persentasi kolonisasi hifa, pembentukan arbuskular,vesikel endomikoriza pada akar-akar inangnya (Proborini, 1998; Reddy et al., 1998). Menurut Oehl, et al.(2006) eksplorasi spora dan jenis-jenis endomikoriza pada tanah dan rhizosfer tanaman inang yang dilakukan pada waktu atau musim yang berbeda akan memperlihatkan keberadaan atau kerapatan jumlah spora dan jenis-jenis endomikoriza karena keberadaan jenis-jenis mikoriza di alam sangat dipengaruhi oleh perbedaan musim, temperatur yang ekstrim (minus 5oC atau diatas 40oC), banyak sedikitnya curah hujan dan tekstur tanah pada daerah tersebut.
.Wilayah yang memiliki empat musim (temperate season) dalam satu siklus musimnya (12 bulan) menunjukkan bahwa persentase dan laju kolonisasi endomikoriza pada akar tanaman pada saat musim dingin sangat rendah tetapi kolonisasi tampak tinggi pada saat musim semi karena pada musim semi, hampir semua tanaman tumbuh secara pesat dan diasumsikan hifa-hifa
endomikoriza tersebar di dalam korteks akar untuk mengabsorbsi gula hasil fotosintesis tanaman inang (Proborini, 1998). Menurut Delvian (2006b) dan Hartoyo et al. (2011) pada saat musim kemarau, kolonisasi endomikoriza terlihat sangat sedikit atau menurun dibandingkan pada saat musim penghujan.
Musim penghujan spora-spora endomikoriza akan germinasi membentuk hifa dan hifa akan tersebar ditanah dan akan mengkolonisasi akar-akar tanaman disekitarnya, karena menurut hapsoh (2008) dan Smith et al (2010) cendawan endomikoriza mampu mengkolonisasi lebih dari 80% tanaman, namun lahan atau tanah yang selalu terendam air sepanjang tahun tidak terdapat kolonisasi endomikoriza pada sistem perakarannya (Proborini, 1998). Perbedaan kolonisasi endomikoriza pada akar tanaman areal kering dan terendam air dilaporkan oleh Carvalho et al. (2001), infeksi/kolonisasi hifa endomikoriza tanaman Aster tripolium, Puccinellia maritima dan Inula crithmoides yang di tanam pada areal yang basah (wet areas) hanya terdapat kolonisasi <10 %, sebaliknya tanaman Spartina maritima, Halimione portulacoides, Arthrocnemum fruticosum dan Arthrocnemum perenne yang hidup di daerah cukup kering (semi-arid areas) >25%. Disimpulkan bahwa ekplorasi endomikoriza pada suatu kawasan dan dilakukan secara periodic dalam satu musim dan kondisi tanah yang berbeda akan sangat bermanfaat untuk melihat tanaman jenis apa saja yang mampu bersimbiosis dengan endomikoriza.
2.2.2. Tanaman inang
Pertumbuhan dan aktivitas mikoriza dapat berbeda antar spesies dan lingkungan. Cendawan mikoriza akan mengkolonisasi inang yang kompatibel (Brundrett et al., 2008). Cendawan mikoriza arbuskular (endomikoriza) adalah
mikroorganisme yang bersifat obligat karena tanpa tanaman inang (asimbiotik) pertumbuhan hifanya sangat sedikit dan hifa hanya mampu bertahan hidup diluar inang selama 20-30 hari (Smith et al., 2003). Menurut Hapsoh (2008) dan Gautam et al. (2009) lebih dari 80% tanaman berpembuluh atau tanaman tingkat tinggi dapat bersimbiosis dengan endomikoriza. Tanaman Jagung, Shorgum dan Gandum merupakan contoh tanaman inang yang sangat kompatibel yang dapat terkolonisasi sangat efektif oleh endomikoriza. Beberapa tanaman budidaya dan tanaman perkebunan lainnya yang dapat terkolonisasi endomikoriza adalah jenis Kacang-kacangan, Kedelai, Barley, Bawang, Nenas, Padi Gogo, Pepaya, Singkong, Tebu, Teh, Tembakau, Palem, Kopi, Karet, Kapas, Jeruk, Kakao, Apel, Mente dan Anggur (Adiningsih et al., 1994; Setiadi, 2002, Hapsoh, 2008).
2.2.3. Suhu
Perkembangan dan aktifitas cendawan endomikoriza dipengaruhi oleh suhu lingkungan. Suhu yang sangat rendah (< 00C) akan menghambat germinasi spora (Brundrett et al., 2008). Menurut Smith et al. (2010), aktivitas dan perkecambahan spora endomikoriza di daerah tropis relatif lebih tinggi dibandingkan daerah sub-tropis karena daerah tropis memiliki kisaran suhu rata-rata diatas 28 0 C dan endomikoriza relatif lebih tahan pada suhu yang cukup tinggi (30 -380C).
Suhu optimum untuk perkecambahan spora endomikoriza bervariasi dan tergantung jenis mikoriza yang dikultur. Mosse (1991) menyatakan bahwa suhu yang cukup tinggi pada siang hari namun masih dibawah 350C tidak menghambat perkembangan akar dan aktivitas fisiologi endomikoriza. Aktifitas
mikoriza menurun pada suhu diatas 40 oC karena pada suhu tersebut akan mengganggu proses metabolisme dan fisiologis tanaman inang sehingga akan berpengaruh pula terhadap aktifitas endomikoriza di dalam korteks akar tanaman inangnya.
Perbedaan suhu yang sangat sangat rendah (- 150C) atau terlalu tinggi dapat (> 50 0C) akan mempengaruhi perkecambahan spora endomikoriza, namun secara genetis ada perbedaan ketahanan enzim pada masing-masing spesies mikoriza terhadap kisaran suhu yang bervariasi sehingga dapat disimpulkan bahwa faktor suhu terhadap perkembangan mikoriza bersifat spesies spesificity (Smith dan Read, 1997). Pada proses perkecambahan spora Gigaspora coralloidea akan optimum jika spora tersebut ditumbuhkan pada media yang mempunyai kisaran suhu 340C, sedangkan untuk spesies Gigaspora margarita dan Gigaspora gigantea akan optimum bergeminasi pada suhu 310C dan tidak berkecambah pada suhu 150C. Pada Glomus epigaeum berkecambah pada suhu 18-25oC, sedangkan Glomus moseae mempunyai toleransi suhu berkisar 25-33 o
C. Secara umum spora-spora endomikoriza akan mati pada suhu 60oC hanya dalam kurun waktu 5-15 menit (Mosse, 1981).
2.2.4. Kadar air tanah
Hasil survey di lapangan menunjukkan bahwa tanaman-tanaman semusim dan perennial (menahun) yang tumbuh di lahan kering Bali Utara (Gerograk Buleleng dan Kubu Karang Asem) mengalami kekurangan air bahkan hampir tidak memperoleh air sama sekali terutama pada saat musim kemarau. Kolonisasi hifa-hifa endomikoriza dalam kortek akar-akar serabutnya sangat menguntungkan tanaman-tanaman tersebut karena dengan adanya hifa-hifa
endomikoriza dapat menjaga kemampuan tanaman untuk bertahan hidup pada kondisi tanah yang kekeringan. Hal ini didukung Smith dan Read (1997) dan Smith et al. (2010), bahwa assosiasi tanaman dengan endomikoriza dapat memperbaiki dan meningkatkan kapasitas serapan air oleh tanaman inang terutama tanaman-tanaman yang tumbuh didaerah marginal atau lahan kering karena hifa eksternal mikoriza dalam menembus dan menyebar ke dalam tanah lebih dari 8 meter yang memungkinkan masih terdapat kandungan air tanah pada kedalaman tersebut.
Hasil penelitian Geeta et al. (2007) menunjukkan bahwa terdapat anomali pada struktur morfologi dan anatomi bibit tanaman alpukat yang diinokulasi endomikoriza. Pada tengah hari saat kadar air rendah, daun bibit alpukat yang diinokulasi endomikoriza tetap terbuka (tidak layu) sedangkan tanaman yang tidak dinokulasi endomikoriza (tanaman kontrol) tertutup daunnya (layu). Hal ini menunjukkan bahwa akar tanaman yang tidak terkolonisasi oleh endomikoriza mempunyai kapasitas serapan air yang lebih rendah dibanding dengan akar tanaman yang berendomikoriza karena hifa-hifa eksternal endomikoriza membantu dalam absorbsi air sedangkan hifa internal yang terdapat dalam kortek akar akan membantu dalam peyimpanan ketersediaan air.
Meningkatnya kapasitas serapan air pada tanaman alpukat yang telah diinokulasi endomikoriza dapat meningkatkan ketahanan bibit terhadap kekeringan dan bibit lebih cepat beradaptasi apabila dipindahkan dari areal pembibitan (nursery ground) kedaerah lain atau di perkebunan (field areas). Hasil penelitian membuktikan bahwa tanaman yang terkolonisasi endomikoriza lebih tahan terhadap kekeringan dibanding tanaman tidak terkolonisasi antara
lain: (1) mampu menurunkan resistensi akar terhadap gerakan air sehingga transpor air ke akar meningkat, (2) meningkatkan status/kadar Fosfor pada tanaman dan (3) hifa-hifa ekternal endomikoriza mampu berekspansi sehingga dapat membantu tanaman memperluas radius penyerapan air diareal-areal marginal yang sangat tandus (Smith dan Read, 2000).
Menurut Smith et al. (2010), penemuan yang menarik adalah adanya korelasi positif antara potensial air tanah dan aktivitas hidup endomikoriza. Tanaman bermikoriza memerlukan jumlah air lebih sedikit dibandingkan dengan tanaman yang tidak bermikoriza untuk memproduksi satu (1) gram bobot kering tanaman. Hal ini disebabkan tanaman bermikoriza lebih tahan terhadap kekeringan dan lebih ekonomis dalam pemakaian air. Selain itu, adanya miselium eksternal pada perakaran tanaman yang bermikoriza menyebabkan endomikoriza lebih efektif dalam mengagregasi butir-butir tanah sehingga hifa-hifa (miselium) endomikoriza dapat meningkatkan kemampuan tanah dalam menyimpan air dibandingkan kondisi tanah tanpa miselium eksternal.
2.2.5. Tingkat keasamam tanah (pH)
Tingkat keasaman tanah memiliki peranan penting dalam perkecambahan spora endomikoriza. Menurut Powell dan Bagyaraj (1984), pH mempengaruhi aktifitas enzim pada proses germinasi spora mikoriza arbuskula. Menurut Widiastuti ( 2004), pada tanah dengan kondisi pH rendah ( keadaan asam) dapat menghambat hifa eksternal endomikoriza untuk dapat mengabsorbsi P yang penting untuk germinasi spora mikoriza. Hal ini berdampak pada proses perkecambahan spora endomikoriza. Perkecambahan spora-spora mikoriza
termasuk endomikoriza bersifat spesies spesifik, misalnya Glomus mosseae dapat berkecambah pada pH 6 – 9, sedangkan Gigaspora coralloidea dan G. heterogama mampu berkecambah pada kisaran pH 4 -6 (Mikola, 1980).
2.2.6. Bahan organik dalam tanah
Bahan organik merupakan komponen penyusun tanah yang penting selain bahan anorganik, air dan udara. Jumlah spora endomikoriza (VA-Mikoriza) yang maksimum akan ditemukan pada tanah yang mengandung bahan organik 1-2 persen dan jumlah spora ditemukan dalam jumlah sedikit pada tanah berbahan organik kurang dari 0.5 persen (Hameeda et.al., 2007). Serasah pada permukaan dan di dalam tanah yang terkolonisasi oleh hifa mikoriza dapat berperan sebagai media atau inokulum mikoriza sehingga dapat mengkolonisasi akar tanaman lainnya pada satu areal (Adiningsih et al., 1994).
2.2.7. Mikroba tanah
Mikroba tanah memiliki pengaruh terhadap perkecambahan spora mikoriza. Percobaan menggunakan spesies Glomus sp. yang ditumbuhkan pada media agar ditambahkan tanah non steril dan air secukupnya diperoleh hasil perkecambahan meningkat namun pada serangkaian percobaan lainnya menggunakan spora Glomus epigaeum yang ditumbuhkan pada tanah steril tidak terjadi perkecambahan spora mikoriza (Mikola, 1980 ; Imas et al., 1989 ).. Menurut Daniel dan Trappe (1980), kegagalan germinasi pada spora endomikoriza dikarenakan pada tanah yang steril tidak terdapat kehidupan berbagai mikroba tanah termasuk bakteri endofitik diazotrop. Bakteri tersebut mampu memproduksi zat perangsang mirip dengan hormon pertumbuhan yang berguna untuk memacu perkecambahan spora mikoriza. Hal serupa
dikemukakan oleh Hameeda et al. (2007) bahwa peningkatan perkecambahan terjadi dikarenakan pada tanah nonsteril terdapat mikroba tanah yang memberikan zat perangsang pertumbuhan bagi perkecambahan spora mikoriza. Namun hasil penelitian Douds et al. (2010) menunjukkan bahwa diperoleh jenis-jenis mikroba patogen baik bakteri maupun cendawan yang terdapat dalam tanah non steril. Bakteri dan cendawan patogen tersebut cukup significan dalam menghambat perkecambahan spora endomikoriza karena mikroba tersebut banyak menginfeksi hifa dan spora mikoriza sehingga menyebabkan kemampuan germinasi spora-spora tersebut terhambat.
2.2.8. Cahaya dan ketersediaan hara
Intensitas cahaya yang tinggi, kekurangan Nitrogen ataupun Fosfor pada level/konsentrasi sedang akan meningkatkan jumlah karbohidrat di dalam akar sehingga tanaman lebih peka terhadap kolonisasi endomikoriza. Prosentase kolonisasi yang tinggi ditemukan pada tanah-tanah yang mempunyai kesuburan rendah (Smith et al., 2010). Tanaman yang tumbuh pada daerah subur dan memiliki pertumbuhan perakaran yang sangat intensif justru akan mengalami penurunan jumlah persentase kolonisasi endomikoriza pada akar tersebut, sebaliknya jika tanaman tumbuh pada lahan miskin hara mineral ditanah dengan intensitas pertumbuhan cabang akar yang rendah menunjukkan peningkatan kolonisasi endomikoriza pada akar, terutama akar-akar serabut (Oehl et al., 2004). Hal ini membuktikan bahwa endomikoriza sangat bermanfaat pada tanaman yang tiumbuh pada daerah kurang subur atau miskin hara.
Peran endomikoriza diasumsikan terkait dengan penyedian dan status Fosfor dalam tanah. Daerah atau lahanberiklim sedang yang mengandung
konsentrasi Fosfor yang tinggi, kolonisasi endomikoriza pada perakaran tanaman disekitarnya akan menurun atau rendah. Hal ini mungkin disebabkan konsentrasi Fosfor internal yang tinggi dalam jaringan inang sehingga iang tidak mengadakan simbiosis dengan endomikoriza (Smith dan Read, 2008). Menurut Miyasaka et al. (2009), cendawan endomikoriza mampu mengeluarkan enzim fosfatase dan asam-asam organik sehingga apabila terdapat banyak hifa atau spora dari cendawan endomikoriza pada tanah yang miskin unsur fosfor (P), endomikoriza dapat melepas P yang terikat sehingga membantu penyediaan unsur P bagi tanaman inangnya. Hifa-hifa dari cendawan mikoriza akan mengkolonisasi bagian apoplast dan sel korteks untuk memperoleh karbon hasil fotosintesis dari tanaman inangnya sebagai satu keuntungan yang diperoleh oleh cendawan endomikoriza apabila bersimbiosis dengan tanaman inangnya (Delvian, 2006b).
2.2.9. Pengaruh logam berat dan unsur lain di tanah
Menurut Delvian (2006 b), tanah-tanah di lahan tropis sering mengalami permasalahan dalam hal salinitas baik tanaman yang hidup pada lahan dengan kadar garam yang tinggi atau rendah.atau terdapatnya kontaminasi Alumunium maupun Mangan pada tanah. Pengaruh unsur-unsur Sodium, Klorida, Aluminium dan Mangan pada keberadaan endomikoriza baik jumlah spora atau kolonisasi pada tanaman masih sedikit diketahui. Menurut Setiadi (2000) pengaruh secara langsung maupun tidak langsung dari masing-masing ion tersebut terhadap perkembangan endomikoriza berkaitan dengan proses fotosintesis dan pertumbuhan tanaman ataupun metabolisme inang. Selanjutnya Widiastuti et al. (2002) menyatakan bahwa infeksi atau kolonisasi
endomikoriza terlihat lebih tinggi pada tanaman yang tumbuh dan hidup di tanah dengan kandungan logam rendah karena pada tanah dengan kandungan logam yang tinggi dapat meracuni tanaman tersebut, namun dengan inokulasi spora dan propagul endomikoriza pada fase pembibitan, tanaman tumbuh dengan baik pada tanah dengan kondisi yang keasamannya tinggi karena pengaruh logam Fe.
2.2.10. Fungisida
Fungisida memiliki pengaruh terhadap perkecambahan endomikoriza. Menurut Cruz et al. (2000), pemberian konsentrasi pestisida benomyl yang relatif rendah (0,001-0,1μm/ml) dapat meningkatkan perkecambahan spora Glomus mosseae, pada konsentrasi yang relatif tinggi (1-2,12μm/ml) akan menghambat perkecambahan spora namun konsentrasi yang sangat tinggi (10-21,25μm/ml) dapat menyebabkan kegagalan spora untuk berkecambah.
Proses perkecambahan spora Glomus mosseae pada pemberian fungisida benomyl konsentrasi relatif rendah (0,001-0,1μm/ml) tidak mempengaruhi perkecambahan spora. Hal ini dikarenakan konsentrasi tersebut masih belum menghalangi osmosis air dari larutan fungisida ke larutan sel spora, dan pemberian fungisida tersebut diduga justru akan merangsang tanaman inang menghasilkan eksudat akar yang berpengaruh dalam mempercepat perkecambahan spora G. mosseae. Eksudat akar tersebut kemudian dikenal dengan faktor M yang berfungsi seperti halnya hormon. Faktor M yang dihasilkan eksudat akar dan spora endomikoriza dapat meningkatkan terjadinya kolonisasi mikoriza dengan inang. Akar-akar tanaman dan endomikoriza yang telah bersimbiosis membentuk kolinisasi/infeksi pada kortek akar dapat pula
meningkatkan produksi hormon pertumbuhan seperti auksin, sitokinin dan giberelin bagi tanaman inangnya (Imas et al., 1989 ; Smith and Read, 1997). Auksin berfungsi memperlambat proses penuaan akar sehingga fungsi akar sebagai penyerap unsur hara dan air akan bertahan lebih lama (Imas, et al., 1989). Hal ini tentunya akan sangat menguntungkan tanaman sehingga tanaman akan lebih lama melakukan aktifitas hidupnya.
Pemberian fungisida jenis benomyl dengan konsentrasi yang cukup rendah (1-2,12μm/ml) dapat berpengaruh pada penurunan laju osmosis air dari larutan fungisida ke larutan sel spora sehingga pada konsentrasi fungisida yang rendahpun sudah mampu menghambat perkecambahan spora endomikoriza. Konsentrasi benomyl yang sangat tinggi (10- 21,25μm/ml) sangat tidak direkomendasikan karena akan menghentikan osmosis air pada spora sehingga menggagalkan germinasi spora endomikoriza (Alizadeh, 2011). Fungisida jenis Agrosan, Benlate, Plantavax dalam konsentrasi yang sangat rendah (2.5 mg/g tanah) dapat menurunkan kolonisasi endomikoriza pada akar tanaman sehingga mengakibatkan penghambatan pertumbuhan tanaman (tahap pembibitan) karena kemampuan penyerapan Fosfat yang rendah pada tanaman tersebut. ( Brundrett et al., 2008 ; Smith et al., 2003).
Penelitian lapangan yang bersifat sinergis dan terpadu pada pengendalian penyakit tanaman secara biologis perlu dikembangkan sehingga tidak memberikan efek negatif terhadap kolonisasi endomikoriza pada akar. Hal ini disebabkan spora dan hifa cendawan mikoriza peka terhadap fungisida sintetik (Powell dan Bagyaraj, 1984; Alizadeh, 2011).
2.3. Tanaman mente (Anacardium occidentale L.)
Tanaman mente bukan merupakan tanaman asli Indonesia, Tanaman ini berasaI dari pegunungan di benua Amerika yang beriklim tropis (Brasilia). Tanaman mente tersebar di daerah tropis, terutama di Asia Selatan seperti India, Burma, Filipina, Semenanjung Melayu dan Indonesia. Mente memiliki banyak nama/istilah seperti Cashew-nut (Inggris), Jambu Moyet, Jambu Mente (Indonesia); Jambu Mente (Jawa), Jambu Mede (Sunda), Gaju (Lampung) (Mandal, 2000).
Penyebaran mente ke Indonesia dibawa oleh pelaut Portugis sekitar 400 tahun yang lalu, dan dibudidayakan di wilayah-lahankering di Kawasan Timur Indonesia. Produktivitas jambu mente dipengaruhi oleh banyak faktor antara lain mutu/kualitas benih yang digunakan, faktor abiotik (tekstur tanah, kelembaban, suhu, cahaya) (Opoku-Ameyaw, et.al., 2007) faktor biotik (mikrobia) (Zaubin dan Daras, 2002).
Tanaman mente diketahui sebagai tanaman budidaya tahunan. Tanaman tersebut seringkali akan mengalami gagal panen apabila dalam periode generative (periode pembungaan) kondisi lingkungannya tidak mendukung seperti curah hujan tidak memadai dan tidak menentu atau terjadi kekeringan selama 1-2 minggu berturut-turut (Mandal, 2000; Sudarto et al., 2004). Selain hal tersebut diatas, salah satu kendala yang mempengaruhi kegagalan dalam pembibitan yaitu pada periode penyimpanan biji mente (Ibiremo, 2010).
Periode penyimpanan biji mente yang terlalu lama (lebih dari 12 bulan) dapat menurunkan kemampuan biji mente dalam berkecambah (Aliyu and Akintaro, 2007 ; Ibiremo, 2010). Hasil penelitian menunjukkan bahwa
penyimpanan biji mente yang dilakukan dari satu tahun setelah pemanenan menghasilkan viabilitas yang rendah (<75%) dalam perkecambahan biji. Berdasarkan pengamatan pada uji pendahuluan penelitian ini, biji mente hasil penyimpanan produk panen yang disimpan lebih dari 12 bulan sangat terhambat kemampuan germinasinya. Hasil pengamatan terlihat jelas bahwa meskipun biji-biji mente tersebut mampu bergerminasi dan tumbuh menjadi bibit namun membutuhkan waktu yang lama yaitu antara 15- 20 hari.
Berdasarkan survey dilapangan, faktor dominan yang sangat mempengaruhi produktivitas tanaman mente masih sulit diketahui. Hal ini karena adanya keterkaitan antara satu faktor dengan faktor lainnya. Rendahnya produktivitas buah mente diduga berawal dari persepsi umum bahwa jambu mente tidak menuntut persyaratan tumbuh yang ketat karena tanaman mente mampu beradaptasi baik pada berbagai kondisi tanah baik tanah yang subur, tidak subur, basah atau kering sehingga problem pada produktivitas mente belum terselesaikan sampai saat ini (Sukawidana, 2010).
Hasil studi lapangan diketahui bahwa bibit jambu mente terutama yang berumur kurang dari 1 tahun memiliki sistem perakaran yang sangat sedikit dibanding bibit tanaman berkayu lainnya (misal: jati belanda dan intaran) sehingga kemampuan bibit untuk bisa tumbuh sangat lambat pada periode tanam yang sama. Salah satu alternatif untuk mengatasi hal tersebut adalah dengan cara memanipulasi daerah disekitar perakaran tanaman (daerah rhizosfer) dengan menginokulasi endomikoriza terutama pada fase pembibitan mente (Sukawidana, 2010). Inokulasi endomikoriza akan efektif dan mampu mengkolonisasi akar jika biji-biji atau gelondong mente ditanam pada
kedalaman 7 – 10 cm (Kartini, 1997; Darmais, pers.com., 2011). Menurut Baon (1997), bibit mente yang telah diinokulasi dengan spora cendawan mikoriza dan akarnya telah terkolonisasi oleh hifa endomikoriza, bibit tersebut dapat dipindahkan ke lapangan dan endomikoriza akan terkolonisasi terus pada system perakaran tanaman tersebut sampai tanaman dewasa.
Pengembangan jambu mente di Indonesia umumnya ditanam pada lahan/areal marginal yang memiliki kandungan bahan organik rendah. Penggunaan pupuk organik sangat dianjurkan dalam budi daya jambu mente di lahan marginal. Sebuah penelitian menunjukkan bahwa pemupukan dengan menggunakan bahan organik dan non organik serta ditambahkan inoklasi spora dan propagul endomikoriza dapat memperbaiki kandungan hara tanah dan dapat meningkatkan pertumbuhan dan hasil produksi jambu mente (Daras, 2007; Opoku-Ameyaw et.al., 2007).
2.4. Manfaat Mikoriza Bagi Tanaman
Keuntungan yang didapat dari simbiosis mutualistik antara cendawan endomikriza dan tanaman adalah tanaman memberi gula dan karbon untuk cendawan dan cendawan membantu dalam penyerapan air, fosfat, mineral dan nutrisi lainnya yang diperlukan bagi tumbuhan (Brundrett et al., 2008 ; Smith et al., 2010). Fosfat merupakan unsur essensial yang diperlukan tanaman dalam jumlah banyak (Smith dan Read, 2008 ; Smith et.al., 2010). Sementara pada areal yang memiliki kondisi tanah cenderung bersifat asam dapat menyebabkan fosfat yang terdapat ditanah dalam bentuk tidak tersedia bagi tanaman karena kondisi asam menyebabkan fosfat terikat kuat dengan Fe sebagai senyawa. Keberadaan endomikoriza pada tanah tanah tersebut mampu mengubah fosfat
yang tidak tersedia menjadi tersedia bagi tanaman (Powell dan Bagyaraj, 1984; Widiastuti, 2004).
Akar-akar tanaman yang telah terkolonisasi oleh mikoriza mampu melakukan aktifitas fotosintesis dengan baik sehingga tanaman lebih sehat dan pada akhirnya mampu menghambat infeksi patogen melalui mekanisme enzimatis yang dihasilkan oleh tanaman dan mikoriza. Proses penghambatan masuknya patogen ke akar tanaman yaitu dengan memanipulasi kondisi yang tidak menguntungkan untuk pertumbuhan patogen pada tanaman yaitu dengan jalan menggunakan karbohidrat dan eksudat akar oleh mikoriza, sehingga mikoriza dapat memanfaatkan karbohidrat dan eksudat-eksudat untuk selanjutnya mikoriza mampu mengeluarkan zat-zat tertentu yang dapat berfungsi untuk menekan atau mematikan patogen sehingga patogen tidak mampu menginfeksi akar tanaman (Smith dan Read, 1997).
BAB III.
KERANGKA BERFIKIR, KONSEP DAN HIPOTESIS PENELITIAN
3.1. Kerangka Berpikir
Kawasan di Bali yang tergolong “lahan kering” misalnya sebagian Kabupaten Buleleng dan Karangasem sangat mengandalkan musim hujan dan pupuk untuk meningkatkan hasil pertaniannya karena kawasan tersebut relatif tidak subur (Antara, 2004 ; Daryana, 2010). Salah satu upaya dalam meningkatkan produksi pertanian lahan kering dapat dilakukan dengan pengembangan model pertanian organik menggunakan pupuk hayati mikoriza (Anas, 2000 ; Priono, 2008 ; Sukasta et al., 2010).
Salah satu tanaman yang banyak dibudiyakan pada lahan kering di Bali adalah tanaman mente (A. occidentale L ). Para petani mente umumnya jarang melakukan regenarasi tanaman mente yang telah tua (berumur lebih dari 25 tahun) dan mengganti tanaman tua dengan melakukan pembibitan mente (Suharto, 2012). Hasil survey di lapangan, beberapa kendala yang dihadapi para petani mente dalam melakukan pembibitan mente antara lain: akar bibit mente mudah sekali patah pada waktu pemindahan dari kebun bibit ke areal perkebunan, bibit mudah layu dan mati bila sudah dipindahkan ke areal perkebunan. Menurut Sukawidana (2010), pembibitan mente terdapat banyak kendala antara lain pertumbuhan bibit yang tidak seragam, jumlah perakaran (serabut) yang dihasilkan sangat sedikit sehingga menyebabkan kemampuan absorbsi hara dan mineral dari tanah rendah Kendala-kendala tersebut pada akhirnya menyebabkan produktivitas mente di Bali dan Nusatenggara, termasuk
di lahan kering perkebunan mente Desa Sendang Kecamatan Gerokgak Kabupaten Buleleng dan Desa Sukadana Kecamatan Kubu Kabupaten Karangasem menjadi tidak/kurang maksimum. Meskipun sejauh ini belum diketahui secara pasti penyebab kematian tersebut, diduga bahwa kematian bibit mente tersebut disebabkan oleh rendahnya kemampuan dan daya tahan bibit mente setelah fase pemindahan dari persemaian terutama di Sukadana yang kondisi tanahnya sangat kering (Sukawidana, 2010 ; Darmais-2011 pers com). Mengacu pada kemampuan mikoriza dalam membantu absorbsi air dan mineral tanaman di lahan kering (Novriani dan Madjid, 2009), pemanfaatan mikoriza untuk peningkatan kualitas produk pertanian perlu dikembangkan sehingga diharapkan penelitian ini dapat mengatasi salah satu faktor pembatas pertumbuhan mente pada lahan kering di Bali,
Cendawan endomikoriza mampu berasosiasi pada lebih dari 80% jenis-jenis tanaman berpembuluh. Kolonisasi cendawan endomikoriza pada akar tanaman inang dapat membantu dalam meningkatkan efisiensi penyerapan unsur hara Fosfat pada lahan kering atau lahan marginal (Hapsoh, 2008, Smith et al., 2010) Tanaman yang bermikoriza lebih tahan terhadap kekeringan karena adanya peranan hifa-hifa ekternal mikoriza mampu menyerap air pada pori-pori tanah yang tidak bisa dicapai oleh akar tanaman inangnya (Smith dan Read, 1997).
Pemanfaatan endomikoriza sebagai pupuk hayati telah dicoba pada beberapa tanaman pertanian dan perkebunan di Bali namun endomikoriza yang digunakan sebagai pupuk tersebut adalah merupakan produk komersial yang diperoleh dari luar Bali, yaitu pupuk mikoriza “Biosfer” dalam pembibitan tanaman Panili (Tirta, 2006); kombinasi spora endomikoriza komersial dan
cendawan Trichoderma untuk mengurangi laju infeksi Jamur Akar Putih (JAP) pada tanaman Mente (Priono, 2008); penggunaan pupuk mikoriza “Technofert” meningkatkan produksi tanaman pakan sapi (Sukasta et al., 2010); pemupukan mikoriza pada tanaman gamal di Karangasem (Antara, 2010).
Penelitian tentang pemanfaatan pupuk hayati endomikoriza indigenus yang dieksplorasi dari lahan kering di Bali masih sangat sedikit bahkan bisa dikatakan belum ada data–datanya. Penelitian ekplorasi endomikoriza di lahan-lahan kering di Bali khususnya di rhizosfer mente adalah sangat diperlukan untuk dapat mengetahui isolat endomikoriza jenis apa yang bisa diaplikasikan pada tanaman mente pertanian lahan kering di Bali karena menurut Suryatmana,et al. (2009), sebagian besar mikroba bersifat spesifik area, artinya mikroba yang dieksplorasi dari daerah setempat relatif lebih cepat beradaptasi dan tumbuh pada daerah tersebut dibanding mikroba yang diambil dari daerah lain.
3.2. Konsep Penelitian
Salah satu kendala pemanfaatan endomikoriza untuk pemupukan tanaman yang dibudidayakan di lahan kering termasuk Bali adalah ketersediaan isolat indigenus endomikoriza sebagai pupuk yang sudah siap dalam bentuk pupuk hayati. Cendawan endomikoriza mampu bersimbiosis dengan berbagai tanaman (pertanian, kehutanan dan perkebunan) dan membantu dalam meningkatkan efisiensi penyerapan air dari dalam tanah serta meningkatkan serapan hara mineral pada lahan marginal (Hapsoh, 2008; Suryatmana et al.,2009; Ibiremo, 2010). Keanekaragaman jenis endomikoriza`di alam sangat dipengaruhi oleh kondisi tanah yang basah, kering, tingkat porositas tanah atau bahkan tanah
yang sangat liat (Danesh et al., 2007). Selain keanekaragaman jenis, kerapatan jumlah spora dan persentase kolonisasi endomikoriza di alam sangat dipengaruhi oleh faktor perbedaan tempat (spatial), musim (temporal) dan tinggi rendahnya curah hujan (Bohrer et al.,2001). Keunikan endomikoriza adalah merupakan cendawan tanah yang bersifat simbiosis obligat ( Mikola, 1980; Smith dan Read, 1997; Brundrett et al., 2008). artinya cendawan tersebut tidak bisa diisolasi pada medium buatan sehingga untuk dapat memperbanyak spora dan hifa cendawan endomikoriza harus dilakukan propagasi atau perbanyakan menggunakan inang (Simanungkalit, 2003)
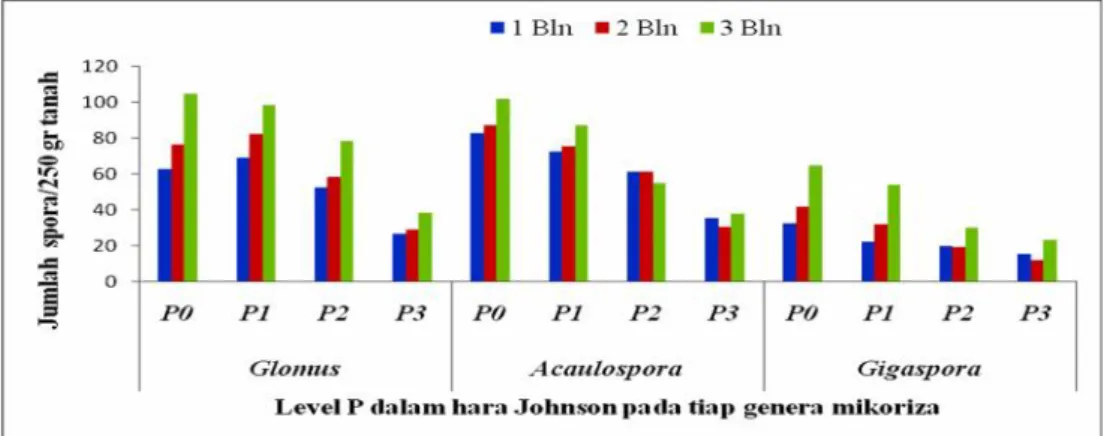
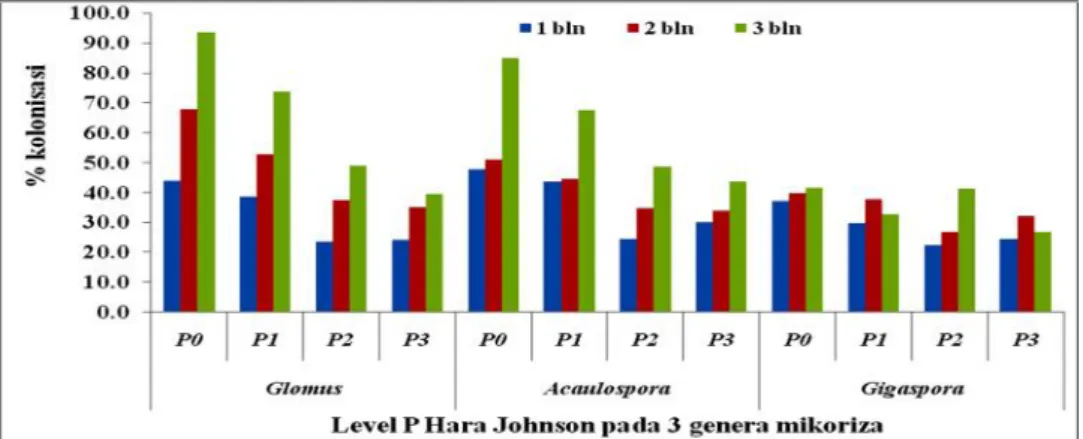
Perbanyakan spora dan propagul endomikoriza dipengaruhi banyak faktor seperti ketersediaan hara tanah (Novriani dan Madjid, 2009); jenis tanaman inang untuk propagasi/perbanyakan spora (Widiastuti, 2002; Simanungkalit, 2003) dan penambahan pupuk (Widiatuti, 2004). Pemanfaatan berbagai macam jenis pupuk telah dicoba seperti vermin-kompos (Sukawidana, 2010) dan pupuk NPK (Tirta, 2006) namun hasil menunjukkan bahwa kedua jenis pupuk tersebut tidak berpengaruh nyata terhadap pertumbuhan sporadan hifa endomikoriza. Menurut Widiastuti (2004) pupuk standar digunakan untuk perbanyakan spora adalah hara Johnson karena formulasi dalam hara Johnson dapat dimanipulasi kandungan Fosfatnya sehingga tidak menghambat pertumbuhan hifa dan kolonisasi endomikoriza diakar. Untuk itu hara Johnson perlu dicoba dan dimanipulasi konsentrasi P didalamnya sehingga tidak menghambat pertumbuhan hifa dan spora indigenus Bali pada fase perbanyakan.
Rendahnya laju pertumbuhan jambu mente di lahan wilayah Nusa Tenggara termasuk Bali diakibatkan oleh kekurangan air, hara, kelembaban tanah serta
kandungan organik tanah yang rendah (Adiningsih et al., 1994; Setiadi, 2000; Rohimat, 2002; Antara 2004). Menurut Widiastuti (2004), inokulasi endomikoriza menggunakan propagul campuran (spora dan akar terkolonisasi hifa) dapat meningkatkan laju pertumbuhan tanaman target. Berdasarkan fakta-fakta tersebut dan guna meningkatkan kualitas pertumbuhan bibit tanaman mente di Bali khususnya maka penelitian tentang pengaplikasian endomikoriza pada pembibitan mente relevan dan penting dilakukan. Diagram alur Konsep penelitian pada Gambar 3.1.
3.3. Hipothesis
Hipothesis dari penelitian ini adalah:
1. Spesies, kerapatan spora dan kolonisasi endomikoriza berbeda antar tempat dan waktu
2. Perbedaan konsentrasi P dalam hara Johnson berpengaruh terhadap jumlah spora dan kolonisasi endomikoriza pada perbanyakan di tanaman Jagung. 3. Perbedaan spesies endomikoriza berpengaruh terhadap respon pertumbuhan
bibit mente (Anacardium occidentale L.).
4. Perbedaan media pembawa inokulum endomikoriza berpengaruh terhadap respon pertumbuhan bibit mente (A. occidentale L.)
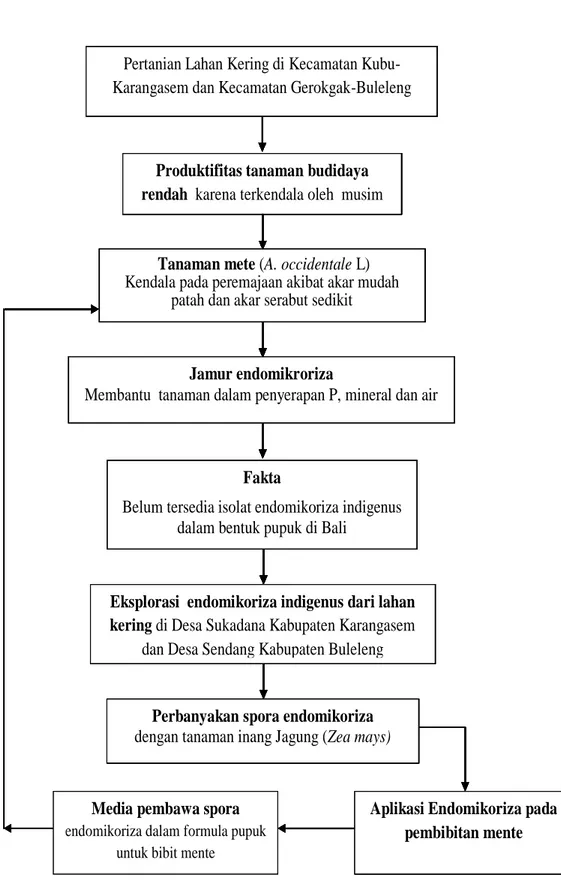
Tanaman mete (A. occidentale L) Kendala pada peremajaan akibat akar mudah
patah dan akar serabut sedikit
Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air
Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng
Aplikasi Endomikoriza pada pembibitan mente Media pembawa spora
endomikoriza dalam formula pupuk untuk bibit mente
Perbanyakan spora endomikoriza dengan tanaman inang Jagung (Zea mays) Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Tanaman mete (A. occidentale L) Kendala pada peremajaan akibat akar mudah
patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Tanaman mete (A. occidentale L) Kendala pada peremajaan akibat akar mudah
patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Tanaman mete (A. occidentale L) Kendala pada peremajaan akibat akar mudah
patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Perbanyakan spora endomikoriza dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Aplikasi Endomikoriza pada pembibitan mente Perbanyakan spora endomikoriza
dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Aplikasi Endomikoriza pada pembibitan mente Perbanyakan spora endomikoriza
dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Aplikasi Endomikoriza pada pembibitan mente Perbanyakan spora endomikoriza
dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Tanaman mete (A. occidentale L) Kendala pada peremajaan akibat akar mudah
patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Perbanyakan spora endomikoriza dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Perbanyakan spora endomikoriza dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Perbanyakan spora endomikoriza dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
Media pembawa spora
endomikoriza dalam formula pupuk untuk bibit mente
Aplikasi Endomikoriza pada pembibitan mente Perbanyakan spora endomikoriza
dengan tanaman inang Jagung (Zea mays) Eksplorasi endomikoriza indigenus dari lahan kering di Desa Sukadana Kabupaten Karangasem
dan Desa Sendang Kabupaten Buleleng Fakta
Belum tersedia isolat endomikoriza indigenus dalam bentuk pupuk di Bali
pupuk
Jamur endomikroriza
Membantu tanaman dalam penyerapan P, mineral dan air Tanaman mete (A. occidentale L)
Kendala pada peremajaan akibat akar mudah patah dan akar serabut sedikit Produktifitas tanaman budidaya rendah karena terkendala oleh musim Pertanian Lahan Kering di Kecamatan Kubu-Karangasem dan Kecamatan Gerokgak-Buleleng
BAB IV
METODE PENELITIAN
4.1.Tempat dan Waktu Penelitian
Penelitian dilaksanakan dalam tiga tahap yaitu: penelitian lapangan (ekplorasi), laboratorium dan Rumah kaca. Penelitian lapangan meliputi pengambilan sampel tanaman dan tanah. Penelitian laboratorium mencakup penghitungan jumlah spora, identifikasi endomikoriza, pengamatan kolonisasi akar, dan analisa tanah. Penelitian rumah kaca meliputi optimasi propagasi/perbanyakan spora endomikoriza, uji efektifitas spora dan propagul pada bibit mente (A.occidentale L.) dan optimasi media pembawa endomikoriza. Penelitian secara keseluruhan bulan April 2011 – Juni 2012.
Lokasi pengambilan sampel dilakukan pada dua lokasi perkebunan mente yaitu di Desa Sendang Kecamatan Grograk Kabupaten Buleleng dan Desa Sukadana Kecamatan Kubu Kabupaten Karang Asem. Penelitian laboratorium dilakukan pada Laboratorium Tanah Fakultas pertanian Unud untuk analisa sifat fisika-kimia tanah, Laboratorium UPT Analitik Universitas Udayana untuk analisa kandungan P tanaman uji dan Laboratorium Taksonomi Tumbuhan Jurusan Biologi F-MIPA Unud untuk penyaringan dan penghitungan spora pada tanah sampel, karakterisasi dan identifikasi endomikoriza, prosessing seluruh sampel akar tanaman, mengamati dan menghitung prosentase kolonisasi endomikoriza. Penelitian lanjutan dilakukan di Laboratoriun Taksonomi Tumbuhan Rendah LIPI Cibinong Bogor untuk identifikasi endomikoriza tingkat spesies. Penelitian di rumah kaca Fakultas Pertanian Universitas
Udayana untuk perbanyakan spora endomikoriza, aplikasi dan optimasi media pembawa endomikoriza pada bibit mente (Anacardium occidentale L.). Alur penelitian pada Gambar 4.1:
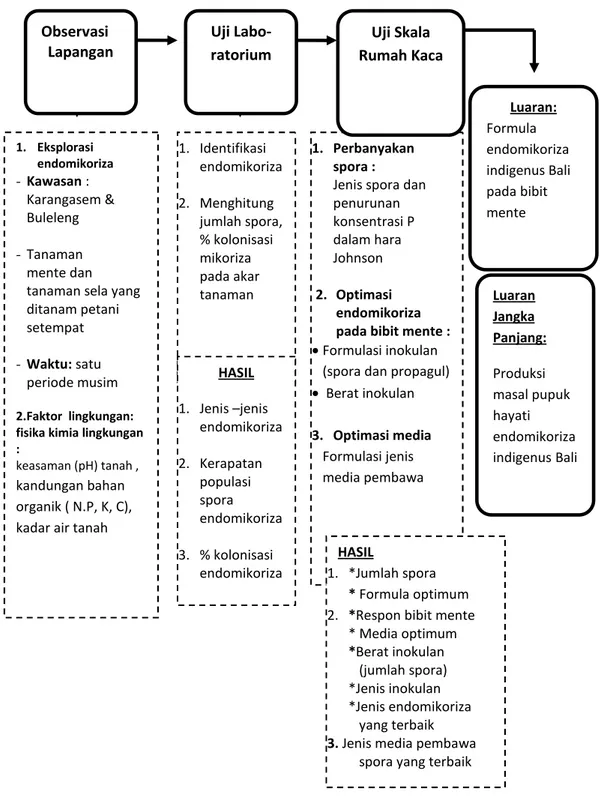
Gambar 4.1. Alur Skema Penelitian 1. Eksplorasi endomikoriza - Kawasan : Karangasem & Buleleng - Tanaman mente dan tanaman sela yang ditanam petani setempat
- Waktu: satu
periode musim
2.Faktor lingkungan: fisika kimia lingkungan :
keasaman (pH) tanah , kandungan bahan organik ( N.P, K, C), kadar air tanah
1. Identifikasi endomikoriza 2. Menghitung jumlah spora, % kolonisasi mikoriza pada akar tanaman 1. Perbanyakan spora :
Jenis spora dan penurunan konsentrasi P dalam hara Johnson 2. Optimasi endomikoriza pada bibit mente : Formulasi inokulan
(spora dan propagul)
Berat inokulan 3. Optimasi media Formulasi jenis media pembawa HASIL 1. *Jumlah spora * Formula optimum 2. *Respon bibit mente
* Media optimum *Berat inokulan (jumlah spora) *Jenis inokulan *Jenis endomikoriza yang terbaik 3. Jenis media pembawa
spora yang terbaik
Observasi
Lapangan Uji Labo-ratorium Rumah KacaUji Skala
HASIL 1. Jenis –jenis endomikoriza 2. Kerapatan populasi spora endomikoriza 3. % kolonisasi endomikoriza Luaran Jangka Panjang: Produksi masal pupuk hayati endomikoriza indigenus Bali Luaran: Formula endomikoriza indigenus Bali pada bibit mente