256 (2001) 99–121
www.elsevier.nl / locate / jembe
Effects of leaf litter addition on meiofaunal colonization of
azoic sediments in a subtropical mangrove in Hong Kong
* Hong Zhou
The Swire Institute of Marine Science and Department of Ecology and Biodiversity, The University of Hong Kong, Hong Kong, China
Received 20 July 2000; received in revised form 17 October 2000; accepted 18 October 2000
Abstract
The responses of major meiofaunal taxa and nematode species assemblage to the decaying leaf litter of the mangrove Kandelia candel were investigated through a field colonization experiment in subtropical Hong Kong. Sixty-four replicate azoic and organic-free sediment cores were treated with leaf litter additions of 03, 0.53, 13 and 23 natural sediment organic concentration, respectively, and retrieved 1, 10, 30 and 60 days post-placement. Replicate cores of ambient sediment were also taken at each sampling date to provide baseline information. Results of ANOVAs suggested that either different meiofaunal taxa responded to the leaf litter in different ways or the response of the same taxon changed over decomposition time. Multivariate ordination performed on nematodes revealed an alteration in community structure after 10, 30 and 60 days between controls and treatments. This alteration was attributed to some deposit feeding nematodes, particularly a bacterivorous species, Diplolaimella sp., which bloomed in all the cores treated with leaf litter, testifying to the important role such meiofauna plays in the process of detritus decomposition. 2001 Elsevier Science B.V. All rights reserved.
Keywords: Mangrove; Kandelia candel; Leaf litter; Decomposition; Meiofauna; Colonization
1. Introduction
Detritus is a major energy source for most marine benthic systems (Findlay and Tenore, 1982). Detritally-enriched marine habitats support a high abundance and diversity of meiofauna. The role meiofauna, particularly free-living nematodes, plays in the detritus-based food chain is, however, still not well understood.
*Tel.: 1852-2809-2179; fax:1852-2809-2197. E-mail address: [email protected] (H. Zhou).
Earlier studies concerning the meiofauna–microbe–detritus relationship were mostly conducted in laboratory cultures (Tietjen and Lee, 1975; Tenore et al., 1977; Milton, 1981; Rieper and Flotow, 1981; Findlay, 1982; Findlay and Tenore, 1982; Pamatmat and Findlay, 1983) and have been reviewed by Warwick (1987). These studies generally suggested that meiofauna, particularly nematodes, could maintain bacterial and diatom assemblages in log phase growth and play an important role in detritus decomposition and even make it available to macro-consumers.
In temperate areas, two closely related species of the nematode family Monhysteridae, namely Diplolaimella dievengatensis and Diplolaimelloides bruciei, are prevalent in salt marshes and their role in stimulating detritus mineralization has been demonstrated (Hopper, 1970; Alkemade et al., 1992a,b, 1993, 1994). Compared with temperate salt marshes, subtropical and tropical mangroves, however, have received relatively less attention and there exists a question about the significance of meiofauna in these detrital systems. For example, Tietjen and Alongi (1990) investigated the relationship between the population growth of two nematode species (Monhystera sp. and Chromadorina sp.), bacterial abundance and the age (chemical composition) of Rhizophora and Avicennia leaf litter. Their results suggested that the nematode populations were more affected by the chemical composition of the leaves than by the bacteria. They concluded that the low field densities (,5 per leaf, Alongi, 1987a) and the inability of nematodes to influence bacterial abundance indicates that meiofauna may not play a major role in the cycling of organic matter in tropical mangrove forests. Gee and Somerfield (1997), however, studied the effects of mangrove diversity and the process of leaf litter decay on the diversity and community structure of meiofauna using a field experiment. Results from this experiment in conjunction with a field investigation on the same mangrove (Somerfield et al., 1998) indicated that the meiofaunal communities on mangrove leaf litter are distinctly different from those in the mud. Nematodes are characterized by a succession of mainly bacterial and deposit-feeding species over the whole decay process while copepods are principally influenced by the life cycle of members of the Darcythompsoniidae.
organic-free sediment with different levels of added leaf litter were inserted into the mangrove sediment and retrieved at different times to determine rates of meiofaunal colonization at varying levels of leaf litter enrichment. Hypotheses to be tested were:
• H Meiofaunal colonization of sediment within the mangrove study area is unaffected0 by the presence of decaying leaves;
• H Meiofaunal colonization and community structure within the mangrove study area0 is unaffected by the quantity of decaying organic matter in the sediment;
• H All meiofaunal taxa within the mangrove study area respond in the same way to0 the presence of decaying organic matter in the sediment.
2. Materials and methods
2.1. Study site
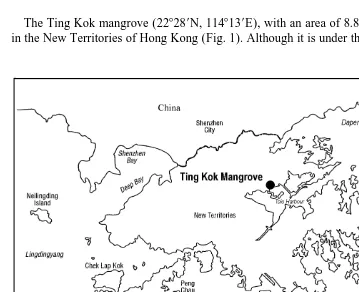
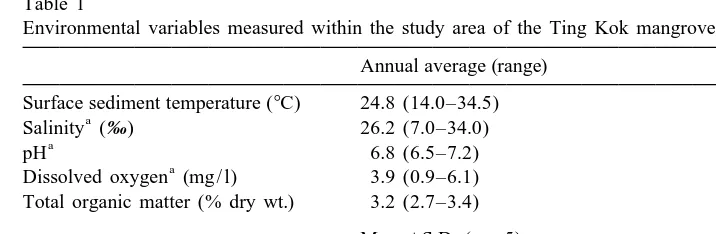
The Ting Kok mangrove (228289N, 1148139E), with an area of 8.8 hectares, is located in the New Territories of Hong Kong (Fig. 1). Although it is under the influence of some
Table 1
Environmental variables measured within the study area of the Ting Kok mangrove forest in 1999 Annual average (range)
Surface sediment temperature (8C) 24.8 (14.0–34.5)
a
Salinity (‰) 26.2 (7.0–34.0)
a
pH 6.8 (6.5–7.2)
a
Dissolved oxygen (mg / l) 3.9 (0.9–6.1) Total organic matter (% dry wt.) 3.2 (2.7–3.4)
Mean6S.D. (n55) Silt / clay (,63mm, % dry wt.) 18.761.6
Sorting coefficient (w) 3.260.1
Mdw 1.260.1
a
Measured from surface pore water.
streams and surface pore water salinity is occasionally diluted to as low as 7.0‰ by monsoon rainfall during summer, salinity is generally oceanic and the forest bottom is of sandy-mud (silt / clay ,20%) with a standing organic matter content of 3.2% (Table 1). Unlike an estuarine soft bottom (muddy) mangal in which litter input is from domestic allochthonous as well as autochthonous sources (Lee, 1988), the Ting Kok mangal receives the bulk of its litter from autochthonous sources, mainly leaves and roots. Ting Kok is dominated by Kandelia candel (L.) Druce, which though occurring only within the Indo-Malaysian region (Duke, 1992), is widely distributed along the coast of Southern China and is the most dominant pioneer mangrove species in Hong Kong (Morton and Morton, 1983). In Hong Kong, tides are mixed semi-diurnal with an annual average tidal range of 1 m. The experiment was conducted in the eastern part of the mangrove around mid-water, where the substratum appeared more uniform without patches of cobbles and spatial variations in the strength and frequency of tidal inundation upon the mangrove were relatively small. This site was chosen for the experiment also because some preliminary work has been done in terms of meiofaunal composition and their spatial distribution (unpublished data) and this basic knowledge will help to interpret the pattern observed in the present study.
2.2. Experimental sediment and leaf litter preparation
Large volumes of surface sediment (8 cm in depth) were collected near the study area. After collection, they were combusted in a laboratory muffle furnace at 5008C for 6 h in order to obtain azoic and organic-free sediment for use in the colonization experiment. Ready-to-fall, senescent Kandelia candel leaves were picked at the study site. After air-drying for 2 weeks, the leaves were powdered into |0.7 mm grains using an electric grinder.
2.3. Estimation of organic content
was determined by incinerating five replicate dried (608C for 24 h) sediment samples of known weight (|10 g) at 5008C for 6 h and calculating the percentage loss of dry weight (Buchanan, 1984). TOM of the prepared leaf detritus was estimated in the same way. These estimates acted as a standard for calculating leaf litter additions to the experimental sediment.
2.4. Experimental design
A two-factor experimental design was adopted. Experimental treatments were represented by different additions of leaf litter to tubes of azoic sediment, corresponding to 03, 0.53, 13 and 23 the TOM of the mangrove sediment and identified as C, L, M and H, respectively. Meiofaunal colonization rates at different stages of leaf litter decomposition were determined by sampling over the time intervals of 1, 10, 30 and 60 days post-placement. Each treatment was replicated four times for each sampling date so that the overall design was 43434, i.e. four replicates with each combination of four levels of treatment and four levels of exposure time.
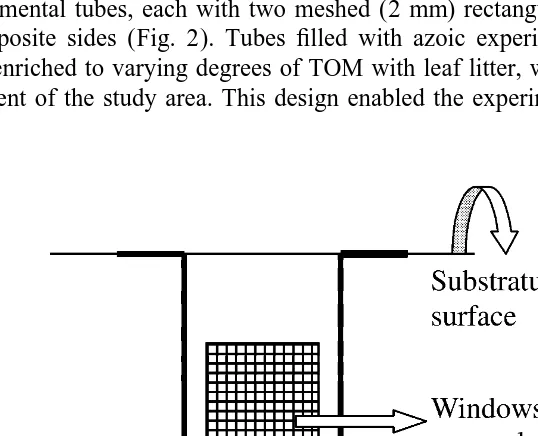
Appropriate amounts of leaf litter corresponding to different doses were homogenized with the azoic and organic-free sediment by stirring using a glass rod. The tubular barrels of 50 cc medical syringes (2.8 cm in diameter and 8.0 cm in depth) were used as experimental tubes, each with two meshed (2 mm) rectangular windows (2 cm33 cm) on opposite sides (Fig. 2). Tubes filled with azoic experimental sediment, which had been enriched to varying degrees of TOM with leaf litter, were embedded in the natural sediment of the study area. This design enabled the experimental sediment to exchange
water and, thus, salts and gases with the surroundings and allowed the meiofauna to settle in the experimental sediment either from the overlying seawater or by migration from the surrounding substratum.
A total of 64 tubes filled with experimental sediment were labeled and placed 2
haphazardly in an area of |8 m around the mid-tide level (11.5 m C.D.) in the Ting Kok mangrove during a low tide on 12 July 1999. Tubes were inserted into the sediment immediately after a similar size core had been removed so that disturbance to the surrounding meiofaunal community during placement was reduced to a minimum. Four replicate tubes of each treatment were retrieved, again haphazardly, on days 1, 10, 30 and 60 post-placement, respectively. After retrieval, each tube was put into a plastic bag and sealed. On each sampling date, four replicate field control cores were also taken haphazardly from within the experimental area to 8 cm sediment depth with a 50 cc syringe corer to provide baseline information on the resident meiofaunal community.
2.5. Sample processing in the laboratory
Upon arrival in the laboratory, each meiofaunal core was fixed immediately with 5% seawater buffered formalin. Faunal extraction procedures followed Somerfield and Warwick (1996) and Warwick et al. (1998). This was a combination of decanting (through 1000 and 63 mm sieves with tap water 10 times), flotation (in diluted Ludox-TM 50 at a specific gravity of 1.15) and centrifugation (three times at 3000 rpm for 3 min each). Major meiofaunal taxa were identified and enumerated under a dissecting microscope. Nematodes were transferred to a glycerine–ethanol mixture (pure glycerine / 70% ethanol, 1:9 in v / v) which was evaporated to pure glycerine, and mounted onto slides for further identification to putative species (Gerlach and Riemann, 1973; Platt and Warwick, 1983, 1988; Warwick et al., 1998) as well as assignment to trophic groups (Wieser, 1953).
2.6. Data analysis
22
Experimental effects on the densities (ind. per core5ind. 6.15 cm ) of major meiofaunal taxa and nematode species numbers were examined using univariate analysis based on a two-way ANOVA linear model: y5m 1Time1Treatment1Time3
Treatment (SPSS for Windows 9.0, 1998). If an interaction of main effects, i.e. time and treatment, was detected, a one-way ANOVA was conducted to examine treatment effects at a specific stage of the experiment. Prior to the analysis of variance, data were ln(x11) transformed to conform to an approximate normal distribution and make the variances constant (Zar, 1999) and then Levene’s test was employed to check the assumption of homogeneity. When the assumption of equal variances was rejected, a non-parametric Kruskal–Wallis test was used instead of the one-way ANOVA. Factors detected to be significant by ANOVAs were further analyzed using a Bonferroni multiple comparison procedure which adjusts the observed significance level by multiplying it by
ˇ
the number of comparisons being made (Norusis, 1999).
and Warwick, 1994). Changes in nematode community structure were examined using non-parametric multivariate techniques contained in the PRIMER (Plymouth Routines in Multivariate Ecological Research) package. MDS (non-metric Multidimensional Scaling Ordination) was based on the Bray–Curtis similarity of either single square root transformed or untransformed, nematode species abundance data. Untransformed data weight more contributions on common rather than rare species compared with the single square root transformation (Clarke and Warwick, 1994). The ANOSIM (Analysis of Similarity) technique was used to test treatment and time effects on the nematode community succession process. A SIMPER (Similarity Percentages) program was then employed to identify those species contributing to differences between field and experimental samples and between treatments observed in the MDS and ANOSIM analysis (Clarke and Warwick, 1994).
For both the univariate and multivariate analyses, a significance level of P#0.05 was used as the rejection value. Field control samples (F) were also analyzed with experimental samples (C, L, M and H) whenever appropriate.
3. Results
3.1. Major taxa
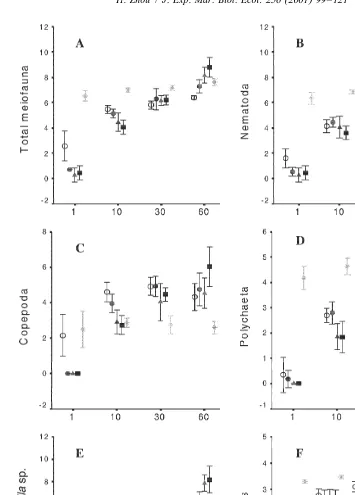
Sixteen taxa were recorded during this experiment. Nematodes were the most dominant group in general, averaging 88% of the total meiofauna over all sampling dates in the field control samples, and 31, 35, 57 and 78% in the control, low, medium and high dose tubes, respectively. Copepods (nauplii in parentheses) accounted for only 2% (0%) of the total meiofauna in the ambient sediment, but 46% (6%), 42% (6%), 28% (3%) and 13% (1%) in the control, low, medium and high dose tubes, respectively. Polychaetes had a relatively constant average dominance around 8% over all sampling dates in the field controls and all levels of treatment. Other groups, including oligochaetes, ostracods, halacaroids, insect larvae, kinorynchs, turbellarians, amphipods, bivalves, isopods, pycnogonids and tanaids, were found in low numbers only and were, thus, not focused upon in this study.
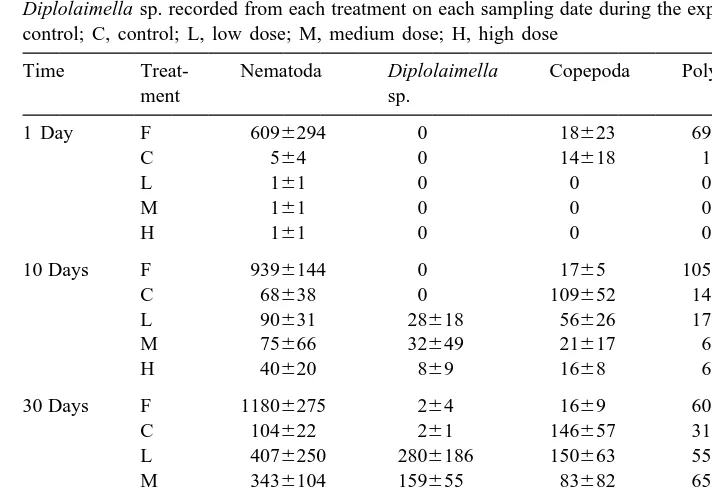
Initial meiofaunal colonization occurred 1 day post-placement, but at a slower rate in the detritus samples than the controls (Table 2; Fig. 3). Copepods colonized faster than nematodes and polychaetes, but seemed more sensitive to the newly-added detritus. That is, copepod numbers in the controls (14618 ind. per core) were close to that of field control samples (18623 ind. per core) after only 1 day, while samples with detritus additions had no copepods. Compared with the numbers in the field control samples (6096294), nematodes initially colonized the experimental sediments in low numbers (564 in controls and 161 in detritus samples).
Table 2
2
Mean numbers (6S.D., n54) per core (6.15 cm ) of major meiofaunal taxa and a bacterial nematode Diplolaimella sp. recorded from each treatment on each sampling date during the experimental period. F, field control; C, control; L, low dose; M, medium dose; H, high dose
Time Treat- Nematoda Diplolaimella Copepoda Polychaeta Total
ment sp. meiofauna
Copepod numbers (109652) were, however, as much as 63 the background levels (1765) after 10 days and reached the highest value (146657) 30 days post-placement, declining to 91663 on the 60th day. In contrast, nematode numbers increased much faster in the detritus samples than in the controls. By the end of the experiment (60 days post-placement), numbers reached the highest value of only 0.23 the background level in the control, but 0.73, 23 and 43 background levels for the low, medium and high dose additions, respectively.
Table 3
Results from two-way ANOVA for the time and treatment effects on numbers (ind. per core) of major meiofaunal taxa and a bacterial feeding nematode Diplolaimella sp. and nematode species numbers. Field controls are not included in these analyses. Day 1 is not included as few meiofauna colonized each tube. A Bonferroni multiple comparison for nematode species numbers is shown in parentheses
Source df P of
Total Nematoda Copepoda Polychaeta Diplolaimella Nematode variation
meiofauna sp. species
Time (days) 2 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 (10530,60)
Treatment 3 0.169 0.053 0.055 0.631 ,0.001 0.030
(C.L5M5H)
Time 6 ,0.001 ,0.001 0.002 0.031 0.031 0.842
3Treatment Total 48
meiofauna to leaf litter additions became significant at the later stages of the experiment, i.e. total meiofauna on the 60th day (H5M.L.C) and nematodes on the 30th (H5M5L.C) and 60th day (H5M.L.C) (Table 4; Fig. 3). No significantly positive effects for copepods and polychaetes could be detected by the end of the experiment.
The rapid increase in nematode numbers in the detritus samples was due to one species, i.e. Diplolaimella sp., which was otherwise rare in controls and ambient mangrove sediments. It reached an average density of 5824 ind. per core in the high dose addition samples on the 60th day (Table 2). An interaction of time and treatment effects on the numbers of Diplolaimella sp. was also revealed by two-way ANOVA (Table 3). The colonization of Diplolaimella sp. started 10 days post-placement and showed significant treatment effects up to the end of the experiment. After 60 days, a positive treatment gradient was clear, i.e. H5M.L4C (Table 4; Fig. 3).
Table 4
Significance levels from one-way ANOVA for the treatment effects on densities of major meiofaunal taxa and a bacterial feeding nematode, Diplolaimella sp., at each specific stage of the experiment. Field controls are not included in these analyses
Days post- Total Nematoda Copepoda Polychaeta Diplolaimella
placement meiofauna sp.
a b a
1 0.020 – 0.002 – –
10 0.110 0.298 0.002 0.023 0.006
30 0.590 0.049 0.251 0.415 ,0.001
a a
60 0.008 0.007 0.089 0.173 ,0.001
a
Kruskal–Wallis test instead of ANOVA.
b
3.2. Nematode community
3.2.1. Diversity
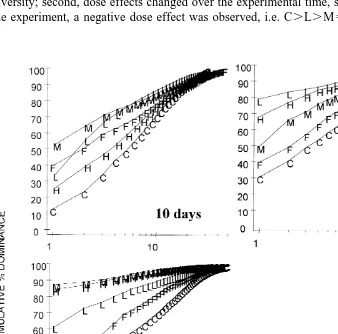
Two-way ANOVA (Table 3) indicated significant time and treatment effects of leaf litter addition on nematode species numbers during the experimental period. The time and treatment interaction was not significant, however. The Bonferroni test (Table 3) showed that nematode species numbers of 10 and 30 days post-placement were significantly less than after 60 days and leaf litter addition significantly reduced them also (C.L5M5H). K-dominance curves (Fig. 4) plotted for different stages of the experiment revealed that: first, the presence of leaf litter reduced nematode species diversity; second, dose effects changed over the experimental time, so that by the end of the experiment, a negative dose effect was observed, i.e. C.L.M5H; and third, after
30 days, diversity of the ambient nematode community (F) was higher than that of the detritus samples (L, M and H) but lower than the controls (C).
3.2.2. Community structure
Nematode community structure in the experimental sediment changed over the colonization process while the community structure in the colonizing pool was rather stable, as revealed by MDS (Figs. 5 and 6). Detritus addition affected community colonization until the end of the experiment and there were always large differences between field and experimental samples during the whole study period (Fig. 6).
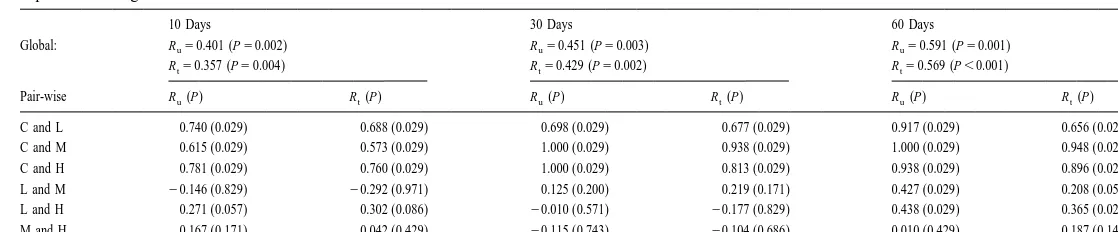
A formal test of experimental effects on the nematode community structure using two-way crossed ANOSIM showed a highly significant time effect for both the global and pair-wise tests between different times (Table 5). A treatment effect was highly significant only between controls and detritus samples. One-way ANOSIM further revealed that treatment effects were significant between controls and the detritus samples after 10 days (Table 6). Dose effects were not so obvious and could only be detected on the 60th day between L and M (Table 6) and L and H (Tables 5 and 6) dose regimes.
3.2.3. Species composition and trophic structure
Most of the species in the experimental sediments are common in the natural
Fig. 6. MDS ordinations of nematode community structure over the experimental period based on untrans-formed (A) and single square root transuntrans-formed (B) abundance data using Bray–Curtis similarity. Field controls (F) are also included and replicates pooled over each sampling occasion for simplification. Abbreviations as used in Fig. 5.
environment of the study site, except for the two extreme cases, i.e. Diplolaimella sp., which was dominant in the detritus samples from day 10 onwards but rare in controls and in the ambient mangrove sediment, and Parastomanema sp., a mouthless nematode which, conversely, was abundant in the natural environment but rarely colonized into the experimental sediments. The top three species contributing to the identified differences in community structure between field and experimental samples and between different levels of treatment at a specific stage of the experiment were indicated by SIMPER analysis and listed in Tables 7 and 8. Experimental samples differed from field controls due to Parastomanema sp. (10, 30 and 60 days), Sabateria praedatrix De Man (10, 30 and 60 days), Metalinhomoeus biformis Juario (30 days) and Chromaspirina sp. (10 days), which were more abundant in the natural environment. The major contributor to the differences between controls and detritus samples was Diplolaimella sp. (10, 30 and
Table 5
Results from two-way crossed ANOSIM global and pair-wise tests using Bray–Curtis similarity, showing the overall time and treatment effects on nematode community structure during the colonization experiment. Ru
and R : R values based on untransformed and single square root transformed abundance data, respectively. C,t
control; L, low dose; M, medium dose; H, high dose
Time (days) Treatment
Global: Ru50.744 (P,0.001) Global: Ru50.429 (P,0.001) Rt50.717 (P,0.001) Rt50.400 (P,0.001) Pair-wise R (P)u R (P)t Pair-wise R (P)u R (P)t
H
.
Zhou
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
256
(2001
)
99
–
121
Table 6
Results from one-way ANOSIM global and pair-wise tests using Bray–Curtis similarity, showing the treatment effects on nematode community structure at a specific experimental stage. Abbreviations as used in Table 5
10 Days 30 Days 60 Days
Global: Ru50.401 (P50.002) Ru50.451 (P50.003) Ru50.591 (P50.001)
Rt50.357 (P50.004) Rt50.429 (P50.002) Rt50.569 (P,0.001)
Pair-wise R (P)u R (P)t R (P)u R (P)t R (P)u R (P)t
C and L 0.740 (0.029) 0.688 (0.029) 0.698 (0.029) 0.677 (0.029) 0.917 (0.029) 0.656 (0.029)
C and M 0.615 (0.029) 0.573 (0.029) 1.000 (0.029) 0.938 (0.029) 1.000 (0.029) 0.948 (0.029)
C and H 0.781 (0.029) 0.760 (0.029) 1.000 (0.029) 0.813 (0.029) 0.938 (0.029) 0.896 (0.029)
L and M 20.146 (0.829) 20.292 (0.971) 0.125 (0.200) 0.219 (0.171) 0.427 (0.029) 0.208 (0.057)
L and H 0.271 (0.057) 0.302 (0.086) 20.010 (0.571) 20.177 (0.829) 0.438 (0.029) 0.365 (0.029)
Table 7
SIMPER analysis of the nematode community identifying the top three species making the biggest percentage contributions (%) to Bray–Curtis dissimilarities (based on single square root transformed abundance data) between field controls and experimental samples at a specific stage of the experiment. F, field control; C, control; L, low dose; M, medium dose; H, high dose
Samples 10 Days 30 Days 60 Days
compared
Species % Species % Species %
F and C Parastomanema sp. 15.5 Parastomanema sp. 15.2 Parastomanema sp. 11.9
a
Sabateria praedatrix 6.7 Sabateria praedatrix 7.5 Diplolaimella sp. 6.5 Chromaspirina sp. 6.5 Metalinhomoeus biformis 5.1 Sabateria praedatrix 5.1
a
F and L Parastomanema sp. 13.6 Parastomanema sp. 13.3 Diplolaimella sp. 13.0
a
Sabateria praedatrix 5.9 Diplolaimella sp. 9.4 Parastomanema sp. 10.9 Chromaspirina sp. 5.8 Sabateria praedatrix 6.3 Sabateria praedatrix 4.7
a
F and M Parastomanema sp. 13.9 Parastomanema sp. 13.0 Diplolaimella sp. 24.6
a
Chromaspirina sp. 6.3 Diplolaimella sp. 7.4 Parastomanema sp. 9.0 Sabateria praedatrix 6.1 Sabateria praedatrix 6.2 Sabateria praedatrix 3.9
a
F and H Parastomanema sp. 12.2 Parastomanema sp. 12.7 Diplolaimella sp. 27.3
a
Chromaspirina sp. 5.5 Diplolaimella sp. 8.4 Parastomanema sp. 7.9
a Sabateria praedatrix 4.9 Sabateria praedatrix 6.2 Anoplostoma viviparum 4.1
a
Average density of the species is higher in the latter sample.
60 days). Other important species included Diplolaimelloides sp. (10, 30 and 60 days),
Theristus sp. (10 and 30 days), Haliplectus wheeleri Coles (30 days), Megadesmolaimus
sp. (60 days) and Anoplostoma viviparum (Bastian) (60 days) in the detritus samples and an unidentified species (10 days) in the controls. Some epi-growth feeders (Type 2A) also showed a positive response to the detritus 60 days post-placement, i.e. Desmodora
cazca Gerlach (L.C), Dichromadora sp. (H.C), Chromaspirina sp. (H.L) and
Paracanthochus sp. (H.M).
Table 8
SIMPER analysis of the nematode community identifying the top three species making the biggest percentage contributions (%) to Bray–Curtis dissimilarities (based on single square root transformed abundance data) between treatments at a specific stage of the experiment. C, control; L, low dose; M, medium dose; H, high dose
Samples 10 Days 30 Days 60 Days
compared
Species % Species % Species %
a a a
C and L Diplolaimella sp. 15.6 Diplolaimella sp. 28.4 Diplolaimella sp. 21.8
a a
Theristus sp. 11.6 Chromaspirina sp. 5.2 Chromaspirina sp. 6.0
a a
Unidentified species 5.6 Haliplectus wheeleri 3.0 Desmodora cazca 4.4
a a a
C and M Diplolaimella sp. 13.1 Diplolaimella sp. 18.6 Diplolaimella sp. 37.2
a a a
Theristus sp. 7.6 Theristus sp. 10.0 Megadesmolaimus sp. 3.5
a
Unidentified species 5.6 Chromaspirina sp. 6.8 Diplolaimelloides sp. 2.9
a a a
C and H Diplolaimella sp. 8.0 Diplolaimella sp. 23.5 Diplolaimella sp. 35.2
a a
Unidentified species 7.1 Diplolaimelloides sp. 5.5 Anoplostoma viviparum 4.4
a a a
Diplolaimelloides sp. 6.7 Theristus sp. 5.5 Dichromadora sp. 3.3
a a
L and M Diplolaimella sp. 12.6 Diplolaimella sp. 13.7 Diplolaimella sp. 24.3
a
Theristus sp. 9.3 Theristus sp. 10.3 Chromaspirina sp. 5.3
a a
Diploliamelloides sp. 6.6 Chromaspirina sp. 5.8 Megadesmolaimus sp. 4.3
a
L and H Diplolaimella sp. 12.2 Diplolaimella sp. 16.2 Diplolaimella sp. 27.4
a a
Theristus sp. 10.8 Theristus sp. 7.2 Anoplostoma viviparum 4.7
a a
Diplolaimelloides sp. 8.8 Diplolaimelloides sp. 6.1 Chromaspirina sp. 3.9
a
M and H Diplolaimella sp. 12.1 Theristus sp. 11.1 Diplolaimella sp. 23.1
a a
Theristus sp. 7.3 Diplolaimella sp. 8.0 Anoplostoma viviparum 4.9
a a
Diplolaimellodies sp. 6.3 Chromaspirina sp. 5.9 Paracanthochus sp. 4.1
a
Average density of the species is higher in the latter sample.
field control and different levels of treatment if the two numerically dominant species, namely Parastomanema sp. and Diplolaimella sp., were not included (Fig. 8A,B).
4. Discussion
Meiofaunal responses to leaf litter additions not only differed among different major taxa but also varied over decomposition time. The meiofaunal colonization of ex-perimental sediments during earlier stages, i.e. days 1 to 10, indicated a negative effect of newly-added leaf litter on the colonization of total meiofauna, copepods and polychaetes. This was possibly related to mangrove-derived tannins or other polyphenic acids. In five mangrove estuaries in Australia, nearly all of the dominant meiofaunal taxa correlated negatively with concentrations of sediment tannins in the low- and mid-intertidal zones (Alongi, 1987b). The field evidence of a negative effect of mangrove-derived tannins on meiofauna was supported by a nematode growth experiment, in which the population growth of Terschellingia longicaudata was limited by Rhizophora
stylosa which contained the highest concentration of tannins of three potential food types
Fig. 7. Changes in nematode trophic composition over the experimental period. Numbers (ind. per core) are obtained by averaging replicates for each treatment at each sampling date. 1A, selective deposit feeder; 1B, non-selective deposit feeder; 2A, epi-growth feeder; 2B, predator / omnivore. To make the plots comparable, Diplolaimella sp. is not included due to its extremely high numbers. C, control; L, low dose; M, medium dose; H, high dose.
leaves from Ting Kok mangrove (12.11% dry wt. for young leaves and 7.25% for mature leaves), and which were close to those of R. stylosa (11.45% for the fresh and 8.23% for 1 week aged leaves) reported upon by Tietjen and Alongi (1990). However, nitrogen content was higher in K. candel (1.86% for young leaves and 1.83% for mature leaves) than in R. stylosa (1.04% for fresh and 1.28% for 1 week aged leaves). Litter bag studies indicated that rapid (14–40 days) loss of tannins from ready-to-fall, senescent
mangrove leaves (R. stylosa, Avicennia marina and Ceriops tagal ) can coincide with rapid increases in densities of bacteria on them (Robertson, 1988). Community development on the leaf litter could be affected and controlled by changes in leaf chemistry and the successional development of the microphyte community (Gee and Somerfield, 1997). Despite a high tannin concentration in K. candel leaves, the also high nitrogen content may be of more importance for meiofaunal community development during the later stages of colonization (30–60 days), as in this study. With an increasing decay time for leaf litter, negative effects shifted to positive ones. By the end of this experiment (60 days post-placement), nematodes and the total meiofauna showed significantly positive responses to the addition of detritus in terms of their relative numbers among the control and different levels of leaf litter addition. A positive response was less evident for copepods and polychaetes. In a north-eastern Malaysian mangrove, Gee and Somerfield (1997) observed that changes in the copepod community on the leaf litter (Rhizophora apiculata and Bruguiera parviflora) were basically an expression of the life cycle of Darcythompsonia. In the present study, the lower P value of 60 days (Table 4, P50.089) compared with 30 days post-placement (Table 4,
P50.251) indicated that a positive response by copepods to leaf litter may exist but could not be detected because of a lack of power in the experimental design (few replicates). Another explanation for this may exist if different colonization mechanisms by the meiofaunal taxa are taken into account. For a small-scale defaunation, nematode dispersal occurs either via suspended or infaunal movement in a uniform way, whereas copepods recolonize sediments predominantly from the water column. The relatively surficial distribution of copepods enhances the probability of passive suspension from current action and limits the possible pathways for infaunal movement (Chandler and Fleeger, 1983). During a vertical distribution study conducted in this mangrove, 53% of nematodes and 96% of copepods were found in the 0–2 cm depth. In this experiment, therefore, leaf litter supplies in the experimental sediment column (0–8 cm in depth) may not be fully available to copepods and, accordingly, their numbers may not be a true reflection of treatment effects.
Nematode community succession was tightly linked to the leaf litter decomposition process in terms of density, species diversity, community structure and trophic composition. Individual numbers increased but species numbers and dominance-based diversity (indicated by the K-dominance curve) decreased with an increased level of leaf litter addition. Community structure and species composition differed among controls and detritus samples. All the differences associated with leaf litter addition could be attributed to the occurrence and rapid increase of a small bacterivorous species,
Diplolaimella sp., in the detritus-enriched samples. Many studies (Lorenzen, 1969;
hand, their presence can stimulate bacterial growth and thus, in turn, enhance the rate of organic detritus breakdown (Warwick, 1987).
Diplolaimella sp., even though dominant in all the detritus samples, was rare in the
ambient mangrove sediment in the study site. In a colonization experiment of PCP-contaminated sand, Cantelmo and Rao (1978) found the abundances of Diplolaimella
punicea in high and medium dose treatments were significantly higher than the control
and low dose ones. Initial colonizers, after defaunation caused by physical disturbances, are normally opportunistic species, which are well adapted to the niches hence made available to them and are able to utilize the newly available resources quicker than their competitors (Grassle and Grassle, 1974). As an opportunistic species, Diplolaimella sp. has an r-strategy life history that is characterized by a relative short generation time, a high reproductive potential and rapid rate of population growth (Warwick, 1987). Moreover, Diplolaimella sp. can take advantage of the food available, i.e. bacteria, which are the first colonizers and can grow rapidly during decomposition of the organic resource (leaf litter or even PCP). In laboratory microcosms, in the absence of meiofauna, colonization of debris derived from algal and salt-marsh plants is succession-al, with bacteria followed by flagellates and ciliates (Newell, 1982).
Diplolaimella sp. was highly dominant numerically in the detritus samples, and thus
dose effect on the successional development of nematode species assemblage could be controlled by this species. Lack of evidence of strong dose effect (between M and H dose regimes in particular) on the population growth of Diplolaimella sp. and, accordingly, nematode diversity, community structure and trophic composition may be because of the limited space and exchange using the tubes for a full development of the
Diplolaimella sp. population (proportional to the high food quantity). But no attempt
was made to evaluate procedural effects.
The generation time of Diplolaimella ocellata, a species associated with mangrove leaves, is only 5–7 days at 308C (Hopper et al., 1973). In a temperate salt marsh, Alkemade et al. (1993) showed that the number of Diplolaimella dievengatensis was most related to the decomposition rate of Spartina anglica leaves in spring, summer and autumn, but not in winter. At the study site in summer, when this experiment took place, the average sediment temperature was 328C. At this temperature, population growths of bacteria and Diplolaimella sp. could be rapid in the detritus samples, but the same colonization pattern may not take place if the experiment was conducted in winter.
The differentiation in nematode community structure and trophic composition between experimental samples and natural environment and between various levels of detritus enrichment was basically an expression of relative abundance of
Paras-tomanema sp. and Diplolaimella sp. in different samples and most possibly a reflection
of their different sources of nourishment, crawling abilities and life histories.
Paras-tomanema sp., a mouthless species, has a large body length / width ratio (a5120) and a
deeper distribution (.2 cm in depth). Its high dominance (40% of total nematode density) indicated low oxygen tension in the mangrove sediment as well as its successful adaptation to this harsh environment by means of epidermal uptake of dissolved organic matter (Jensen, 1987b).
Anoplostoma viviparum and Haliplectus wheeleri were also more abundant than in the
controls. These species are all deposit feeders (Types 1B and 1A) in Wieser’s trophic group scheme. These results are in agreement with those of previous studies in terms of species and trophic composition, though the colonization time scale related to the decaying leaf litter was not well comparable because of the different methods employed. In a temperate salt marsh, Alkemade et al. (1993) demonstrated that the numbers of some deposit-feeding species, namely Diplolaimella dievengatensis, Diplolaimelloides
bruciei, Theristus acer, Desmolaimus zeelandicus and Monhystera parva, were
positive-ly correlated with decomposition rates of Spartina anglica leaves. Similarpositive-ly, Gee and Somerfield (1997) showed that the majority of nematode species colonizing mangrove leaves are bacterial and deposit feeders like Perspiria sp., Theristus sp. and
Diplolaimel-loides sp., etc. The fact that some epi-growth feeders, such as Desmodora cazca and Dichromadora sp., also responded positively to leaf litter additions and all the trophic
groups reached highest numbers in the high dose treatment by the end of the experiment indicated a successional order of nematode trophic groups over the leaf litter decomposi-tion process. This study suggests that deposit-feeding nematodes, especially bacterial feeders, are pioneer colonizers of the leaf litter and may play an important role linking detritus and decomposer to higher trophic levels. Nevertheless, lack of spatial replication in the experimental design could limit the generality of the findings and, thus, complementary field and / or laboratory studies are still needed.
5. Conclusions
1. Meiofaunal colonization of sediments within a mangrove was affected both by the presence and the quantity of decaying leaves in the sediment.
2. Responses to Kandelia candel leaf litter differed among the major meiofaunal taxa and varied over the decomposition time: a negative response to the early stages of decomposition (1–10 days), followed by a transition period (10–30 days) and, finally, a positive response to the later stages of decay (30–60 days). This was probably a reflection of changes in leaf chemistry and the successional development of microphytes.
3. Within the nematode community, species diversity decreased with an increase in individual numbers as a response to leaf litter addition and deposit feeders, especially bacterial feeders such as Diplolaimella sp., are pioneer colonizers of Kandelia candel leaf litter. This suggests that such meiofauna can utilize the microphytes developing during leaf litter degradation as sources of food and may thus play an important role in its further decomposition.
Acknowledgements
Somerfield of Plymouth Marine Laboratory, UK, for their critical comments on the manuscript. [RW]
References
Alkemade, R., Wielemaker, A., Hemminga, M.A., 1992a. Stimulation of decomposition of Spartina anglica leaves by the bacterivorous marine nematode Diplolaimelloides bruciei (Monhysteridae). J. Exp. Mar. Biol. Ecol. 159, 267–278.
Alkemade, R., Wielemaker, A., de Jong, S.A., Sandee, A.J.J., 1992b. Experimental evidence for the role of bioturbation by the marine nematode Diplolaimella dievengatensis in stimulating the mineralization of Spartina anglica detritus. Mar. Ecol. Prog. Ser. 90, 149–155.
Alkemade, R., Wielemaker, A., Hemminga, M.A., 1993. Correlation between nematode abundance and decomposition rate of Spartina anglica leaves. Mar. Ecol. Prog. Ser. 99, 293–300.
Alkemade, R., Wielemaker, A., Herman, P.M.J., Hemminga, M.A., 1994. Population dynamics of Diplolaimel-loides bruciei, a nematode associated with the salt marsh plant Spartina anglica. Mar. Ecol. Prog. Ser. 105, 277–284.
Alongi, D.M., 1987a. Inter-estuary variation and intertidal zonation of free-living nematode communities in tropical mangrove systems. Mar. Ecol. Prog. Ser. 41, 103–114.
Alongi, D.M., 1987b. The influence of mangrove-derived tannins on intertidal meiobenthos in tropical estuaries. Oecologia (Berlin) 71, 537–540.
Alongi, D.M., Boesch, D.F., Diaz, R.J., 1983. Colonization of meiobenthos in oil-contaminated subtidal sands in the lower Chesapeake Bay. Mar. Biol. 72, 325–335.
Austen, M., Warwick, R.M., 1995. Effects of manipulation of food supply on estuarine meiobenthos. Hydrobiologia 311, 175–184.
Buchanan, J.B., 1984. Sediment analysis. In: Holme, N.A., McIntyre, A.D. (Eds.), Methods For the Study of Marine Benthos. Blackwell Scientific, Oxford, pp. 41–65.
Cantelmo, F.R., Rao, K.R., 1978. Effect of pentachlorophenol (PCP) on meiobenthic communities established in an experimental system. Mar. Biol. 46, 17–22.
Chandler, G.T., Fleeger, J.W., 1983. Meiofaunal colonization of azoic estuarine sediment in Louisiana: mechanisms of dispersal. J. Exp. Mar. Biol. Ecol. 69, 175–188.
Clarke, K.R., Warwick, R.M., 1994. Change in Marine Communities: An Approach To Statistical Analysis and Interpretation. Plymouth Marine Laboratory, Plymouth, UK.
Colangelo, M.A., Macri, T., Ceccherelli, V.U., 1996. A field experiment on the effect of two types of sediment disturbance on the rate of recovery of a meiobenthic community in a eutrophicated lagoon. Hydrobiologia 329, 57–67.
Decker, C.J., Fleeger, J.W., 1984. The effect of crude oil on the colonization of meiofauna into salt marsh sediments. Hydrobiologia 118, 49–58.
Deutsch, A., 1978. Gut ultrastructure and digestive physiology of two marine nematodes, Chromadorina ¨
germanica (Butschli, 1874) and Diplolaimella sp. Biol. Bull. 155, 317–335.
Duke, N.C., 1992. Mangrove floristics and biogeography. In: Robertson, A.I., Alongi, D.M. (Eds.), Tropical Mangrove Ecosystems. American Geophysical Union, Washington, DC, pp. 63–100.
Findlay, S., 1982. Effect of detrital nutritional quality on population dynamics of a marine nematode (Diplolaimella chitwoodi ). Mar. Biol. 68, 223–227.
Findlay, S., Tenore, K.R., 1982. Effect of a free-living marine nematode (Diplolaimella chitwoodi ) on detrital carbon mineralization. Mar. Ecol. Prog. Ser. 8, 161–166.
Gee, J.M., Somerfield, P.J., 1997. Do mangrove diversity and leaf litter decay promote meiofaunal diversity? J. Exp. Mar. Biol. Ecol. 218, 13–33.
Gerlach, S.A., Riemann, F., 1973. The Bremerhaven checklist of aquatic nematodes. A catalogue of Nematoda Adenophorea excluding the Dorylaimida. Part 1 / 2. Meeresforschung in Bremerhaven, Supplement 4. Kommissionsverlag Franz Leuwer, Bremen, pp. 1–735.
Grassle, J.F., Grassle, J.P., 1974. Opportunistic life histories and genetic system in marine benthic polychaetes. J. Mar. Res. 32, 253–284.
Hopper, B.E., 1970. Diplolaimelloides bruciei: n. sp. (Monhysteridae: Nematoda), prevalent in marsh grass, Spartina alterniflora Loisel. Can. J. Zool. 48, 573–575.
Hopper, B.E., Fell, J.W., Cefalu, R.C., 1973. Effect of temperature on life cycles of nematodes associated with the mangrove (Rhizophora mangle) detrital system. Mar. Biol. 23, 293–296.
Jensen, P., 1987a. Feeding ecology of free-living aquatic nematodes. Mar. Ecol. Prog. Ser. 35, 187–196. Jensen, P., 1987b. Differences in microhabitat, abundance, biomass and body size between oxybiotic and
thiobiotic free-living marine nematodes. Oecologia 71, 564–567.
Lambshead, P.J.D., Platt, H.M., Shaw, K.M., 1983. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. J. Nat. Hist. 17, 859–874. Lee, S.Y., 1988. The ecology of a traditional tidal shrimp pond in Hong Kong, the production and fate of
microdetritus, and implications for management. Ph.D. Thesis, The University of Hong Kong.
Lorenzen, S., 1969. Freilebende meeresnematoden aus dem Schlickwatt und den Salzwieesen der Nor-dseekuste. Veroff. Inst. Meeresforsch. Bremerh. 11, 195–238.
Milton, R., 1981. The effect of nematode (Diplolaimella shiewoodi ) presence on density of marine sediment bacteria. Am. Zool. 21, 972.
Moens, T., Vincx, M., 1997. Observations on the feeding ecology of estuarine nematodes. J. Mar. Biol. Assoc. UK 77, 211–277.
Moens, T., Verbeeck, L., de Maeyer, A., Swings, J., Vincx, M., 1999. Selective attraction of marine bacterivorous nematodes to their bacterial food. Mar. Ecol. Prog. Ser. 176, 165–178.
Morton, B., Morton, J. (Eds.), 1983. The Sea Shore Ecology of Hong Kong. Hong Kong University Press, Hong Kong, pp. 220–229.
Newell, R.C., 1982. The energetics of detritus utilization in coastal lagoons and nearshore waters. In: Oceanologica Acta, Proceedings International Symposium on Coastal Lagoons, SCOR / IABO / UNESCO, Bordeaux, France, 8–14 September 1981, pp. 347–355.
ˇ
Norusis, M.J. (Ed.), 1999. SPSS 9.0 Guide To Data Analysis. Prentice-Hall, Engelwood Cliffs, NJ, pp. 271–273.
Pamatmat, M.M., Findlay, S., 1983. Metabolism of microbes, nematodes, polychaetes, and their interactions in sediment, as detected by heat flow measurements. Mar. Ecol. Prog. Ser. 11, 31–38.
Platt, H.M., Warwick, R.M., 1983. Free-living Marine Nematodes. Part I. British Enoplids. Synopses of the British Fauna (New Series), Vol. No. 28. Cambridge University Press, Cambridge.
Platt, H.M., Warwick, R.M., 1988. Free-living Marine Nematodes. Part II. British Chromadorids. Synopses of the British Fauna (New Series), Vol. No. 38. Bath Press, Avon, UK.
Rieper, M., Flotow, C., 1981. Feeding experiments with bacteria, ciliates and harpacticoid copepods. Kiel. Meeresforsch. 5, 370–375.
Robertson, A.I., 1988. Decomposition of mangrove leaf litter in tropical Australia. J. Exp. Mar. Biol. Ecol. 116, 235–247.
Somerfield, P.J., Warwick, R.M., 1996. Meiofauna in Marine Pollution Monitoring Programmes. A Laboratory Manual. Directorate of Fisheries Research, Lowestoft.
Somerfield, P.J., Gee, J.M., Aryuthaka, C., 1998. Meiofaunal communities in a Malaysian mangrove forest. J. Mar. Biol. Assoc. UK 78, 717–732.
Tenore, K.R., Tietjen, J.H., Lee, J.J., 1977. Effect of meiofauna on incorporation of aged eelgrass, Zostera marina, detritus by the polychaete Nepthys incisa. J. Fish. Res. Board Can. 34, 563–567.
Thistle, D., Lambshead, P.J.D., Sherman, K.M., 1995. Nematode tail-shape groups respond to environmental differences in the deep sea. Vie Milieu 45, 107–115.
Tietjen, J.H., Alongi, D.M., 1990. Population growth and effects of nematodes on nutrient regeneration and bacteria associated with mangrove detritus from northeastern Queensland (Australia). Mar. Ecol. Prog. Ser. 68, 169–179.
Tietjen, J.H., Lee, J.J., 1977. Feeding behavior of marine nematodes. In: Coull, B.C. (Ed.), Ecology of Marine Benthos. University of South Carolina Press, Columbia.
Tong, Y.F., 1998. Herbivory on the mangrove Kandelia candel (L.) Druce in Hong Kong. M. Phil. Thesis, The University of Hong Kong.
Warwick, R.M., 1987. Meiofauna: their role in marine detrital systems. In: Moriarty, D.J.W., Pullin, R.S.V. (Eds.), Detritus and Microbial Ecology in Aquaculture. ICLARM Conference Proceedings, Vol. 14. International Center for Living Aquatic Resources Management, Manila, Philippines, pp. 282–295. Warwick, R.M., Platt, H.M., Somerfield, P.J., 1998. Free-living Marine Nematodes. Part III. British
Monhysterids. Synopses of the British Fauna (New Series), Vol. No. 53. Field Studies Council, Shrewsbury.
¨ ¨
Wieser, W., 1953. Die Beziehung zwischen Mundhohlengestalt. Ernahrungsweise und Vorkommen bei freilebenden marnen Nematoden. Ark Zool. 4, 439–484.