FISIOLOGI TUMBUHAN
HORMON
MAKALAH
UNTUK MEMENUHI TUGAS MATA KULIAH Fisiologi Tumbuhan
yang dibina oleh Ir. Nugrahaningsih, M.P. disajikan Hari Senin, 31 Oktober 2016
Oleh : Kelompok 2
S1 Biologi/ Kelas H Tahun 2015
Ida Nurpitasari 150342604029
Reynanda Dwi Aprilia 150342607133
Sugi Hartono 150342608273
Uun Rohmawati 150342604651
Woro Ayu Ambarwulan 150342607204
UNIVERSITAS NEGERI MALANG
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM JURUSAN BIOLOGI
KATA PENGANTAR
Puji syukur penulis panjatkan kehadirat Allah SWT, yang telah melimpahkan rahmat, taufik, dan hidayah-Nya sehingga penulis dapat menyelesaikan makalah yang berjudul “Hormon”. Makalah ini dibuat dalam rangka memenuhi tugas mata kuliah Fisiologi Tumbuhan. Meskipun terdapat beberapa hambatan dalam proses pengerjaan makalah ini, tetapi kami berhasil menyelesaikannya dengan tepat waktu.
Pada kesempatan ini penulis mengucapkan terima kasih kepada:
1. Ir. Nugrahaningsih, M.P. selaku dosen mata kuliah Fisiologi Tumbuhan, 2. kedua orang tua kami yang telah memberikan dukungan moril dan spiritual,
3. seluruh teman seperjuangan Biologi kelas H tahun 2015, yang banyak membantu dan memberi masukan dalam pengerjakan makalah ini, dan
4. semua pihak yang tidak dapat kami sebutkan satu persatu.
Penulis berharap adanya masukan yang bersifat membangun sehingga makalah ini dapat lebih sempurna. Penulis juga berharap agar makalah ini nantinya dapat berguna bagi semua kalangan.
Malang, 30 Oktober 2016
Penulis
Kata Pengantar ...……….…....ii
Daftar Isi ...iii
BAB I. PENDAHULUAN...l A. Latai Belakang…...………...1
B. Rumusan Masalah…………...…………...………...2
C. Manfaat...2
BAB II. KAJIAN TEORI ...3
A. Hormon Auksin ………...………..………...……...3
B. Hormon Giberelin ...9
C. Hormon Sitokinin ………...…...………...19
D. Hormon Asam Absisat ………...………...………...28
E. Hormon Etilen ………...………...33 F. Hormon Brasinosteroid ………...……...………...35 BAB 3. PENUTUP ...38 A. Kesimpulan ………...………...38 B. Saran ………...………...………...38 Daftar Pustaka ………...………...39
BAB 1 PENDAHULUAN A. Latar Belakang
Hormon merupakan zat pengatur tumbuh, yaitu molekul organik yang dihasilkan oleh satu bagian tumbuhan dan ditransportasikan ke bagian lain yang dipengaruhinya. Hormon pada tumbuhan (fitohormon) adalah sekumpulan senyawa organik bukan hara (nutrien), baik yang terbentuk secara alami maupun dibuat oleh manusia, yang dalam kadar sangat kecil (di bawah satu milimol per liter, bahkan dapat hanya satu mikromol per liter) mendorong, menghambat, atau mengubah pertumbuhan, perkembangan, dan pergerakan (taksis) tumbuhan. Hormon tumbuhan merupakan bagian dari sistem pengaturan pertumbuhan dan perkembangan tumbuhan. Kehadirannya di dalam sel pada kadar yang sangat rendah menjadi prekursor (“pemicu”) proses transkripsi RNA. Hormon tumbuhan sendiri dirangsang pembentukannya melalui signal berupa aktivitas senyawa-senyawa reseptor sebagai tanggapan atas perubahan lingkungan yang terjadi di luar sel. Kehadiran reseptor akan mendorong reaksi pembentukan hormon tertentu. Apabila konsentrasi suatu hormon di dalam sel telah mencapai tingkat tertentu, atau mencapai suatu nisbah tertentu dengan hormon lainnya, sejumlah gen yang semula tidak aktif akan mulai berekspresi. Dari sudut pandang evolusi, hormon tumbuhan merupakan bagian dari proses adaptasi dan pertahanan diri tumbuh-tumbuhan untuk mempertahankan kelangsungan hidup jenisnya.
Hormon tumbuhan merupakan bagian dari proses regulasi genetik dan berfungsi sebagai prekursor. Rangsangan lingkungan memicu terbentuknya hormon tumbuhan. Bila konsentrasi hormon telah mencapai tingkat tertentu, sejumlah gen yang semula tidak aktif akan mulai ekspresi.
Pemahaman terhadap fitohormon pada masa kini telah membantu peningkatan hasil pertanian dengan ditemukannya berbagai macam zat sintetis yang memiliki pengaruh yang sama dengan fitohormon alami. Aplikasi zat pengatur tumbuh dalam pertanian modern mencakup pengamanan hasil (seperti penggunaan cycocel untuk meningkatkan ketahanan tanaman terhadap lingkungan yang kurang mendukung), memperbesar ukuran dan meningkatkan kualitas produk (misalnya
dalam teknologi semangka tanpa biji), atau menyeragamkan waktu berbunga (misalnya dalam aplikasi etilena untuk penyeragamanpembungaan tanaman buah musiman).
Hormon tumbuhan tidak dihasilkan oleh suatu kelenjar sebagaimana pada hewan, melainkan dibentuk oleh sel-sel yang terletak di titik-titik tertentu pada tumbuhan, terutama titik tumbuh di bagian pucuk tunas maupun ujung akar. Selanjutnya, hormon akan bekerja pada jaringan di sekitarnya atau, lebih umum, ditranslokasi ke bagian tumbuhan yang lain untuk aktif bekerja di sana. Pergerakan hormon dapat terjadi melalui pembuluh tapis, pembuluh kayu, maupun ruang-ruang antarsel. Hormon dalam menjalankan perannya, dapat berperan secara tunggal maupun dalam koordinasi dengan kelompok hormon lainnya. Oleh karena itu, perlu adanya pembahan tersendiri megenai hormone pada tumbuhan mengingat fungsinya yang penting bagi kehidupan tumbuhan.
B. Rumusan Masalah
Rumusan makalah ini adalah sebagai berikut. 1. Apa saja hormon pada tumbuhan ?
2. Apa fungsi utama masing-masing kelompok hormon tumbuhan ?
3. Bagaimana mekanisme kerja antar hormon tumbuhan dalam mengatur proses pertumbuhan dan perkembangan tumbuhan ?
C. Manfaat
Manfaat makalah ini adalah sebagai berikut. 1. Mengetahui hormon pada tumbuhan
2. Mengetahui fungsi utama masing-masing kelompok hormon tumbuhan
3. Mengetahui mekanisme kerja antar hormon tumbuhan dalam mengatur proses pertumbuhan dan perkembangan tumbuhan ?
BAB II KAJIAN TEORI
A. HORMON AUKSIN a. Hormon Auksin
Istilah auksin ( dari bahasa Yunani auxien, “meningkatkan” ) pertama kali digunakan oleh Frits Went, seorang mahasiswa pascasarjana di negeri Belanda pada tahun 1926 yang menemukan bahwa suatu senyawa yang belum dapat diketahui mungkin menyebabkan pembengkokan ini, yang disebut fototropisme. Senyawa yang ditemukan Went didapati cukup banyak di ujung koleoptil dan menunjukkan upaya Went untuk menjelaskan hal tersebut. Hal penting yang ingin diperlihatkan bahwa bahan tersebut berdifusi dari ujung koleoptil menuju potongan kecil. Aktivitas auksin dilacak melalui pembengkokan koleoptil yang terjadi akibat terpacunya pemanjangan pada sisi tumbuhan (Taiz dan Zeiger, 2002 ).
Auksin adalah zat hormon tumbuhan yang ditemukan pada ujung batang, akar, dan pembentukan bunga yang berfungsi untuk sebagai pengatur pembesaran sel dan memicu pemanjangan sel di daerah belakang meristem ujung. Auksin berperan penting dalam pertumbuhan tumbuhan. Peran auksin pertama kali ditemukan oleh ilmuwan Belanda bernama
Fritz Went (1903-1990). Hormon auksin merupakan zat pengatur tumbuh yang mempengaruhi
pemanjangan koleoptil gandum, yang telah dikemukakan oleh Charles Darwin pada abad ke-19. Percobaan definitive yang membuktikan adanya zat yang berdifusi dan merangsang pembesaran sel, telah dikerjakan oleh Fritz Went di Holand pada tahun 1920, dan pada tahun 1930 struktur dan identitas auksin diketahui sebagai asam indol-3-asetat (IAA) (Taiz dan Zeiger, 2002 ).
IAA (auksin) adalah zat yang mempunyai sifat khas yaitu mendorong perpanjangan sel pucuk (Surachmat Kusumo, 1984). Secara keseluruhan, IAA akan meningkatkan tinggi tanaman, kecuali pada konsentrasi IAA yang diberikan tidak optimal. Pemberian yang tidak optimal itu justru akan menghambat pertumbuhan tanaman itu sendiri (Hopkins, 1995). Noggle dan Fritz (1983) menambahkan bahwa pemberian IAA akan dapat meningkatkan pemanjangan sel terutama ke arah vertikal sehingga akan meningkatkan tinggi tanaman (Taiz dan Zeiger, 2002 ).
Auksin disintesis di pucuk batang dekat meristem pucuk, jaringan muda (misal, daun muda), dan selalu bergerak ke arah bawah batang (polar), sehingga terjadi perbedaan auksin di ujung batang dan di akar. Auksin banyak diproduksi di jaringan meristem pada bagian ujung-ujung tumbuhan, seperti kuncup bunga, pucuk daun dan ujung-ujung batang. Selain itu di embrio biji. Auksin tersebut disebarkan ke seluruh bagian tumbuhan, tetapi tidak semua bagian mendapat bagian yang sama. Bagian yang jauh dari ujung akan mendapatkan auksin lebih sedikit. Aktivitasnya meliputi perangsangan dan penghambatan pertumbuhan, tergantung pada
konsentrasi auksinnya. Jaringan yang berbeda memberikan respon yang berbeda pula terhadap kadar auksin yang merangsang atau menghambat pertumbuhan tanaman (Taiz dan Zeiger, 2002 ). Auksin dan pemanjangan sel, meristem apikal suatu tunas merupakan tempat utama sintesis auksin. Karena auksin dari apeks tunas bergerak turun ke daerah pemanjangan sel, sehingga hormon akan merangsang pertumbuhan sel – sel tersebut. Auksin berpengaruh hanya pada kisaran konsentrasi tertentu, yaitu sekitar 10 -8 sampai 10-3 M. Pada konsentrasi yang lebih tinggi, auksin bisa menghambat pemanjangan sel. Hal ini disebabkan oleh tingginya level auksin yang menginduksi sintesis hormon lain, yaitu etilen, yang umumnya bekerja sebagai inhibitor pertumbuhan tumbuhan akibat pemanjangan sel. Jika terkena cahaya matahari, auksin akan mengalami kerusakan sehingga menghambat pertumbuhan tumbuhan. Hal ini menyebabkan batang membelok ke arah datangnya cahaya karena pertumbuhan bagian yang tidak terkena cahaya, lebih cepat dari pada bagian yang terkena cahaya. Menurut Waier at al. (1982), struktur kimia IAA adalah sebagai berikut:
b. Biosintises Asam Indol-3-Asetat
Hormon IAA disintesis dari triptofan hasil-hasil intermediet yang terdapat antara triptofan dan IAA adalah asam amino indol piruvat, triptoamin, dan indol asetaldehida (Davies, 1995). Triptofan sendiri terbentuk dari PEP (fosfoenol piruvat) dan eritrosa-4-fosfat. Jalur biosintesis IAA mulai dari PEP sampai dengan triptofan juga merupakan jalur biosintesis dari senyawa-senyawa fenolik IAA juga dapat dibentuk secara langsung dari asam amino serine dengan indol (Wattimena, 1988). Sedangkan menurut Salisbury dan Ross (1995), mekanisme pembentukan IAA pada jaringan tumbuhan adalah sebagai berikut:
c. Transpor Asam Indol-3-Asetat
Pengangkutan auksin pada tumbuhan disebut pengangkutan polar. Arah pengangkutan auksin pada batang adalah basipetal, yaitu dari ujung ke basal, sedangkan pada akar adalah akropetal, yaitu dari basal ke ujung. IAA tidak hanya dipindahkan lewat sistem pembuluh tapis floem maupum xilem, tetapi dapat melalui parenkim yang bersinggungan dengan berkas pembuluh. Pergerakan auksin adalah lambat. Pengangkutan ini memerlukan energi yang berupa ATP terhambat (tidak ada oksigen). Senyawa penghambat kuat pengangkutan auksin (anti auksin) adalah asam 2,3,5-triidobenzoat (TIBA) dan asam a-naftiltamat (Salisbury dan Ross, 1995). Selanjutnya dikatakan oleh Salisbury dan Ross (1995) bahwa pengangkutan auksin secara polar terjadi dengan menggunakan ATP-ase membran plasma pada sel untuk memompa H + dari sitosol menuju dinding sel. Nilai pH dinding sel yang lebih rendah (sekitar 5) menyebabkan dipertahankannya gugus karbolsil auksin yang di sitosol (dengan pH 7 hingga 7,5). Auksin yang tidak bermuatan bergerak dari dinding menuju ke sitosol melalui ko-transpor dengan ion H+. Nilai pH sitosol yang lebih tinggi menyebabkan gugus karbolsil auksin terdisosiasi menjadi bermuatan negatif. Peningkatan konsentrasi auksin bermuatan di sitosol menyebabkan
d. Peran IAA dalam Pertumbuhan dan Deferensiasi
Respon auksin pada tanaman dapat bersifat menghambat apabila dalam konsentrasi tinggi, respon auksin dipengaruhi oleh konsentrasinya. Respon auksin terhadap pertumbuhan sangat bervariasi tergantung pada kepekaan organ tanaman (Gardner et al., 1991).
.
e. Fungsi Hormon Auksin
Secara umum fungsi hormon auksin pada tumbuhan yaitu: 1. Berperan dalam pembelahan dan pembesaran sel 2. Merangasng pembelahan kambium vaskuler
3. Memicu terbentuknya akar lateral dan akar adventif dan memicu pertumbuhan akar dengan lebih baik
4. Berperan dalam pertumbuhan tunas samping 5. Mengurangi jumlah biji dalam buah
6. Merangsang persentase terbentuknya buah dan bunga 7. Berperan dalam mengurangi gugur buah
8. Memecah dormansi benih dan merangsang proses perkecambahan, serta dapat memecah dormansi pucuk/apikal agar dapat berkembang.
9. Merangsang terjadinya partenokarpi (kemampuan tumbuhan untuk mebentuk buah tanpa proses penyerbukan).
Gambar. Mekanisme Kerja Auksin (Taiz dan Zeiger, 2002 ).
Mekanisme kerja auksin sangat dipengaruhi oleh cahaya dan aktif jika tidak terkena cahaya. Hal ini menyebabkan fototropisme pada tumbuhan. Fototropisme merupakan peristiwa membengkoknya batang tanaman ke arah cahaya. Hal ini terjadi karena adanya perbedaan rangsangan perpanjangan sel akibat penyebaran auksin yang tidak merata dan tidak diproduksinya auksin pada bagian yang terkena cahaya. Bagian yang tidak terkena cahaya aktif memproduksi auksin sehingga terjadi penimbunan auksin. Penimbunan auksin pada sisi yang tidak terkena cahaya ini mengakibatkan pemanjangan sel di sisi tersebut lebih cepat sehingga batang membengkok ke arah datangnya cahaya.
Selain fototropisme, auksin juga mempengaruhi peristiwa geotropisme atau pertumbuhan ke arah bumi. Gaya gravitasi menyebabkan konsentrasi auksin di bagian bawah lebih tinggi sehingga geotropisme di batang menjadi negatif dan geotropisme di bagian akar menjadi positif sehingga akar membelok ke arah bumi.
Gambar. Mekanisme Kerja Auksin (Taiz dan Zeiger, 2002 ). g. Proses Pengangkutan Auksin
Cara kerja hormon Auksin adalah menginisiasi pemanjangan sel dan juga memacu protein tertentu yang ada di membran plasma sel tumbuhan untuk memompa ion H+ ke dinding sel. Ion H+ mengaktifkan enzim ter-tentu sehingga memutuskan beberapa ikatan silang hidrogen rantai molekul selulosa penyusun dinding sel. Sel tumbuhan kemudian memanjang akibat air yang masuk secara osmosis. Auksin merupakan salah satu hormon tanaman yang dapat meregulasi banyak proses fisiologi, seperti pertumbuhan, pembelahan dan diferensiasi sel serta sintesa protein Auksin (Taiz dan Zeiger, 2002 ).
Auksin diproduksi dalam jaringan meristimatik yang aktif (yaitu tunas, daun muda dan buah). Kemudian auxin menyebar luas dalam seluruh tubuh tanaman, penyebarluasannya dengan arah dari atas ke bawah hingga titik tumbuh akar, melalui jaringan pembuluh tapis (floom) atau jaringan parenkhim. Auksin atau dikenal juga dengan IAA = Asam Indolasetat (yaitu sebagai auxin utama pada tanaman), dibiosintesis dari asam amino prekursor triptopan, dengan hasil perantara sejumlah substansi yang secara alami mirip auxin (analog) tetapi mempunyai aktifitas lebih kecil dari IAA seperti IAN = Indolaseto nitril, TpyA = Asam Indolpiruvat dan IAAld = Indolasetatdehid. Proses biosintesis auxin dibantu oleh enzim IAA-oksidase.
Cara pengangkutan auksin memiliki keistimewaan yang berbeda dengan pengangkutan floem, di antaranya :
2. Pengangkutan auksin berlangsung secara polar. Pada batang auksin ditransport secara basipetal (away from apex), sedangkan pada akar, transport auksin secara akropetal ke arah ujung melalui parenkim vaskuler.
3. Pergerakan auksin memerlukan energi metabolisme, seperti ditunjukkan oleh kemampuan zat penghambat sintesis ATP atau keadaan kurang oksigen dalam menghambat pergerakan itu.
B. HORMON GIBERELIN a. Hormon Giberelin
Asam giberelat atau Gibberellic Acid ditemukan di Jepang dalam penelitian tentang penyakit padii yang disebut penyakit “bakanae”. Bakanae ini disebabkan oleh jamur Gibberellafujikuroi, yang menunjukan bahwa batang dan daun padi yang memanjang secara tidak normal (Gadner et al. 1991; Cleland, 1989). Pada tahun 1930-an, Yabuta dan Hayashi memisahkan suatu senyawa aktif dari cendawan tersebut dan diberi nama giberelin (Salisbury dan Ross, 1995).
Asam Giberelat adalah zat kimia yang dikelompokkan ke dalam terpenoid. Semua asam giberelat termasuk asam diterpenoid tetrasiklik dan merupakan turunan rangka ent-giberelan (Davies, 1995). Semua giberelin bersifat asam dan dinamakan GA ( asam gibetrelat ) yang dinomori untuk membeda-bedakannya. Semua giberelin mempunyai 19 dan 20 atom karbon, yang bergabung dalam sistem cincin 4 atau 5. Giberelin dinomori seperti pada ent-giberelin, kecuali gugus metil karbon 20 pada ent-giberelin yang telah teroksidasi dan kemudian lepas menjadi CO2 menjadi giberelin 19-karbon. Sistem cincin kelima ( tidak terdapat pada giberelin ) adalah cincin lakton menempel pada cincin A giberelin. Gugus metil karbon 19 ent-giberelin teroksidasi menjadi karboksil dan kemudian digunakan untuk membentuk cincin lakton ( Salisbury dan Ross, 1995). Semua kelompok terpenoid terbentuk dari unit isoprene yang terdiri dari lima atom karbon (Abidin, 1990). Bagian dasar kimia asam giberelat adalah kerangka giban dan kelompok karboksil bebas (Gardner et al., 1991). Menurut Salisbury dan Ross (1995), struktur rangka ent-giberelan adalah sebagai berikut,
Gambar.
Struktur rangka ent-giberelan. (Salisbury dan Ross, 1995) Menurut Weayer dalam Abidin (1990) perbedaan utama pada giberelin meliputi:
a. Beberapa giberelin mempunyai 19 buah atom karbon dan yang lainnya mempunyai 20 atom karbon
b. Gugus hidroksil berada dalam posisi 3 dan 13.
c. Semua giberelin dengan 19 atom karbon adalah asam monohidroksil yang mengandung gugus COOH pada posisi 7 dan mempunyai cincin lakton.
Tiap jenis tanaman mempunyai beberapa asam giberelat tertentu (Abidin, 1990). Macam-macam bentuk asam giberelat berbeda dapat juga karena penggantian kelompok-kelompok hidroksil, metil atau etil pada kerangka giban dan karena adanya cincin laktona yang dihasilkan oleh kondensasi karbon 20 ke karbon 19 dalam struktur giban. Adanya cincin laktona menyebabkan aktivitas biologis yang lebih besar dibandingkan dengan yang tidak mempunyai cincin laktona (Gardner et a., 1991). Menurut Hopkins ( 2009 ), struktur kimia dari GA3 dan GA1 yang merupakan salah satu jenis asam giberelat yang aktif serta GA8 tidak aktif karena penambahan kelompok hidroksil dalam 2 posisi adalah sebagai berikut.
Gambar. Struktur Kimia GA3, GA1 ( aktif ) dan GA8 (tidak aktif ) (Hopkins, 2009) b. Biosintesis Asam Giberelat
Biosintesis berbagai asam giberelat terutama berlangsung di dalam buah dan biji yang sedang berkembang, dalam tunas, dan akar (Davies, 1995). Walaupun asam giberelat diketahui menghambat pertumbuhan akar, akar merupakan sumber asam giberelat bagi organ-organ yang lain (Gardner et.al., 1991). Asam giberelat merupakan diterpen yang disintesis dari unit-unit asetat asetil ko-enzim A oleh lintasan asam mevalonat. Genarilgenaril pirofosfat yaitu senyawa 20-karbon bertindak sebagai donor bagi semua atom karbon pada giberelin. Senyawa itu diubah menjadi kopalilpirofosfat yang memiliki sistem dua cincn dan senyawa terakhir tersebut kemudian diubah menjadi kauren yang mempunyai sistem empat cincin. Perubahan lebih lanjut disepanjang lintasan meliputi oksidasi yang terjadi di reticulum endoplasma, menghasilkan senyawa antara kaurenol, kaurenal dan asam kaurenoat (Salisbury dan Ross, 1995).
Aseto aseti ko-enzim A dan asetil ko-enzim A diubah menjadi hidroksimetil glutaril ko-enzim A. Hidroksimetil glutaril ko- enzim A diubah menjadi asam mevalonat yang selanjutnya diubah menjadi mevalonat-5-pirofosfat. Mevalonat-5-pirofosfat dikarboksilasi menjadi isopentenil pirofosfat. Isopentenil pirofosfat diubah menjadi dimetilalil pirofosfat yang merupakan unit
pemula untuk biosintesis terpen. Dimetilalil pirofosfat diubah menjadi geranil pirofosfat yang selanjutnya diubah menjadi farnesil pirofosfat. Kondensasi farnesil pirofosfat dengan 3 molekul isopentenil pirofosfat menghasilkan geranil-geranil pirofosfat (Davies, 1995). Geranil-geranil pirofosfat merupakan senyawa denga 20 atom karbon yang berperan sebagai pemberi (donor) untuk semua atom karbon pada asam giberelat. Senyawa itu diubah menjadi kopalil pirofosfat yang mempunyai dua sistem cincin, lalu diubah menjadii kauren. Perubahan kauren lebih lanjut di sepanjang lintasan meliputi oksidasi yang terjadi di retikulum endoplasma. Lintasan ini menghasilkan senyawa antara seperti kaurenol (suatu senyawa alkohol), kaurenal (senyawa aldehid) dan asam kaurenoat. Dari senyawa ini akan disintesis asam giberelat lain dengan 19 dan 20 atom karbon. Beberapa reaksi dalam biosintesis asam giberelat adalah
Gambar. Beberapa reaksi dalam biosintesis asam giberelat (Salisbury dan Ross, 1995) Senyawa pertama dengan sistem cincin giberelan yang sejati adalah aldehid GA12, suatu molekul 20-karbon. Dari senyawa itu terbentuk giberelin 20-karbon dan giberelin 19-karbon. Aldehid GA12 terbentuk dengan cara menerobos salah satu karbon cincin B pada asam kaurenoat dan mengerutkan cincin tersebut (Salisbury dan Ross, 1995).
Umumnya giberelin 19-karbon lebih aktif daripada giberelin 20-karbon dan karbon yang hilang dari molekul 20-karbon adalah karbon dari gugus metil yang menempel diantara cincin A dan cincin B aldehid GA12. Karbon tersebut teroksidasi menjadi gugus karboksil yang kemudian lepas menjadi CO2. Pada sebagian besar giberelin cincin kelima ( lakton ) terbentuk dari karbon 19 gugus karboksil pada aldehid GA12 untuk menghasilkan GA9. Zat perlambat pertumbuhan tertentu yang diperdagangkan menghambat pemanjangan batang dan menyebabkan pengkerdilan bekerja untuk menghambat giberelin. Produk tersebut meliputi Phosphon D, Amo-1618, CCC atau Cycocel, ansimidol, dan paklobutrazol. Phosphon D menghambat proses selanjutnya yakni pembentukan kauren, sedangkan ansimidol dan paklobutrazol menghambat reaksi oksidasi antara kauren dan asam kaurenoat. Pada banyak jenis tumbuhan penghambat tumbuh oleh salah satu zat dapat diatasi oleh GA3 yang efek utamanya adalah menghambat sintesis gibelin (Salisbury dan Ross, 1995).
c. Bagian Tumbuhan yang Mensintesis Giberelin
Biji yang belum matang mengandung giberelin dalam jumlah yang cukup tinggi dibandingkan pada bagian tubuh lainnya. Sebagian besar kandungan giberelin yang tinggi di dalam biji dihasilkan dari biosintesis bukan diangkut ke bagian tersebut. Kemampuan bagian tumbuhan mensintesis giberelin kurang begitu dipahami sebab hanya sedikit data langsung yang tersedia mengenai biokimianya. Walaupun demikian, sebagian besar sel tumbuhan mungkin mampu mensistesis giberelin (Salisbury dan Ross, 1995).
Daun muda diduga menjadi tempat utama sintesis giberelin seperti halnya auksin. Apabila ujung tajuk dan daun muda dipangkas dan ditunggul batangnya diberi auksin atau giberelin maka pemanjangan batang terpacu jika dibandingkan dengan batang terpotong yang tidak diberi hormone. Daun muda dapat memacu pemanjangan batang karena daun muda mengirim kedua jenis hormone tersebut ke batang. Akar juga mensintesis giberelin namun giberelin eksogen menimbulkan efek kecil pada pertumbuhan akar dan menghambat pembentukan akar liar (Salisbury dan Ross, 1995).
d. Pengangkutan Makanan dan Unsur Mineral dalam Sel Penyimpanan pada Biji yang Dipacu oleh Giberelin
Segera setelah biji berkecambah sistem akar dan tajuk muda mulai menggunakan hara mineral, lemak, pati dan protein yang terdapat di sel penyimpanan pada biji. Kecambah muda
bergantung pada cadangan makanan ini sebelum mampu menyerap garam mineral dari tanah dan sebelum memanjangkan sistem tajuknya menuju cahaya. Garam mineral dapat mudah berpindah dalam melalui floem menuju ke seluruh akar dan tajuk muda, jika garam tersebut bersifat lasak. Kecambah menghadapi kesulitan dengan lemak, polisakarida, dan protein sebab molekul tersebut tidak dapat dipindahkan ( Salisbury dan Ross, 1995 ).
Embrio ( nutfah ) biji serelia-bulir dan rumput lainnya dikelilingi cadangan makanan yang terdapat di sel yang secara metabolic tidak aktif yakni endosperma. Endosperma sendiri diselimuti lapisan tipis yang hidup, yang biasanya mempunyai ketebalan dua hingga emat sel disebut selaput aleuron. Setelah perkecambahan terjadi terutama akibat meningkatnya kelembapan, sel aleuron mengelurakan sejumlah enzim hidrolisis yang mencerna pati, protein, fitin, RNA dan bahan dinding sel tertentu yang terdapat dalam endosperma ( Salisbury dan Ross, 1995 ).
Salah satu enzim yang diperlukan dalam proses pencernaan ialah α-amilase yang dapat menghidrolisis pati. Jika embrio dihilangkan dari biji, sel aleuron tidak menghasilkan dan mengelurakan sebagian besar enzim hidrolitiknya, termasuk α-amilase. Hal itu memperlihatkan bahwa embrio biasanya mengeluarkan menyediakan suatu hormon untuk selaput aleuron dan memacu sel aleuron untuk membuat enzim hidrolitik. Hormon giberelin mendorong sekresi enzim hidrolitik ke endosperma tempat enzim mencerna cadangan makanan dan dinding sel. Unsur mineral cadangan menjadi lebih muda tersedia sebagai hasil kerja giberelin ( Salisbury dan Ross, 1995 ).
Gambar. Skema Ilustrasi Sintesis Giberelin selama Perkecambahan Benih ( Hopkins, 2009 ).
Terjadinya imbibisi air merangsang sintesis GA. Giberelin bergerak dari embrio (1) ke aleuron di mana merangsang sintesis enzim α-amilase dan protease. (2) protease mengkonversi β-amilase yang semula tidak aktif untuk aktif. Enzim α- dan β-amilase bersama-sama mencerna pati menjadi glukosa, (3) Glukosa digunakan untuk memenuhi metabolisme embrio ( Hopkins, 2009 ).
Pada biji rumputan giberelin disintesis di skuletum ( kotiledon ) dan mungkin dibagian lain pada embrio. Walaupun selaput aleuron bertanggung jawab dalam memproduksi enzim yang mencerna sebagian cadangan makanan di endosperma, terdapat bukti bahwa di skutelum juga mengeluarkan enzim pencerna. Sisi skuletum yang menghadap endosperma tersusun dari selapis sel kolumna yang struktur dalamnya kaya akan reticulum endoplasma dan diktiosom yang khas pada sel yang melakukan sekresi. Terbukti bahwa skuletum lebih penting daripada aleuron dalam beberapa spesies ( Salisbury dan Ross, 1995 ).
e. Peranan Asam Giberelat dalam Pertumbuhan
Asam giberelat sebagai hormon tumbuh pada tanaman berpengaruh terhadap pembungaan, penyinaran, partenokarpi, mobilisasi karbohidrat selama perkecambahan dan aspek fisiologis lainnya. Asam giberelat dapat menyebabkan pemanjangan batang dengan merangsang pembelahan dan pemanjangan sel, sehingga dapat diperoleh tanaman yang lebih tinggi (Davies, 1995). Asam Giberelat memacu pembentangan sel melalui beberapa kemungkinan yaitu meningkatkan potensial osmosis sel, tekanan potensial dinding sel atau turgor, dan permeabilitas membran yang akan menyebabkan osmosis sel, sehingga terjadi pembentangan sel (Kende and Zeevaart, 1997; Kende et al., 1995). Asam giberelat juga dapat menyebabkan pemanjangan batang dengan mempengaruhi respon tanaman terhadap panjang hari penyinaran (Davies, 1995). Hormon GA3 dapat menyebabkan perkecambahan biji pada beberapa biji yang secara normal membutuhkan suhu dingin atau cahaya untuk menginduksinya (Davies, 1995). Pemberian hormon eksogen giberelin dapat meningkatkan pertumbuhan dan senyawa metabolit sekunder. Giberelin selain menambah tinggi tanaman juga menambah luas daun dan berat kering tanaman, dan pertambahan berat kering merupakan hasil peningkatan aktifitas fotosintesis (Surachmat, 1984). Faktor yang 50 mempengaruhi pertumbuhan adalah faktor internal yaitu gen dan hormon serta faktor eksternal yaitu suhu, cahaya, ketersediaan air, ketinggian, kecepatan angin, iklim dan suhu sedangkan faktor utama yang mengendalikan penyebaran dan struktur sebagian komunitas tumbuhan alami di daerah tropik adalah air (Loveless, 1999). Fungsi hormone giberelin adalah sebaga berikut.
a) Mematahkan dormansi atau hambatan pertumbuhan tanaman sehingga tanaman dapat tumbuh normal dengan cara mempercepat proses pembelahan sel.
b) Merangsang pemanjangan sel. c) Meningkatkan proses pembungaan.
d) Menyebabkan perkembangan buah tanpa benih/partenokapri. e) Dapat menunda penuaan daun dan buah.
f. Kombinasi IAA dan GA3 Terhadap Pertumbuhan dan Perkembangan
Variasi musim dan faktor kelembaban sangat penting bagi vegetasi dan situasi ekstrim yang kadang–kadang terjadi variasi ini memiliki arti lebih penting dari pada nilai rata–rata dalam menentukan kehadiran atau ketidak hadiran jenis tertentu (Loveless, 1999). Lingkungan tempat tumbuh, diferensiasi sel dan jaringan sangat berpengaruh terhadap pembentukan senyawa metabolit sekunder. Pengetahuan tentang hubungan antara pertumbuhan, diferensiasi sel dan peningkatan senyawa metabolit sekunder tertentu dapat digunakan sebagai acuan untuk penelitian tentang pengaruh pemberian GA3 dan IAA untuk memanipulasi metabolisme tanaman sehingga didapat peningkatan pertumbuhan, peningkatan jumlah sel–sel sekretori dan peningkatan senyawa metabolit sekunder diantaranya saponin. Secara alamiah tanaman sudah mengandung hormon pertumbuhan seperti auksin, giberelin dan sitokinin (hormon endogen). Hormon endogen tanaman berada pada jaringan jaringan meristem yaitu jaringan yang aktif tumbuh seperti ujung-ujung tunas atau tajuk dan akar. Tetapi karena pola budidaya yang intensif disertai pengelolaan tanah yang kurang tepat, maka kandungan hormon endogen tersebut menjadi berkurang atau rendah bagi proses pertumbuhan vegetatif dan generatif tanaman sehingga dijumpai tanaman tumbuh lambat, kerdil, umbi kecil dan sebagainya selain disebabkan karena kurang unsur hara. Oleh karenanya diperlukan penambahan hormon eksogen untuk menghasilkan pertumbuhan tanaman yang optimal. Jenis hormon eksogen antara lain auksin akan meningkatkan permeabilitas dinding sel yang akan mempertinggi penyerapan unsur utamanya yaitu N, Mg, Fe, Cu untuk membentuk klorofil yang sangat diperlukan untuk mempertinggi fotosintesis. Dengan fotosintesis yang semakin meningkat, akan dihasilkan fotosintet yang meningkat. Auksin akan bergerak ke akar memacu pembentukan giberelin yang akan membantu pembentukan akar. Penambahan kandungan auksin eksogen akan meningkatkan tekanan turgor akar sehingga giberelin dan sitokinin endogen di akar diangkut ke tajuk tanaman, sehingga terjadi peningkatan ukuran sel dan hasil fotosintesis yang meningkat pula. Pada awal pertumbuhan auksin akan mempercepat proses pertumbuhan vegetatif tanaman dan mengatasi kekerdilan tanaman. Bersamaan dengan proses pertumbuhan vegetatif maka hasil fotosintesis akan meningkat pula termasuk peningkatan senyawa kimia metabolit sekunder. Perubahan kandungan giberelin eksogen akan meningkatkan perbandingan C/N. Giberelin eksogen pada fase generatif mampu memperbesar sel jaringan penyimpanan sehingga mampu menyimpan hasil-hasil fotosintesis lebih banyak yang berakibat pada lebih besarnya ukuran jaringan penyimpanan (buah, bulir maupun umbi). Giberelin berperan pada pemanjangan sel melalui:
1. Peningkatan kadar auksin, yakni akan memacu pembentukan enzim yang melunakan dinding sel, terutama enzim proteolitik yang akan melepas amino triptofan sebagai prekusor auksin sehingga kadar auksin meningkat.
2. Merangsang pembentukan polihidroksi asam sinamat, yaitu senyawa yang menghambat kerja enzim IAA oksidase yang merupakan enzim perusak auksin, giberelin merangsang terbentuknya enzim α-amilase yang akan menghidrolisis pati sehingga kadar gula dalam sel akan naik dan menyebabkan air lebih banyak lagi masuk ke dalam sel yang mengakibatkan sel memanjang. C. HORMON SITOKININ
Cytokinin (CK) merupakan derivate dari basa nitrogen adenine yang fungsi utamanya untuk merangsang pembelahan sel dalam kultur jaringan. Sitokinin ini juga mempengaruhi beberapa respon perkembangan tumbuhan diantaranya yaitu diferensiasi akar dan tunas dalam kultur jaringan, pertumbuhan tunas lateral, perluasan daun, perkembangan kloroplast dan penundaan penuaaan. Dan akhir-akhir ini juga telah dipelajari bahwa sitokinin memerankan peranan penting dalam menjaga meristem tunas apical. Berikut ini penjelasan lebih lanjut mengenai hormon Sitokinin
a. Sitokinin Merupakan Derivat Adenin
Semua Sitokinin alami merupakan turunan dari adenine baik rantai sampingnya berikatan dengan isoprena ataupun aromatic (siklik). Sitokinin yang rantai sampingnya isoprena disebut dengan sitokinin isoprenoid dan sitokinin yang rantai sampingnya berikatan dengan aromatic disebut juga dengan sitokinin aromatic. Meskipun ada juga beberapa variasi yang tergantung spesies. Pada tahap perkembangan, sitokinin isoprenoid yang paling umum adalah N6 –(∆2 – isopentenyl)- adenine (iP), trans-zeatin (tZ) dan dihydrozearin (DZ). Kinetin ini merupakan senyawa pertama yang ditemukan pada aktivitas sitokinin yang merupakan sitokinin sintetik dan dibuat dengan pemanasan DNA. Kinetin ini merupakan turunan sintesis yang belum diidentifikasi pada tanaman dan laporan terbaru menyatakan bahwa kinetin ini telah diidentifikasi dalam urin manusia. Isopentenyl adenine (Ip) dan trans-Zeatin merupakan jenis sitokinin isoprenoid dan termasuk ke dalam sitokinin alami. Untuk Benziladenin (BAP) termasuk ke dalam sitokinin aromatic. Benzyladenin ini dapat ditemukan hanya dalam beberapa spesies tumbuhan saja. Berikut ini gambar macam-macam sitokinin isoprenoid
Gambar Contoh Struktur Kimia Sitokinin (Sumber : Hopkins, 2009)
1. Biosintesis Sitokinin Dimulai Dengan Kondensasi Kelompok Isopentenyl Dengan Kelompok Amino Adeno Monofosfat
Enzim yang mengarahkan dalam sintesis sitokinin diisolasi dari jamur lender Dictyostellum discoideum, jaringan kalus tembakau dan jaringan mahkota empedu. Mahkota empedu ini merupakan neoplastik atau tumor. Dimana dalam gambar di bawah ini ditunjukkan pada batang Bryophyllum. Mahkota tersebut merupakan hasil dari infeksi dari bakteri Agrobacterium tumefaclens, dimana bakteri tersebut mengubah sel-sel tanaman inang dengan gen bakteri sehingga menyebabkan kelebihan produksi auksin dan sitokinin. Berikut ini gambarnya
Sumber : Hopkin, 2009
Reaksi utama dalam biosintesis ini yaitu adisi kelompok dimethyallyl difosfate (DMAP) dengan nitrogen pada posisi keenam adenosine 5 monofosfat (AMP). Reaksi adisi tersebut dikatalis oleh enzim adenosine fosfar isopentenil transferase (IPT) dan produk yang dihasilkan yaitu N6 -(∆2-isopentenyl)-adenosin-5’ monofosfat (iPRMP). iPT ini dikatalis juga untuk
membatasi dalam proses biosintesis sitokinin. Karena terdapat factor yang memungkinkan untuk memanipulasi konten sitokinin jaringan dapat mengubah tanaman dengan gen dan menyebabkan tumbuhan mengandung iPT yang berlebih. Dan tahap kedua yaitu kelompok fosfat dan grup ribose dirubah dari {9R-5’P}iP menjadi bentuk sitokinin aktif N6 –(∆2-isopentenyl)-adenin (iP). Dan kemungkinan lain, rantai samping isopentenyl pada {9R-5’P}Ip di hidroxilkan sebelum grup fosfat dan ribose berubah menjadi bentuk zeatin (Z). Zeatin dan iP ini merupakan sitokinin biologis yang paling aktif dalam kebanyakan tumbuhan. Dan reduksi ikatan rangkap rantai samping zeatin akan memberikan turunan dihydrozeatin, yang sangat aktif dalam beberapa spesies legume. Berikut ini reaksi yang terjadi pada biosintesis sitokinin.
. Gambar Reaksi Biosintesis Isopentenyl adenine (IP)
Sumber : Hopkins, 2009
Dari gambar diatas dapat diketahui bahwa secara garis besar biosintesis isopentenyl adenine (IP) dan trans-zeatin (TZ) dari adenosine monofosfar dan isopentenil pirofosfar. Dimana adenosine difosfat dan adenosine trifosfate juga dapat digunakan sebagai precursor. Reaksi 1 merupakan langkah pembatasan dan dikatalisis oleh enzim adenine fosfat isopentenyl transferase (IPT).
Sitokinin ini juga dikenal untuk menjalani interconversions luas antara basa bebas (necleobase), ribosides dan ribotides ketika dieksperimenkan dalam jaringan. Dan enzim yang telah diidentifikasi dalam gandum untuk mengkatalis konversi Ip yaitu ribosidenya ({9R}iP) atau
ribotide ([9R-5’P]iP), serta enzim tersebut yang mengkatalis hidrolisis ke basa yang tidak berikatan (iP).
2. Sitokinin Dapat Dinonaktifkan dengan Konjugasi Atau Oksidasi
Ada dua rute utama untuk mengatur tingkat aktivitas sitokinin oleh penghapusan sitokinin dari kelompok yang aktif yaitu Konjugasi dengan glukosa atau asam amino lain dan Oksidasi. Contohnya yaitu pada glikosilasi dari kelompok rantai samping hidroksil (O-glikolisasi) dari zeatin atau dihydrozeatin misalnya yang melimpah pada tanaman.O-glikosidase ini tidak sendiri dalam mengaktifkan, tetapi mudah dihidrolisis untuk mengaktifkan sitokinin oleh enzi glukosidase. Dan O-glikosilasi ini merupakan mekanisme untuk menyimpan kelebihan sitokinin yang akan digunakan saat kondisi fisiologi membutuhkan. O-glukisilasi ini juga tahan terhadap oksidasi dan berfungsi melindungi hormone dari oksidasi ketika sedang diangkut ke jaringan target. Dalam beberapa tanaman, seperti lobak, jagung atau tembakau, sitokinin juga dapat glikolisasi di salah satu posisi nitrogen pada cincin purin. Kedua glikosida 7 dan 9 ini secara tidak aktif. N-glikosida juga sangat stabil dan tidak muncul untuk mudah dihidrolisis yang akan memberikan basa bebas aktif. Bentuk dari mereka sehingga menampakkan sarana untuk penonaktifan permanen sitokinin daripada penyimpanan.
Sitokinin ini juga membentuk konjugasi dengan asam amino alanin. 9-Alanyl konjugat dari zeatin dan dihydrozeatin telah diidentifikasi dalam jaringan lupinus, buah dan akar nodul, biji apel yang belum matang dan bibit kacang. Konjugat ini sangat stabil dan memungkinkan berfungsi untuk menonaktifkan sitokinin dalam cara yang sama seperti N-glukosida. Berikut ini gambar struktur kimianya
Gambar Struktur Kimia (Sumber : Hopkins, 2009)
Mekanisme lain dan utama untuk menghilangkan sitokinin dari kelompok hormone yang degradasi irreversible yaitu dengan menggunakan enzim sitokinin oksidase atau dehidroginase (CKX). CKX ini akan memotong rantai samping isopentenyl baik dari zeatin atau IP atau turunan ribosyl mereka. Dalam fungsinya, namun CKX mengenal ikatan rangkap dari sisi samping isopreoid yang akibatnya sitokinin dihydrozeatin dan aromatic tahan terhadap degradasi oleh CKX . Dengan cara yang sama bahwa kandungan sitokinin endogen dapat ditingkatkan lebih dari gen isopentenil transferase (IPT) pada tanaman transgenic, konten sitokinin juga dapat dikurangi dalam tanaman transgenic yang overexpress sitokinin oksidase atau dehidroginasenya.
b. Sitokinin Disintesis Terutama di Akar dan Translokasi pada Xilem
Tempat utama biosintesis sitokinin pada tumbuhan tingkat tinggi berada pada akar. Karena sitonin banyak ditemukan di dalam akar terutama pada ujung akar dan dalam getah xylem akar dari berbagai sumber. Jika tidak semua mengandung sitokinin, maka sitokinin akan diangkut ke aerial tanaman melalui xylem. Dan Jika pada tanaman akarnya dihilangkan atau dipotong maka penuaan daun akan semakin cepat. Dan penuaan akan tertunda ketika akar ada, hal tersebut dikarenakan bahwa adanya sitokinin yang dapat menghambat proses penuaan. Sitokinin ini disentesis di akar dan akan diangkut ke daum melalui jaringan vascular.
Buah yang belum matang dan berkembang buahnya juga mengandung kadar sitokinin yang tinggi. Pertama sitokinin alami akan diisolasi dari endosperm jagung dan mengembangkan buah plum. Sementara ada beberapa bukti bahwa biji dan buah-bahan mampu mensinesis sitokinin dan ada juga yang mengatakan sebaliknya. Pada dasarnya, Biji berkembang karena aktivitas metabolisme sangat tinggi dan pertumbuhan yang cepat memungkinkan biji hanya berfungsi sebagai wastafel untuk sitokinin yang diangkut dari akar.
c. Sitokinin Dibutuhkan Untuk Cell Proliferasi
Peran sitokinin yang jelas adalah dalam mengatur pembelahan sel. Hasil dari peran tersebut dapat dibuktikan dari hasil percobaan isolasi jaringan pada wortel dan tembakau pada media. Proliferasi sel ini terjadi hanya ketika auksin ditambahkan ke beberapa medium sebagai factor pembelahan sel. Factor pembelahan sel tersebut akan menjadi kinetin. Kinetin dan sitokinin lainnya selalu di damping dengan kehadirannya auksin dimana akan diransang pembelahan sel di berbagai jaringan.
1. Sitokinin Mengatur Progresi Melalui Siklus Sel
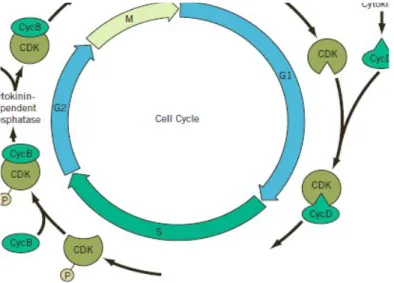
Dalam kultur sel tembakau diperlukan adanya auksin dan sitokinin untuk pembelahan sel lanjutan. Tidak adanya hormone lain menyebabkan sel ditangkap dengan baik dalam G1 atau fase G2 dari siklus sel. Penambahan hormone yang hilang akan menyebabkan pembelahan sel yang dapat dideteksi dalam waktu 12 sampai 24 jam. Pada tahun 1996, K Zhang dan rekan kerjanya melaporkan bahwa sel-sel akan ditangkap di G2 dan tidak adanya sitokinin yang terdapat pada CDK menyebabkan aktivitas berkurang karena terjadi fosforilasi yang tinggi pada residu tirosin. Dan ketika percobaanya diberi sitokinin kembali, tirosin tersebut di fosforilasikan dan enzim diaktifkan kembali serta pembelahan sel dapat diteruskan. Residu tirosin spesifik pada unit katlitik CDK adalah terfosforilasinya selama fase S oleh kinase lain. Meskipun CDK unit katalitik yang terfosforilasi, CDK ini mampu menggabungkan dengan cyclin ( sebuah cyclin tipe B atau CycB). Kompleks tersebut terfosforilasinya tetap tidak aktif sampai inhibitor kelompok fosfat diubah oleh sitokinin yang berikatan dengan fosfatase. Dengan demikian peran utama sitokinnin dalam sel tembakau ini akan menghasilkan kompleks CDK yang aktif dan memulai pembelahan sel dengan menjadi katalis transisi dari fase G2 menuju mitosis (fase M). Berikut ini gambar sederhana untuk mengontrol hormonal dari siklus sel pada tumbuhan. Adanya sitokinin mempromosikan terjadinya mitosis (G2 ke M transisi) dengan mengaktifkan sebuah fosfotase yang menghilangkan kelompok inhibitor fosfat dari CDK atau cyclin B kompleks. Dan adanya auksin serta sitokinin juga mendorong pengumpulan cyclin G1 (Cyclin D) yang diperlukan untuk terjadinya fase S (sintesis)
Sumber : Hopkins, 2009
d. Sitokinin Sebagai Reseptor Dan Signal
Sitokinin reseptor akhirnya ditemukan oleh T.Kakimodo dan rekan-rekannya yang mengembangkan tes hipokotil Arabidopsis untuk scanning mutan. Bagian hipokotil tanaman menanggapi ketika ditambahkan sitokinin dan menghasilkan respon yang khas yaitu adanya proliferasi sel dengan cepat, penghijauan dan pembentukan tunas. Respon sitokinin 1 (CRE 1) mutan tidak terlihat responnya, bahkan ketika konsentrasi sitokinin dinaikkan menjadi 10 kali lipat juga tidak terlihat yang memungkinkan reseptor sitokinin nonfungsional dalam mutan. Percobaan berikutnya menegaskan bahwa jenis protein CRE 1 sebenarnya reseptor sitokinin.
1. Reseptor Sitokinin Merupakan Membran Basa Histidin Kinase
CRE 1 merupakan komponen pertama dari dua komponen sistem pengaturan yang beroperasi di bakteri dan prokariot lainnya. Nama berasal dari konfigurasi bakteri dimana reseptor pertama mengaktifkan pengatur respon (RR) pada komponen kedua. Pengatur respon juga mengatur transkripsi gen target atau memodulasi reaksi metabolism lainnya. Selain sebagai reseptor hormone, dua sistem regulasi juga berfungsi dalam osmosensing, pencahayaan dan sensorik.
CRE1 adalah intraseluler histidin kinase (HK) dengan tiga domain. Domain sensor berada di terminal N akhir dari protein termasuk dua membrane hidrofobik dan mencakup jangkar reseptor yang berada di membrane plasma, dimana diantara ruang tersebut terdapat loop hidrofilik yang membentang ke ruang ekstraseluler. Ruang ekstraseluler ini termasuk tempat dimana sitokinin melekat sejak mutasi pada sisi yang menghalangi sitokinin berikatan dan menyebabkan reseptor tidak berfungsi. Dan domain histidin kinase terletak di sisi sitoplasma membrane. Pertama kali ketika sitokinin melekat, kinase mengidentifikasi adanya HK sebagai enzim yang terlibat dalam reaksi fosforilasi dan untuk histidin kinase kelompok fosforilnya akan ditambahkan ke residu histidin. Fosfat yang menempel pada residu histidin tertentu (HIS 459) termasuk dalam domain histidin kinase. Histidin kinase ini juga mencakup dua domain penerima yaitu Dad an Db. Setelah menjadi jelas bahwa akseptornya adalah kinase histidin Arabidopsis genom sepnuhnya bisa diurutkan dengan mencari komponen potensial lain dari sistem signaling. Arabidopsis ini dapat diketahui bahwa dia mempunyai gene dari 8 histidine kinase yang berbeda-beda, 6 6 protein fosfotransfer protein dan 23 regulasi respon. Hanya 3 dari gen HK yang termasuk dalam sitokinin reseptor yaitu CRE1, AHK2, AHK3. Berikut ini struktur dari sitokinin
reseptor CRE1. Monomer (kiri) memiliki tiga domain dan dua kecil daerah hidrofobik membran-spanning. Mengikat dengan sitokinin (kanan) menginduksi dimerisasi dan autofosforilasi. Lokasi dari histidin dan residu fosfat-mengikat asam aspartat ditunjukkan.
2. Rantai Sinyal Sitokinin Melibatkan Transfer Grup Phosphoryl Ke Respon Regulasi
Gambar Transduksi Sinyal Sitokinin Sumber : Hopkins, 2009
Pertama adanya pengikatan molekul sitokinin ke domain pengirim termasuk dimerisasi dan selanjutnya fosforilasi pada residu histide dalam masing-masing 2 reseptor molekul. Sedangkan sebagian besar enzim kinase mengkatalisis penambahan kelompok fosforil pada molekul kedua tersebut, sedangkan reseptor autophosphorylates histidin kinase akan memfosforilasi sendiri. Kelompok fosforil kemudian secara spontan ditransfer ke residu asam aspartat pada domain penerima Db.
Kelas prokariotik terdapat dua komponen sistem yang terdiri dari kinase reseptor dan regulator respon. Dimana regulasi respon diaktifkan dengan menerima kelompok fosforil langsung dari kinase histidine. Pada tumbuhan, kelompok fosforil dilewatkan melalui satu atau lebih protein histidin-phosphotransfer (HPTs). Kelompok fosforil ditransfer dari domain penerima kinase histidin untuk residu histidin pada protein HP. Protein HP terfosforilasi kemudian bermigrasi ke inti di mana kelompok fosforil ditransfer ke residu Asp dalam regulator respon. Hal tersebut menunjukkan bahwa transfer bergantian dari histidin menjadi asam aspartat untuk histidin menjadi asam aspartat. Sistem kelompok fosforil yang ditransfer yaitu HKS, HPTs, dan RRS dan disebut juga sebagai jaringan phosphorelay.
Ada dua kelas dari respon regulator tipe A dan tipe B. Peran regulator respon masih bekerja, tetapi secara umum, tampak bahwa tipe B regulator respon adalah faktor transkripsi. Ketika diaktifkan oleh fosforilasi respon regulasi tipe B menginduksi ekspresi gen yang bertanggung jawab untuk mengatur beberapa tanggapan sitokinin. Di antara gen target untuk tipe B regulator respon adalah gen untuk jenis regulator respon A. Respon regulator jenis A, bagaimanapun tidak mempengaruhi factor transkripsi dan tidak mengatur ekspresi gen. Respon regulasi tersebut memodulasi respon sitokinin dengan mempengaruhi aspek-aspek lain dari metabolisme.
Sistem sitokinin mempunyai kemampuan untuk mematikan jaringan phosphorelay ketika tidak ada sitokinin hadir. Kesimpulan ini didasarkan pada temuan bahwa CRE1, selain dari fungsi kinase yang, juga menunjukkan aktivitas fosfatase. Aktivitas enzim fosfatase adalah kebalikan dari kinase fosfatase menghilangkan kelompok fosforil. Jadi, dengan tidak adanya sitokinin, CRE1 membalikkan proses, membongkar kelompok fosforil dari HPTs, dan cepat menginaktivasi jalur respon sitokinin.
a. Pengertian dan Sintesis Asam Absisat
Asam absisat (Abscisic Acid, ABA) merupakan seskuiterpenoid berkarbon 15 yang disintesis sebagian di kloroplas dan plastida lain melalui lintasan asam mevalonat. Biosintesis ABA pada sebagian besar tumbuhan terjadi secara tidak langsung melalui penguraian karotenoid tertentu yang ada di plastida. Kloroplas daun mengandung karotenoid yang menjadi bahan dasar ABA, sementara di akar, buah, embrio biji serta bagian tumbuhan tertentu lainnya, karotenoid penting berada di kromoplas lain, leukoplas atau proplastid. Semua reaksi yang membentuk xantoksin mungkin berlangsung di plastid, namun tahap berikutnya mungkin terjadi di suatu tempat di sitosol (Salisbury dan Ross, 1995).
Gambar. Struktur kimia dari ABA (Taiz and Zeiger, 2002).
Karotenoid violaxantin dengan konfigurasi trans pada semua ikatan rangkap, oleh suatu enzim diubah menjadi 9-cis violaxantin yang mempunyai konfigurasi cis yang sama dengan ABA pada karbon 2 dan 3. Selanjutnya, 9-cis violaxantin akan teroksidasi oleh O2 dan pecah, melepaskan senyawa atau beberapa senyawa yang belum dikenal (dengan total 25 karbon) dan
xantoksin yaitu epoksida berkarbon 15 dengan struktur serupa dengan ABA. Xantoksin diubah menjadi ABA aldehid dengan membuka cincin epoksida dan dengan oksidasi (oleh NADP+ atau NAD+ ) gugus hidroksil cincin menjadi gugus keto. Akhirnya gugus aldehid di rantai samping ABA aldehid dioksidasi menjadi gugus karboksil ABA. Yang menarik, oksidasi terakhir ini hampir dipastikan membutuhkan koenzim yang mengandung molibdenum yang menunjukkan adanya fungsi penting lain dari molibdenum bagi tumbuhan (Salisbury dan Ross, 1995).
Gambar 2 reaksi sintesis ABA dari karotenoid violaxantin (Salisbury dan Ross, 1995). ABA dapat dinonaktifkan dengan dua cara. Pertama dengan penempelan glukosa pada gugus karboksilnya membentuk ester ABA-glukosa. Ester ini tampaknya hanya terdapat di vakuola. Proses penonaktifan ini juga terjadi pada auksin, giberilin dan sitokinin. Proses
penonaktifan lainnya ialah oksidasi dengan O2 membentuk asam faseat dan asam dihidrofaseat. ABA diangkut dengan mudah dalam xilem dan floem dan juga dalam sel parenkim, biasanya tak ada polaritas, sehingga pergerakan ABA dalam tumbuhan serupa dengan pergerakan giberilin (Salisbury dan Ross, 1995).
b. Fungsi Asam Absisat
Beberapa fungsi dari asam absisat (ABA) dapat dijelaskan sebagai berikut: a ABA menginduksi penutupan stomata
Kandungan ABA dalam daun monokotil dan dikotil meningkat beberapa kali lipat jika daun mengalami keadaan rawan air, baik jika daun dipisahkan dari akarnya taupun tidak. Diketahui pula bahwa akar yang mengalami rawan air juga membentuk ABA lebih banyak dan bahwa ABA ini diangkut melalui xilem menuju daun dan menutup stomata. ABA y ang dipasok oleh akar sebagian besar berasal dari ujung akar dangkal yang mengalami rawan air dan ABA berlaku sebagai isyarat bagi daun jika air tanah mulai habis. Stomata menutup sebagai respon terhadap ABA yang berasal dari akar atau daun, sehingga terlindung dari kekeringan. ABA menyebabkan stomata menutup dengan menghambat pompa proton yang kerjanya bergantung pada ATP di membran plasma sel penjaga. Pompa ini biasanya mengangkut proton keluar dari sel penjaga, dan menyebabkan terjadinya aliran masuk cepat dan penimbunan K+, kemudian terjadi penyerapan air secara osmotik serta pembukaan stomata. Namun, ABA yang bekerja di ruang bebas pada permukaan luar membran plasma sel penjaga membatasi masuknya K+ , sehingga K+ dan air merembes keluar, turgor berkurang dan stomata menutup (Salisbury dan Ross, 1995). b ABA melindungi terhadap keadaan rawan garam dan rawan dingin
Tingkat ABA naik bukan hanya ketika tumbuhan mengalami tekanan akibat pasokan air yang tidak mencukupi, tapi juga akibat tanah bergaram, suhu dingin, suhu beku dan suhu tinggi. Rawan garam menyebabkan terbentuknya beberapa protein baru, khususnya protein berbobot molekul rendah yaitu osmotin (Salisbury dan Ross, 1995).
c ABA menghambat pertumbuhan embrio pada permulaan dormansi biji
Tahapan dalam kehidupan tumbuhan yang menguntungkan jika pertumbuhan dihentikan adalah pada saat permulaan dormansi biji dan ABA bertindak sebagai penghambat pertumbuhan. Akan berkecambah jika ABA dihambat dengan cara membuatnya tidak aktif, membuangnya atau dengan peningkatan aktivitas giberilin (Campbell, 2003). ABA eksogen merupakan penghambat
kuat bagi perkecambahan biji. Ketika dormansi berakhir, oleh suatu keadaan lingkungan misalnya cahaya dan suhu rendah, ABA akan menurun dan biji dapat berkecambah.
d ABA mempengaruhi pengguguran daun
Semula ABA disebut sebagai penyebab penguguran daun. Namun, sejak terbukti bahwa ABA menstimulasi penguguran organ hanya pada beberapa spesies tumbuhan saja dan bahwa hormon utama yang menyebabkan penguguran adalah etilen. Di sisi lain, ABA jelas terlibat dalam pelayuan daun, dan melalui hal ini secara tidak langsung meningkatkan pembentukan dan stimulasi etilen (Taiz and Zeiger, 2002).
c. Hubungan Asam Absisat dengan Pertumbuhan dan Perkembangan Tumbuhan ABA berperan utama dalam mengatur permulaan dan pemeliharaan dormansi pucuk dan biji serta respon tumbuhan terhadap stres. Selain itu ABA juga mempengaruhi berbagai aspek lain pada perkembangan tumbuhan, dengan berinteraksi (biasanya secara berlawanan) dengan auksin, sitokinin, giberilin dan etilen (Taiz and Zeiger, 2002). Salah satu aspek tersebut adalah pada perkembangan biji (embriogenesis).
Perkembangan embrio dapat dibagi dalam tiga fase utama yaitu:
1 Fase pertama, yang ditandai dengan pembelahan sel dan jaringan, zigot mengalami embriogenesis dan proliferasi jaringan endosperm.
2 Fase kedua, pembelahan sel berhenti dan terjadi penimbunan senyawa (cadangan makanan).
3 Fase ketiga, embrio menjadi toleran terhadap pengeringan dan biji menjadi kering (kehilangan 90% air). Sebagai konsekuensi dari pengeringan, metabolisme berhenti dan biji memasuki masa istirahat. Berbeda dengan biji yang mengalami dormansi, biji yang istirahat akan berkecambah jika terkena air.
Dua fase terakhir menghasilkan biji yang aktif dengan sumber yang memadai untuk mendukung perkecambahan dan tahan hingga berminggu-minggu bahkan bertahun-tahun sebelum memulai lagi perkecambahan. Secara khas, kandungan ABA pada biji sangat rendah di awal embriogenesis, dan kemudian berangsur-angsur menurun hingga biji menjadi matang (Taiz and Zeiger, 2002).
Menurut Quatrano (1987), ABA endogen sangat berkaitan dengan pembentukan awal lintasan pematangan normal dan dengan penghambatan perkecambahan dini. Selanjutnya, pada banyak spesies ABA eksogen dapat menyebabkan atau mempercepat pembentukan beberapa protein sebagai cadangan makanan yang khusus dalam embrio yang dibiakkan. Bukti tersebut
menunjukkan bahwa peningkatan taraf ABA secara normal pada awal dan pertengahan fase perkembangan biji dapat mengatur penimbunan protein cadangan (Salisbury dan Ross, 1995). E. ETILEN
Ethylene merupakan hidrokarbon sederhana dengan struktur kimia H2C=CH2. ethylene tampaknya tidak terlalu dibutuhkan untuk pertumbuhan vegetatif, meskipun dapat memiliki dampak yang signifikan pada pengembangan akar dan tunas. Ethylene umumnya digunakan untuk meningkatkan pematangan pisang dan buah-buahan lain. Ethylene sering dihasilkan ketika konsentrasi auksin tinggi. Ethylene ditemukan pada semua tanaman organ akar, batang, daun, umbi, umbi-umbian, buah-buahan, biji-bijian, dan sebagainya meskipun tingkat produksi dapat bervariasi tergantung dari perkembangannya (Hopkins dan Huner, 2009)
a. Etilen Disintesis Dari Asam Amino Metionin
Terdapat tiga tahap jalur untuk biosintesis etilen pada tumbuhan tingkat tinggi. langkah pertama, gugus adenosine (adenin dan ribosa) yang disumbangkan untuk metionin oleh molekul ATP, sehingga membentuk SAM. Konversi metionin untuk SAM dikatalisis oleh enzim metionin adenosyltransferase atau SAM sintetase. Pemecahan SAM untuk menghasilkan 5'-methylthioadenosine (MTA) dan ACC yang dimediasi oleh enzim ACC sintase. Aspek penting yang lain dari biosintesis etilen adalah metionin yang jumlahnya terbatas. Untuk mempertahankan tingkat produksi etilen normal, sulfur yang dilepaskan selama pembentukan etilena harus didaur ulang kembali membentuk metionin sehingga sering disebut sebagai siklus metionin atau juga dikenal sebagai siklus Yang (Hopkins dan Huner, 2009)
Gambar: skema biosintesis etilen (Hopkins dan Huner, 2009) b. Etilen Berperan Dalam Perkembangan Vegetatif
Ethylene dikenal terutama untuk pematangan buah dan penuaan. Ethylene juga telah terbukti untuk merangsang pemanjangan batang, tangkai daun, akar, dan struktur bunga dari tanaman yang hidup di air dan tanaman semiakuatik. Efeknya terutama dicatat dengan tanaman air karena genangan air mengurangi dispersi gas dan dengan demikian mempertahankan etilena internal dengan tingkat yang lebih tinggi. Ethylene banyak merangsang penghambatan dan pertumbuhan abnormal seperti pembengkakan jaringan batang dan lekukan yang terdapat di bawah daun atau epinasty. Daun epinasty terjadi karena pemanjangan sel yang berlebihan pada bagian adaxial daun. Peran ethylene juga untuk perkecambahan biji, penghambatan tunas, mengurangi dominasi apikal, pematangan buah, kematian sel, dan tanggapan patogen (Hopkins dan Huner, 2009)
c. Reseptor Dan Pensinyalan Ethylene
Dengan tidak adanya etilena, sinyal rantai dimulai dengan protein yang disebut Constitutive Triple Response 1 (CTR1). CTR1 berinteraksi dengan domain histidin kinase dari reseptor ETR1. Interaksi ini menyebabkan fosforilasi CTR1 dan memulai aliran sinyal transduksi. CTR1 adalah serine / treonin protein kinase. CTR1 mempengaruhi protein kinase yang akhirnya mengakibatkan satu atau lebih fosforilasi. Protein kinase mirip ke dengan gugus protein mitogen-activated kinase yang berperan penting dalam transduksi sinyal pada hewan, tumbuhan, dan jamur. Ketika etilena terikat dengan reseptor, mencegah interaksi CTR1 dengan ETR1 yang kemudian menginisiasi protein kinase dan selanjutnya melakukan aktivasi gen. Hasilnya adalah bahwa dalam ketiadaan etilena, ekspresi gen etilena selalu dalam keadaan aktif. Pengaruh etilena adalah untuk mengubah gen yang tidak aktif dengan mencegah aktivasi yang diperlukan oleh faktor transkrips (Hopkins dan Huner, 2009)
Gambar: Regulasi gen oleh etilen dan jalur responnya (Hopkins dan Huner, 2009)
F. BRASINOSTEROID
Brassinosteroids adalah hormon steroid dengan struktur kimia mirip dengan hormon steroid pada hewan. Brassinosteroids menimbulkan beragam jenis respon perkembangan, termasuk peningkatan laju batang dan pemanjangan tabung polen, peningkatan pembelahan sel, perkecambahan biji, morfogenesis daun, dominasi apikal, penghambatan pemanjangan akar, diferensiasi vaskular, mempercepat penuaan, dan kematian sel. Brassinosteroids juga terlibat dalam mediasi respon baik tekankan abiotik maupun biotik, termasuk garam, kekeringan, suhu ekstrem, dan patogen (Hopkins dan Huner, 2009)
a. Brassinosteroids Merupakan Polyhydroxylate Sterol Hasil dari Triterpen Squalene Brassinosteroids adalah zat lipoidal polyhydroxylated sterol yang berhubungan dengan biosintesis giberelin dan asam absisat. Tanaman mensintesis sejumlah besar dan berbagai sterol, termasuk sitosterol, stigmasterol, kolesterol, dan campesterol. Sterol adalah triterpenoid dengan 30 atom C yang berasal dari asetat melalui jalur asam mevalonat. Dalam sintesis terpen, penambahan secara berurutan dari 5-karbon isopentenil pirofosfat (IPP) menghasilkan senyawa terpen dengan 10-, 15-, atau 20- atom karbon. Triterpen terbentuk ketika dua unit C15 (farnesyl) bergabung untuk membentuk molekul squalene C30. Langkah pertama adalah reaksi siklisasi untuk membentuk cycloartenol sebagai prekursor umum. Reaksi dekarboksilasi dan oksidasi juga terlibat, karena umumnya sterol kebanyakan memiliki 26-29 karbon dan satu gugus
hidroksil (OH). Diperkirakan bahwa sebagian besar sterol (kecuali stigmasterol) dapat berfungsi sebagai prekursor untuk berbagai brassinosteroids (Hopkins dan Huner, 2009)
Gambar: langkah-langkah utama dalam biosintesis brassinolide dari triterpenoid squalene (Hopkins dan Huner, 2009)
b. Reseptor dan Pensinyalan Brasinosteroid
Reseptor utama untuk brassinosteroids membutuhkan interaksi dua protein yang berupa plasma membran yang terkait dengan heterodimer. Yang pertama adalah serin / treonin kinase dikenal sebagai Brassinosteroid Insensitive 1 (BRI1). Yang kedua adalah protein BRI1-Associated Reseptor kinase (BAK1). Brassinosteroids mengikat ke domain ekstraselular BRI1, yang pertama menginduksi disosiasi protein penghambat (BKI1) yang menghambat asosiasi BAK1 dengan BRI1 dan kemudian meningkatkan dimerisasi dengan BAK1 dan autofosforilasi dari BRI1. Fosforilasi kompleks memulai BR signaling. Salah satu target persinyalan brassinosteroid adalah protein BZR1 (Brassinazole Resistant 1). BZR1 merupakan faktor transkripsi yang tergantung pada fosforilasinya. Dalam keadaan terfosforilasi, BZR1 terperangkap dalam sitoplasma sementara defosforilasi memungkinkan untuk bergerak ke dalam inti. Fosforilasi BZR1 dimediasi oleh dua faktor yaitu; rantai BR signaling dan protein yang berbeda, yaitu BIN2. BIN2 memediasi fosforilasi BZR1 dan dengan demikian tetap menggunakan protein dalam sitoplasma. Di sisi lain sinyal brassinosteroid , menengahi
defosforilasi BZR1, yang keduanya mengaktifkan faktor transkripsi dan mendorong perpindahan ke dalam inti. Setelah di inti, BZR1-P mengikat target di daerah promotor BR-sensitif gen dan menginisiasi transkripsi. Persinyalan BR mungkin juga menghambat kemampuan fosforilasi dari BIN2, sehingga memastikan aktivasi dan lokalisasi inti BZR1 (Hopkins dan Huner, 2009)
Gambar: Skema persinyalan brassinosteroid (BR) (Hopkins dan Huner, 2009).
BAB III KESIMPULAN
A. Kesimpulan
Berdasarkan makalah yang telah dibuat, hormon pada tumbuhan terdiri dari beberapa hormon dan fungsi yang berbeda-beda. Hormon dapat mempengaruhi pertumbuhan dan perkembangan pada tanaman.
B. Saran
Beberapa saran yang dapat penulis berikan, antara lain agar makalah ini dapat menjadi sumber referensi dan ini dapat bermanfaat bagi yang membaca. Jika terdapat kesalahan dalam penulisannya diharapkan dapat memperbaikinya untuk lebih baik kedepannya.
DAFTAR PUSTAKA
Abidin. 1994. Dasar - Dasar Pengetahuan tentang Zat Pengatur Tumbuh. Penerbit Angkasa, Bandung.
Campbell, Neil. A. 2003. Biologi. Edisi Kelima Jilid 2. Jakarta: Erlangga.
Davies, J.P., 1995. Plant Hormones, Physiology Biochemistry and Molecular Biology. Dortrech: Kluwar Academic Publisher
Gardner, F. P., R. B., Pearce, and R. I. Mitchell. 1991. Fisiologi Tanaman Budidaya. Penerjemah: Susilo, H. UI Press. Jakarta
Hopkins, W.G., and Huner, N.P.A. 2009. Introduction to Plant Physiology Fourth Edition. United States of America: John Wiley & Sons, Inc
Hopkins. W.G. 1999. Introduction to Plant Physiology. John Willey and Sons, Inc. New York. Kende H., E. van der Knap, Hyung-Taeng Cho. 1998. “A Model Plant to Study Stem
Elongation”. Plant Physiol. 118: 1105-1112.
Kende, H. and J.A.D. Zeevart. 1997. “ The Five “Classical” Plant Hormones”. The Plant Cel. 9: 1197-1210.
Loveless, A.R. 1999.Prinsip-prinsip Biologi Tumbuhan untuk Daerah Tropik I (diterjemahkan Oleh Kartawinata, K., Danimiharja, S., dan Soetisna, U.). P.T Gramedia Pustaka Utama, Jakarta. 104
Noggle, G. R. And G. J. Fritz. 1983. Introductory Plant Physiology, New Jersey; Prentice. Hall. Inc.
Salisbury, F.B dan C.W. Ross. 1995. Fisiologi Tumbuhan Jilid 3. Penerjemah Lukman, O.R. dan Sumaryono. Penerbit ITB. Bandung.
Surachmat Kusumo, 1989 “Zat Pengatur Tumbuh Tanaman“ CV Yasa Guna.
Taiz, L dan Zeiger, E.2002. Plant Physiology Edisi 3. Sunderland, Massachusetts: Sinauer Associates, Inc., Publishers
Waier, T.E., Stocking, C.R., Barbour, M.G., and Rost, TL. 1982. Botany: An Introduction to Plant Biology. 6nd edition. University of California. California.