II TINJAUAN PUSTAKA
2.1 Antibodi
Respon imun akan terjadi dalam tubuh hewan yang terpapar oleh suatu antigen. Respon humoral tubuh yang terinfeksi akan menghasilkan antibodi. Antibodi merupakan suatu molekul protein atau globulin yang diproduksi oleh sel B dan saling berikatan secara spesifik dengan antigen. Antibodi ditemukan pada darah atau kelenjar tubuh vertebrata lainnya, dan digunakan oleh sistem kekebalan tubuh untuk mengidentifikasikan dan menetralisasikan benda asing seperti bakteri dan virus. Molekul antibodi adalah globulin, maka umumnya dikenal sebagai imunoglobulin (Ig). Ada 5 kelas utama imunoglobulin dalam serum yaitu IgG, IgA, IgM, IgD dan IgE (Alberts et al 2002; Roitt 2003). Bila serum protein dipisahkan dengan cara elektrophoresis, maka imunoglobulin ditemukan terbanyak dalam fraksi globulin gama, meskipun ada beberapa imunoglobulin yang juga ditemukan dalam fraksi globulin alfa dan beta (Bratawidjaja 2002).
Semua molekul imunoglobulin mempunyai 4 rantai polipeptida yang terdiri dari dua pasang rantai polipeptida berat (heavy = H) dan dua rantai ringan (light = L) yang identik. Keempatnya bergabung melalui ikatan disulfida antar rantai. Struktur molekul antibodi berbentuk seperti huruf “Y”, masing-masing mempunyai daerah variabel (VH dan VL) dan daerah konstan (CH dan CL). Fragmen imunoglobulin yang identik disebut Fab (Fragment antigen binding) yang merupakan bagian imunoglobulin yang mengikat antigen serta bereaksi dengan determinan antigen dan hapten. Bagian tunggal imunoglobulin disebut Fc (Fragment crystallizable) oleh karena mudah dikristalkan. Daerah variabel (V) tersusun dari sekitar 110 sampai 130 asam amino, merupakan gugus NH2 sebagai tempat ikatan antara rantai H dan L. Daerah konstan C pada rantai H meliputi daerah aktivasi komplemen dan molekul reseptor Fc dari berbagai jenis sel (Bratawidjaja 2002; Roitt 2003).
Daerah V dan C mempunyai tiga kelas determinan antigenik yaitu : (1) determinan isotypic, membedakan rantai ringan menjadi dua klas, yaitu kappa (κ) dan
lamda (λ) yang terdiri atas 230 asam amino, sedangkan rantai berat yang terdiri dari 450-600 asam amino mempunyai lima isotipe berbeda yang membagi imunoglobulin menjadi lima klas yang berbeda dengan fungsi yang berbeda-beda pula (IgG, IgM, IgA, IgE, IgD); (2) determinan allotypic, dibedakan dari produk gen pada lokus yang sama (IgG1, IgG2a, IgG2b); dan (3) determinan idiotypic, dihubungkan dengan spesifisitas ikatan antigen (Saldanha 2000) (Gambar 1).
Gambar 1 Tiga Kelas Determinan Antigenik (Sumber: Saldanha (2000))
Rantai-rantai pada molekul antibodi tersebut dapat dipisahkan satu sama lain dengan mereduksi ikatan S-S atau dengan pengasaman. Pada IgG yang merupakan tipe antibodi terbanyak, daerah engsel yang terpapar memanjang oleh adanya kandungan prolein yang tinggi, karena itu mudah terkena efek proteolitik; molekul akan terpotong oleh papain menjadi dua fragmen Fab yang identik, masing-masing mengandung satu tempat pengikatan antigen dan fragmen ketiga yaitu Fc yang tidak mempunyai kemampuan mengikat antigen. Pepsin memotong pada tempat yang berbeda, melepaskan Fc dari bagian molekul lainnya berupa fragmen 5S yang besar, yang dikenal sebagai F(ab)2. Fragmen ini mampu mengikat antigen seperti antibodi asal dan masih bersifat divalen. Daerah spesifik untuk berikatan dengan antigen disebut paratop dan mengekspresikan beberapa kumpulan idiotop yang mampu meningkatkan produksi antibodi (Roitt 2003). Allotypic, variasi asam amino pada lokus yang Idiotypic, variasi dihubungkan Isotypic, variasi rantai berat γ, μ, ε,
Menurut Roitt (2003) dapat dibuat antiserum yang spesifik terhadap molekul antibodi individual dan dapat membedakan antara satu antibodi monoklonal dengan lainnya tanpa tergantung dari struktur isotipik maupun alotipik. Antiserum tersebut dapat menetapkan determinan individual yang khas untuk tiap antibodi dan disebut idiotipe. Determinan idiotipe ini terletak di daerah variabel dari antibodi dan berkaitan dengan daerah hipervariabel.
2.2 Antibodi Anti-idiotipe
Idiotope adalah determinan antigen tunggal pada regio variabel molekul antibodi yang dapat ditemukan pada sel B dan sel T (Kennedy et al. 1983). Lokasi idiotope pada sisi pengikat antigen, pada rantai berat maupun pada rantai ringan antibodi, namun kebanyakan terletak pada rantai berat (Ban et al. 1994).
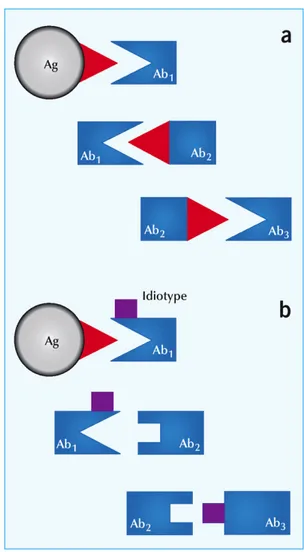
Idiotipe adalah kumpulan idiotope atau epitop spesifik, diekspresikan sebagai reseptor antigen pada sel B dan sel T. Jerne (1985) membuat suatu teori tentang jaringan idiotipik, bahwa imunisasi dengan suatu antigen dapat menginduksi antibodi spesifik terhadap antigen (Ab1). Antibodi yang dihasilkan dari antigen eksternal disebut antibodi idiotipe (Ab1), antibodi yang dihasilkan oleh Ab1 adalah antibodi anti-idiotipe (Ab2), antibodi yang dihasilkan Ab2 adalah antibodi anti anti-idiotipe (Ab3), dan seterusnya, sehingga pada akhirnya dapat menginduksi terbentuknya serangkaian autoantibodi yang dapat saling mengenali satu sama lain membentuk suatu jaringan idiotipik.
Daerah variabel (V) molekul antibodi mempunyai daerah spesifik untuk berikatan dengan antigen (paratop) dan permukaan yang mampu meningkatan produksi antibodi (idiotop). Paratop dan idiotop terletak pada tempat yang sama, maka anti-idiotipe akan meniru (internal image) struktur antigen melalui proses saling melengkapi (complementary) dan homobodies (Kennedy & Attanasio 1990). Jerne (1985) membuat suatu teori tentang jaringan idiotipik, bahwa imunisasi dengan suatu antigen dapat menginduksi antibodi spesifik terhadap antigen (Ab1).
Gambar 2 The idiotypic network: a. Menurut teori jaringan idiotipik dari Jerne, imunisasi dengan antigen dapat mengakibatkan tidak hanya generasi antigen-antibodi spesifik (Ab1) tetapi juga ke antibodi yang mengenali Ab1. Hal ini terjadi karena struktur unik (idiotipe) dari tempat pengikatan antigen dari Ab1 merangsang sistem kekebalan tubuh untuk menghasilkan Ab2 yang meniru struktur antigen. Mekanisme yang sama menghasilkan Ab3. Ab1 dan Ab3 memiliki kapasitas mengikat serupa dan dalam banyak kasus, urutan asam aminonya identik mengikat tempat antigen; b. Seperti yang juga dikemukakan oleh Jerne, struktur unik dalam rangka Ab1 dapat memicu jaringan idiotipik.
(Sumber: Shoenfeld (2004))
Antibodi anti-idiotipe (Ab2) yang memiliki karakteristik serologik internal image sangat potensial digunakan sebagai antigen dalam serodiagnostik, preparasi vaksin atau modulasi respon imun untuk mengontrol infeksi (Zhou et al. 1994). Antibodi anti-idiotipe juga dapat digunakan sebagai prekusor awal sistem imun inang terhadap agen infeksius. Pemberian anti-idiotipe pada simpanse sebelum pemberian antigen HBs meningkatkan titer antibodi terhadap HBs dibandingkan dengan tanpa pemberian anti-idiotipe (Kennedy et al. 1984). Menurut Suartha (2001), antibodi anti-anti-idiotipe mampu memberikan perlindungan 88.8% terhadap serangan bakteri Streptococcus Group C (SGC) ganas.
Antibodi anti-idiotipe juga dapat digunakan sebagai antigen pengganti pada imunisasi dengan antigen yang sulit diperoleh dalam jumlah yang banyak (Roitt 2003). Antibodi anti-idiotipe dapat diproduksi dengan mudah dalam jumlah yang banyak, kesulitan yang berhubungan dengan tenaga, biaya dalam penyediaan antigen dari agen penyakit dapat dieliminasi, bahaya penyebaran agen infeksius dalam pelaksanaan di lapangan dapat dihindari (Lin & Zhou 1995).
Antibodi anti-idiotipe dapat digunakan sebagai vaksin karena dapat meningkatkan respon kebal dari vaksin konvensional yang semula tidak efektif atau lemah (Huang et al. 1988). Paryati et al (2006), mengatakan antibodi anti-idiotipe mampu menginduksi kadar antibodi protektif terhadap rabies. Antibodi anti-idiotipe juga dapat mengurangi efek klinis yang merugikan dari penggunaan vaksin konvensional (Clark et al. 1996), mampu memberikan kekebalan protektif pada sistem imun prematur (Huang et al. 1988), protektif menghambat penyebaran secara vertikal infeksi suatu mikroorganisma (Kennedy et al. 1996). Vaksin antibodi anti-idiotipe juga dapat merangsang klon imun yang toleran dan tersembunyi (McNamara et al. 1984).
Chatterjee et al. (2000) menggunakan antibodi anti-idiotipe untuk melawan
kanker karena penggunaan vaksin anti-idiotipe lebih baik dari vaksin antigen tradisional, sedangkan Reinarzt et al. (2003) mengatakan antibodi anti-idiotipe dapat sebagai pengganti antigen anti tumor dalam strategi vaksinasi karena dapat menstimulasi sel B. Lebih lanjut antibodi anti-idiotpe dapat meningkatkan respon imun terhadap neoplasma dan meningkatkan produksi auto antibodi pada penyakit autoimun (Rico & Hall 1989).
Park et al. (2005) dalam penelitiannya mengembangkan penggunaan antibodi
anti-idiotipe terhadap kapsular polisakarida dari Neisseria meningitides group B sebagai vaksin karena belum ada vaksin terhadap penyakit ini yang merupakan penyakit serius penyebab sepsis dan meningitis pada bayi dan balita.
2.4 Virus Influenza
Penyakit flu pada manusia dan hewan disebabkan oleh virus dalam famili
Orthomyxoviridae, memiliki pembungkus (envelope), bersegmen dan memiliki negative-single strand Rybonucleic acid (RNA). Virus ini berukuran 80-120 nm, merupakan
partikel pleimorphic berukuran sedang yang terdiri atas 2 lapis lemak dan terletak diatas matriks M1 yang mengelilingi genom. Permukaan envelope mempunyai dua tonjolan glikoprotein yaitu hemaglutinin (H) dan neuraminidase (N). Protein lain selain H dan N, virus influenza A juga memiliki protein matriks (M1, M2), nukleoprotein (NP), polimerase (P1, Pb2, PA), non structural (NS1), dan NEP. Masing-masing protein mempunyai fungsi yang berbeda (Tabel 1) (Mulyadi & Prihatini 2005).
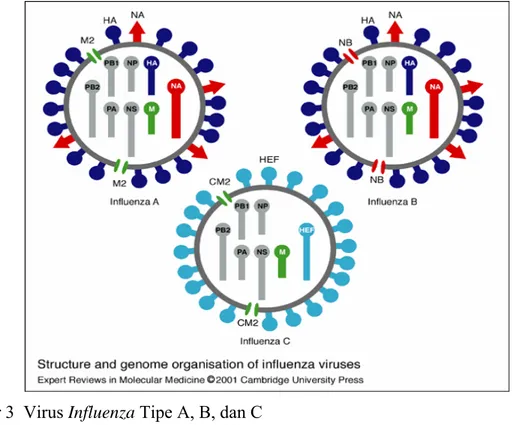
Sampai saat ini famili Orthomyxoviridae terdiri dari 5 genus yaitu Influenzavirus A, Influenzavirus B, Influenzavirus C (tiga genus ini biasa disebut dengan influenza tipe A, B, dan C), Thogotovirus dan Isavirus. Hanya Genus Influenzavirus A yang menginfeksi unggas (Capua & Alexander 2009). Virus influenza tipe A dan B memiliki 8 segmen RNA, namun virus influenza tipe C hanya memiliki 7 segmen (Gambar 3) (Hoffmann et al. 2000).
Determinan antigenik utama dari virus influenza A dan B adalah glikoprotein transmembran hemaglutinin (H atau HA) dan neuroaminidase (N atau NA), yang mampu memicu terjadinya respon imun dan respon yang spesifik terhadap subtipe virus. Respon ini sepenuhnya bersifat protektif di dalam, tetapi bersifat protektif parsial pada lintas subtipe yang berbeda. Berdasarkan sifat antigenisitas dari glikoprotein-glikoprotein tersebut, saat ini virus influenza dikelompokkan ke dalam enambelas subtipe H (H1-H16) dan sembilan N (N1-N9). Kelompok-kelompok tersebut ditetapkan ketika dilakukan analisis filogenetik terhadap nukleotida dan penetapan urutan (sequences) gen-gen HA dan NA melalui cara deduksi asam amino (Fouchier 2005).
Tabel 1 Protein Influenza A Protein Tempat (perkiraan jumlah
virion)
Fungsi Hamaglutinin (HA) Permukaan (500) Perlekatan sel dan
penetrasi, aktivitas penyatuan (fusi)
Neuraminidase (NA) Permukaan (100) Pelepasan virus, aktivitas enzim
Membran/matrik (M1)
Di dalam (interna) (3000) Struktur pembungkus (envelope) utama protein, pertemuan virus
M2 Permukaan (20-60) Virus tidak dibungkus dan
pertemuan, hubungan ion Nucleoprotein (NP) Di dalam (interna) (3000) Berkaitan dengan RNA dan
protein polymerase Polymerase
(PB1,PB2,PA)
Di dalam (interna) (30-60) Replikasi RNA dan transkripsi
NS1 Nonsruktural(sel terinfeksi)
Pengaturan replikasi virus
NEP Di dalam (interna)
(130-200) Faktor ekspor inti (nuclear)
Gambar 3 Virus Influenza Tipe A, B, dan C (Sumber: Hoffmann et al. (2000))
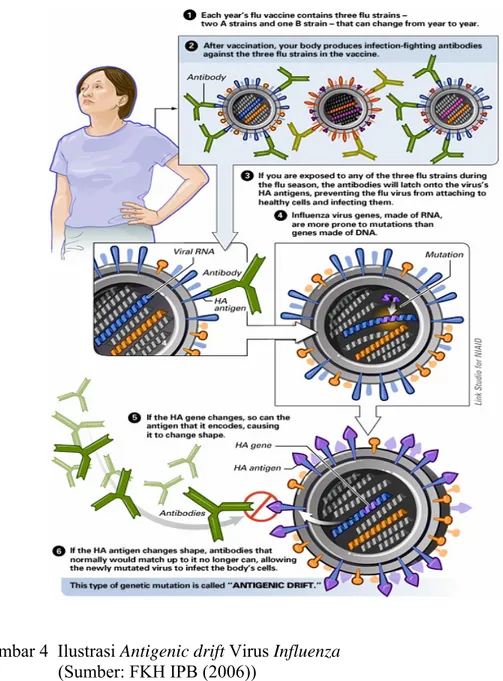
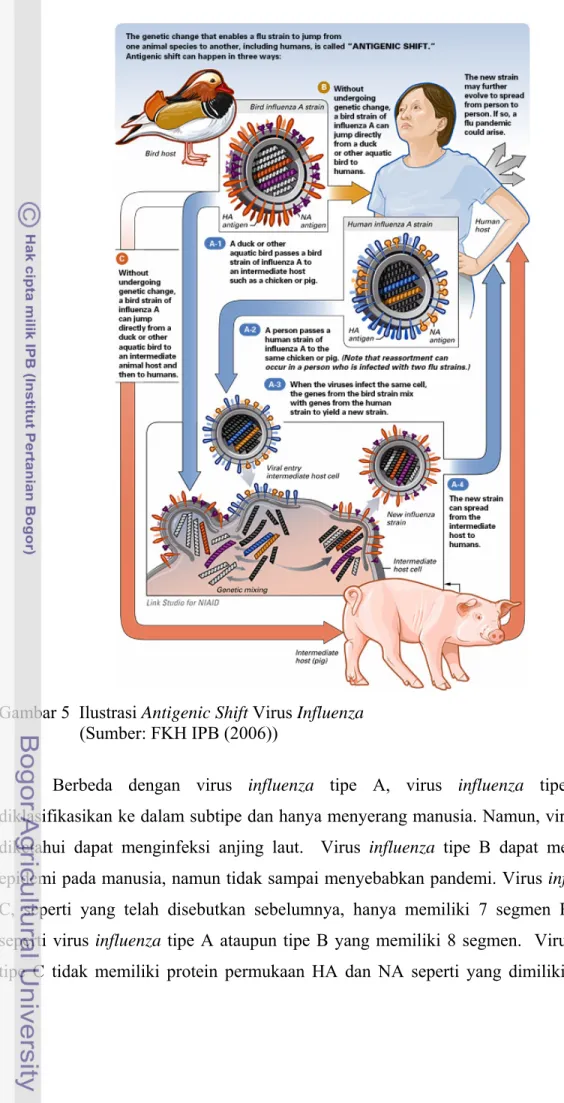
Virus influenza A mempunyai sifat mudah berubah. Antigen permukaan yang dimiliki virus influenza tersebut dapat berubah secara periodik yang lebih dikenal dengan istilah antigenic drift (mutasi titik/minor) dan antigenic shift (pergeseran genetik/mutasi mayor). Antigenic drift merupakan perubahan yang terjadi akibat mutasi genetik struktur protein permukaan virus, sehingga antibodi yang telah terbentuk oleh tubuh akibat vaksinasi sebelumnya tidak dapat mengenali keberadaan virus tersebut, sedangkan
antigenic shift merupakan perubahan genetik virus yang memungkinkan virus ini
menginfeksi secara lintas spesies (Gambar 4 dan 5) . Mutasi mayor merupakan keunikan virus influenza karena genom virus itu terdiri atas delapan potong RNA, sehingga ketika virus influenza dengan tipe berbeda menginfeksi sel yang sama akan terjadi pertukaran segemen RNA dalam sel. Kedua sifat tersebut dapat menyebabkan kejadian pandemi (Stohr 2005).
Gambar 4 Ilustrasi Antigenic drift Virus Influenza (Sumber: FKH IPB (2006))
Gambar 5 Ilustrasi Antigenic Shift Virus Influenza (Sumber: FKH IPB (2006))
Berbeda dengan virus influenza tipe A, virus influenza tipe B tidak diklasifikasikan ke dalam subtipe dan hanya menyerang manusia. Namun, virus ini telah diketahui dapat menginfeksi anjing laut. Virus influenza tipe B dapat menyebabkan epidemi pada manusia, namun tidak sampai menyebabkan pandemi. Virus influenza tipe C, seperti yang telah disebutkan sebelumnya, hanya memiliki 7 segmen RNA, tidak seperti virus influenza tipe A ataupun tipe B yang memiliki 8 segmen. Virus influenza tipe C tidak memiliki protein permukaan HA dan NA seperti yang dimiliki oleh virus
influenza tipe A dan B, akan tetapi kedua segmen tersebut digantikan oleh glikoprotein
tunggal yang disebut dengan haemagglutinin-esterase-fusion (HEF). Virus influenza tipe C hanya menyebabkan gejala penyakit ringan saja dan tidak menyebabkan epidemi maupun pandemi penyakit pada manusia (Nicholson et al. 1998).
2.5 Virus Avian Influenza (AI)
Avian Influenza (AI) menyebabkan angka kematian yang tinggi pada ayam di
Italia pada tahun 1878. Namun baru diketahui pada tahun 1955 bahwa penyebab fowl
plague sebenarnya adalah virus AI yang memiliki komposisi gen yang serupa (hampir
identik) dengan virus influenza manusia.
Virus AI adalah virus influenza tipe A, pada awalnya hanya ditularkan oleh unggas. Unggas tersebut adalah burung, bebek, ayam, selain itu dapat juga ditularkan oleh beberapa hewan lain seperti babi, kuda, anjing laut, ikan paus, musang, dan kucing. Data lain menunjukkan penyakit ini bisa terdapat diburung puyuh dan burung onta. Penyakit ini ditularkan dari burung ke burung, tetapi dapat juga menular ke manusia (Mulyadi & Prihatini 2005). Keberadaan virus AI H5N1 pada unggas air di Indonesia telah dibuktikan dengan ditemukannya isolat dari unggas air di daerah Jawa Barat (Susanti 2008) dan di kucing (Murtini et al. 2008).
Berdasarkan atas patogenitasnya, virus AI dibedakan menjadi 2 bentuk yaitu Low
Pathogenic Avian Influenza (LPAI) dan Highly Pathogenic Avian Influenza (HPAI).
Pada umumnya virus AI subtipe H1-H4, H6, H8-H15 ada dalam bentuk LPAI dan umumnya menyebabkan sedikit gejala klinis atau bahkan tidak memperlihatkan gejala klinis sedikitpun. Sifat mudah mutasi dari virus ini menyebabkan keganasannya ditentukan oleh waktu, tempat dan inang yang terinfeksi. Artinya walaupun sama-sama H5 yang menginfeksi belum tentu menunjukkan tingkat keganasan yang sama (Russel & Webster 2005). Menurut Capua dan Alexander (2009), hanya virus AI subtipe H5, H7 dan H10 yang terlihat sebagai penyebab HPAI tapi tidak semua sub tipe H5, H7 dan H10 bersifat virulen.
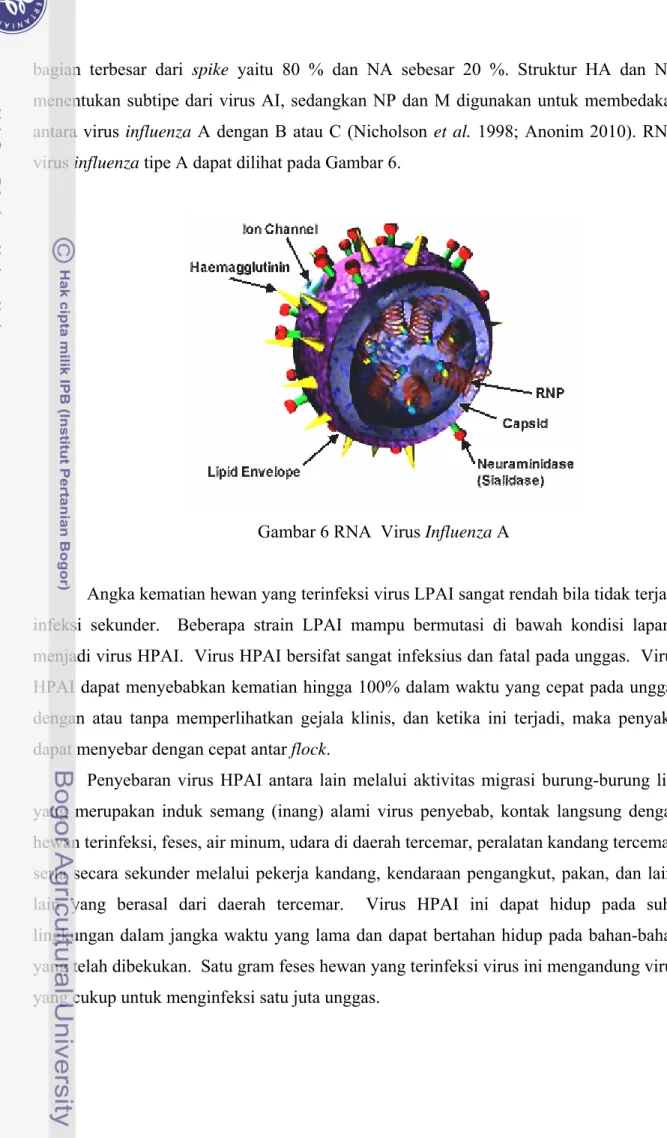
Virus AI dibungkus oleh glikoprotein dan dilapisi oleh lapisan lemak ganda (bilayer lipid). Glikoprotein HA dan NA merupakan protein permukaan yang sangat berperan dalam penempelan dan pelepasan virus dari inang. Protein HA merupakan
bagian terbesar dari spike yaitu 80 % dan NA sebesar 20 %. Struktur HA dan NA menentukan subtipe dari virus AI, sedangkan NP dan M digunakan untuk membedakan antara virus influenza A dengan B atau C (Nicholson et al. 1998; Anonim 2010). RNA virus influenza tipe A dapat dilihat pada Gambar 6.
Gambar 6 RNA Virus Influenza A
Angka kematian hewan yang terinfeksi virus LPAI sangat rendah bila tidak terjadi infeksi sekunder. Beberapa strain LPAI mampu bermutasi di bawah kondisi lapang menjadi virus HPAI. Virus HPAI bersifat sangat infeksius dan fatal pada unggas. Virus HPAI dapat menyebabkan kematian hingga 100% dalam waktu yang cepat pada unggas dengan atau tanpa memperlihatkan gejala klinis, dan ketika ini terjadi, maka penyakit dapat menyebar dengan cepat antar flock.
Penyebaran virus HPAI antara lain melalui aktivitas migrasi burung-burung liar yang merupakan induk semang (inang) alami virus penyebab, kontak langsung dengan hewan terinfeksi, feses, air minum, udara di daerah tercemar, peralatan kandang tercemar, serta secara sekunder melalui pekerja kandang, kendaraan pengangkut, pakan, dan lain-lain yang berasal dari daerah tercemar. Virus HPAI ini dapat hidup pada suhu lingkungan dalam jangka waktu yang lama dan dapat bertahan hidup pada bahan-bahan yang telah dibekukan. Satu gram feses hewan yang terinfeksi virus ini mengandung virus yang cukup untuk menginfeksi satu juta unggas.
Virus influenza yang sangat virulen (H5N1) dapat hidup di air sampai 4 hari pada suhu 22o C dan lebih dari 30 hari pada 0o C, di dalam kotoran dan tubuh unggas yang sakit virus dapat bertahan lebih lama, virus akan mati dengan pemanasan 60o C selama 30 menit atau 56o C selama 3 jam, deterjen, desinfektan (formalin, iodin) (Horimoto & Kawaoka 2001; Cucunawangsih 2006).
2.6 Patogenesis Avian Influenza 2.6.1 Reseptor Virus Avian Influenza
Penularan virus influenza A dapat terjadi antar spesies termasuk manusia, namun virus AI tidak efisien dalam replikasi pada sel manusia, demikian juga sebaliknya, virus
influenza pada manusia tidak efisien replikasi pada unggas. Hal ini disebabkan oleh
perbedaan reseptor yang dimiliki oleh masing-masing sel tersebut. Ikatan pada reseptor merupakan awal patogenesitas virus AI. Virus akan masuk ke sel inang melalui endositosis yang diperantarai oleh reseptor (Elton et al. 2001; Rahardjo 2004).
Reseptor yang dimiliki oleh sel manusia berupa N-acetylsialic acid yang mengikat galaktosa dengan ikatan alfa 2,6 atau disingkat NeuAcα2,6Gal (asam sialat-α2,6), sedangkan pada unggas dan kuda, senyawa galaktosa diikat pada alfa 2,3 atau disingkat NeuAcα2,3Gal (asam sialat-α2,3). Kebanyakan hewan mempunyai reseptor mirip dengan unggas seperti paus dan anjing laut. Khusus untuk babi, sampai saat ini satu-satunya hewan yang mempunyai kedua macam reseptor tersebut, dalam arti bahwa babi sensitif terhadap AI maupun virus influenza manusia (Nicholson et al. 1998; Suzuki et al. 2000). Perbedaan reseptor ini korelasinya dengan letak asam amino nomer 226 yaitu pada virus
influenza manusia diisi oleh leusin (Leu), sedangkan pada pada AI dan virus influenza
pada kuda (Equine Influenza) diisi oleh glisin (Gln) (Rahardjo 2004).
2.6.2 Mekanisme Infeksi Virus Avian Influenza
Proses terjadinya infeksi atau patogenesa tahap pertama terjadi secara inhalasi (menghirup) atau ingesti (memakan) AI. Enzim tripsin dan protease lainnya dalam sel tropisma terutama pada epitel saluran pernapasan, paru-paru dan trakhea tersedia untuk pembelahan (cleavage) protein hemaglutinin. Jadi pada saat AI menginfeksi sel, enzim tripsin dan protease dalam sel akan memecah protein hemaglutinin menjadi dua bagian yaitu HA1 dan HA2 (Rahardjo 2004).
Mekanisme infeksi virus influenza dimulai dengan perlekatan virus pada permukaan membran plasma. Reseptor untuk virus influenza adalah sialoglycolipid atau
gangliosides atau sialoglycoprotein. Terminal sialic acid akan dikenali oleh hemaglutinin
(HA) yang berperan dalam perlekatan virus. Kedudukan reseptor virus pada bagian distal globular hemaglutinin, yaitu daerah molekul yang menunjukkan sedikit perbedaan yang dikelilingi oleh 3 tempat antigen yang berbeda. Virus akan masuk ke dalam sel dengan bantuan ikatan paku HA pada mukoprotein yang mengandung terminal N-acetyl
neuraminic acid (NANA = sialic acid) (Manugerra & Hannoun 1999).
Partikel virus setelah melekat akan ditelan melalui proses endositosis dan virus lewat suatu lubang selubung akan masuk ke dalam gelembung endositotik dan akhirnya endosom. mengatakan secara berangsur-angsur lisosom akan menyatu pH akan menurun (pH kira-kira 5). Protein HA akan berikatan dengan reseptor yang terdapat dinding sel inang. Siklus replikasi terjadi dalam saluran pernapasan dan atau saluran pencernaan dengan mengeluarkan bagian dari virus yang bersifat menimbulkan infeksi (Rahardjo 2004). Protein virus eksternal seperti HA yang tahan terhadap degradasi enzim (protease). Protein HA akan mengalami perubahan pada saat kondisi pH yang cukup asam tersebut, yaitu dengan membuka bagian hydrophobic sub unit HA2 sehingga memungkinkan terjadinya penyatuan antara membran endosomal seluler dan lapisan lemak ganda (lipid bilayer) virus. Untuk virus influenza tipe A protein M2 memungkinkan destabilization lapisan protein M1. Virus akan masuk pada saat nukleokapsid dilepaskan ke dalam sitoplasma seluler. Penghambatan masuknya virus dapat bervariasi tergantung pada tingkat kekebalan dan bahan kimia. Pengaktifan pH oleh aliran ion akan membuat protein M2 menjadi penting dalam proses uncoating (pelepasan selubung). Spesifik target rangkaian asam nukleat (sequence nucleat) pada nukleoprotein (NP) akibat translokasi nukleokapsid ke dalam nukleus (Manugerra & Hannoun 1999)
Virus influenza yang merupakan virus RNA mempunyai 3 tipe virus spesifik RNA (messenger, antigenomic dan viral atau genomic) yang semuanya disintesa di dalam nukleus. Segmen NS melalui sambungan untuk menyandikan protein NS1 (non-struktur) dan NS2. Untuk virus tipe A, segmen M sebagai sandi protein M1 dan M2. Selama infeksi oleh virus influenza, total sintesa protein dipertahankan pada tingkat yang tinggi kemudian terjadi surprising swing dari sintesa protein seluler menjadi protein influenza,
seharusnya dalam bagian untuk menghalangi translasi seluler mRNA. Translasi ada dalam sitoplasma dimana protein virus disintesa. Ini yang menjalani glikosilasi seperti HA dan NA, lewat ke dalam retikulum endoplasmik dan kemudian Golgi dan trans-Golgi
apparatus. Pada jalur ini glikoprotein influenza dilekatkan ke dalam lipid bilayer sampai
pada permukaan sel. Protein virus yang lain akan datang di bawah membran sel plasma melalui difusi yang sederhana. Virion dibentuk oleh kuncup (budding) pada permukaan membran plasma.
Menurut Garman dan Laver (2004), hemaglutinin adalah sebuah protein yang mengalami glikosilasi dan asilasi (glycosylated and acylated protein) terdiri dari 562-566 asam amino yang terikat dalam sampul virus. Kepala membran distalnya yang berbentuk bulat, daerah eskternal yang berbentuk seperti tombol dan berkaitan dengan kemampuannya melekat pada reseptor sel, terdiri dari oligosakharida yang menyalurkan derivat asam neuroaminic. Daerah eksternal (exodomain) dari glikoprotein transmembran yang kedua, neuroamidase (NA), melakukan aktivitas ensimatik sialolitik (sialolytic
enzymatic activity) dan melepaskan progeni virus yang terjebak di permukaan sel yang
terinfeksi sewaktu dilepaskan. Fungsi ini mencegah tertumpuknya virus dan mungkin juga memudahkan gerakan virus dalam selaput lendir dari jaringan epitel yang menjadi sasaran. Selanjutnya virus pun akan menempel ke sasaran. Ini membuat neoroamidase merupakan sasaran yang menarik bagi obat antivirus.
Virion akan masuk dan menyatu kedalam ruang endosom setelah berhasil melekat pada reseptor yang sesuai (Rust 2004). Virion akan mengalami degradasi dengan cara menyatukan membran virus dengan membran endosom melalui pemindahan proton terowongan protein dari matrix-2 (M2) virus, pada nilai pH di endosom sekitar 5.0. Proses selanjutnya akan terjadi serangkaian penataan ulang protein matrix-1 (M1) dan kompleks glikoprotein homotrimerik HA. Hasilnya adalah terbuka (exposed) sebuah bidang (domain) yang sangat lipofilik dan fusogenik dari setiap monomer HA yang masuk ke dalam membran endolisomal, dan dengan demikian memulai terjadinya fusi antara membran virus dengan membran lisomal. Berikutnya, kedelapan segmen RNA genomik dari virus, yang terbungkus dalam lapisan pelindung dari protein (ribonucleoprotein complex, RNP) nukleokapsid (N), dilepaskan ke dalam sitoplasma. Ke delepan segmen tersebut disalurkan ke nukleus untuk melakukan transkripsi mRNA
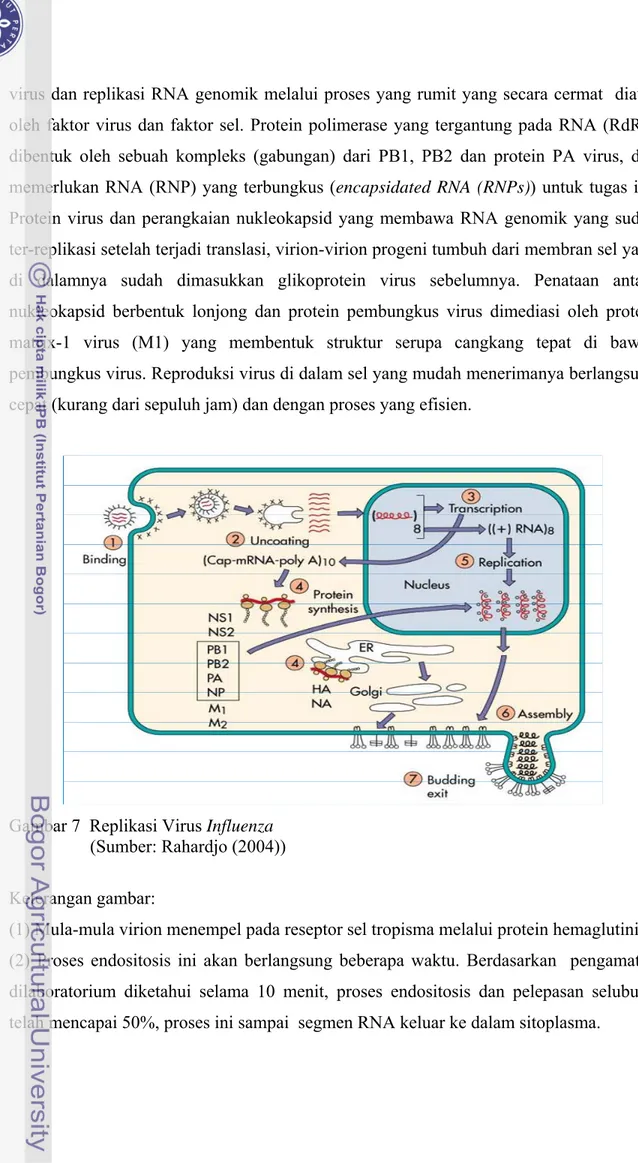
virus dan replikasi RNA genomik melalui proses yang rumit yang secara cermat diatur oleh faktor virus dan faktor sel. Protein polimerase yang tergantung pada RNA (RdRp) dibentuk oleh sebuah kompleks (gabungan) dari PB1, PB2 dan protein PA virus, dan memerlukan RNA (RNP) yang terbungkus (encapsidated RNA (RNPs)) untuk tugas ini. Protein virus dan perangkaian nukleokapsid yang membawa RNA genomik yang sudah ter-replikasi setelah terjadi translasi, virion-virion progeni tumbuh dari membran sel yang di dalamnya sudah dimasukkan glikoprotein virus sebelumnya. Penataan antara nukleokapsid berbentuk lonjong dan protein pembungkus virus dimediasi oleh protein matrix-1 virus (M1) yang membentuk struktur serupa cangkang tepat di bawah pembungkus virus. Reproduksi virus di dalam sel yang mudah menerimanya berlangsung cepat (kurang dari sepuluh jam) dan dengan proses yang efisien.
Gambar 7 Replikasi Virus Influenza (Sumber: Rahardjo (2004)) Keterangan gambar:
(1) Mula-mula virion menempel pada reseptor sel tropisma melalui protein hemaglutinin. (2) Proses endositosis ini akan berlangsung beberapa waktu. Berdasarkan pengamatan dilaboratorium diketahui selama 10 menit, proses endositosis dan pelepasan selubung telah mencapai 50%, proses ini sampai segmen RNA keluar ke dalam sitoplasma.
(3) Segmen-segmen tersebut masuk ke dalam ini sel (nukleus) dan mengalami transkripsi, untuk mengubah bentuk (-) RNA menjadi (+) RNA.
(4) Sebagian segmen keluar kembali ke sitoplasma untuk mempersiapkan protein selubung untuk dipakai oleh virus baru yang akan dihasilkan. Protein yang dimaksud adalah protein hemaglutinin, neuraminidase, matriks dan protein nonstruktural.
(5) Delapan segmen yang berada di inti sel ditambah dengan segmen RNA yang masih tersisa di sitoplasma melakukan replikasi yaitu perbanyakan RNA. Berbeda dengan virus RNA lainnya, dimana replikasinya terjadi di luar inti sel. Dengan berlangsung di dalam inti sel, AI menggunakan bahan-bahan yang diperlukan dari dalam inti sel inang. Proses ini memudahkan terjadinya proses Antigenic drift dan antigenic shift.
(6) Segmen RNA yang sudah mengalami replikasi, keluar ke sitoplasma untuk dibungkus dengan protein HA, NA dan M serta NS, menjadi anak AI yang siap dilepas dari sel inang. Untuk bisa keluar dari sel inang, virus baru ini akan menempel pada reseptor yang terdapat dalam sel inang. Penempelan ini dilakukan oleh protein neuraminidase bukan hemaglutinin seperti pada saat masuk ke sel. Proses ini bisa berlangsung selama 2 jam setelah infeksi (Rahardjo 2004).
2.7 Vaksin dan Vaksinasi Avian Influenza
Salah satu kebijakan pemerintah dalam mengendalikan penyakit AI di Indonesia adalah vaksinasi yang berpedoman pada ketentuan Badan Kesehatan Hewan Dunia (OIE 2004; OIE 2008). Program vaksinasi merupakan 1 (satu) dari 9 (sembilan) langkah strategi penanggulan HPAI yang diterapkan Pemerintah Indonesia sejak tahun 2004. Pada pelaksanaannya, kebijakan vaksinasi ini diterapkan dengan : (a) menggunakan vaksin yang sudah mendapatkan registrasi; (b) pada peternakan sektor 1, 2 dan 3 dilakukan secara swadaya; (c) pada peternakan sektor 4 pengadaan vaksin dan operasional vaksinasinya dibantu pemerintah; dan (d) program vaksinasi dilaksanakan dengan mengikuti SOP (Standar Operasional Prosedur) Pengendalian AI yang telah ditetapkan (Ditjennak 2008).
Vaksin AI mengacu pada ketentuan OIE (2008) adalah sebagai berikut:
1. Vaksin yang direkomendasikan penggunaannya adalah vaksin AI inaktif. Vaksin AI aktif konvensional tidak direkomendasikan.
2. Vaksin AI inaktif konvensional hanya boleh diproduksi dengan menggunakan
seed virus low pathogenic.
3. Vaksin bersifat imunogenik, dilihat dari respon pembentukkan antibodi yang tinggi.
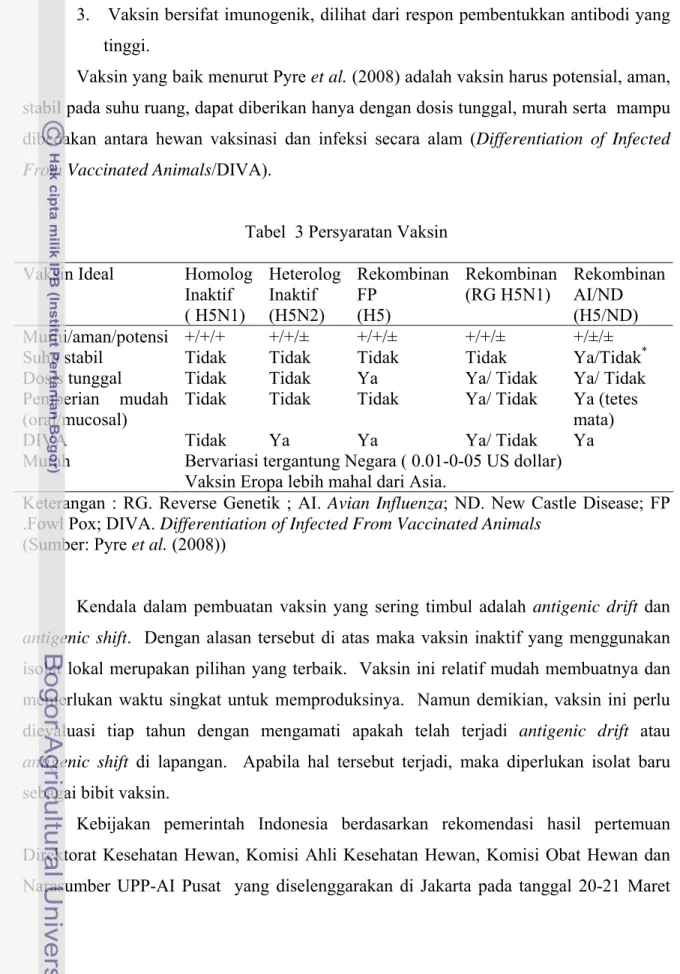
Vaksin yang baik menurut Pyre et al. (2008) adalah vaksin harus potensial, aman, stabil pada suhu ruang, dapat diberikan hanya dengan dosis tunggal, murah serta mampu dibedakan antara hewan vaksinasi dan infeksi secara alam (Differentiation of Infected
From Vaccinated Animals/DIVA).
Tabel 3 Persyaratan Vaksin Vaksin Ideal Homolog
Inaktif ( H5N1) Heterolog Inaktif (H5N2) Rekombinan FP (H5) Rekombinan (RG H5N1) Rekombinan AI/ND (H5/ND) Murni/aman/potensi +/+/+ +/+/± +/+/± +/+/± +/±/±
Suhu stabil Tidak Tidak Tidak Tidak Ya/Tidak*
Dosis tunggal Tidak Tidak Ya Ya/ Tidak Ya/ Tidak Pemberian mudah
(oral/mucosal)
Tidak Tidak Tidak Ya/ Tidak Ya (tetes mata)
DIVA Tidak Ya Ya Ya/ Tidak Ya
Murah Bervariasi tergantung Negara ( 0.01-0-05 US dollar) Vaksin Eropa lebih mahal dari Asia.
Keterangan : RG. Reverse Genetik ; AI. Avian Influenza; ND. New Castle Disease; FP .Fowl Pox; DIVA. Differentiation of Infected From Vaccinated Animals
(Sumber: Pyre et al. (2008))
Kendala dalam pembuatan vaksin yang sering timbul adalah antigenic drift dan
antigenic shift. Dengan alasan tersebut di atas maka vaksin inaktif yang menggunakan
isolat lokal merupakan pilihan yang terbaik. Vaksin ini relatif mudah membuatnya dan memerlukan waktu singkat untuk memproduksinya. Namun demikian, vaksin ini perlu dievaluasi tiap tahun dengan mengamati apakah telah terjadi antigenic drift atau
antigenic shift di lapangan. Apabila hal tersebut terjadi, maka diperlukan isolat baru
sebagai bibit vaksin.
Kebijakan pemerintah Indonesia berdasarkan rekomendasi hasil pertemuan Direktorat Kesehatan Hewan, Komisi Ahli Kesehatan Hewan, Komisi Obat Hewan dan Narasumber UPP-AI Pusat yang diselenggarakan di Jakarta pada tanggal 20-21 Maret
2009 adalah untuk menghasilkan vaksin yang baik dengan kualitas, efikasi dan keamanan yang tinggi serta potensi yang optimal diperlukan pemilihan master seed baru dengan prinsip kehati-hatian yang tinggi. Untuk memenuhi prinsip kehati-hatian tersebut, ditentukan kriteria master seed baru sebagai berikut:
1. Subtipe H5N1
2. Sifat imunogenesitas tinggi
3. Sifat antigenesitas dengan cakupan geografis yang luas. 4. Sifat genetik antigenik yang stabil.
5. Tingkat proteksi yang tinggi terhadap uji tantang dengan beberapa isolat virus yang berbeda karakter genetik dan antigeniknya (Ditjennak 2009).
Hingga tahun 2009, Indonesia telah menggunakan vaksin AI impor dan produksi dalam negeri yang masih menggunakan master seed Legok 2003 (HPAI H5N1) dan H5N2. Jumlah vaksin AI di Indonesia yang telah mendapat registrasi sementara dari Departemen Pertanian RI adalah sebanyak 24 vaksin (Lampiran 1) yang menggunakan
master seed, yaitu H5N1 (HPAI, isolat lokal, strain Legok), H5N1 (HPAI, isolat lokal,
strain Purwakarta), H5N2 (LPAI, strain England N-28), H5N2 (LPAI, strain Mexico 232), H5N2+ND (LPAI, strain England N-28 dan ND strain Lasota), H5N9 (LPAI, strain Wisconsin 68), dan H5N1 reverse genetic (isolat lokal Legok dengan Puerto Rico), dan vaksin vektor (H5N8 dengan FP 89) (Ditjennak 2009).
Strategi vaksinasi AI yang dilakukan di Indonesia berdasarkan prosedur operasional standar pengendalian penyakit AI (Ditjennak 2008) adalah sebagai berikut:
1. Vaksin AI yang digunakan adalah vaksin inaktif yang strain virusnya homolog dengan subtipe virus isolat lokal (H5) baik yang diproduksi secara konvensional maupun rekayasa genetika.
2. Vaksin yang digunakan harus telah mendapatkan nomor registrasi dari pemerintah c.q Direktorat jenderal Peternakan Departemen Pertanian.
3. Strategi vaksinasi dilaksanakan dengan ketentuan sebagai berikut: a. Pada daerah bebas dilarang melakukan vaksinasi;
c. Pada daerah kasus tinggi atau endemis, dilakukan vaksinasi secara tertarget.
4. Pelaksanaan vaksinasi tertarget tersebut dipilih pada populasi tertentu di daerah tertentu dengan kasus tinggi penyakit pada AI pada unggas, terdapat kasus manusia dan atau terdapat peternakan sektor1,2 dan 3 di sekitarnya. 5. Vaksinasi dilakukan 3-4 kali dalam satu tahun, dan atau sesuai petunjuk
produsen yang tertera pada etiket atau brosur. 6. Vaksinasi hanya dilakukan pada unggas yang sehat.
7. Cakupan vaksinasi meliputi seluruh populasi unggas terancam di daerah tertular yakni ayam buras, bebek, itik, entok, kalkun, angsa, burung merpati, burung puyuh, ayam ras petelur dan ayam ras pedaging yang termasuk peternakan sektor 4.
8. Vaksinasi dilakukan oleh petugas dinas dan atau kader/relawan desa terdidik, yang telah dilatih dan telah mengikuti pelatihan vaksinasi (vaksinator).
9. Program vaksinasi dilakukan dibawah pengawasan dokter hewan setempat. Strategi vaksinasi diharapkan dapat menurunkan kerentanan terhadap infeksi virus sekaligus mengurangi pengeluaran virus dari tubuh unggas, baik dari segi jumlah maupun lamanya waktu, sehingga merupakan alat yang tepat untuk menekan terjadinya kasus baru dan sirkulasi virus di lingkungan. Kesuksesan program vaksinasi dapat tercapai apabila vaksinasi dianggap sebagai alat untuk memaksimalkan tindak biosekuriti tanpa mengesampingkan pelaksanaan surveilans agar setiap perubahan antigenik virus yang bersirkulasi dilapangan dapat segera terdeteksi (Capua & Marangon 2006; Maas et al. 2007).
Efek perlindungan pasca vaksinasi merupakan respon kebal terhadap protein hemaglutinin (HA) pada permukaan virus dan atau neuraminidase (NA). Efek perlindungan yang diberikan oleh sebuah vaksin hanya terhadap subtipe HA individual yang terdapat dalam vaksin.
Pemeriksaan serologi dapat dilakukan untuk mengetahui adanya pembentukkan antibodi terhadap virus influenza A yang dapat diamati pada hari ke7 sampai ke 10 pasca infeksi. Pemeriksaan serologik yang dipakai adalah uji hambatan hemaglutinasi (Hemaglutination Inhibition/HI) untuk mengetahui adanya antibodi terhadap
hemaglutinin (H) dan uji Agar Gel Presipitasi (AGP) untuk mengetahui adanya antibodi terhadap neuraminidase (N) (Rahardjo 2004; WHO 2002; OIE 2008).