Vol. 4 No. 1. Hal 1-9 ISSN: 2087-7706
MORFOLOGI BUNGA YANG SESUAI BAGI KULTUR MIKROSPORA PADA
TANAMAN JARAK PAGAR (
Jatropha curcas
L.)
Appropriate Morphology of Physic Nut (
Jatropha curcas
L.) Flowers for
Microspore Culture
SUAIB*), MAKMUR JAYA ARMA, DAN MUHIDIN
Jurusan Agroteknologi Fakultas Pertanian Universitas Halu Oleo, Kendari
ABSTRACT
An experiment aimed in gaining information on morphological characteristics of
Jatropha curcas(L.) containing high frequency of uninucleate microspores has been done. This was important due to the successful application of experiment in haploid technique through anther or microspore culture is strongly determined by the availability of information from basic research such as appropriate morphology of the flowers. This scientific article, in our knowledge, was an early explanation of the initial steps of whole procedures in the microspore culture ofJatropha curcas(L.). Results of the research showed that the morphological features of flower that contained high frequency of mid- and late-uninucleate microspores was that the flowers must be in the range of 2 to 3 mm in length. Such flowers contained yellowish anthers.
Key w or ds: Flow er s length, mid- and late-uninucleate, mor phology, physic nut, micr ospor e
1
PENDAHULUAN
Kultur anter a atau kultur mikr ospor a mer upakan teknik per banyakan tanaman yang jauh lebih efisien dar i kultur jar ingan mer istem pucuk/ tunas, kultur suspensi sel, dan kultur pr otoplas. Hal ini dapat dicapai kar ena jumlah mikr ospor a dalam satu bunga, jauh lebih banyak jika dibandingkan dengan jumlah sel pada jar ingan mer istem pucuk/ tunas, suspensi sel, dan pr otoplas pada penggunaan satu pohon sumber eksplan. Setiap mikr o-spor a ber peluang untuk ber kembang menjadi individu tanaman lengkap apabila dikultur -kan di dalam medium yang kaya nutr isi kar ena sel ter sebut memiliki sifat totipotensi. Dengan demikian pengkajian penggunaan kultur mikr ospor a bagi per banyakan bibit unggul tanaman jar ak pagar sangat per lu dilakukan.
Penelitian kultur anter a dan atau kultur mikr ospor a pada tanaman jar ak pagar hi ngga saat ini belum per nah dilapor kan. Akan tetapi, apabila kultur mikr ospor a ber hasil dengan *)Alamat kor espondensi:
Email : suaib_06@yahoo.co.id
baik pada tanaman jar ak pagar sebagaimana telah ber hasil dengan baik pada ber bagai jenis tanaman lainnya, akan ber implikasi paling sedikit pada dua manfaat. Pertama, diper oleh bibit dalam jumlah yang lebih banyak pada w aktu yang lebih singkat, dan ke dua, diper oleh bibit bar u ber upa tanaman haploid ganda (doubled haploid) dengan konstitusi genetik homozigot sempur na. Sifat atau sifat -sifat yang dimiliki oleh tanaman haploid ganda disandikan oleh gen atau gen-gen homozigot. Dengan demikian, tanaman yang ber konstitusi genetik homozigot sempur na akan lebih optimal dalam mengekspr esi-kan sifat atau sifat-sifatnya jika dibandingkan dengan tanaman yang ber konstitusi genet ik heter ozigot.
Setiap pohon tanaman jar ak pagar dapat menghasilkan beber apa tangkai bunga, dan setiap bunga ter dir i dar i banyak kuncup bunga. Setiap kuncup bunga memiliki beber apa kepala sar i/ anter a (anthers) yang mengandung dua r uang sar i (theca). Setiap satu r uang sar i ter dapat paling sedikit 1.000 butir mikr ospor a (microspores) ter gantung var ietas dan ukur an kepala sar inya. Dengan demikian setiap tangkai bunga akan ter dapat
jutaan mikr ospor a, suatu jumlah yang jauh melebihi jumlah sel mer istematik bagi satu tanaman sumber eksplan.
Mikr ospor a adalah sel tunggal yang memiliki infor masi genetik yang lengkap dalam for masi separ uh dar i genom lengkapnya (haploid). Melalui kultur in vitro, setiap mikr ospor a ber potensi untuk ber difer ensiasi menjadi satu tanaman lengkap dengan konstitusi genetik haploid. Tanaman haploid yang dihasilkan akan mempr oduksi tepung sar i yang ster il sehingga akan gagal menghasilkan buah atau biji. Akan tetapi, melalui penggandaan kr omosom selama kultur mikr ospor a akan menghasilkan tanaman haploid ganda yang fer til dan dapat ber buah.
Untuk mendapatkan tanaman fer til dar i kultur mikr ospor a, pada fase ter tentu selama kultur in vitro dilakukan penggandaan kr omosom menggunakan bahan kimia mutagen. Bahan kimia mutagen yang umum digunakan adalah colchicine, apabila fr ekuensi tanaman haploid ganda yang dihasilkan secar a spontan sangat r endah. Selain colchicine, beber apa jenis bahan kimia lain dar i kelompok her bisida juga dilapor kan mempunyai car a ker ja yang sama dengan
colchicine dalam menggandakan kr omosom tanaman. Her bisida dimaksud adalah
amiprophosmethyle, oryzalin, dan trifluralin
(Hansen dan Ander sen, 1996).
Mikr ospor a yang dikultur kan secar a in vitro akan ber difer ensiasi membentuk pr oembr io, embr io, dan tanaman hijau dan lengkap apabila kebutuhan nutr isi dan faktor lain bagi per kembangannya ter penuhi selama kultur , sebaliknya akan melalui jalur pem-bentukan kalus apabila tidak ter penuhi. Pada fase kultur inilah diper lukan pengkajian yang seksama untuk mengidentifikasi dan mer umuskan faktor -faktor yang menentukan difer ensiasi mikr ospor a menjadi embr io, dan tanaman hijau dan lengkap.
Budidaya tanaman jar ak pagar sangat mudah dilakukan kar ena mudah tumbuh, mempunyai spektr um wilayah per tumbuhan yang luas, dan tidak ter lalu mensyar atkan suatu jenis tanah yang khusus. Akan tetapi, untuk kegiatan pembudidayaan yang luas dihadapkan pada masalah penyediaan bibit yang banyak pada waktu yang singkat. Untuk memecahkan masalah di atas, penyediaan bibit melalui teknik kultur jar ingan dapat
diter apkan sebagaimana telah ber hasil diper oleh dar i banyak penelitian. Satu di antar a banyak penelitian itu adalah yang dilakukan oleh Rajor e dan Batr a (2005) dimana dengan menggunakan mer istem pucuk dapat mer egener asikan sejumlah tanaman bar u.
Akan tetapi, melalui kultur sel-sel mer istem mempunyai kelemahan jika dibandingkan dengan kultur anter a dan atau kultur mikr ospor a. Kelemahan-kelemahan kultur sel mer istem antar a lain: (1) jumlah sel mer istem pada satu pohon sumber eksplan lebih sedikit dar i pada jumlah sel mikr ospor a yang ter dapat di dalam anter a, (2) tidak memper baiki sifat mewar is pada tanaman, sedangkan melalui kultur anter a dan atau mikr ospor a dapat dicapai per baikan genetik tanaman kar ena status genetiknya ber ubah menjadi haploid ganda, dan (3) ber peluang menghasilkan individu bar u tanaman yang memiliki sifat infer ior kar ena fenomena var iasi somaklon.
Ber bagai faktor yang mempengar uhi embr iogenesis mikr ospor a dan r egener asi “plantlet” menentukan efisiensi dan efektifitas kultur mikr ospor a. Kush dan Vir mani (1996), Palmer dan Keller (1997), Bhojw ani dan Bhatnagar (1999), Wang et al. (2000), dan Segui-Simar r o dan Nuez (2007) menyatakan bahw a paling sedikit ter dapat lima faktor yang mempengar uhi pembentukan embr io melalui kultur mikr ospor a. Kelima faktor dimaksud adalah: (1) genotipe tanaman donor , (2) kondisi per tumbuhan tanaman donor , (3) tahap per kembangan mikr ospor a, (4) pr aper lakuan, dan (5) medium kultur .
Khusus untuk tahap per kembangan mikr ospor a, secar a alamiah mikr ospor a akan ber kembang menjadi sel gamet jantan dan ber per an dalam zigot bagi pr oses pembentukan buah. Akan tetapi, secar a ar tifisial mikr ospor a dapat diinduksi untuk membentuk kalus atau embr io selama kultur
in vitro. Selanjutnya dir egener asikan hingga menjadi embr io dan tanaman. Mikr ospor a har us ber ada pada tahap per kembangan yang sesuai, yakni mulai dar i tahap uni -nukleat tengah “mid-uninucleate” hingga tahap binukleat aw al “ear ly-binucleate” bagi tanaman yang r esponsif.
Dalam kultur mikr ospor a akan dihasilkan tanaman bar u dengan konstitusi genetik separ uh (haploid) dar i genom penuhnya.
Haploid yang dilambangkan sebagai n adalah or ganisme, sel, atau inti sel yang memiliki hanya satu salinan kr omosom dar i setiap kr omosomnya (Maclean, 1987). Dengan demikian, tanaman haploid adalah tanaman yang hanya memiliki setengah dar i genom lengkapnya, sedangkan tanaman haploid ganda adalah tanaman yang memiliki genom haploid yang ber ganda.
Menur ut Johr i dan Rao (1984) dan Mor r ison dan Evans (1988), bahw a kultur mikr ospor a ber manfaat untuk : (1) per baikan klon, (2) menyingkat pr ogr am pemuliaan tanaman, dan (3) memudahkan seleksi secar a langsung pada suatu sifat kar ena disandikan oleh alel atau alel-alel homozigot sehingga ekspr esi sifat ter sebut menjadi optimal. Sementar a itu, Shar iatpanahi et al.(2006) dan Rimber a et al. (2007) mengemukakan bahwa manfaat pembentukan tanaman haploid melalui kultur mikr ospor a pada tanaman menyer buk sendir i adalah untuk: (1) memper cepat pr ogr am pemuliaan tanaman, (2) memper baiki efisiensi seleksi, (3) mendeteksi keter tautan dan inter aksi gen, (4) menduga var ians genetik dan jumlah gen bagi sifat-sifat kuantitatif, (5) menghasilkan tr anslokasi genetik, (6) menghasilkan galur -galur dengan penambahan kr omosom dan substitusi, dan (7) memudahkan studi tr ansfor masi genetik dan mutasi.
Suatu sifat yang disandikan oleh alel r esesif akan ter tutupi oleh alel dominan bila ber ada pada konstitusi heter ozigot. Akan tetapi, bila suatu sifat disandikan oleh alel heter ozigot maka tanaman ter sebut akan mempunyai individu mikr ospor a dengan konstitusi genetik haploid yang ber beda dan masing-masing membaw a alel r esesif atau dominan dar i sifat ter sebut. Bila mikr ospor a dimaksud dikultur kan, alel r esesif ter sebut akan seger a ter deteksi hanya dalam gener asi per tama hasil kultur mikr ospor a. Dengan demikian, suatu sifat yang disandikan oleh alel r esesif atau alel dominan, dengan mudah dapat dilakukan seleksi lebih awal.
Tulisan ini membahas cir i mor fologi bunga tanaman jar ak pagar yang banyak (lebih dar i 50 %) mengandung mikr ospor a pada tahap uninukleat tengah hingga uninukleat akhir . Infor masi cir i mor fologi ini sangat menentukan keber hasilan pelaksanaan kultur anter a dan atau kultur mikr ospor a kar ena pada fase mikr ospor a seper ti inilah yang bisa
membelokkan ar ah per kembangan mikr ospor a dar i status fungsional sebagai sel jantan (tepung sar i, “pollen gr ains”) menjadi embr io dan “plantlet” selama kultur in vitr o.
BAHAN DAN METODE
Satu batang tanaman donor sepanjang 20-30 cm yang diper oleh dar i petani, ditanam di dalam polibag ber ukur an 35-45 cm yang ber isi medium tanaman tanah lapisan atas dan pupuk kandang dengan per bandingan yang sama. Sebanyak 17 polibag yang ber isi masing-masing satu batang tanaman donor , ditata di lapangan tanpa pola r ancangan ter tentu. Setelah tumbuh, tanaman dipelihar a hingga ber bunga dengan pember ian pupuk NPK pada dosis yang sesuai, penyir aman medium tanaman secar a ber kala sesuai kebutuhan, dan pencegahan pengganggu dengan pestisida.
Setelah ber bunga, tangkai bunga dipetik dar i pohon utamanya, kemudian ber agam ukur an (diameter dan panjang) bunga diukur . Dengan menggunakan sepasang pinset ber ujung r uncing, anter a dikeluar kan dar i dalam bunga kemudian diletakkan di atas “deckglass” yang telah ditetesi lar utan 3% sukr osa. Mikr ospor a dikeluar kan dar i dalam anter a dengan membelah dinding anter a, sedangkan kotor an ber upa ser pihan anter a dibuang kemudian ditutup dengan “cover slip”. Tahap per kembangan mikr ospor a diamati dengan mikr oskop cahaya dalam dua kategor i, yaitu: (1) “uninucleate”, dan (2) “bi- dan multi-nucleate”. Cir i masing-masing tahap per kembangan mikr ospor a mengacu pada Zhang et al., (2005) dan Gonzalez dan Jouve (2005).
Var iabel yang diamati pada penelitian ini adalah: (1) jumlah bunga setiap tandan; (2) panjang dan diameter bunga; (3) panjang dan lebar anter a untuk semua ukur an bunga; (4) klasifikasi mikr ospor a ber dasar kan fase-fase per kembangannya melalui pew ar naan inti, dan (5) melakukan pencatatan infor masi kualitatif lainnya yang ber kaitan dengan mor fologi bunga.
Data hasil per hitungan ditabulasi, kemudian dihitung nilai r er ata dan simpangan bakunya. Semua per hitungan dilakukan dengan paket Statistika SAS (SAS Institute Inc., 1989-1996) menggunakan komputer .
HASIL
Penanaman tanaman jar ak pagar sebagai sumber mikr ospor a dilakukan sebanyak dua kali. Per tama dilakukan untuk tiga pot tanaman, dan kedua untuk 14 pot tanaman. Setiap pot ter dir i dar i satu batang atau satu pohon, sehingga selur uhnya ter dapat 17 pot atau 17 pohon. Sedangkan pemelihar aan tanaman dilakukan dalam bentuk pemupukan Ur ea, TSP, dan KCl, ber tur ut-tur ut dengan dosis 200, 100, dan 100 kg ha-1. Penyir aman
medium tanam dilakukan secar a ber kala sesuai fase per tumbuhan dan kebutuhan tanaman. Pencegahan jazad pengganggu per tumbuhan tanaman dilakukan secar a fisik dengan membunuh langsung jazad sasar an, dan secar a kimiaw i dilakukan dengan penyempr otan menggunakan insektisida atau pestisida.
Penampilan per tumbuhan dan per kembangan tanaman jar ak pagar sumber mikr o-spor a sebagai eksplan dalam penelitian ini di per lihatkan pada Gambar 1.
Gambar 1. Per tumbuhan dan per kembangan tanaman jar ak pagar sebagai sumber eksplan Penentuan cir i mor fologi bunga yang
mengandung mikr ospor a yang sesuai bagi kultur mikr ospor a, meliputi: (1) menghitung jumlah bunga setiap tandan; (2) mengukur panjang dan diameter setiap bunga; (3) mengukur panjang dan lebar anter a untuk semua ukur an bunga; (4) mengisolasi, menghitung, dan mengklasifikasikan mikr ospor a ber dasar kan fase per kembangannya melalui pew ar naan inti, dan (5) melakukan pencatatan infor masi kualitatif lainnya yang ber kaitan dengan mor fologi bunga. Umur pohon mulai ber bunga
per tama, lama w aktu yang diper lukan sejak inisiasi kuncup bunga hingga panen tandan atau r angkaian bunga untuk siap digunakan bagi pengamatan mor fologi tandan atau r angkaian bunga.
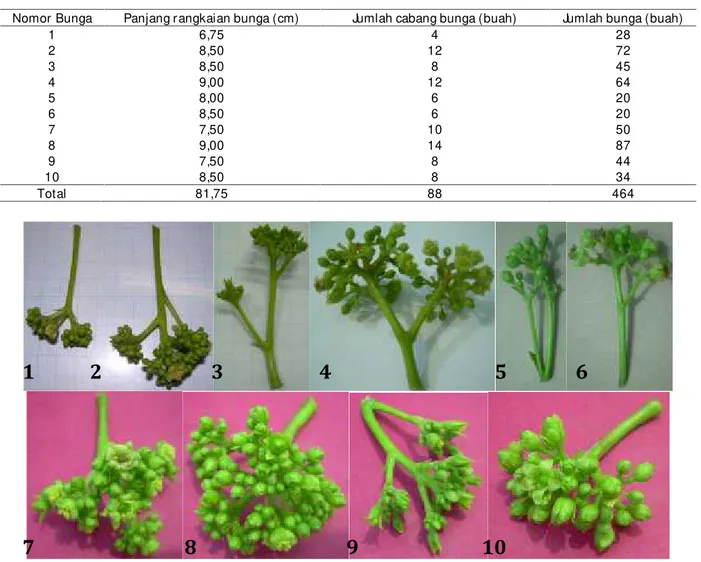
Rer ata panjang, jumlah cabang, dan jumlah bunga dar i 10 r angkaian bunga disajikan pada Tabel 1, sedangkan mor fologi ke sepuluh r angkaian bunga sebagai sampel dalam penelitian ini disajikan pada Gambar 2. Sementar a itu, r er ata lebar dan panjang bunga ber tur ut-tur ut dicantumkan pada Tabel 2 dan Tabel 3.
Tabel 1. Rer ata panjang r angkaian bunga, jumlah cabang bunga, dan jumlah bunga
Nomor Bunga Panjang r angkaian bunga (cm) Jumlah cabang bunga (buah) Jumlah bunga (buah)
1 6,75 4 28 2 8,50 12 72 3 8,50 8 45 4 9,00 12 64 5 8,00 6 20 6 8,50 6 20 7 7,50 10 50 8 9,00 14 87 9 7,50 8 44 10 8,50 8 34 Total 81,75 88 464
Gambar 2. Mor fologi 10 r angkaian bunga tanaman jar ak pagar yang diamati dalam penelitian.
Tabel 2. Rer ata lebar bunga
Nomor Bunga Jumlah Bunga Menur ut Ukur an Lebar (milimeter ) Total < 2 2 - < 3 3 - < 4 4 - < 5 5 - < 6 6 > 1 2 3 7 6 5 5 28 2 6 13 18 20 10 5 72 3 11 9 8 7 6 4 45 4 8 6 9 10 13 18 64 5 3 2 5 3 4 3 20 6 2 3 3 6 3 3 20 7 3 4 3 14 12 14 50 8 4 14 16 16 17 20 87 9 2 4 9 8 10 11 44 10 2 3 5 6 4 14 34 Total 43 61 83 96 84 97 464 Rer ata 4,3 6,1 8,3 9,6 8,4 9,7 46,6 Std. Dev. 3,09 4,38 5,10 5,38 4,70 6,50 22,41
1
2
3
4
5
6
7
8
9
10
Demikian pula, penampilan masing-masing bunga menur ut enam klasifikasi panjang dan lebar bunga ditampilkan pada Gambar 3.
Pengukur an r er ata panjang bunga menur ut 10 r angkaian bunga yang digunakan dalam penelitian ini menunjukkan ukur an yang ber agam. Hasil analisis masing-masing kelas ukur an panjang individu bunga, juga menunjukkan var iabilitas yang ber agam. Bunga tanaman jar ak pagar mempunyai lebar dan panjang yang hampir sama sehingga ukur an panjang r elatif sama dengan lebar nya, kecuali bagi bunga yang telah memasuki fase
penyer bukan atau fase dimana bakal biji telah ter bentuk. Bunga pada tahap ini mempunyai ukur an panjang yang lebih besar dar i lebar bunga sehingga bunga ter sebut nampak lebih panjang.
Ber dasar kan tahap per kembangan mikr ospor a, fr ekuensi mikr ospor a uninukleat tengah dan akhir diper oleh pada bunga dengan lebar dan panjang 2-3 mm (Tabel 5). Bunga yang ber ukur an panjang dan atau lebar lebih dar i 3 mm adalah mor fologi yang tidak sesuai bagi pelaksanaan kultur mikr ospor a secar ain vitro.
Gambar 3. Penampilan ukur an panjang dan lebar individu bunga tanaman jar ak pagar . Tabel 3. Rer ata panjang bunga
Nomor Bunga Jumlah Bunga Menur ut Ukur an Panjang (milimeter ) Total < 2 2 - < 3 3 - < 4 4 - < 5 5 - < 6 6 > 1 4 5 6 8 4 1 28 2 8 16 20 24 12 2 72 3 12 8 11 6 8 0 45 4 0 4 6 9 30 15 64 5 2 3 4 6 5 0 20 6 0 0 4 8 2 6 20 7 2 1 3 14 16 14 50 8 8 16 15 14 10 24 87 9 4 6 12 14 8 0 44 10 0 3 6 8 3 14 34
C
A
B
Tabel 4. Rer ata per sentase (%) mikr ospor a uninukleat t engah dan akhir menur ut panjang dan lebar bunga
Ulangan
Per sentase Mikr ospor a Uninukleat Tengah dan Akhir Menur ut Ukur an Panjang dan Lebar Bunga (milimeter )
< 2 2 - < 3 3 - < 4 4 - < 5 5 - < 6 6 > 1 20,46 80,68 10,24 2,86 0,00 0,00 2 18,26 89,76 5,83 0,82 0,00 0,00 3 10,36 86,94 4,42 1,84 0,00 0,00 Total 49,08 257,38 20,49 5,52 0,00 0,00 Rer ata 16,36 85,79 6,83 1,84 0,00 0,00 Std. Dev. 5,31 4,65 3,04 1,02 0,00 0,00
Gambar 4. Ster ilisasi bunga (A) sebelum isolasi ant er a (B dan C) dan mikr ospor a (D, E dan F). Sementar a itu, penampilan mikr ospor a
embr iogenik setelah mendapat pr aper laku-an str es menunjukkan per ubahan mor fologi, yaitu semakin ber tambahnya ukur an mikr ospor a. Besar nya per ubahan ukur an
kar ena ber tambahnya volume mikr ospor a minimal sebesar 100% atau paling kecil dua kali lebih besar dar i ukur an mikr ospor a non-embr iogenik atau mikr ospor a yang belum mendapat pr aper lakuan str es (Gambar 5).
Gambar 5. Mor fologi ber bagai mikr ospor a non-embr iogenik (panel atas) dan mikr ospor a embr iogenik (panel baw ah) set elah mendapat kombinasi pr aper lakuan str es suhu, lama inkubasi, dan medium star vasi.
A
B
C
PEMBAHASAN
Pengamatan mor fologi bunga yang mengandung mikr ospor a uninukleat ter banyak tidak ditentukan oleh panjang r angkaian bunga, akan tetapi lebih ditentukan oleh panjang dan lebar bunga. Hal ini ter lihat dar i populasi mikr ospor a uninukleat yang lebih besar yang sangat ber hubungan dengan ukur an bunga, bukan ukur an r angkaian bunga. Bunga dengan lebar dan panjang 2-3 mm, mengandung mikr ospor a uninukleat tengah dan akhir yang lebih tinggi jika dibandingkan dengan bunga dengan lebar dan panjang kur ang dar i 2 mm atau lebih dar i 3 mm. Bahkan bunga dengan panjang dan lebar < 2 mm mengandung mikr ospor a uninukleat aw al yang lebih tinggi, sedangkan bunga dengan panjang dan lebar > 3 mm mengandung tepung sar i dengan fr ekuensi yang lebih tinggi (Tabel 5).
Fenomena di atas umumnya telah dilapor kan oleh banyak peneliti atau kelompok penelit pada ber bagai tanaman budidya. Tanaman-tanaman dimaksud seper ti tanaman tembakau (Volcov et al., 2005),
Brassica (Zhang et al., 2006), kedelai (Rodr igueset al., 2006), tomat (Segui -Semar r o dan Nuez, 2007), dan kentang (Panahandeh et al., 2008). Bagi tanaman jar ak pagar , selain mor fologi bunga ber upa panjang dan lebar bunga, umur bunga juga menentukan fr ekuensi mikr ospor a uninukleat tengah dan akhir . Semakin tua umur bunga akan semakin tinggi fr ekuensi mikr ospor a pada tahap lebih lanjut atau polen sehingga tidak sesuai untuk digunakan sebagai eksplan bagi penyelenggar aan kultur mikr ospor a secar a in vitro.
Selain itu, ukur an anter a bagi beber apa spesies tanaman juga ber hubungan er at dengan fase per kembangan mikr ospor a. Pada tanaman cabai, ukur an dan w ar na anter a diketahui ber pengar uh ter hadap fase per kembangan mikr ospor a (Indr ianto et al., 2004; Supena et al., 2006). Indr ianto et al. (2004) mengur aikan bahw a mikr ospor a tanaman cabai besar pada tahap tetradhanya dijumpai pada kuncup bunga yang ber ukur an 0,3-0,5 cm. Anter a yang ber w ar na hijau muda, sebagian hijau dan sebagian ungu, akan mengan-dung mikr ospor a uninukleat. Setelah bunga ber ukur an 0,6-0,8 cm dan anter anya ber w ar na ungu muda, bunga hanya akan
mengandung mikr ospor a binukleat. Dengan demikian, tahap per kembangan mikr ospor a di dalam anter a sangat ditentukan oleh ukur an bunga dan anter a, sehingga penggunaan cir i mor fologi bunga atau ukur an kuncup mer upakan car a yang tepat dalam menentukan tahap per kembangan mikr ospor a.
SIMPULAN DAN SARAN
Hasil per cobaan dapat disimpulkan bahw a mor fologi bunga tanaman jar ak pagar yang sesuai bagi kultur mikr ospor a adalah ketika bunga mempunyai panjang dan lebar antar a 2-3 mm, dan ber umur hanya setelah beber apa har i setelah inisiasi bunga, dan dengan anter a ber w ar na kuning muda.
Dar i hasil penelitian ini bisa dijadikan sebagai infor masi awal bagi penelitian-penelitian lebih lanjut sehingga bisa ditemukan for mula yang lebih sesuai bagi tanaman jar ak pagar .
DAFTAR PUSTAKA
Bhojw ani SS, Bhatnagar SP, 1999. The embr yology of angiosper ms. 4th r evised and enlar ged edition, pp. 308-321. Vikas Publishing House PVT. LTD.
Gonzalez JM, Jouve N. 2005. Micr ospor e development dur ing in vit r o andr oge-nesis in tr iticale. Biologia Plantar um, 49(1): 23-28. Hansen NJP, Ander sen SB, 1996. In vit r o
chr omosome doubling pot ent ial of colchicine, or yzalin, tr iflur alin, and APM in Br assica napus micr ospor e cultur e. Euphyt ica, 88: 159-164. Indr ianto A, Semiar ti E, Sur ifah, 2004. Pr oduksi
galur mur ni melalui induksi embr iogenik mikr ospor a cabai mer ah dengan str es. Zur iat, 15(2): 133-139.
Johr i BM, Rao PS. 1984.Experimental embryology. Dalam: Johr i BM (ed.). Embr yology of Angiosper ms. pp. 735-802. Spr inger -Ver lag, Ber lin, Heidelber g, New Yor k, Tokyo.
Kush GS, Vir mani SS. 1996. Haploids in plant breeding. Dalam: Jain SM, Sopor y SK, Veilleux RE. (eds.) In VitroHaploid Pr oduction in Higher Plants. Vol.1. Fundamental Aspects and Methods. pp.12-17. Kluw er Academic Publis-her s, Dor dr echt, Boston, London.
Maclean N. 1987.Macmillan Dictionary Of Genetics and Cell Biology. Macmillan Refer ence Books. The Macmillan Pr ess Ltd. London Basingstoke, 422pp.
Mor r ison RA, Evans DA. 1988. Haploid plants fr om tissue cultur e: New plant var ieties in a
shor tened time fr ame. Bio/ Technology, 6: 684-690.
Panahandeh J, Valizadeh M, Khosroshahly M, Yermishin AP, Khoei FR, Mahna N. 2008. Micr ospor ogenesis and cr ossing behavior of a tet r aploid, inter specific int er -EBN hybr id potato. Scientiae Hor ticult ur ae, doi:10.1016/ j.scienta.2008.02.006
Palmer CE, Keller WA. 1997. Pollen embryos. Dalam: Shr ivanna KR, Saw ney VK (eds.). Pollen Biotechnology for Cr op Pr oduction and Impr ovement. pp. 392-422. Cambr idge Univer sity Pr ess. U.K.
Rajor e S, Batr a A. 2005. Efficient plant r egener ation via shoot tip cultur e in Jatr opha cur cas L. Jour nal of Plant Biochemist r y and Biotechnology, 14: 73-75.
Rimber a FK, Adaniya S, Ishimine Y, Etoh T. 2007. Mor phology of papaya plant s der ived via anther cultur e.Scientia Hor ticultur ae, 111: 213-219. Rodr igues LR, Camar go-For t e B.de,
Bodanese-Zanettini MH. 2006. Isolat ion and cultur e of soybean (Glycine max L. Mer r ill) micr ospor es and pollen gr ains.Br azilian Ar chives of Biology and Technology, 49(4): 537-545.
SAS Institut e Inc. 1989-1996. SAS/STAT User’s Guide Release 6.12. SAS Inst it ute, Inc., Car y, N.C. Segui-Simar r o JM, Nuez F. 2007. Embr yogenesis
induction, callogenesis, and plant r egener ation by in vitr o cultur e of tomato isolated micr ospor es and w hole anther s. Jour nal of Exper imental Botany, doi: 10.1093/ jxb/ er l271.
Shar iatpanahi ME. Belogr adova K, Hessamvasir i L, Heber le-Bor s E, Tour aev A. 2006. Efficient embr yogenesis and r egener ation in fr eshly isolated and cultur ed w heat (Triticum aestivum L.) micr ospor es w ithout str ess pr et r eatment. Plant Cell Repor ts, DOI 10.1007/ s00299-006-0205-7.
Supena EDJ, Musw ita W, Suhar sono S, Custer s JBM. 2006. Evaluation of cr ucial factor s for implementing shed-micr ospor e cultur e of Indonesian hot pepper (Capsicum annuum L.) cultivar s. Scientia Hor ticultur ae, 107: 226–232. Volcov RA, Ir ina, Panchuk I, Scho¨ffl F. 2005. Small
heat shock pr oteins ar e differ ent ially r egulated dur ing pollen development and follow ing heat str ess in tobacco. Plant Molecular Biology, 57:487–502.
Wang M, Van Ber gen S, Van Duijn B. 2000. Insight into a key development al sw itch and its impor tance for efficient plant br eeding. Plant Physiology, 124: 523-530.
Zhang Z, Lu Y, Liu X, Feng J, 2005. Nuclear and cell migr ation dur ing pollen development in r ice (Oryza sativa L). Sex Plant Repr oduct ion, 17: 297-302.
Zhang SF, Ma CZ, Zhu JC, Wang JP, Wen YC, Fu TD, 2006. Genetic analysis of oil cont ent in Brassica napus L. using mixed model of major gene and polygene. Acta Genetica Sinica, 33 (2): 171-180.