www.elsevier.com / locate / bres
Research report
Hypoxic modulation of striatal lesions induced by administration of
endothelin-1
*
Laibaik Park , Jim Thornhill
Department of Physiology and Saskatchewan Stroke Research Center, University of Saskatchewan, 107 Wiggins Road, Saskatoon, Saskatchewan,
Canada S7N 5E5
Accepted 22 August 2000
Abstract
Levels of endothelin-1 (ET-1), a potent endogenous vasoconstrictor, are elevated in plasma and cerebrospinal fluid (CSF) following cerebral ischemia and reperfusion injury. The present study sought insight into the potential differential vasoactive effects on the cerebral vasculature and resultant neural damage of ET-1 during normoxic vs. ischemic conditions and upon reperfusion. Under normoxic conditions, intrastriatal stereotaxic injection of exogenous ET-1 (40 pmol) induced a significant (P,0.05) reduction (#29612%) in the regional (striatal) cerebral blood flow measured by Laser Doppler flowmetry (CBFLDF) for up to 40 min in halothane-anesthetized male Long–Evans rats. Intrastriatal injection of ET-1 10 min after the onset of hypoxia (12% O , balance N ) tended to blunt, but not2 2 significantly, the striatal CBFLDF responses to the 35 min period of hypoxia. ET-1 given during reoxygenation significantly (P,0.05) reduced striatal CBFLDF, which was similar to the effect of ET-1 during normoxia. ET-1-induced infarction when administered prior to hypoxia, but not during or post-hypoxia, was significantly (P,0.05) exacerbated compared to infarction of ET-1 without hypoxia. These results suggest that exogenous ET-1 administered into the brain parenchyma can induce an infarction associated with modulation of CBFLDFduring the normoxic or reoxygenation period, but not during the hypoxic period and that the increased release of ET-1 in any pathological phase of cerebral ischemia contributes to irreversible neural damage with associated hemodynamic disturbances. 2000 Elsevier Science B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Ischemia
Keywords: Endothelin; Hypoxia; Laser Doppler flowmetry; Reoxygenation; Neural damage
1. Introduction induce vasodilation via the production of vasodilators [43].
The ET receptors are also expressed on neuronal cell types Endothelin (ET) family peptides (ET-1, ET-2, and ET- such as neurons, glial cells, epithelial cells of choroid 3), originally isolated from the supernatant of cultured plexus, and ependymal cells lining the ventricles of the endothelial cells, are potent vasoconstrictor agents that are brain [2,20]. Astrocytes are activated via ETB receptors present in several species, including human [56]. The [22] and the expression of these receptors was increased in activities of ET peptides are mediated by two types of rat neostriatum when cells were in a fully differentiated heterotrimeric G protein-coupled ET receptors (ETA and state [16].
ET ) [14]. The ETB A receptors have higher affinity for With global and focal ischemia and during subarachnoid ET-1 and ET-2 than ET-3, and are predominantly located hemorrhage of animals and humans, ET peptides are on the cerebrovascular smooth muscle cells to mediate elevated in plasma and CSF [1,3,26,39,58]. The increased vasoconstriction. ETB receptors have equal affinity for release of ET-1 in ischemic stroke suggests that this ET-1, ET-2, and ET-3 and are found on endothelial cells to endogenous peptide may be a contributing factor in the pathogenesis of this disease [15,58], possibly due to its potent vasoconstrictor properties. As ET-1 levels in
plas-*Corresponding author. Tel.: 11-306-966-6569; fax: 1
1-306-966-ma, CSF, and brain tissue are phasically increased with
6532.
E-mail address: [email protected] (L. Park). ischemic stroke or subarachnoid hemorrhage, it is
lated that elevated ET-1 levels that occur during the fluid replacement of blood taken through arterial catheter. various phases (e.g. hypoxic vs. reperfusion periods of Core body temperatures were servo-controlled at ischemic stroke) may compromise tissue perfusion of the 3760.28C via a heating pad connected to the temperature ischemic area in different ways. Under normoxic con- controller (Acadia Clinical).
ditions, topical or intraparenchymal application of ET-1 or
ET-3 induces potent and long-lasting vasoconstriction of 2.1.2. Induction of focal cerebral ischemia with ET-1 cerebral vessels to the point of inducing severe ischemia Procedures for the induction of focal cerebral ischemia and consequently irreversible ischemic injury using ET-1 (Sigma, St. Louis, MO, USA) were modified [9,10,12,32,41,46,52], but the vasoconstrictive property of from those published previously [9,10,52]. After catheteri-ET-1 under hypoxic and reoxygenation conditions is not zation of right femoral artery and vein, experimental known. In addition, pharmacological studies have attempt- animals were placed in a stereotaxic frame (David Kopf ed to correlate the hypoperfusion effects of ET-1 during Instruments). An incision was made along the midline of hypoxic-reperfusion periods of the ischemic stroke the scalp. Two burr holes were made, one for a Laser [2,8,35,36,49] with its resultant pathophysiological effects Doppler Pencil Probe (P-434) using coordinates of: an-on cerebral tissues during these different phases of is- terior 0.70 mm from bregma, lateral 20.18 mm from chemia, but the correlation remains inconsistent [33,37] midline, and ventral 6.0 mm from dura, according to the and largely unknown. Paxinos and Watson Rat Atlas [38] and the other for the The purpose of the present study was to determine: (a) 23-gauge guide cannula (coordinates noted below) in the the vasoactive effects of exogenous ET-1 given intrastriat- right frontal hemisphere. A Laser Doppler probe was ally during normoxic, hypoxic (12% O ), or reoxygenation2 attached to the guide cannula and was 2.0 mm lateral from periods on cerebral blood flow; and (b) if the timing the guide cannula. Relative changes of striatal cerebral between ET-1 administration and hypoxia altered the blood flow (CBFLDF) were continuously monitored via the resultant neuronal damage. Laser Doppler perfusion monitor (Laserflo Model BPM 403A, TSI, MN, USA) throughout the experiment in all groups. After steady state readings were obtained for
2. Materials and methods approximately 2–5 min, baseline control measurements of
CBFLDF were recorded for 30 min before intrastriatal 2.1. Animal preparation administration of sterile saline (SS) or ET-1. Since relative changes, but not absolute values, of CBFLDF are reliably 2.1.1. General measured with Laser Doppler flowmetry [6,17,41], the Experiments were done using Long–Evans rats aged relative change in cerebral blood flow over time following from 9 to 11 weeks obtained from Charles River Lab- SS, ET-1, hypoxia, or reoxygenation were determined oratories, Montreal, Quebec. Experimental animals were from respective baseline control values for each animal. maintained under constant environmental conditions with
ambient temperatures of 21618C, relative humidity of 2.2. Experimental protocol 30%, and a 12 h light and dark cycle. Animals had free
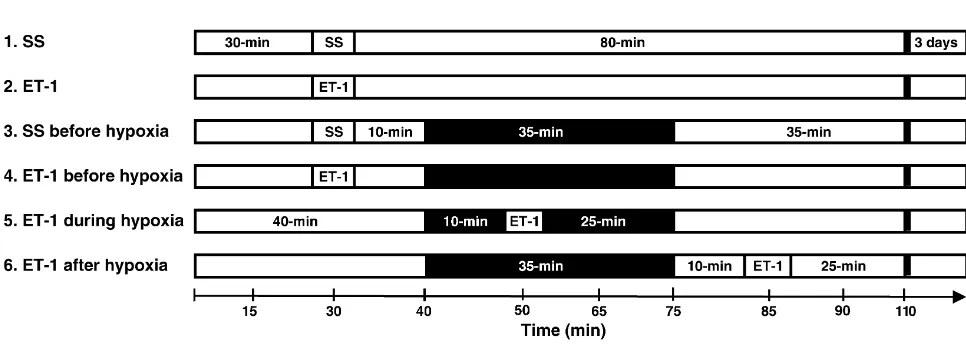
access to standard food pellets and tap water. The Guide- Rats were randomly assigned to one of six groups: lines of the Canadian Council of Animal Care in conjunc- Group 1, sterile saline (SS) (1 ml) was stereotaxically tion with the University Animal Care Committee were injected into the striatum and acted as a control group followed for all experimental procedures. without hypoxia (n510); Group 2, 1 ml of ET-1 (40 Animals were anesthetized with halothane (4% for pmol), as based on former studies for focal lesion induc-induction and 1.2% for maintenance) via nose cone in a tion [9,10,52], was stereotaxically injected into the mixture of 30% oxygen, balance nitrogen. Halothane levels striatum but no hypoxia was induced (n58); Group 3, SS were decreased to 0.7% in those groups receiving 35-min (1ml) was given into the striatum during normoxic period of hypoxia (12% O balance N ). Halothane concentration2 2 followed by the 35-min period of hypoxia (12% O2
needle was left in place for a further 5 min to prevent (physiological variables, CBFLDF and infarct volume) were reflex up the needle tract. Hypoxia was induced via a nose analyzed using the Student’s unpaired t-test, one-way or cone with 12% oxygen balanced in nitrogen for 35 min. the two-way analysis of variance (ANOVA) with repeated Schematic presentation of experimental protocol for each measures, or Mann–Whitney U-tests as appropriate. of the six groups is shown in Fig. 1. Statistical significance was accepted at the level of 0.01 or 0.05 of probability. All values in the text and figures are 2.3. Histopathological assessment presented as mean6S.E.M. In all instances, n refers to the number of individual animals in which observations were Three days after the experiment, the striatal lesions of all made.
groups were assessed. Rats were anesthetized with an intraperitoneal injection of sodium pentobarbital (100 mg /
kg) and the brains were perfusion-fixed transcardially with 3. Results
saline followed by a FAM solution (10% formaldehyde:
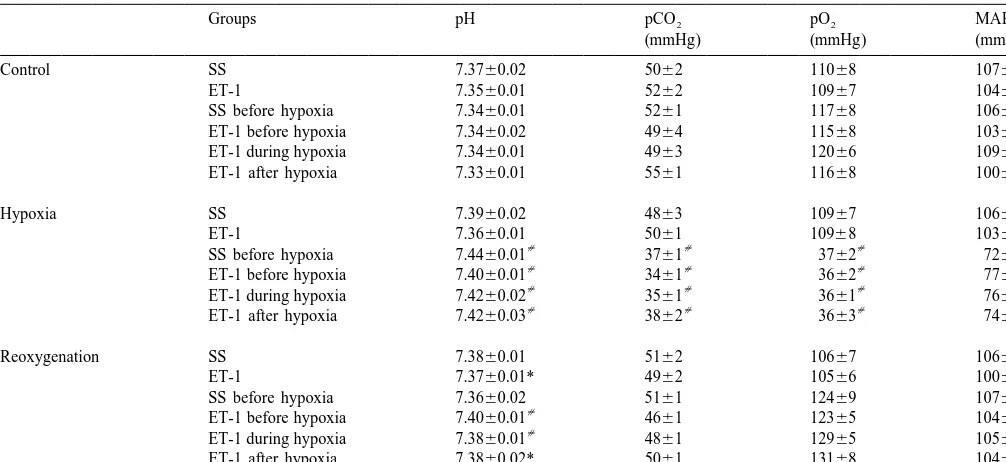
10% acetic acid: 80% methanol). Brains were then re- Table 1 shows changes of physiological variables (pH, moved and post-fixed in the same fixative overnight. Fixed pO , pCO , and MABP) in the six experimental groups.2 2
brains were embedded in paraffin, cut in 6 mm coronal There were no significant differences in respective physio-sections with an interval distance of 250 mm, and stained logical variables between the six experimental groups with hematoxylin–eosin (H&E) for assessment of infarct during control period. Intrastriatal injection of SS or ET-1 volume. The lesioned area of each section as defined by to the two groups that had no hypoxic episode caused no the respective staining and its size was determined by changes in the physiological variables measured. In those morphological analysis via an NIH image analyzer (Ver- four groups given a 35-min episode of hypoxia (12% O ),2
sion 1.61). The infarct volume of each brain was calcu- pH was markedly (P,0.01) increased, whereas pO ,2
lated by adding lesioned areas of each section multiplied pCO , and MABP were significantly (P2 ,0.01) decreased by the sum of the sectional interval (250 mm) and the from respective pre-hypoxic control values. Upon reoxyge-thickness of each section (6 mm). nation to normoxia in these four groups, pO , pCO , and2 2
MABP values returned to respective pre-hypoxic control 2.4. Statistical analysis values. However, pH values in all ET-1 groups with or without hypoxia remained elevated during the reoxygena-CBFLDFdata of the baseline control period were normal- tion period, compared to respective pre-hypoxic control ized by averaging CBFLDF values over a 30-min time values.
period before intrastriatal administration of SS or ET-1. Intrastriatal injection of SS caused no change in striatal Changes in CBFLDFfollowing intrastriatal SS or ET-1 with CBFLDFover the initial 30-min post-injection compared to or without hypoxia were expressed as a percentage of the pre-injection control period (Fig. 2A). From 30 to 70 mean CBFLDF of the 30-min baseline control period using min post-SS-injection, striatal CBFLDF slowly rose over the following formula: percentage change5[(df2dF ) / dF ]? time such that striatal CBFLDF was significantly above 100, where df is the flow after injection of ET-1 or SS and (P,0.05) pre-injection control values by 35 min after the dF is the mean flow of the baseline control period. Data injection. This gradual rise in striatal CBFLDF was also
Table 1
a
Physiological variables
Groups pH pCO2 pO2 MABP
(mmHg) (mmHg) (mmHg)
Control SS 7.3760.02 5062 11068 10762
ET-1 7.3560.01 5262 10967 10462
SS before hypoxia 7.3460.01 5261 11768 10662
ET-1 before hypoxia 7.3460.02 4964 11568 10362
ET-1 during hypoxia 7.3460.01 4963 12066 10963
ET-1 after hypoxia 7.3360.01 5561 11668 10063
Hypoxia SS 7.3960.02 4863 10967 10662
ET-1 7.3660.01 5061 10968 10361
[ [ [ [
SS before hypoxia 7.4460.01 3761 3762 7265
[ [ [ [
ET-1 before hypoxia 7.4060.01 3461 3662 7762
[ [ [ [
ET-1 during hypoxia 7.4260.02 3561 3661 7662
[ [ [ [
ET-1 after hypoxia 7.4260.03 3862 3663 7466
Reoxygenation SS 7.3860.01 5162 10667 10664
ET-1 7.3760.01* 4962 10566 10062
SS before hypoxia 7.3660.02 5161 12469 10764
[
ET-1 before hypoxia 7.4060.01 4661 12365 10462
[
ET-1 during hypoxia 7.3860.01 4861 12965 10562
ET-1 after hypoxia 7.3860.02* 5061 13168 10465
a [
Values are the mean6S.E.M. (N55 for each group). * P,0.05 or P,0.01 compared with the control values of each group. MABP, mean arterial blood pressure.
Fig. 2. Regional (striatal) cerebral blood flow measured by laser Doppler flowmetry (CBFLDF) are shown following intrastriatal injections of (A) SS (1ml) (♦, n510) or ET-1 [40 pmol] (m, n58) without hypoxia or (B) SS (1ml) (♦, n55) with hypoxia. Data are presented as mean percentage changes of CBFLDFof the striatum6S.E.M. compared to respective pre-injection values. * P,0.05 denotes significant difference in striatal CBFLDFwithin that group
[
seen in other sham treated rats in which no SS was injected the absence of a hypoxic episode. Animals given intra-intrastriatally (data not shown), thus this response of the striatal ET-1 during hypoxia or upon reoxygenation had control group appears related to the effects of anesthesia infarct volumes similar to that of the group given ET-1 [25]. Intrastriatal injection of ET-1 (40 pmol) significantly without hypoxia.
(P,0.05) decreased the striatal CBFLDF(mean peak fall of 29612%) compared to pre-injection values over the initial
40 min post-injection period (Fig. 2A). The CBFLDF after 4. Discussion
ET-1 administration remained significantly (P,0.01)
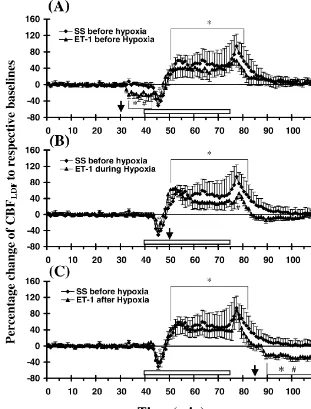
below CBFLDF responses of the SS control group even As ET-1 is a potent and long-lasting vasoconstrictor of after CBF responses of that group returned to pre-injection arteries [56], the involvement of ET-1 was studied as a control values. potential causal factor in the pathogenesis of hypoxic Induction of hypoxia (12% O ) 10 min after an intra-2 ischemia. The present data show that the exogenous ET-1 striatal injection of SS (Fig. 2B) evoked a triphasic administered into the striatum at different time points with response in striatal CBFLDF. An initial, transient decrease respect to an episode of global hypoxia significantly (P,0.05) in striatal blood flow occurred, compared to reduced cerebral blood flow during normoxia and reoxyge-pre-hypoxic control values of that group, followed by a nation, but not during hypoxia (Figs. 2 and 3). All groups more prolonged increase above baseline values over the given ET-1 with or without hypoxia were significantly last 25 min of the hypoxic period. There was a further damaged from respective saline controls with the most escalation in CBFLDF upon return to normoxic conditions severe neural damage induced when ET-1 was given in this group. Further analysis of this consistent triphasic before the hypoxic challenge (Fig. 4).
CBFLDF response to hypoxia revealed that striatal CBFLDF Immunohistochemistry and in situ hybridization have initially dropped to 51625% of baseline values with confirmed the presence of ET, along with its receptor hypoxia, rebounded to be 60628% above baseline values subtypes ET and ET , in neurons, glia, and cerebrovascu-A B
during the latter phase of the hypoxic period, and then lar endothelium [13,27,31,50,57]. ET-1 is processed by rapidly increased further to 94630% above baseline endothelin-converting enzyme from inactive big ET-1 to control values upon reoxygenation, before returning to active ET-1 and is produced by both regulated and control values after 10 min of reoxygenation. constitutive pathways in response to important stimuli such Fig. 3A shows that the intrastriatal injection of ET-1 10 as hypoxia, ischemia, or shear stress [29]. ETs induce min before hypoxia decreased striatal CBFLDFcompared to potent and long-lasting constriction of cerebral arteries in its own pre-injection control values and the CBFLDFvalues vivo and in vitro, normally by the activation of ETA
21
of the SS group. However, upon the induction of hypoxia, receptors and, in part, by enhanced Ca entry via
voltage-21
the CBFLDF responses to the ET-1 injection became gated Ca channels [15,44]. The activation of ETB
insignificant from those of the SS group, though the receptors can produce either vasodilation, via ETB1 re-CBFLDF responses were blunted in general. When ET-1 ceptors on the endothelium, or vasoconstriction through was given during the hypoxic period (Fig. 3B), the ETB2 receptors on smooth muscle cells. Most of ET-1 is CBFLDF responses of this group during hypoxia were released locally to the abluminal interface between endo-similar, though blunted, to CBFLDF responses of the group thelium and smooth muscle where it can activate ETA / B2
given SS 10 min before hypoxia induction. However, the receptors on smooth muscle cells and mediate vasocon-increase in CBFLDF seen upon reoxygenation in the ET-1 striction in a paracrine manner.
treated group was significantly (P,0.05) reduced from the The plasma level of ET-1 in stroke patients is markedly initial CBFLDFincrease seen upon reoxygenation in the SS elevated and correlated with more severe neurologic group. ET-1 given during reoxygenation (Fig. 3C) sig- deficits [58]. As well, ET-1 levels in the CSF of stroke nificantly (P,0.05) decreased striatal CBFLDF from re- patients is significantly increased during the 18 h after spective pre-hypoxic and SS-injected control values. The stroke [26]. Following middle cerebral artery occlusion in magnitude of CBFLDF reduction in this group was similar rabbits, acute, marked increases of ET-1 in brain tissue and to the response evoked by ET-1 given without hypoxia. plasma were associated with the focal ischemic event [3]. The mean focal infarct volumes as assessed from serial Brain extracellular fluid ir-ET-1 levels have also been H&E stained sections 3 days after experimentation are shown to be significantly increased during ischemic and shown in Fig. 4. There was no significant difference in reperfusion periods in a rat model of transient forebrain infarct volume between SS-treated groups with or without ischemia [1]. In transient forebrain ischemia of gerbils, the hypoxia. Infarct volume was significantly increased (P, treatment of BQ-123, an ET receptor antagonist, caused aA
0.01) in all ET-1 treatment groups compared to SS-treated marked neuroprotection in the CA1 region of the
hip-3 3
groups with (0.960.8 mm ) or without (0.760.2 mm ) a pocampus [8]. Furthermore, BQ-123 given prior to is-hypoxic episode. Infarct volume of the group given ET-1 chemia reversed the early incomplete recovery of cerebral
3
Fig. 3. Regional (striatal) CBFLDFresponses are shown following intrastriatal injection of ET-1 (40 pmol) (A) 10 min before (n58), (B) during (n58), and (C) 10 min after the 35 min period of hypoxia (n510). Data are presented as mean percentage changes of CBFLDF6S.E.M. compared to respective pre-injection values. CBFLDF responses of SS group to hypoxia were shown for comparison to respective ET-1 groups. * P,0.05 denotes significant
[
difference in striatal CBFLDF within that group compared to respective pre-injection values and P,0.05 represents significant difference in striatal CBFLDFbetween the ET-1 and SS control groups at that time. Rectangular box above X-axis indicates the 35 min period of hypoxia. Arrows indicate the time of SS or ET-1 injections.
gerbils treated with N√-nitro-L-arginine (NLA), a NOS ischemic neural damage with an associated marked
reduc-inhibitor [49]. Pretreatment of spontaneously hypotensive tion of cerebral blood flow when administered during rats with SB 217242, a nonselective ET / ET -receptorA B hypoxic and reperfusion periods.
antagonist, also reduced ischemic brain injury [2]. These Ischemic effects of administered exogenous ETs have findings suggest that the production / release of ET-1 been previously shown to be caused by the profound increases during the development of ischemic stroke. This reduction of local cerebral blood flow [10,11,18,32,42,46], enhanced release of ET-1, together with a possible in- responses mostly thought to be mediated by ET receptorsA
creased availability of ET-1 binding sites through exter- [18,36] although mild contractions may be elicited by ETB
adminis-3
Fig. 4. Mean volumes of ischemic infarct (mm6S.E.M.) are shown for the six experimental groups. * Indicates significant increase (P,0.01) in infarct
[
volume compared to either SS treatment group and denotes significant increase (P,0.05) in infarct volume from all other ET-1 treatment groups.
tered during normoxic and reoxygenation periods, but not of a cerebral blood vessel depends on the degree and when given during 12% hypoxia. Previous studies demon- duration of impaired blood flow [40]. The exact explana-strated that bound (incubated) ET-1 is difficult to displace tion for this increased infarction with administration of by specific receptor antagonists due to the high binding ET-1 prior to hypoxia remains unclear, but might result affinity and the slow dissociation rate of ET-1 [55]. In from the secondary metabolic stress exposed by hypoxia addition, the occupancy of ETA receptors by endogenous on selectively vulnerable cells within a rim of moderately ET-1 is sufficient to promote rapid endocytosis [4] so that ischemic tissue between normally perfused areas and the receptor binding affinity of ET-1 may be reduced. It is evolving infarct in which pathological processes are spa-likely that the binding sites available for exogenous ET-1 tially and temporally dynamic.
[7] H. Ehrenreich, T. Costa, K.A. Clouse, R.M. Pluta, Y. Ogino, J.E.
required to induce an ischemic insult. Indeed, even higher
Coligan, P.R. Burd, Thrombin is a regulator of astrocytic ET-1,
doses of exogenous ET-1 could have been utilized to
Brain Res. 600 (1993) 201–207.
induce focal ischemia [46]. It must be noted that the [8] G. Feuerstein, J.L. Gu, E.H. Ohlstein, F.C. Barone, T.L. Yue, abluminal production of ET-1 is reported to be up-reg- Peptidic endothelin-1 receptor anatagonist, BQ-123, and
neuro-protection, Peptides 15 (3) (1994) 467–469.
ulated by numerous cells including astrocytes, neurons,
[9] K. Fuxe, B. Bjelke, B. Andbjer, H. Grahn, R. Rimondini, L.F.
pituitary cells, smooth muscle cells, and infiltrating
macro-Agnati, Endothelin-1 induced lesions of the fronto-parietal cortex of
phages in response to ischemic stimuli [7,13,23,27,39,57]. the rat. A possible model of focal cortical ischemia, Neuroreport 8 Thus, high concentrations of ET-1 can be locally released (1997) 2623–2629.
˚ ¨
[10] K. Fuxe, N. Kurosawa, N. Cintra, A. Hallstrom, M. Goiny, L.
under conditions of ischemia, yet are difficult to detect and
`
Rosen, L.F. Agnati, L.E. Ungerstedt, Involement of local ischemia
reliably measure. This local release may be more harmful
in endothelin-1 induced lesions of the neostriatum of the
anes-to ischemic brain tissue since ET-1 release, abluminally thetized rat, Exp. Brain Res. 88 (1992) 131–139.
rather than luminally, causes more prolonged vasoconstric- [11] K. Fuxe, W. Staines, A. Hemsen, L. Hersh, J.M. Lundberg,`
tion and hypoperfusion of cerebral microvasculature Demonstration and nature of endothelin-3-like immunoreactivity in somatostatin and choline acetyltransferase-immunoreactive nerve
[42,48,53,54].
cells of the neostriatum of the rat, Neurosci. Lett. 123 (1991)
In conclusion, these results indicate that exogenous
ET-107–111.
1 does induce ischemic injury, with or without a concur- [12] G. Gartshore, J. Patterson, I.M. Macrae, Influence of ischemia and rent significant reduction of CBF, and infers that en- reperfusion on the course of brain tissue swelling and blood–brain barrier permeability in a rodent model of transient focal cerebral
dogenously released ET-1 from the ischemic episode itself
ischemia, Exp. Neurol. 147 (1997) 353–360.
could further compromise hemodynamic disturbances
in-[13] A. Giaid, S.J. Gibson, N.B.N. Ibrahim, S. Legon, S.R. Bloom, M.
cluding no-reflow phenomenon and (post) ischemic hypo- Yanagisawa, T. Masaki, I.M. Varndell, J.M. Polak, Endothelin 1, an perfusion [21] during any pathological phase of cerebral endothelium-derived peptide, is expressed in neurons of the human spinal cord and dorsal root ganglia, Proc. Natl. Acad. Sci. USA 86
ischemia. However, further studies using specific ET
(1989) 7634–7638.
receptor antagonists will be of great importance for
[14] K. Goto, H. Hama, Y. Kasuya, Molecular pharmacology and
elucidating a more specific role of ET-1 in the pathological pathophysiological significance of endothelin, Jpn. J. Pharmacol. 72 phases of cerebral ischemia. (1996) 261–290.
[15] D.A. Greenberg, J. Chan, Endothelins and the central nervous system, Neurology 42 (1992) 25–31.
[16] H. Hama, T. Sakurai, Y. Kausya, M. Fujiki, T. Masaki, K. Goto,
Acknowledgements Astrocytes was liberated in its state from fully differentiated version
by endothelin-1, Jpn. J. Pharmacol. 58 (1992) 158.
[17] A. Heimann, S. Kroppenstedt, P. Ulrich, O.S. Kempski, Cerebral
This work was supported by the Heart and Stroke
blood flow autoregulation during hypobaric hypotension assessed by
Foundation of Saskatchewan. The authors wish to thank
laser Doppler scanning, J. Cereb. Blood Flow Metab. 14 (1994)
Johanne Asselin for technical assistance and Gloria
1100–1105.
Schneider for typing the manuscript. [18] D.C. Henshall, S.P. Butcher, J. Sharkey, A rat model of
endothelin-3-induced middle cerebral artery occlusion with controlled reperfu-sion, Brain Res. 843 (1999) 105–111.
[19] F.M. Hofman, P. Chen, R. Jeyaseelan, F. Incardona, M. Fisher, R.
References Zidovetzki, Endothelin-1 induces the production of the neurophil
chemotactic factor IL-8 by human brain-derived endothelial cells, [1] F.C. Barone, Y.T. Globus, W.J. Price, R.F. White, B.L. Storer, G.Z. Blood 92 (1998) 3064–3072.
¨ ¨
Feuerstein, R. Busto, E.H. Ohlstein, Endothelin levels increases in [20] E. Hosli, L. Hosli, Autoradiographic evidence for endothelin re-rat focal and global ischemia, J. Cereb. Blood Flow Metab. 14 ceptors on astrocytes in cultures of rat cerebellum, brain stem, and
(1993) 337–342. spinal cord, Neurosci. Lett. 129 (1991) 55–58.
[2] F.C. Barone, R.F. White, J.D. Elliott, G.Z. Feuerstein, E.H. Ohlstein, [21] K.A. Hossman, Ischemia-mediated neuronal injury, Resuscitation 26 The endothelin receptor antagonist SB 217242 reduces cerebral (1993) 225–235.
focal ischemic brain injury, J. Cardiovasc. Pharmacol. 26 (S3) [22] N. Ishikawa, M. Takemura, Y. Koyama, Y. Shigenaga, T. Okada, A. (1995) S404–S407. Baba, Endothelins promotes the activation of astrocytes in rat [3] L.G. Bian, T.X. Zhang, W.G. Zhao, J.K. Shen, G.Y. Yang, Increased neostriatum through ET(B) receptors, Eur. J. Neurosci. 9 (1997)
endothelin-1 in the rabbit model of middle cerebral artery occlusion, 895–901.
Neurosci. Lett. 174 (1994) 47–50. [23] H. Kasuya, B.K.A. Weir, D.M. White, K. Stefansson, Mechanism of [4] N. Bhowmick, P. Narayan, D. Puett, The endothelin subtype A oxyhemoglobin-induced release of ET-1 from cultured endothelial receptor undergoes agonist- and antagonist-mediated internalization cells and and smooth-muscle cells, J. Neurosurg. 79 (1993) 892– in the absence of signaling, Endocrinology 139 (1998) 3185–3192. 898.
¨ ¨
[5] F. Blomstrand, C. Giaume, E. Hansson, L. Ronnback, Distinct [24] M.S. Kramer, P.E. Vinalli, L.I. Katolik, F.A. Simeone, Comparison pharmacological properties of ET-1 and ET-3 on astroglial gap of cerebral blood flow by laser-Doppler flowmetry and hydrogen
21
junctions and Ca signaling, Am. J. Physiol. 277 (Cell Physiol. 46) clearance in cats after cerebral insult and hypervolemic
hemodilu-(1999) C616–C627. tion, Neurosurgery 38 (1996) 355–361.
[6] U. Dirnagl, B. Kaplan, M. Jacewicz, W. Pulsinelli, Continuous [25] Y. Kuroda, M. Murakami, J. Tsuruta, T. Murakawa, T. Sakabe, measurement of cerebral cortical blood flow by laser-Doppler Blood flow velocity of middle cerebral artery during prolonged flowmetry in a rat stroke model, J. Cereb. Blood Flow Metab. 9 anesthesia with halothane, isoflurane, and sevoflurane in humans,
[26] Y. Lampl, G. Fleminger, R. Gilad, R. Galron, I. Sarova-Pinhas, M. [44] S.B. Salom, G. Torregrosa, E. Alborch, Endothelins and cerebral Sokolovsky, Endothelin in cerebrospinal fluid and plasma of patients circulation, Cerebrovasc. Brain Metab. Rev. 7 (1995) 131–152.
¨
in the early stage of ischemic stroke, Stroke 28 (1997) 1951–1955. [45] Y. Sasaki, M. Takimoto, K. Oda, T. Fruh, M. Takai, T. Okada, S. [27] M.E. Lee, S.M. dela Monte, S.C. Ng, K.D. Bloch, T. Quertermous, Hori, Endothelin evokes efflux of glutamate in cultures of rat
Expression of the potent vasoconstrictor endothelin in the human astrocytes, J. Neurochem. 68 (1997) 2194–2200.
central nervous system, J. Clin. Invest. 86 (1990) 141–147. [46] J. Sharkey, S.P. Butcher, J.S. Kelly, Endothelin-1 induced middle [28] K.H. Leseth, M. Adner, H.K. Berg, L.R. White, J. Aasly, L. cerebral artery occlusion: pathological consequences and neuro-Edvinsson, Cytokines increase endothelin ETB receptor contractile protective effects of MK801, J. Auton. Nerv. Syst. 49 (1994) activity in rat cerebral artery, Neuroreport 10 (1999) 2355–2359. S177–S185.
[29] E.R. Levin, Endothelins, New Engl. J. Med. 333 (1995) 356–363. [47] H. Shibaguchi, Y. Kataoka, S.S. Koizumi, M. Kohzuma, M. Obana, [30] J. Liu, D.J. Casley, W.G. Nayler, Ischemia causes externalization of A. Himeno, K. Yamashita, K. Taniyama, Nitric oxide participates in ET-1 binding sites in rat cardiac membranes, Biochem. Biophys. the stimulatory and neurotoxic action of endothelin on rat striatal Res. Commun. 164 (1989) 1220–1225. dopaminergic neurons, Cell. Mol. Neurobiol. 17 (1997) 471–481. [31] M. MacCumber, C. Ross, S.H. Snyder, Endothelin in brain: re- [48] T. Shigeno, T. Mima, K. Takakura, M. Yanagisawa, A. Saito, K.
ceptors, mitogenesis, and biosynthesis in glial cells, Proc. Natl. Goto, T. Masaki, Endothelin-1 acts in cerebral arteries from the Acad. Sci. USA 87 (1990) 2359–2363. adventitial but not from the luminal side, J. Cardiovasc. Pharmacol. [32] I.M. Macrae, M.J. Robinson, D.I. Graham, J.L. Reid, J. McCulloch, 13 (Suppl. 5) (1991) S174–S176.
Endothelin-1-induced reductions in cerebral blood flow: dose depen- [49] M. Spatz, Y. Yasuma, A. Strasser, R.M. McCarron, Cerebral dency, time course, and neurological consequeces, J. Cereb. Blood postischemic hypoperfusion is mediated by ETA receptors, Brain Flow Metab. 13 (1993) 276–284. Res. 726 (1996) 242–246.
[33] M.A. McAuley, V. Breu, D.I. Graham, J. McCulloch, The effects of [50] K. Takahashi, M. Ghatei, P.M. Jones, J.K. Murphy, H.C. Lam, D.J. bosentan on cerebral blood flow and histopathology following O’Halloran, S.R. Bloom, Endothelin in human brain and pituitary middle cerebral artery occlusion in the rat, Eur. J. Pharmacol. 307 gland: presence of immunoreactive endothelin, endothelin mRNA (1996) 171–181. and endothelin receptors, J. Clin. Endocrinol. Metab. 72 (1990) [34] M.A. Murray, F.M. Faraci, D.D. Heistad, Effect of protein kinase C 693–699.
inhibitors on endothelin- and vasopressin-induced constriction of the [51] O. Touzani, S. Galbraith, P. Siegl, J. McCulloch, Endothelin-B rat bisilar artery, Am. J. Physiol. 263 (1992) H1643–H1649. receptors in cerebral resistance arteries and their functional signifi-[35] E.H. Ohlstein, P. Nambi, S.A. Douglas et al., SB 209670, arationally cance after focal cerebral ischemia in cats, J. Cereb. Blood Flow
designed potent nonpeptide endothelin receptor anatgonist, Proc. Metab. 17 (1997) 1157–1165. ˚
´ ¨
Natl. Acad. Sci. USA 91 (1994) 8052–8056. [52] A. Ueki, L. Rosen, B. Andbjer, A. Hallstrom, M. Goiny, S. [36] T.R. Patel, Therapeutic potential of endothelin antagonists in Tanganelli, U. Ungerstedt, M. Fuxe, Evidence for a preventive cerebrovascular disease, CNS Drugs 5 (1996) 293–310. action of the vigilance-promoting drug nodafinil against striatal [37] T.R. Patel, J. McCulloch, Failure of an endothelin antagonist to ischemic injury induced by endothelin-1 in the rat, Exp. Brain Res.
modify hypoperfusion after transient global ischemia in the rat, J. 96 (1993) 89–99.
Cereb. Blood Flow Metab. 16 (1996) 490–499. [53] R.N. Willette, C.F. Sauermelch, Abluminal effects of endothelin in [38] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates, cerebral microvasculature assessed by laser Doppler flowmetry, Am.
2nd Edition, Academic Press, San Diego, CA, USA, 1986. J. Physiol. 28 (1990) H1688–H1693.
[39] R.M. Pluta, R.J. Boock, J.K. Afshar, K. Clouse, M. Bacic, H. [54] R.N. Willette, C.F. Sauermelch, M. Ezekiel, G. Feuerstein, E.H. Ehrenreich, E.H. Oldfield, Source and cause of endothelin-1 release Ohlstein, Effect of endothelin on cortical microvascular perfusion in into cerebrospinal fluid after subarachnoid hemorrhage, J. Neuro- rats, Stroke 21 (1990) 451–458.
surg. 87 (1997) 287–293. [55] J.R. Wu-Wong, W.J. Chiou, K.E. Naugles Jr., T.J. Opgenorth, [40] W. Pulsinelli, Pathophysiology of acute ischaemic stroke, Lancet Endothelin receptor antagonists exhibit diminishing potency
follow-339 (1992) 533–536. ing incubation with agonist, Life Sci. 54 (1994) 1727–1734. [41] W. Reith, M. Forsting, J. Weber, R. Stingele, W. Hacke, K. Sartor, [56] M. Yanagisawa, H. Kurihara, S. Kimura, Y. Tomobe, M. Kobayashi,
Laser Doppler flowmetry of focal cerebral ischemia and reperfusion Y. Mitsui, Y. Yasaki, K. Goto, T. Masaki, A novel potent vasocon-in deep bravasocon-in structures vasocon-in rats, Acta Neurochir. (Wien) 131 (1994) strictor peptide produced by vascular endothelial cells, Nature 332
151–156. (1988) 411–415.
[42] M.J. Robinson, I.M. Macrae, M. Todd, J.L. Reid, J. McCulloch, [57] T. Yoshizawa, O. Shinmi, A. Giaid, M. Yanagisawa, S.J. Gibson, S. Reduction in local cerebral blood flow induced by endothelin-1 Kimura, Y. Uchiyama, J.M. Polak, T. Masaki, I. Kanazawa, Endo-applied topically to the middle cerebral artery in the rat, J. thelin: a novel peptide in the posterior pituitary system, Science 247 Cardiovasc Pharmacol. 17 (Suppl. 7) (1991) S354–357. (1990) 462–464.