REPRODUKSI SEL dan PEWARISAN GEN
REPRODUKSI SEL
Sel yang membelah disebut sel induk, dan hasil pembelahannya disebut sel
anak. Sel induk memindahkan salinan informasi genetiknya (DNA) ke sel anak.
Untuk menyampaikan informasi genetik tersebut sel induk harus melipatgandakan
informasi genetik yang dimilikinya (DNA) melalui replikasi (duplikasi) sebelum
melaksanakan pembelahan atau reproduksi sel.
Sel merupakan unit terkecil tempat berlangsungnya proses metabolisme
dan reproduksi. Kedua proses ini dikendalikan oleh bahan genetik, yaitu DNA
yang menyusun keseluruhan gen. Bahan genetic akan mengendalikan
pembentukan enzim yang menjalankan metabolisme,dan mampu bereplikasi
memperbanyak diri dalam proses reproduksi. Dari jenis selnya makhluk selular
dibagi menjadi prokariot dan eukariot. Pada sel eukariot terdapat pembagian
ruang menjadi inti dan sitoplasma; pada inti terdapat bahan genetik (kromosom),
sedangkan pada sitoplasma terdapat organel-organel tempat berlangsungnya
proses metabolisme. Sel prokariot tidak terdapat pembagian sel menjadi inti dan
sitoplasma sehingga material genetik dapat langsung kontak dengan organel.
Tubuh virus tidak memenuhi syarat untuk disebut sel, hanya terdiri dari bahan
genetik yang dibungkus mantel. Oleh karena itu, virus tidak dapat melakukan
metabolisme, kecuali kalau berada di dalam sel inang.
Bakteri bereproduksi melalui pembelahan biner, satu sel membelah
menjadi dua, yang dimulai dengan replikasi kromosom bersamaan dengan
pembesaran sel yang diikuti dengan pembelahan sel. Virus akan bereproduksi di
dalam sel inang melalui dua siklus, siklus litik dan siklus lisogenik. Pada siklus
litik virus setelah menginfeksi inang langsung akan menggandakan bahan
genetiknya yang kemudian disusul dengan pembentukan virus-virus utuh. Pada
siklus lisogenik bahan genetik virus akan berintegrtasi dengan kromosom inang,
dan ikut bereproduksi bersamaan reproduksi sel inang. Siklus reproduksi sel
eukariot mempunyai tahapan G1/ S/ G2/M. Pada tahapan S dilakukan replikasi
kromosom dan pada tahapan M terjadi pembelahan sel, yaitu melalui proses
mitosis atau proses meiosis. Proses mitosis berlangsung terutama pada
sama. Pada mitosis setiap kromosom akan membentuk kromatid bersaudara, yang
selanjutnya kedua kromatid tersebut akan berpisah menjadi dua kromosom, yang
kemudian masing-masing akan bermigrasi ke dua kutub yang berbeda dan
akhirnya menjadi dua sel yang berbeda. Pada meiosis selain masing-masing
kromosom membentuk dua kromatid juga terjadi perpasangan kromosom
homolog. Pada meiosis I akan terjadi pemisahan kromosom homolog, kemudian
pada meiosis II terjadi pemisahan kromatid bersaudara. Pada akhir meiosis akan
terbentuk empat sel dengan jumlah kromosom separuh dari kromosom tetua, dan
antarsel terdapat perbedaan genetik.
Kita mengenal tiga jenis reproduski sel, yaitu Amitosis, Mitosis dan
Meiosis (pembelahan reduksi). Sel terus berkembang dan berkembang biak ,
sel-sel baru menggantikan sel-sel-sel-sel tua yang mati. Reproduksi dimulai dari inti yaitu
mengalami replikasi (duplikasi) DNA dalam kromosom. Kemudian pembelahan
dua pasang DNA antara dua inti yang membelah. Yang terakhir adalah
pembelahan sel membentuk dua anak sel baru ( proses mitosis ). Mitosis adalah
pembelahan sel nukleus setelah terjadinya duplikasi kromosom. Setiap sel anak
mengadakan seperangkat lengkap kromosom yang identik dengan sel induknya (
Diploid=2n ). Jadi mitosis merupakan cara untuk transfer informasi tanpa
mengalami perubahan atau pengurangan dari sel induk ke sel anaknya. Semua
macam reproduksi aseksual dilakukan secara mitosis. Mitosis dapat dijumpai atau
dilihat pada sel-sel embrionik baik pada hewan maupun tumbuhan, namun ada
individu yang cara pembelahannya sederhana yakni pada amoeba yang di kenal
dengan amitosis.
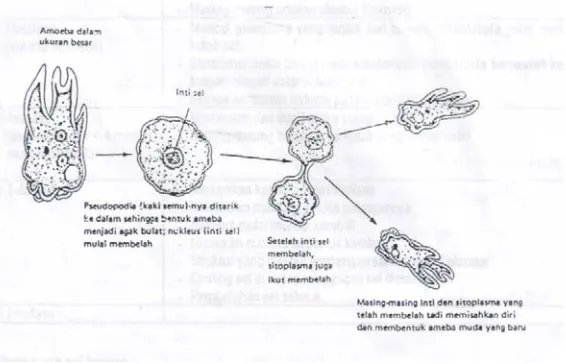
Amitosis ( a = tidak , mitos= benang ) adalah proses pembelahan sel secara
langsung dan sederhana, didahului dengan pembelahan inti tanpa pembentukan
benang-benang kumparan ( kromosom ). Sel membelah diri secara langsung tanpa
melalui tahap-tahap pembelahan sel. Pembelahan cara ini banyak dijumpai pada
Gambar : Pembelahan amitosis pada amoeba
Apabila gen telah mengadakan duplikasi dan tiap kromosom dinamakan
kromatid. Proses mitosis pertama terjadi dalam sitoplasma, terjadi selama bagian
akhir interfase ( periode antara dua mitosis ) pada stuktur kecil yang disebut
dengan sentriole, yang selama interfase tidak aktif sampai sebelum mitosis
berlangsung. Ini disebabkan oleh protein mikrotubulus yang tumbuh antara
masing-masing pasang yang sebenarnya mendorong mereka saling menjauhi.
Pada saat yang sama mikrotubulus tumbuh secara radial menjauhi setiap
pasangan. Sebagian protein mengadakan penetrasi ke inti, sekumpulan
mikrotubulus yang menghubungkan ke dua pasang sentriole dinamakan spindel,
dan seluruh kumpulan mikrotubulus ditambah dua pasangan sentriole dinamakan
aparatus mitosis.
MITOSIS adalah cara reproduksi sel dimana sel membelah melalui tahap-tahap
yang teratur, yaitu Profase-Metafase-Anafase-Telofase. Antara tahap telofase ke
tahap profase berikutnya terdapat masa istirahat sel yang dinarnakan Interfase
(tahap ini tidak termasuk tahap pembelahan sel). Pada tahap interfase, inti sel
melakukan sintesis bahan-bahan inti.
Secara garis besar ciri dari setiap tahap pembelahan pada mitosis adalah sebagai
Tingkat-tingkat pembelahan sel :
Profase ( A, B, C ). Spindel sedang terbentuk , zat krolmatid inti yang
panjang memendek mejadi kromosom nyata.
Promentase ( D ). Pada tingkat pertama selubung inti melarut
mikrotubulus dari apparatus mitosis yang terbetuk melekat pada
kromosom. Pelekatan selalu terjadi pada tempat yang sama pada tiap
kromosom, pada bagian kecil yang disebut sentromer.
Metafase ( E ) . Pasangan sentriole terdorong saling menjauhi perlekatan
mikrotubulus ke pusat sel, berbaris pada bidang ekuator spindel mitosis.
Anafase ( F ).Dengan masih bertumbuhnya spindel, tiap pasang kromosom
saling berpisahan. Mikrotubulus yang menghubungkan satu pasang
sentriole menarik satu kromatid dan mikrotubulus yang menghubungkan
pasangan sentriole lainnya menaeik kromatid ke arah yang berlawanan.
Telofase ( G & H ). Spindel mitosis terus tumbuh lebih panjang, menarik
dua kumpulan kromosom anak terpisah sama sekali. Kemudian apparatus
mitosis melarut dan membetuk membram inti baru disekitar tiap kumpulan
kromosom. Membran ini mungkin terbetuk dari bagian-bagian retikulum
endoplasma yang telah tersedia dalam sitoplasma. Setelah itu sel
membetuk batas pada dua garis tengah antara ke dua inti :
.Reproduksi sel dimulai pada inti : adanya duplikasi DNA (
Deoksiribosa Nukleid acaid ) dalam kromosom.
Kemudian pembelelahan dua pasang DNA : antara dua inti
yang membelah menjadi 2 sel.
Ada pun tahapan-tahapan mitosis :
Fase-fase Aktivitas yang terjadi
1. Profase ( merupakan fase terpanjang )
2. Metafase ( kurang 30 menit ).
¾ Nukleus menghilang, membram inti melebur.
¾ Kromosom timbul menjadi pilihan ( heliks ). Untaian tersebut lebih pendek dan lebih tebal sehingga tampak lebih nyata. Membram nukleus mulai menghilang.
¾ Kromosom membetuk duplikat. Duplikatnya saling melekat pada daerah khusus yang di sebut dengan sentromer atau kinektor.
¾ Masing-masing untaian di sebut Kromatid.
¾ Munculnya gelendong yang terjadi dari sebaris mirotubula yang meluas di antara kutub sel.
¾ Sentromer mulai terikat pada sekelompok mikrotubula berpindah ke suatu titik ditengah-tengah antar kutub.
¾ Semua sentromer terletak pada suatu bidang ekuator.
3. Anafase ( fase terpendek < 4 menit ; mudah di kenali ).
¾ Kromosom dan duplikatnya saling berpisah.
¾ Masing-masing bergerak ke kutub yang berlawanan.
4 Telofase ¾ Merupakan kebalikan dari profase.
¾ Kromosom mulai membuka gulungannya.
¾ Nukleus mulai muncul kembali.
¾ Membram nukleus mulai muncul kembali.
¾ Struktur yang disebut lempengan sel muncul di ekuator.
¾ Dinding sel disetiap lempengan sel disekresikan.
¾ Pembelahan sel selesai.
MEIOSIS (Pembelahan Reduksi) adalah reproduksi sel melalui tahap-tahap
pembelahan seperti pada mitosis, tetapi dalam prosesnya terjadi pengurangan
(reduksi) jumlah kromosom. Meiosis terbagi menjadi dua tahap besar yaitu
Meiosis I dan Meiosis II. Baik meiosis I maupun meiosis II terbagi lagi menjadi
tahap-tahap seperti pada mitosis. Secara lengkap pembagian tahap pada
pembelahan reduksi adalah sebagai berikut:
Berbeda dengan pembelahan mitosis, pada pembelahan meiosis antara telofase I
dengan profase II tidak terdapat fase istirahat (interface). Setelah selesai telofase
II dan akan dilanjutkan ke profase I barulah terdapat fase istirahat atau interface.
Gambar : Pembelahan Meiosis
PERBEDAAN ANTARA MITOSIS DENGAN MEIOSIS
Aspek yang dibedakan Mitosis Meiosis
Tujuan Untuk pertumbuhan Sifat mempertahan-kan diploid
Pada hewan dikenal adanya peristiwa meiosis dalam pembentukan gamet,
yaitu Oogenesis dan Spermatogenesis. Sedangkan pada tumbuhan dikenal
Makrosporogenesis (Megasporogenesis) dan Mikrosporogenesis.
Pada makhluk hidup uniseluler (bersel satu) populasinya dapat
bertambah dalam waktu yang sangat singkat disebabkan juga karena
masing-masing sel terus-menerus membelah. Tapi pembelahan sel yang tidak terkendali
justru membawa suatu dampak yang merugikan makhluk hidup, seperti
munculnya penyakit kanker. Demikian halnya jika pembelahan sel terlalu lambat,
maka pertumbuhan dari makhluk hidup tersebut menjadi lambat sehingga
terbentuk orang-orang kerdil (orang kate)
Daur Hidup dan Penentuan Jenis Seks
Daur hidup eukariot dapat dipenuhi dengan reproduksi vegetatif (siklus
aseksual) atau reproduksi generatif (siklus seksual). Pada reproduksi vegetatif
tidak dilibatkan proses pembentukan gamet dan perkawinan, sedangkan pada
reproduksi generatif terdapat proses pembentukan gamet dan perkawinan. Adanya
pembentukan gamet dan perkawinan menyebabkan adanya fase haploid dan fase
diploid dalam siklus hidup. Makhluk yang sebagian besar dari siklus hidupnya
berada pada fase haploid disebut haplobion, sedangkan yang fase diploidnya yang
lebih panjang disebut diplobion. Cendawan merupakan makhluk haplobion,
sedangkan tumbuhan dan hewan merupakan diplobion. Khamir merupakan
cendawan uniselular, dapat berkembang secara aseksual maupun seksual. Dua sel
dengan tipe perjodohan yang berbeda dapat melakukan perkawinan membentuk
sel diploid, selanjutnya bermeiosis membentuk sel haploid yang selanjutnya dapat
berkembang menjadi cendawan baru. Sel diploid juga dapat bertahan,
berkembang secara vegetatif menghasilkan cendawan diploid. Neurospora crassa
merupakan cendawan multiselular, mempunyai siklus seksual dan siklus aseksual.
Pada siklus aseksual konidium akan berkecambah membentuk hifa, yang
kemudian dapat berkembang menghasilkan konidium. Konidium ini juga dapat
digunakan sebagai sarana untuk perkawinan. Konidium apabila jatuh ke dalam
askogonium dengan tipe perjodohan yang berbeda akan berkembang menjadi sel
induk spora diploid. Melalui proses meiosis akan dihasilkan spora yang tersusun
dan askus.
Askospora ini akan berkecambah menjadi individu baru. Tumbuhan dan
hewan merupakan eukaroit multiselular diplobion. Tumbuhan dewasa diploid
akan menghasilkan bunga yang mengandung anter dan putik.Pada anter terdapat
sel induk mikrospora yang melalui proses meiosis akan menghasilkan polen
haploid. Sedangkan pada putik terdapat sel induk megaspora, yang melalui proses
meiosis akan dihasilkan sel telur haploid. Melalui proses perkawinan akan terjadi
penggabungan inti polen dengan sel telur, menghasilkan zigot diploid. Zigot ini
akan berkembang menjadi biji, yang selanjutnya akan berkembang menjadi
keistimewaan bahwa sebagian terbesar hewan ada pemisahan seks menjadi
individu jantan dan individu betina. Jenis seks organisme diatur dengan berbagai
sistem. Pada bakteri jenis seks diatur dengan adanya plasmid F, yang
membedakan bakteri menjadi F+ dan F-. Adanya pembedaan tersebut
memungkinkan suatu bakteri melakukan konjugasi, setara dengan
perkawinan.Pada cendawan dan tumbuhan jenis seks ditentukan oleh satu atau
beberapa lokus (gen). Perbedaan alel pada lokus-lokus tersebut menyebabkan
organisme menjadi jantan atau betina. Pada hewan jenis seks ditentukan oleh jenis
kromosom seks. Komposisi kromosom seks tertentu menentukan jenis seks
betina, sedangkan komposisi yang lain menjadi jantan. Pada lebah jenis seks
ditentukan oleh perbedaan tingkat ploidi. Jantan berasal dari telur yang tidak
dibuahi (haploid), dan betina berasal dari telur yang dibuahi (diploid). Pada ikan
dan cacing laut tertentu jenis seks ditentukan oleh lingkungan, seperti hormon
yang dikeluarkan induknya.
HUKUM PEWARISAN MENURUT MENDEL
Hukum Segregasi
Sifat organisme dikendalikan oleh gen yang dapat diwariskan dari satu generasi
ke generasi berikutnya. Setiap sifat dikendalikan oleh sepasang alel yang terdapat
pada satu lokus dari suatu kromosom. Antara dua alel pada satu lokus mungkin
mempunyai hubungan dominan-resesif atau kodominan. Pada persilangan antara
dua tetua homozigot yang berbeda akan diperoleh F1 yang bersifat heterozigot.
Dalam kasus alel dominan-resesif, fenotipe F1 akan sama dengan fenotipe tetua
dominan, tetapi dalam kasus alel kodominan genotipe F1 yang berbeda dari kedua
genotipe tetuanya akan menghasilkan fenotipe yang berbeda pula. Persilangan
sendiri antar F1 akan menghasilkan generasi F2. Pada percobaan monohibrid atau
persilangan dengan pembeda satu sifat atau satu lokus, akan diperoleh nisbah
genotipe AA : Aa : aa sama dengan 1:2:1, bila F1nya bergenotipe Aa atau
tetua-tetua awalnya AA dan aa. Dalam kasus alel dominan-resesif dari nisbah genotipe
tersebut akan dihasilkan nisbah fenotipe 3:1 untuk dominan (A-) : resesif (aa),
mewakili fenotipe tetua-1 (homozigot), F1 (heterozigot), tetua-2 (homozigot)
dengan nisbah sama dengan nisbah genotipe. Dari data F2 monohibrid, Mendel
menyusun Hukum Segregasi yang bermakna bahwa pasangan alel yang
bergabung melalui perkawinan akan bersegregasi dengan bebas dalam proses
pembentukan gamet. Kebebasan ini ditunjukkan oleh nisbah yang sama antara
gamet F1 beralel A dengan yang beralel a atau 1/2 A dan 1/2 a sehingga melalui
proses penggabungan gamet secara acak dalam pembentukan populasi F2 akan
diperoleh genotipe AA, Aa, dan aa dengan perbandingan AA, 1/2Aa, dan 1/4 aa.
Hukum Perpaduan Bebas
Berdasarkan data F2 dihibrid, Mendel menyusun Hukum Perpaduan Bebas yang
berisi bahwa segregasi alel-alel pada suatu lokus, bebas dari pengaruh segregasi
alel-alel lokus lainnya. Dari F1 bergenotipe AaBb dalam proses pembentukan
gamet alel A dapat bebas berpadu dengan B atau b, juga a bebas memilih B atau
b. Akibat perpaduan bebas ini maka setiap jenis gamet yang terbentuk, yaitu AB,
Ab, aB, dan ab akan mempunyai frekuensi yang sama. Dalam kasus dihibrid akan
mempunyai frekuensi masing-masing 0,25. Akibat perpaduan bebas dari alel-alel
dalam pembentukan gamet, dan penggabungan bebas gametgamet dalam
perkawinan maka dalam kasus alel dominan-resesif, F2 akan mempunyai fenotipe
dengan perbandingan 9:3:3:1. Untuk membuktikan Hukum Perpaduan Bebas
dilakukan uji silang dihibrid dengan menyilangkan F1 terhadap tetua resesif.
Terbukti kebenaran Hukum ini dengan munculnya turunan uji silang dengan
perbandingan 1:1:1:1 untuk fenotipe yang menggambarkan gamet AB, Ab, aB,
dan ab. Kebenaran Hukum Mendel diperkuat oleh hasil pengamatan kromosom
yang memperlihatkan kesetaraan Hukum Mendel dengan perilaku kromosom
dalam proses meiosis. Kesetaraan tersebut terlihat sebagai berikut: kesetaraan
perpasangan alel dengan perpasangan kromosom homolog, kesetaraan segregasi
sepasang alel dengan perpisahan pasangan kromosom homolog menuju kutub
yang berbeda, dan kesetaraan antara perpaduan bebas alel-alel dengan kebebasan
berbagai kromosom dalam memilih kutub yang dituju dalam meiosis.
Dengan kemajuan penemuan genetika terlihat adanya berbagai fenomena yang
menyimpang dari kaidah Mendel. Penyebab pertama dari penyimpangan tersebut
karena adanya interaksi alel-alel antarlokus dalam ekspresi pembentukan fenotipe.
Interaksi seperti ini secara umum disebut epistasis, yang terbagi atas interaksi
komplementasi, modifikasi dan duplikasi. Penyebab kedua adalah terjadinya
pautan antarlokus atau segregasi alel-alel suatu lokus yang dipengaruhi oleh
segregasi pada lokus yang lain pada kromosom yang sama. Dan yang terakhir
disebabkan oleh gen yang bersangkutan terletak pada organel di luar inti
(ekstrakromosom). Migrasi organel pada waktu pembelahan sel berbeda dengan
proses migrasi kromosom pada saat meiosis sehingga pewarisannya tidak
mengikuti kaidah Mendel.
PEMETAAN KROMOSOM
Peta Genetika Ekuariot Diploid
Penentuan posisi gen pada kromosom dilakukan dengan melihat pada kromosom
mana gen-gen tersebut terletak, dan bagaimana jaraknya satu dengan yang lain.
Pengelompokan gen pada kromosom dilihat dengan keterpautan antara satu lokus
dengan yang lainnya. Bila dua lokus berpautan maka disimpulkan bahwa kedua
lokus tersebut terletak pada satu kromosom. Bila lokus tersebut bersegregasi
bebas kemungkinan lokus-lokus tersebut terpisah pada kromosom yang berbeda.
Pada satu kromosom jarak antara satu lokus dengan lokus yang lainnya diukur
dengan menghitung persentase rekombinasi, yaitu banyaknya gamet tipe
rekombinan dibandingkan dengan total gamet. Bila rekombinan bernilai lebih
besar atau sama dengan 0.5 maka kedua lokus dinyatakan bebas, dan lebih kecil
dari 0.5 maka dinyatakan berpautan. Semakin besar jarak antara dua lokus maka
akan semakin besar frekuensi pindah silang dan akan semakin meningkatkan
persentase gamet rekombinan.
Pemetaan Genetik Eukariot Haploid
Cendawan merupakan makhluk haplobion sehingga analisis genetic dapat
langsung dilakukan terhadap spora hasil meiosis. Spora yang terdapat dalam askus
askus ada yang teratur (disebut tetrad teratur) dan ada pula yang tidak teratur
(tetrad tidak teratur). Pada cendawan dengan tetrad teratur, susunan spora pada
askus dapat menggambarkan proses meiosis yang terjadi. Apabila tidak terjadi
pindah silang akan dihasilkan askus tipe DP, dan akan muncul tipe T bila terjadi
pindah silang. Dalam kasus analisis untuk dua lokus akan muncul tipe DR yang
merupakan hasil pindah silang dua pasang kromatid atau akibat perpaduan bebas.
Jarak satu lokus terhadap sentromernya dihitung dengan rumus r = (0.5T/Total)
100%; dan jarak antara dua lokus dihitung dengan rumus r = {(DR + 0.5T)/Total}
100%. Dalam kasus cendawan dengan tetrad tidak teratur jarak antarlokus
dihitung dari persentase spora rekombinan terhadap total spora. Jarak dihitung
berdasarkan persamaan r = (n-spora rekombinan/ Total spora) 100%.
Peta Sitologis
Peta sitologi merupakan peta fisik berdasarkan pengamatan pada kromosom di
bawah mikroskop. Untuk memperoleh gambar kromosom dilakukan pengambilan
sampel dari jaringan dengan sel yang sedang aktif membelah. Gambar kromosom
diperoleh dari pengamatan kromosom saat metafase, kemudian kromosom
tersebut diatur berdasarkan tipenya. Gambar seperti ini disebut kariotipe. Dalam
analisis akan dihitung jumlah kromosom tiap sel, kemudian dari masing-masing
kromosom dipelajari morfologinya, seperti letak sentromer, keberadaan satelit,
dan pita kromatin. Pita-pita menunjukkan kondensasi DNA, yang dapat diartikan
sebagai posisi gen. Hubungan pita dengan gen dipelajari dengan melihat
keberadaan atau kehilangan pita (delesi) dengan Untuk memperdalam pemahaman
Anda mengenai materi di atas, kerjakanlah latihan berikut ! kehilangan sifat
tertentu dari organisme. Keparalelan antara peta sitologi dengan peta genetik
dilakukan dengan membandingkan urutan-urutan gen yang dihasilkan oleh kedua
pita tersebut.
STRUKTUR DAN REPLIKASI BAHAN GENETIK
Melalui kejadian transformasi genetik pada bakteri telah dibuktikan bahwa DNA
merupakan unsur genetik atau penentu sifat makhluk hidup. DNA merupakan
sedangkan nukleotida itu sendiri disusun oleh tiga komponen, yaitu gula
(deoksiribonukleotida), fosfat, dan basa. Ada empat jenis basa pada DNA, yaitu
Sitosin, Guanin, Adenin, dan Timin. Di samping DNA ada asam nukleat lain,
yaitu RNA yang dibedakan dari DNA oleh gula dan basanya. Pada RNA terdapat
Ribosa sebagai pengganti Deoksiribosa, dan Urasil sebagai pengganti Timin pada
DNA. Nukleotida yang satu dirangkaikan dengan nukleotida lain oleh ikatan 3′ –
5′ fosfodiester membentuk suatu rantai polinukleotida. Dalam satu molekul DNA
terdapat dua utas polinukleotida yang diikat oleh ikatan hidrogen yang terbentuk
antara basa-basanya. Perpasangan A-T diikat oleh dua ikatan hidrogen dan
perpasangan G-C oleh tiga ikatan hidrogen. Pasangan kedua utasan tersebut
berpilin membentuk heliks ganda; pada satu pilinan terdapat 10 pasang basa
dengan jarak antar nukleotida sebesar 3.4 A0 atau satu pilinan mempunyai
panjang sekitar 34 A0.
Pilinan heliks ganda mempunyai garis tengah 20 A0. Struktur heliks ganda
dengan gula fosfat yang terletak di bagian luar heliks dan basa di bagian dalam
heliks, menjamin kestabilan struktur DNA. Kestabilan ini meliputi, pertama
ketahanan DNA terhadap kerusakan akibat keadaan lingkungannya, dan yang
kedua kestabilan genetik yang berhubungan dengan ketepatan dalam proses
replikasi. Struktur heliks-ganda memungkinkan adanya replikasi semikonservatif
di dalam sel DNA berasosiasi dengan protein, seperti histon pada eukariot.
Besarnya DNA yang menyusun genom beragam dari satu organisme ke organisme
lain. Genom virus berkisar dari 3 103 sampai 105 pasang basa, bakteri dari 5 105
sampai 107 pasang basa, eukariot dari 107 sampai 1011 pasang basa. Genom
bakteri terdiri dari kromosom dan plasmid; dan sebagian besar bakteri mempunyai
satu kromosom. Genom eukariot terdiri dari gen inti yang disusun oleh sejumlah
kromosom, dan gen sitoplasma yang terdapat dalam bentuk DNA mitokondria dan
DNA kloroplas.
Replikasi DNA
Replikasi DNA mengikuti pola semikonservatif yang melibatkan sejumlah enzim
dan protein di dalamnya. Heliks ganda akan diudar menjadi utas tunggal dengan
bantuan helikase, girase, dan protein SSB. Utas tunggal yang terbentuk akan
membentuk percabangan replikasi dan akan digunakan sebagai utas model
cetakan. Karena pertumbuhan sintesis DNA berjalan dengan arah 5-3 maka pada
utas leading (berujung 3′OH) sintesis akan bergerak dari ujung ke arah pangkal
percabangan replikasi. Sebaliknya pada utas lagging (berujung 5′P), sintesis
berjalan dari pangkal ke ujung percabangan. Sintesis ini berjalan secara bertahap
dalam bentuk fragmen Okazaki. Polimerase DNA mempunyai tingkat ketepatan
yang tinggi karena dilengkapi dengan perangkat baca ulang. Dengan situs
eksonuklease 3-5 nukleotida yang salah akan dibuang dan diganti dengan yang
seharusnya. Dalam replikasi kromosomnya, bakteri menggunakan replikasi model
q yang dimulai dari satu titik Ori, yaitu Ori-C. Replikasi kromosom eukariot
dimulai dari banyak titik Ori, dan berjalan dwiarah. Virus menggunakan cara
EKSPRESI GEN
Transkripsi: Informasi dari Gen ke RNA
Gen diekspresikan melalui peranan dalam pengendalian sifat-sifat organisme.
Peran ini dijalankan melalui pengendalian proses pembentukan protein dan
enzim-enzim yang mengkatalisis reaksi kimia pada berbagai tahapan
metabolisme. Gen diekspresikan melalui dua tahapan, yaitu transkripsi dan
translasi. Transkripsi adalah penyalinan informasi dari gen ke dalam molekul
RNA, yang dalam pelaksanaannya adalah berupa penyusunan basa-basa pada
rantai RNA dengan menggunakan runtunan basa DNA gen sebagai modelnya.
Translasi adalah penterjemahan informasi berupa runtunan basa RNA menjadi
rangkaian asam-amino pada protein. Dalam transkripsi terdapat dua perangkat,
yaitu ruas DNA yang menjadi model dan enzim transkriptase yang mengkatalisis
proses sintesis RNA. Ruas DNA yang dijadikan model juga disebut ruas penyandi
ialah ruas yang dibatasi oleh promotor dan terminator. Promotor ialah segmen
DNA yang berfungsi sebagai tanda bagi enzim transkriptase untuk mengawali
proses transkripsi atau penyalinan basa DNA menjadi basa RNA. Terminator
ialah segmen DNA yang menjadi tanda untuk berakhirnya proses transkripsi.
Panjang ruas antara promoter dan terminator (ruas penyandi) akan sama dengan
panjang RNA yang dihasilkan. Terdapat empat jenis RNA hasil transkripsi, yaitu
mRNA, tRNA, rRNA, dan snRNA. Tiga RNA selain snRNA berperanan dalam
menyusun runtunan asam-amino rantai polipeptida atau protein. tRNA berperan
sebagai pengangkut asam-amino dan penterjemah rangkaian kodon-kodon yang
terdapat pada mRNA menjadi rangkaian asam-amino. rRNA berfungsi sebagai
rangka ribosom dan mengenali tRNA dan mRNA.
Protein dan Sandi Genetik
Protein merupakan makromolekul dengan asam-amino sebagai molekul dasarnya.
Semua asam-amino mempunyai rumus bangun molekul yang sama, kecuali
prolin, terdiri dari gugus R, Ca, gugus – NH2, dan gugus -COOH. Perbedaan satu
asam-amino dengan yang lainnya terletak pada gugus R-nya. Dikenal 20 jenis
asam-amino yang terlibat pada tahap translasi. Asam-amino dirangkaikan satu
dengan yang lain membentuk rantai polipeptida, yang berbentuk liniear. Struktur
polipeptida yang linear merupakan struktur primer protein, yang kemudian akan
berkembang menjadi struktur sekunder berkat terbentuknya ikatan hidrogen antara
asam-amino pada jarak tertentu. Terdapat dua jenis struktur sekunder yaitu
heliks-a dheliks-an lembheliks-arheliks-an-b. Pheliks-adheliks-a theliks-ahheliks-ap heliks-akhir heliks-akheliks-an terjheliks-adi lheliks-agi pelipheliks-atheliks-an dheliks-ari struktur
struktur tersier sudah terdapat situs-situs fungsional protein. Untuk protein
monomer (disusun oleh satu polipeptida) struktur tersier merupakan bentuk akhir
protein yang menentukan fungsinya, sedangkan pada protein oligomer (disusun
beberapa polipeptida) bentuk akhirnya adalah struktur kuartener, yang merupakan
gabungan dari struktur tersier. Struktur akhir protein menentukan fungsinya,
perubahan struktur dapat menyebabkan protein kehilangan fungsinya.
Berdasarkan fungsinya protein terbagi menjadi enzim (berperan sebagai
katalisator), protein cadangan, protein sistem pengangkut, protein sistem
kekebalan, hormon, dan protein struktur. Hubungan antara protein dengan gen
diatur melalui sandi genetik, yaitu suatu aturan yang menghubungkan antara suatu
kodon pada mRNA dengan asam-amino pada polipeptida. Satu kodon akan
menyandikan satu jenis asam-amino, sehingga dengan begitu satu gen akan
menyandikan satu jenis polipeptida. Perubahan pada kodon akan menyebabkan
terjadinya perubahan pada protein.
Translasi: RNA menjadi Protein
Translasi adalah proses penterjemahan informasi genetik berupa rangkaian kodon
mRNA menjadi rangkaian asam-amino polipeptida. Ada tiga unsur yang terlibat
proses translasi tersebut, yaitu mRNA, tRNA, dan rRNA. Unsur pertama, mRNA
berperan sebagai model untuk menyusun runtunan asam-amino polipeptida. Pada
mRNA terdapat ruas penyandi, yaitu bagian yang dibatasi oleh kodon awal dan
kodon akhir yang akan menjadi model untuk penyusunan protein. Unsur kedua,
tRNA berperan menterjemahkan kodon-kodon mRNA menjadi asamamino
polipeptida dan mengangkut asam-amino ke kompleks translasi. Kemampuan
menterjemahkan dipunyai tRNA berkat adanya simpul antikodon, sedangkan
kemampuan sebagai pengangkut karena ada ujung penerima asam-amino sehingga
tRNA dapat berasosiasi dengan asam amino membentuk aminoasil-tRNA.
Ribosom berperan sebagai tempat pertemuan mRNA dengan tRNA serta
penterjemahan kodon serta reaksi perangkaian asam-amino. Pada ribosom
terdapat berbagai situs yang berfungsi untuk mendukung peran di atas; situs
tersebut ialah situs mRNA, situs P dan situs A untuk tRNA, serta situs
perpanjangan polipeptida, dan proses akhir. Inisiasi dimulai dengan subunit
ribosom kecil mengenali mRNA berkat kemampuan rRNA16S berpasangan
dengan ruas Shine Dalgarno di hulu kodon awal, selanjutnya tRNA inisiator akan
menempel pada kodon awal yang terdapat pada mRNA, terakhir subunit ribosom
besar akan bergabung menghasilkan ribosom sempurna.
Setelah insisasi pada ribosom akan menempel mRNA dan
aminoasil-tRNA-inisiator pada situs P yang juga tepat pada posisi kodon awal, kemudian ke situs A
akan menempel satu aminoasil-tRNA yang cocok dengan kodon yang terdapat
pada situs tersebut. Setelah ada dua tRNA pada ribosom akan terjadi reaksi
transpeptidasi yang memindahkan asam-amino dari situs P merangkai dengan
asam amino pada situs A, membentuk aminoasil-tRNA dan terjadi penambahan
satu asam-amino pada rantai polipeptida. Selanjutnya terjadi pergeseran ribosom
satu kodon ke arah hilir mRNA yang menyebabkan peptidiltRNA dari situs A
pindah ke situs P dan situs A kosong dan siap menerima aminoasil-tRNA
berikutnya. Proses akan terus berulang sampai ribosom menemukan kodon akhir.
Ketika ribosom mencapai kodon akhir tidak akan ada aminoasiltRNA yang masuk
ke situs A dan muncul protein FR yang akan memisahkan ribosom, tRNA, mRNA
dan polipeptida. Subunit ribosom kecil akan terpisah dari subunit besar, mRNA
akan terurai menjadi nukleotida bebas, dan polipeptida siap masuk ke proses
berikutnya untuk membentuk protein yang berfungsi. Untuk dapat mencapai
struktur akhir protein yang fungsional polipeptida hasil translasi kadang-kadang
harus melewati suatu proses modifikasi pascatranslasi, yang dapat berupa
pemotongan rantai polipeptida, perubahan asam-amino tertentu atau penambahan
senyawasenyawa tertentu. Berbagai protein hasil translasi menurut fungsinya,
yaitu enzim, hormon, protein pengangkut, protein toksin, antibodi, protein
penyimpan dan cadangan, protein kontraksi, serta protein penyangga struktur.
Mutasi Tingkat Gen
Mutasi ialah perubahan struktur DNA kromosom atau gen-gen, yang
menyebabkan perubahan pada tingkat ekspresi. Perubahan pada struktur gen akan
protein. Oleh karena itu, mutasi akan menyebabkan terjadinya perubahan proses
metabolisme, yang dikatalisis oleh enzimenzim yang disandikan oleh gen-gen
yang bermutasi tersebut. Perubahan struktur dan ekspresi gen serta perubahan
metabolisme tersebut dapat terlihat pada perubahan morfologi, perubahan kimia
atau perubahan pertumbuhan dan daya adaptasi. Ukuran perubahan struktur dapat
dilihat dari banyaknya basa yang berubah; apabila hanya satu basa yang berubah
maka disebut mutasi titik, dan apabila sejumlah basa yang berdampingan yang
berubah disebut mutasi basa ganda. Mutasi basa ganda dapat mengenai satu gen
atau melebihi satu gen. Mutasi yang hanya mengenai satu gen disebut mutasi gen,
dan apabila mengenai lebih dari satu gen biasa disebut mutasi kromosom. Mutasi
titik terjadi akibat adanya pergantian (substitusi) suatu pasangan basa pada DNA
dengan pasangan basa lainnya atau akibat terjadinya penyisipan atau pengurangan
satu pasang basa. Substitusi pasangan basa dapat berupa transisi (purin diganti
oleh purin atau pirimidin diganti oleh pirimidin) atau transversi (purin diganti oleh
pirimidin atau pirimidin diganti oleh purin). Pasangan TA melalui transisi akan
diganti oleh CG, dan melalui transversi diganti oleh AT dan GC. Akibat
perubahan satu basa ini maka akan terjadi perubahan satu kodon pada mRNA dan
perubahan satu asam-amino pada rantai polipeptida. Ada tiga kemungkinan jenis
kodon baru yang muncul, kodon sinonim (disebut mutasi bisu), kodon lain bukan
sinonim (mutasi ubah arah), dan kodon akhir (mutasi hilang arah). Penambahan
atau pengurangan basa DNA akan menyebabkan perubahan sejumlah kodon yang
terdapat di belakang tempat penyisipan atau pengurangan tersebut.
Perubahan kodon tersebut dapat juga memunculkan kodon akhir di posisi baru.
Oleh karena itu, mutasi akibat penambahan atau pengurangan akan menyebabkan
terjadinya perubahan sejumlah asam-amino dibelakang titik mutasi serta
mengubah ukuran panjang rantai polipeptida. Mutasi ini dapat terjadi secara
spontan atau akibat adanya rangsangan dari luar. Mutasi spontan terjadi karena
adanya fenomena kimia tautomerik, yaitu perubahan konfigurasi molekul akibat
perubahan kondisi larutan. Perubahan tautomerik menyebabkan Adenin menjadi
Aimino, Sitosin menjadi Cimino, Guanin menjadi Genol, dan Timin menjadi
Tenol. Dalam proses replikasi turunan-turunan basa tersebut dapat membentuk
Sejumlah senyawa kimia (seperti hidroksilamin, EES, EMS, akridin) dapat
merangsang terjadinya perubahan basa DNA. Perubahan basa ini dapat
menyebabkan terjadi perpasangan basa atau basa tersebut dilewati dalam replikasi
atau basa tersebut dibuang dari DNA. Sinar dengan gelombang tertentu dapat
merupakan bahan yang merangsang terjadinya perubahan struktur DNA dan
memunculkan mutasi.
Mutasi Tingkat Kromosom
Mutasi kromosom ialah mutasi basa ganda yang lingkupannya melebihi rentang
suatu gen. Mutasi kromosom terbagi menjadi perubahan struktur kromosom dan
perubahan jumlah kromosom. Perubahan struktur kromosom terdiri dari duplikasi
(pertambahan atau penggandaan segmen kromosom tertentu), delesi (kehilangan
segmen kromosom), inversi (perubahan arah segmen kromosom), dan translokasi
(perpindahan posisi segmen baik dalam satu kromosom maupun antarkromosom).
Rekombinasi merupakan kekuatan utama terjadinya perubahan struktur
kromosom, yaitu akibat terjadinya penyimpangan dari proses rekombinasi
homolog. Rekombinasi antara pasangan kromosom homolog pada situs yang
bukan situs aslinya dapat memunculkan duplikasi dan delesi. Rekombinasi
antardua situs yang terdapat dalam satu kromosom yang sama dapat
memunculkan delesi dan inversi. Rekombinasi nirhomolog dapat menyebabkan
terjadinya translokasi intrakromosom. Rekombinasi antara dua kromosom
heterolog dapat memunculkan translokasi antar kromosom. Perubahan jumlah
kromosom terdiri dari perubahan aneuploid (penambahan kromosom tertentu) dan
perubahan euploid (penambahan dengan semua kromosom satu ploidi).
Terjadinya penyimpangan dalam proses mitosis dapat menyebabkan terjadinya
perubahan jumlah kromosom. Perubahan euploid terbagi menjadi alopoliploid
(penambahan dengan ploidi yang berbeda) dan autopolyploid (penambahan
REGULASI EKSPRESI GEN
Regulasi Ekspresi Gen pada Mikrobia
Ekspresi suatu gen dapat diatur, “dinyalakan atau dimatikan” melalui suatu sistem
yang disebut ekspresi gen. Pada makhluk bersel tunggal terdapat berbagai tipe
atau tingkat regulasi, antara lain regulasi pada tingkat inisiasi transkripsi (operon
laktosa), regulasi pada tingkat RNA translasi (operon triptofan), regulasi pada
tingkat struktur promoter (pada pergantian protein permukaan flagela S.
typhimurium) dan regulasi pada tingkat struktur gen (pada perubahan tipe
perjodohan Saccharomyces cereviseae). Pada operon laktosa terdapat tiga gen
struktural yang salah satunya ialah lak-Z yang menyandikan enzim
b-galaktosidase, yang berperan mengkatalisis penguraian laktosa menjadi glukosa
dan galaktosa. Enzim b-galaktosidase akan diekspresikan apabila tersedia laktosa
dan tidak ada glukosa, dan ekspresinya harus dihentikan bila glukosa cukup
tersedia atau tidak tersedia laktosa. Ekspresi operon laktosa diatur dengan dua
cara yaitu dengan protein represor yang berinteraksi dengan laktosa, atau dengan
protein aktivator yang berinteraksi dengan glukosa. Gen regulator lak-i akan
menyandikan protein represor, yang bila tidak ada laktosa akan mencegah
transkriptase melakukan inisiasi transkripsi. Bila ada laktosa maka senyawa ini
akan mengubah konfigurasi protein represor sehingga tidak dapat menghalangi
transkriptase menginisiasi transkripsi. Protein aktivator CAP yang disandikan oleh
gen regulator crp berfungsi mengaktifkan transkriptase. Protein CAP dapat
berperan sebagai aktivator bila berasosiasi dengan cAMP, yang ketersediaannya
dalam sel ditentukan oleh keberadaan glukosa.
Apabila kuantitas glukosa tinggi, cAMP rendah maka CAP tidak dapat
mengaktifkan transkriptase, dan sebaliknya bila glukosa rendah, cAMP tinggi
maka CAP dapat mengaktifkan transkriptase. Operon triptofan keberlangsungan
ekspresinya diatur pada tingkat RNA yang ditranskripsikan; dan pengaturan
tersebut dilakukan oleh triptofan hasil metabolisme. Pada operon triptofan
terdapat lima gen struktural dan satu gen regulator (trp-L); dengan trp-L berada
tepat di belakang promotor. Inisiasi transkripsi akan berjalan dengan lancar, dan
enzim transkriptase berada pada ruas trp-L, yaitu dengan menentukan apakah
transkripsi akan berhenti pada trp-L atau dilanjutkan ke ruas gen yang ada di