TESIS
KERAGAMAN DAN DINAMIKA POPULASI LALAT BUAH (DIPTERA: TEPHRITIDAE) YANG MENYERANG
TANAMAN BUAH-BUAHAN DI BALI
NI KADEK NITA KARLINA ASTRIYANI
PROGRAM PASCASARJANA UNIVERSITAS UDAYANA
DENPASAR
2014
i
TESIS
KERAGAMAN DAN DINAMIKA POPULASI LALAT BUAH (DIPTERA: TEPHRITIDAE) YANG MENYERANG
TANAMAN BUAH-BUAHAN DI BALI
NI KADEK NITA KARLINA ASTRIYANI NIM 1290861001
PROGRAM MAGISTER
PROGRAM STUDI BIOTEKNOLOGI PERTANIAN PROGRAM PASCASARJANA
UNIVERSITAS UDAYANA DENPASAR
2014
ii
KERAGAMAN DAN DINAMIKA POPULASI LALAT BUAH (DIPTERA: TEPHRITIDAE) YANG MENYERANG
TANAMAN BUAH-BUAHAN DI BALI
Tesis untuk Memperoleh Gelar Magister
Pada Program Magister, Program Studi Bioteknologi Pertanian, Program Pascasarjana Universitas Udayana
NI KADEK NITA KARLINA ASTRIYANI NIM 1290861001
PROGRAM MAGISTER
PROGRAM STUDI BIOTEKNOLOGI PERTANIAN PROGRAM PASCASARJANA
UNIVERSITAS UDAYANA DENPASAR
2014
iii
Lembar Pengesahan
TESIS INI TELAH DISETUJUI PADA TANGGAL : 16 Juni 2014
Pembimbing I, Pembimbing II,
Prof. Dr. Ir. I Wayan Supartha, MS. I Putu Sudiarta, SP.,MSi.,Ph.D.
NIP. 19570330 198601 1 001 NIP. 19791107 200501 1 002
Mengetahui
Ketua Direktur
Program Studi Bioteknologi Pertanian Program Pascasarjana Program Pascasarjana Universitas Udayana Universitas Udayana
Prof. Dr. Ir. I Gede Rai Maya Temaja, MP. Prof. Dr. dr. A.A. Raka Sudewi, Sp. S (K) NIP. 19621009 198803 1 002 NIP. 19590215 198510 2 001
iv
PENETAPAN PANITIA PENGUJI
Tesis ini Telah Diuji Pada Tanggal 16 Juni 2014
Panitia Penguji Tesis Berdasarkan SK Rektor
Universitas Udayana, No. 1849/UN14.4/HK/2014, Tanggal 20 Juni 2014
Ketua : Prof. Dr. Ir. I Wayan Supartha, M.S.
Anggota :
1. I Putu Sudiarta , S.P.,M.Si.,Ph.D 2. Prof. Ir. I Wayan Susila, M.S 3. Prof. Dr. Ir. I Nyoman Wijaya, M.S 4. Dr. G.N. Alit Susanta Wirya, S.P., M.Agr
v
SURAT PERNYATAAN BEBAS PLAGIAT
Saya yang bertandatangan di bawah ini :
Nama : Ni Kadek Nita Karlina Astriyani
NIM : 1290861001
Program Studi : Bioteknologi Pertanian
Judul Tesis : Keragaman dan Dinamika Populasi Lalat Buah (Diptera:
Tephritidae) yang Menyerang Tanaman Buah-Buahan di Bali.
Dengan ini menyatakan bahwa karya ilmiah tesis ini bebas plagiat. Apabila dikemudian hari terbukti plagiat dalam karya ilmiah ini, maka saya bersedia menerima sanksi peraturan Mendiknas RI No.17 Tahun 2010 dan Peraturan Perundang-Undangan yang berlaku.
Denpasar, 20 Juni 2014 Yang membuat pernyataan
(Ni Kadek Nita Karlina Astriyani)
vi
UCAPAN TERIMA KASIH
Pertama-tama perkenankanlah penulis memanjatkan puji syukur ke hadapan Ida Sang Hyang Widhi Wasa, karena hanya atas asung wara nugraha- Nya, tesis ini dapat diselesaikan.
Pada kesempatan ini perkenankanlah penulis mengucapkan terima kasih yang sebesar-besarnya kepada Prof. Dr. Ir. I Wayan Supartha, MS. selaku Pembimbing I yang dengan penuh perhatian dan kesabaran memberikan dorongan, semangat, bimbingan, saran dan pengarahan kepada penulis selama penyusunan tesis ini;, I Putu Sudiarta,S.P.,M.Si.,Ph.D selaku Pembimbing II yang dengan penuh perhatian dan kesabaran telah memberikan bimbingan dan saran kepada penulis sekaligus sebagai pembimbing akademik yang dengan sabar menjadi pembimbing akademik penulis selama menjadi mahasiswa pada Program Pascasarjana Universitas Udayana.
Ucapan yang sama juga ditujukan kepada Rektor Universitas Udayana Prof. Dr. dr. Ketut Suastika, Sp.PD., KEMD, Ibu Direktur Pascasarjana Universitas Udayana Prof. Dr. dr. A.A. Raka Sudewi, Sp. S(K) dan Prof. Dr. Ir. I Gede Rai Maya Temaja, M.P selaku Ketua Program Studi Bioteknologi Pertanian Program Pascasarjana Universitas Udayana atas kesempatan yang diberikan kepada penulis untuk menjadi mahasiswa Program Magister pada Program Pascasarjana Universitas Udayana serta seluruh staf dosen dan staf administrasi yang telah banyak membantu penulis selama menempuh pendidikan. Pada kesempatan ini penulis juga mengucapkan terima kasih yang sebesar-besarnya kepada Bapak dan Ibu pegawai Balai Karantina Denpasar Kelas 1 atas informasi,
vii
kerjasama dan kesempatan untuk belajar dan kemudahan dalam penggunaan Laboratorium Entomologi selama penulis melakukan penelitian. Ucapan terima kasih penulis sampaikan pula kepada para penguji tesis Prof. Ir. I Wayan Susila, M.S, Prof. Dr. Ir. I Nyoman Wijaya, M.S dan Dr. G.N. Alit Susanta Wirya, S.P., M.Agr yang telah dengan sabar memberikan masukan, saran dan koreksi sehingga tesis ini dapat terwujud menjadi lebih baik.
Ucapan terima kasih yang tulus penulis sampaikan kepada orangtua penulis yaitu Ir. I Nengah Widiada, dan Ni Luh Ami, kakak penulis Ni Putu Widyami Yanthi, SE, Adik-adik I Nyoman Bagus Kamayana dan Ni Ketut Santhi Sannidhi, begitu juga terimakasih kepada Kadek Cahyadi Putra, S.Pd yang selalu memberikan semangat kepada penulis, serta seluruh keluarga yang tidak dapat penulis sebutkan satu persatu atas dukungan, semangat dan doa selama penulis menyelesaikan pendidikan terutama penyusunan tesis ini. Akhirnya ucapan terima kasih penulis sampaikan juga kepada teman-teman Agroekoteknologi ’08 (Mika, Ayu, Ayu Rahma, dan Gek Surya), Biotek ’12 (Ocha, Dewa, Rian, Adi Candra, Bli Dika, Wira dan Agus), Biotek’ 13 (Putri,Ogink, Kayan dan Rahde) serta semua teman-teman yang tidak dapat penulis sebutkan satu persatu atas doa, kerjasama dan loyalitasnya selama ini.
Semoga Ida Sang Hyang Widhi Wasa selalu melimpahkan rahmat-Nya kepada semua pihak yang telah membantu pelaksanaan dan penyelesaian tesis ini.
Semoga tesis ini dapat memberikan sumbangan bagi pengembangan ilmu pengetahuan dan bermanfaat bagi semua.
Denpasar, Juni 2014 Penulis
viii ABSTRAK
Keragaman dan Dinamika Populasi Lalat Buah (Diptera: Tephritidae) yang Menyerang Tanaman Buah-Buahan di Bali
Lalat buah (Diptera: Tephritidae) merupakan hama yang memiliki arti penting bagi pertanian. Terdapat sekitar 4000 spesies lalat buah di dunia dan 35%
di antaranya merupakan hama penting pada buah-buahan termasuk di dalamnya buah-buahan komersial yang mempunyai nilai ekonomi tinggi. Informasi tentang keberadaan jenis-jenis lalat buah yang ada di suatu daerah perlu diketahui dan dilaporkan sebagai langkah antisipasi dan pengendalian pada tanaman buah yang dibudidayakan terutama di Bali. Tujuan penelitian ini adalah untuk mengetahui keragaman, indeks keragaman dan indeks kesamaan, spesies lalat buah yang dominan di pasar dan di sentra buah-buahan, hubungan kelimpahan populasi dengan persentase serangan serta keragaman dan tingkat parasitisasi parasitoid yang berasosiasi dengan masing-masing spesies lalat buah di lapangan.
Penelitian dilakukan di Lapangan dan di Laboratorium. Penelitian di Laboratorium dilaksanakan di Laboratorium Pengendalian Hama dan Penyakit Terpadu Tanaman Program Studi Agroekoteknologi Fakultas Pertanian Universitas Udayana Bali dengan ketinggian 30 meter diatas permukaan laut pada bulan Januari sampai Maret 2014. Penelitian dilakukan dengan pemasangan perangkap (trapping) dan pengambilan sampel buah yang terserang lalat buah di Pasar Klungkung, Pasar Gianyar, Pasar Kreneng, Pasar Badung, Pasar Anyar, dan Sentra mangga, Sentra jeruk, Sentra cabai besar, Sentra cabai kecil, serta Sentra semangka.
Hasil penelitian menunjukkan bahwa ada 6 spesies yang ditemukan di lokasi pasar dan sentra buah buahan di Bali yaitu Bactrocera papayae Drew &
Hancock, Bactrocera carambolae Drew & Hancock, Bactrocera umbrosa Fabricius, Bactrocera cucurbitae Coquillete, Bactrocera caudata Fabricius dan Bactrocera albistrigata de Maijere (Diptera:Tephritidae). Keragaman spesies tersebut tergolong rendah yaitu > 1.5. Spesies yang dominan diantara keenam spesies lalat buah tersebut adalah B. carambolae dan B. papayae. Indeks kesamaan antara lokasi penelitian mencapai nilai 80%-100%. Kelimpahan populasi lalat buah mempunyai hubungan positif dengan persentase serangan lalat buah. Terdapat dua spesies parasitoid yang ditemukan berasosiasi dengan lalat buah di lapangan yaitu Fopius sp. dari famili Braconidae. Diantara dua spesies tersebut memiliki tingkat parasitasi yang rendah, tapi Fopius sp. pada tanaman belimbing memiliki tingkat parasitasi yang lebih tinggi yaitu sebesar 56%, sedangkan tingkat parasitasi dari parasitoid Fopius sp. pada tanaman cabai merah hanya sebesar 33%.
Keragaman, kesamaan, dominansi lalat buah dan juga keragaman parasitoid serta tingkat parasitasinya berbeda-beda tiap lokasi dan tiap sampel buah-buahan. Disarankan untuk mengeahui mengenai jenis-jenis lalat buah yang ada dan menyerang buah-buahan di Bali yang dilakukan dalam rentang waktu yang lebih lama dan dalam berbagai fase tanaman inang serta jenis-jenis parasitoid yang efektif dalam pengendalian lalat buah di lapang.
Kata Kunci: Lalat buah, keragaman, kesamaan, dominansi
ix ABSTRACT
Diversity and Population Dynamics of Fruit Flies (Diptera: Tephritidae) Invading Fruit Plants in Bali
Fruit flies (Diptera: Tephritidae) are pests that have significant role for the existence of agriculture. There are about 4000 species of fruit flies in the world and 35% of them are important pests on fruits including commercial fruits that have high economic value. Information about the existence of the types of fruit flies in an area need to be identified and reported as a anticipation and control to fruit crops which are mainly cultivated in Bali. The purpose of this study was to determine the diversity, diversity index and similarity index, the dominant species of fruit fly that exist in the market and at the fruits’ sort center, relation of the population abundance and diversity and levels of parasitoids parasitization’s attack rate associated with each species of fruit flies in the field.
The study was conducted at the Field and in the Laboratory . Laboratory research was conducted at the Laboratory Integrated Pest and Disease Control Management, Faculty of Agriculture, Udayana University, Bali with height 30 meters above sea level in January to March 2014 . Study was conducted by trapping and sampling the fruit flies attacking fruit at Klungkung market, Gianyar market, Kreneng market, Badung market, Anyar market and Center of fruit mango, orange, long chili, rawit chili and watermelon.
The results showed that there were 6 species found in the market and fruits’ center in Bali, namely Bactrocera papayae Drew & Hancock, Bactrocera carambolae Drew & Hancock,Bactrocera umbrosa Fabricius, Bactrocera cucurbitae Coquillete, Bactrocera caudata Fabricius and Bactrocera albistrigatade Maijere (Diptera: Tephritidae). The species diversity was low >
1.5. The dominant species among the six species of the fruit fly were B.carambolae and B.papayae . Index of similarity between the study site reached a value of 80 % -100 % . The abundance of fruit flies population had a positive relationship with the level of fruit flies’ attack. There were two species of parasitoids were found in fruit flies in the field, namely Diasmimorpha longicaudacus and Fopius vandenboschi of the famili Braconidae. Between the two species, the level of parasitism was still low, however Fopius sp. in Averrhoa carambola L had higher parasitism rate that is equal to 56 %, while the rate of parasitism of the parasitoid Fopius sp. in Capsicum annuum L. was only by 20 % . Diversity, similarity, dominance and diversity parasitoids of fruit flies and parasitization levels. it differed from each sample location and fruits. It was advised to know the types of existing fruit flies attacking fruit in Bali which were are done in longer time and in various phases of the plant host and the types of parasitoids which were effective to control fruit flies in the field .
Keywords : Fruit flies , diversity , similarity , dominance
x
RINGKASAN
Lalat buah merupakan hama yang menjadi perhatian dunia di dalam kegiatan ekspor import buah-buahan yang dilakukan oleh suatu Negara. Perhatian itu diberikan karena kegiatan ekspor import komoditas buah segar yang dilakukan oleh masing- masing Negara membawa resiko terhadap masuknya lalat buat dari satu Negara ke Negara lain. Indonesia pernah mengalami masalah adanya komoditas buah-buahan yang menunjukkan gejala serangan lalat buah (Suputa et al., 2006). Permasalahan klasik tersebut sering dihadapi Indonesia karena menyangkut standar mutu (kualitas) produk. Kerusakan yang diakibatkan lalat buah menyebabkan munculnya gejala tusukan lalat buah berupa titik hitam pada buah serta gugurnya buah sebelum mencapai kematangan yang diinginkan, sehingga produksi baik kualitas maupun kuantitas menurun. Berbagai upaya pengendalian lalat buah telah dilakukan, baik secara tradisional maupun dengan menggunakan insektisida kimia. Disamping itu, petani mengendalikan lalat buah dengan atraktan, yaitu senyawa yang dapat menarik lalat buah jantan.
Informasi tentang keberadaan jenis-jenis lalat buah yang ada di suatu daerah perlu diketahui dan dilaporkan sebagai langkah antisipasi dan pengendalian pada tanaman buah yang dibudidayakan terutama di Bali. Informasi tersebut penting karena spesies lalat buah tertentu mempunyai preferensi terhadap jenis inang tertentu. Oleh karena itu perlu penelitian mengenai keragaman dan dinamika populasi lalat buah di area produksi dan membuat daftar spesies, pemetaan dan deteksi lalat buah.
Penelitian dilakukan di Lapangan dan di Laboratorium. Penelitian di Laboratorium dilaksanakan di Laboratorium Pengendalian Hama dan Penyakit Terpadu Tanaman Program Studi Agroekoteknologi Fakultas Pertanian Universitas Udayana Bali dengan ketinggian 30 meter diatas permukaan laut pada bulan Januari sampai Maret 2014. Penelitian dilakukan dengan pemasangan perangkap (trapping) dan pengambilan sampel buah yang terserang lalat buah di Pasar Klungkung, Pasar Gianyar, Pasar Kreneng, Pasar Badung, Pasar Anyar, dan Sentra mangga, Sentra jeruk, Sentra cabai besar, Sentra cabai kecil, serta Sentra semangka.
Hasil penelitian menunjukkan bahwa ada 6 spesies yang ditemukan di lokasi pasar dan sentra buah buahan di Bali yaitu Bactrocera papayae Drew &
Hancock, Bactrocera carambolae Drew & Hancock, Bactrocera umbrosa Fabricius, Bactrocera cucurbitae Coquillete, Bactrocera caudata Fabricius dan Bactrocera albistrigata de Maijere (Diptera:Tephritidae). Keragaman spesies tersebut tergolong rendah yaitu > 1.5. Spesies yang dominan diantara keenam spesies lalat buah tersebut adalah B. carambolae dan B. papayae. Indeks kesamaan antara lokasi penelitian mencapai nilai 80%-100%. Kelimpahan populasi lalat buah mempunyai hubungan positif dengan persentase serangan lalat buah. Terdapat dua spesies parasitoid yang ditemukan berasosiasi dengan lalat buah di lapangan yaitu Fopius sp. dari famili Braconidae. Diantara dua spesies tersebut memiliki tingkat parasitasi yang rendah, tapi Fopius sp. pada tanaman belimbing memiliki tingkat parasitasi yang lebih tinggi yaitu sebesar 56%, sedangkan tingkat parasitasi dari parasitoid Fopius sp. pada tanaman cabai merah hanya sebesar 33%.
xi DAFTAR ISI
Halaman
SAMPUL DALAM...i
PRASYARAT GELAR...ii
LEMBAR PENGESAHAN...…..iii
PENETAPAN PANITIA PENGUJI...iv
SURAT PERNYATAAN BEBAS PLAGIAT...v
UCAPAN TERIMA KASIH...vi
ABSTRAK...viii
ABSTRACT...ix
RINGKASAN...x
DAFTAR ISI...……xi
DAFTAR TABEL ………..………...xiii
DAFTAR GAMBAR...xiv
DAFTAR LAMPIRAN ……….………..………...xv
BAB I PENDAHULUAN 1.1 Latar Belakang………...………1
1.2 Rumusan Masalah ……….………..…5
1.3 Tujuan Penelitian ………...……..5
1.4 Manfaat Penelitian ………..……6
BAB II KAJIAN PUSTAKA 2.1 Lalat Buah ………...………7
2.1.1 Klasifikasi ……….….7
2.1.2 Morfologi Lalat Buah…...……….…10
2.1.3 Bioekologi Lalat Buah.……….…15
2.1.4 Gejala Serangan Lalat Buah .………….………...16
2.1.5 Peran Tanaman Inang dalam Menentukan Besarnya Populasi Lalat Buah ……….……….18
2.1.6 Faktor yang Mempengaruhi Perkembangan Lalat Buah ……….…19
2.1.7 Peran Parasitoid dalam Fluktuasi Populasi Lalat Buah ………23
2.1.8 Persebaran Lalat Buah ………....…28
2.2 Pengaruh Tanaman Inang terhadap Perilaku Serangga ………29
BAB III KERANGKA BERPIKIR DAN HIPOTESIS PENELITIAN 3.1 Kerangka Berpikir ……….33
3.2 Konsep ………...38
3.3 Hipotesis ………41
BAB IV METODE PENELITIAN 4.1 Tempat dan Waktu Penelitian ……….…43
xii
4.2 Alat dan Bahan ………...44
4.2.1 Alat ………44
4.2.2 Bahan ……….…44
4.3 Pelaksanaan Penelitian ………44
4.3.1 Keragaman dan Dominansi Spesies Lalat Buah ……….…45
4.3.1.1 Keragaman dan Indeks Keragaman Lalat Buah di Pasar ….…45 4.3.1.2 Keragaman dan Indeks Keragaman Lalat Buah di Sentra ……49
4.3.1.3 Dominansi Spesies Lalat Buah ……….49
4.3.2 Indeks Kesamaan Lalat Buah di Pasar dan di Sentra Buah …………50
4.3.3 Kelimpahan dan Persentase Serangan Lalat Buah ………..50
4.3.4 Keragaman dan Tingkat Parasitasi Parasitoid ……….…51
BAB V HASIL PENELITIAN 5.1 Keragaman dan Dominansi Spesies Lalat Buah ……….53
5.1.1 Keragaman Spesies Lalat Buah di Pasar Buah-buahan ………...53
5.1.2 Keragaman Spesies Lalat Buah di Sentra Buah-buahan ………….…57
5.1.3 Dominansi Spesies Lalat Buah ………60
5.2 Indeks Kesamaan Lalat Buah di Pasar dan di Sentra Buah ………62
5.3 Hubungan Kelimpahan Populasi dengan Persentase Serangan Lalat Buah62 5.4 Keragaman dan Tingkat Parasitasi Parasitoid yang Berasosiasi dengan Lalat Buah ………63
BAB VI PEMBAHASAN……….…66
BAB VII SIMPULAN DAN SARAN 7.1 Simpulan ………...…73
7.2 Saran ………..…74
DAFTAR PUSTAKA ………..…75
LAMPIRAN ……….79
xiii
DAFTAR TABEL
No Teks Halaman
5.1 Indeks Keragaman Jenis Lalat Buah di 5 Lokasi Pasar ………...54 5.2 Buah-Buahan yang dipasarkan di Lokasi Pasar ………...…..56 5. 3 Indeks Keragaman Jenis Lalat Buah di Sentra Buah dengan Perangkap...57 5.4 Matriks Hubungan antara Spesies Lalat Buah dengan Tanaman Inang…59 5.5 Spesies Lalat Buah yang Menyerang Buah-Buahan ………....….61 5.6 Indeks Kesamaan Spesies Lalat Buah di Lokasi Pasar dan Sentra Buah...62 5.7 Tingkat Parasitasi Parasitoid Terhadap Lalat Buah ………..64
xiv
DAFTAR GAMBAR
No Teks Halaman
2.1a Morfologi Lalat Buah (Famili Tephritidae) ... 7
2.1b Morfologi Bagian-bagian Tubuh Lalat Buah …... 8
2.2 Taksonomi Spesies Lalat Buah ... 9
2.3 Morfologi Genus Bactrocera sp. ... 10
2.4 Morfologi Genus Dacus sp. ... 10
2.5 Morfologi Genus Anastrepha sp. ... 11
2.6 Morfologi Genus Ceratitis sp... 12
2.7 Morfologi Genus Rhagoletis sp. ... 13
2.8 Gejala Serangan Lalat Buah ... 17
2.9 Gejala Membusuknya Buah Akibat Serangan Lalat Buah ... 18
3.1 Kerangka Berpikir Penelitian ... 38
3.2 Kerangka Konsep Penelitian ... 41
4.1 Stoples Rearing ... 48
5.1 Spesies Lalat Buah di Lokasi Pasar ……….…... 55
5.2 Indeks Dominansi Lalat Buah di Lokasi Penelitian …..…... 60
5.3 Hubungan Kelimpahan dengan Persentase Seranga..…... 63
5.4 Spesies Parasitoid Fopius sp. ………..…...…. 65
xv
DAFTAR LAMPIRAN
No. Teks Halaman
1. Spesifikasi Spesies Lalat Buah dengan Atraktan……….……79
2. Morfologi Spesies Lalat Buah ………..…84
3. Karakter Morfologi dari Bagian-Bagian Tubuh Lalat Buah ………90
4. Jumlah Hasil Perangkap di Lokasi Penelitian ………..96
5. Indeks Keragaman Lalat Buah di Lokasi Penelitian ………..…...101
6. Kelimpahan Lalat Buah di Lokasi Penelitian ………..…...104
7. Tanaman-Tanaman yang ada di Sekitar Sentra Buah-Buahan ……...105
8. Gejala Serangan Lalat Buah ……….………...106
BAB I PENDAHULUAN
1.1 Latar Belakang Masalah
Indonesia merupakan salah satu negara penghasil buah tropis yang memiliki keragaman dan keunggulan cita rasa yang cukup baik. Cita rasa dan beragamnya jenis buah-buahan di Indonesia menyebabkan buah-buahan lokal dapat bersaing dengan buah-buahan impor. Selain itu, buah-buahan lokal memiliki harga yang lebih terjangkau bila dibandingkan dengan buah-buahan impor. Tingginya kebutuhan terhadap buah-buahan lokal membuat pengembangan tanaman buah-buahan di Indonesia mengalami peningkatan. Namun, dalam pengembangannya eksport buah-buahan lokal mengalami kendala penyediaan benih bermutu, budidaya sampai penanganan panen. Salah satu kendala dalam budidaya tanaman buah-buahan adalah adanya serangan hama lalat buah.
Lalat buah merupakan hama yang menjadi perhatian dunia di dalam kegiatan ekspor import buah-buahan yang dilakukan oleh suatu negara. Perhatian itu diberikan karena kegiatan ekspor import komoditas buah segar yang dilakukan oleh masing- masing negara membawa resiko terhadap masuknya lalat buat dari satu negara ke negara lain. Indonesia pernah mengalami masalah adanya komoditas buah-buahan yang menunjukkan gejala serangan lalat buah (Suputa et al., 2006). Permasalahan klasik tersebut sering dihadapi Indonesia karena menyangkut standar mutu (kualitas) produk. Standar yang ditetapkan adalah suatu produk tidak mengandung residu berbahaya melebihi ambang batas, tidak mengandung hama penyakit (OPT), dan suatu negara harus menyediakan daftar spesies (pest list) atau deskripsi yang cukup tentang OPT suatu komoditas apabila ingin memperluas pasar perdagangan
komoditas pertanian tersebut (BKP, 2007a). Globalisasi perdagangan buah segar membuat semua negara harus memperhatikan kesehatan tanaman dari serangan hama khususnya lalat buah.
Lalat buah (Diptera: Tephritidae) merupakan hama yang memiliki arti penting bagi pertanian. Terdapat sekitar 4000 spesies lalat buah di dunia dan 35%
di antaranya merupakan hama penting pada buah-buahan termasuk di dalamnya buah-buahan komersial yang mempunyai nilai ekonomi tinggi. Sekitar 75 % tanaman buah-buahan di Indonesia telah terserang lalat buah (Sutrisno, 1999 dalam Sahabudin, 2004). Di samping menyerang buah-buahan, sekitar 40 % larva lalat buah juga hidup dan berkembang pada tanaman sayur-sayuran, famili asteraceae (Compositae) (Kuswadi, 2001).
Di Indonesia, lalat buah sebagai hama telah diketahui sejak tahun 1920, dan telah dilaporkan menyerang mangga di Pulau Jawa. Pada tahun 1938, lalat buah juga dilaporkan menyerang cabai, jambu, belimbing dan sawo. Survei lalat buah di Indonesia yang dilakukan oleh Hardy pada tahun 1985 menemukan 66 spesies. Survei berikutnya yang dilakukan oleh Balai Karantina Pusat dari tahun 1992 - 1994 menemukan sekitar 47 spesies, dan 20 spesies di antaranya merupakan kompleks Bactrocera dorsalis (Drew & Hancock 1994; Hamzah, 2004). Sementara laporan AQIS (2008) menyebutkan bahwa terdapat 63 spesies lalat buah di Indonesia namun tidak termasuk Ceratitis capitata Wied.yang dikenal dengan sebutan Mediterranean Fruit Fly atau Medfly sebagai hama penting tanaman jeruk di wilayah sekitarlaut Tengah (White & Harris, 1992).
Orr (2002) melaporkan bahwa ada sekitar 90 spesies lalat buah di Indonesia bagian barat termasuk lalat buah jenis lokal (indegenous). Delapan spesies di
antaranya merupakan hama penting yaitu Bactrocera albistrigata (de Maijere), B.
dorsalis Hendel, B. carambolae Drew and Hancock, B. papayae Drew and Hancock, B. umbrosa (Fabricius), B. (Zeugodacus) caudata (Fabricius) dengan sinonim Bactrocera (Z) tau (Walker), Bactrocera (Z) cucurbitace (Coquillete) dan Dacus (Callantra) longicornis (Wiedemann).
Kerusakan yang diakibatkan lalat buah menyebabkan munculnya gejala tusukan lalat buah berupa titik hitam pada buah serta gugurnya buah sebelum mencapai kematangan yang diinginkan, sehingga produksi baik kualitas maupun kuantitas menurun. Kehilangan hasil yang diakibatkan oleh serangan hama lalat buah bervariiasi antara 30-100% bergantung pada kondisi lingkungan dan kerentanan jenis buah yang diserangnya (Gupta & Verma, 1978; Dhilton et al., 2005a, 2005b dan 2005c). Intensitas serangan lalat buah di Bali menunjukkan variasi yang cukup besar, yaitu antara 6,4 - 70% (Sarwono, 2003). Sodiq (2004) menyatakan bahwa intensitas serangan lalat buah pada mangga berkisar antara 14,8%-23%, namun tidak jarang kerusakan yang diakibatkan lalat buah khususnya pada belimbing dan jambu biji dapat mencapai 100%.
Berbagai upaya pengendalian lalat buah telah dilakukan, baik secara tradisional dengan membungkus buah dengan kantong plastik, kertas koran atau daun kelapa maupun dengan menggunakan insektisida kimia. Disamping itu, petani mengendalikan lalat buah dengan atraktan, yaitu senyawa yang dapat menarik lalat buah jantan. Teknik ini efektif mengendalikan lalat buah jantan yang masuk ke dalam perangkap beratraktan. Teknik berikutnya yaitu teknik jantan mandul yang merupakan cara pengendalian dengan membuat lalat buah jantan menjadi infertil, artinya lalat buah jantan masih dapat membuahi betina, namun
telur yang dihasilkan steril dan larva dalam keadaan rusak (Vijaysegaran &
Osman 1991 dalam Shiga, 1991).
Pengendalian lalat buah lainnya yaitu dengan menggunakan musuh alami sebagai pengatur keseimbangan di alam. Musuh alami dapat berupa predator, pathogen dan parasitoid. Parasitoid yang berasal famili Braconidae (Hymenoptera), yaitu Fopius sp. dan Biosteres, sp (Siwi et.al., 2006). Parasitoid Famili Braconidae dapat mencapai tingkat parasitisasi sebesar 57% dan parasitisasi oleh parasitoid Famili Euphelmidae pada B.oleae dapat mencapai 80 sampai dengan 95% (Malau, 1968; Delrio, 1978; Delrio dan Gavalloro, 1977;
Delio dan Prota, 1976 dalam Flecher, 1987). Di Kamerun, diperoleh sejumlah besar parasitoid (Fopius. sp. dan Biosteres, sp.) pada buah kopi dengan derajat parasitisasi pada pupa lalat buah berkisar antara 10 sampai 56% dengan rata-rata 35% (Garry et al., 1986). Di Yogyakarta didapatkan 33,9% puparium B.
carambolae yang menginfestasi buah belimbing terparasit oleh B.vandenboschi (Soesilohadi, 1995).
Informasi tentang keberadaan jenis-jenis lalat buah yang ada di suatu daerah perlu diketahui dan dilaporkan sebagai langkah antisipasi dan pengendalian pada tanaman buah yang dibudidayakan terutama di Bali. Informasi tersebut penting karena spesies lalat buah tertentu mempunyai preferensi terhadap jenis inang tertentu (Muryati et al., 2005). Oleh karena itu perlu penelitian mengenai keragaman dan dinamika populasi lalat buah di area produksi dan membuat daftar spesies, pemetaan dan deteksi lalat buah. Diketahuinya keragaman dan dinamika populasi lalat buah di Bali mempunyai arti penting dalam perencanaan dan pelaksanaan tindakan monitoring maupun pengendalian
yang akan dilakukan agar lebih efektif dan efisien. Disamping itu, informasi tentang keberadaan jenis-jenis lalat buah, parasitoid dan tanaman inang yang ada di suatu daerah perlu diketahui dan dilaporkan untuk mengantisipasi ledakan hama tersebut di lapangan.
1.2 Rumusan Masalah
Terdapat lima masalah yang ingin dijawab dalam penelitian ini yang dirumuskan sebagai berikut:
1. Bagaimanakah keragaman, dan dominansi spesies lalat buah yang berada di pasar dan di sentra buah-buahan di Bali?
2. Bagaimanakah kesamaan spesies lalat buah yang berada di pasar dan di sentra buah-buahan di Bali?
3. Bagaimanakah hubungan kelimpahan populasi dan persentase serangan lalat buah yang menyerang tanaman buah-buahan di lapang?
4. Bagaimanakah keragaman dan tingkat parasitisasi parasitoid yang berasosiasi dengan masing-masing spesies lalat buah di lapangan?
1.3 Tujuan
Penelitian ini bertujuan untuk:
1. Untuk mengetahui keragaman dan dominansi spesies lalat buah yang berada di pasar dan di sentra buah-buahan di Bali.
2. Untuk mengetahui kesamaan spesies lalat buah yang berada di pasar dan di sentra buah-buahan di Bali.
3. Untuk mengetahui hubungan kelimpahan populasi dan persentase serangan lalat buah yang menyerang tanaman buah-buahan di lapang.
4. Untuk mengetahui keragaman dan tingkat parasitisasi parasitoid yang berasosiasi dengan masing-masing spesies lalat buah di lapangan.
1.4 Manfaat Penelitian
Dari penelitian ini diharapkan dapat diperoleh luaran seperti:
1. Secara akademis, hasil penelitian ini akan memperkaya pengetahuan mengenai lalat buah dan parasitoid yang meliputi keragaman, kesamaan, kelimpahan, dominansi lalat buah dan parasitoidnya pada tanaman buah- buahan di Bali.
2. Secara praktis, hasil penelitian ini dapat membantu dalam upaya pengendalian hama lalat buah pada tanaman buah-buahan yang dibudidayakan.
BAB II
KAJIAN PUSTAKA
2.1 Lalat buah 2.1.1 Klasifikasi
Lalat buah diklasifikasikan sebagai berikut:
Kingdom : Animalia Filum : Arthropoda Kelas : Insecta Ordo : Diptera Famili : Tephritidae
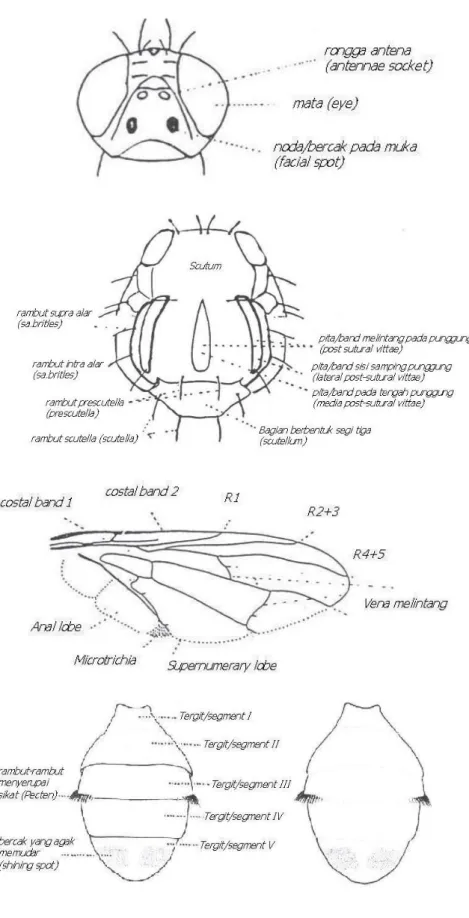
Bentuk morfologi famili Tephritidae antara spesies satu dengan yang lainnya hampir mirip. Maka dari itu perlu dilakukan identifikasi yang teliti dan seksama dalam menentukan spesies dari famili Tephritidae. Secara umum morfologi famili Tephtitidae dapat dilihat pada Gambar 2.1a
Gambar 2.1.a Morfologi Lalat Buah (Famili Tephritidae)
Gambar 2.1.b Morfologi Bagian-Bagian Tubuh Lalat Buah Famili Tephritidae
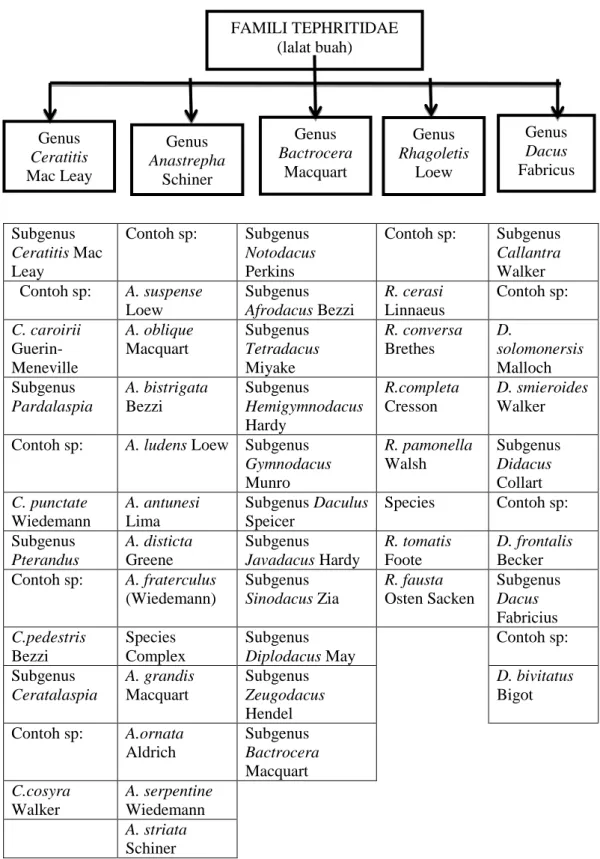
Menurut White and Harris (1992), lalat buah memiliki 5 genus yaitu pada Genus Ceratitis Mac Leay, Genus Anastrepha Schiner, Genus Bactrocera Macquart, Genus Rhagoletis Loew dan Genus Dacus Fabricius (Gambar 2.1b).
Subgenus Ceratitis Mac Leay
Contoh sp: Subgenus Notodacus Perkins
Contoh sp: Subgenus Callantra Walker Contoh sp: A. suspense
Loew
Subgenus Afrodacus Bezzi
R. cerasi Linnaeus
Contoh sp:
C. caroirii Guerin- Meneville
A. oblique Macquart
Subgenus Tetradacus Miyake
R. conversa Brethes
D.
solomonersis Malloch Subgenus
Pardalaspia
A. bistrigata Bezzi
Subgenus
Hemigymnodacus Hardy
R.completa Cresson
D. smieroides Walker Contoh sp: A. ludens Loew Subgenus
Gymnodacus Munro
R. pamonella Walsh
Subgenus Didacus Collart C. punctate
Wiedemann
A. antunesi Lima
Subgenus Daculus Speicer
Species Contoh sp:
Subgenus Pterandus
A. disticta Greene
Subgenus
Javadacus Hardy
R. tomatis Foote
D. frontalis Becker Contoh sp: A. fraterculus
(Wiedemann)
Subgenus Sinodacus Zia
R. fausta Osten Sacken
Subgenus Dacus Fabricius C.pedestris
Bezzi
Species Complex
Subgenus Diplodacus May
Contoh sp:
Subgenus Ceratalaspia
A. grandis Macquart
Subgenus Zeugodacus Hendel
D. bivitatus Bigot Contoh sp: A.ornata
Aldrich
Subgenus Bactrocera Macquart C.cosyra
Walker
A. serpentine Wiedemann A. striata Schiner
Gambar 2.2 Taksonomi Spesies Lalat Buah (Harris, 1992) FAMILI TEPHRITIDAE
(lalat buah)
Genus Anastrepha
Schiner
Genus Bactrocera
Macquart
Genus Rhagoletis
Loew
Genus Dacus Fabricus Genus
Ceratitis Mac Leay
2.1.2 Morfologi Lalat Buah
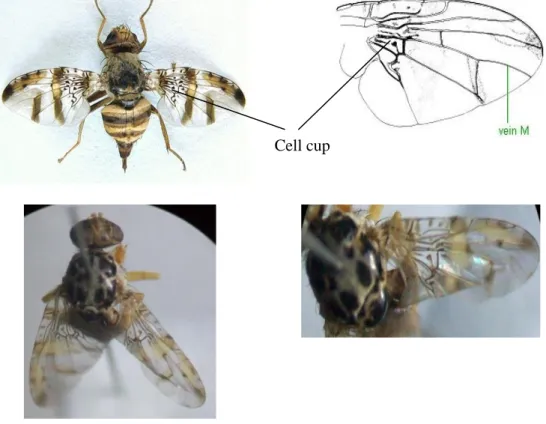
Seperti yang disebutkan oleh White and Harris (1992), famili Tephritidae memiliki 5 genus yang morfologinya berbeda-beda. Morfologi kelima genus tersebut adalah sebagai berikut:
1. Genus Bactrocera :
Cell cup sempit dengan extension sangat panjang.
Pola sayap biasanya berupa costal band dan anal streak
Abdomen oval dengan tergum I – V tidak bergabung (not fused)
Gambar 2.3 Morfologi Genus Bactrocera sp.
2. Genus Dacus :
Cell cup sempit dengan extension sangat panjang.
Cell cup
Ceromata
Pola sayap biasanya berupa costal band dan anal streak Abdomen oval dengan tergum I – V bergabung (fused)
Gambar 2.4 Morfologi Genus Dacus sp.
3. Genus Anastrepha :
Cell cup lebar dengan extension agak panjang
Biasanya terdapat crossband membentuk pola warna pada sayap Vena M membentuk curva sebelum mencapai pinggir sayap
Cell cup
Cell cup
Gambar 2.5 Morfologi Genus Anastrepha sp.
4. Genus Ceratitis :
Cell cup lebar extension pendek, Vena M pada sayap hampir membentuk sudut siku-siku
Biasanya terdapat spot dan bintik pada basal cell sayap
Gambar 2.6 Morfologi Genus Ceratitis sp.
Cell cup
5. Genus Rhagoletis :
Cell cup lebar dengan extension pendek,
Vena M pada sayap hampir membentuk sudut siku-siku
Tidak terdapat spot dan bintik-bintik pada basal cell dari sayap
Gambar 2.7 Morfologi Genus Rhagoletis sp.
Telur lalat buah secara umum berwarna putih atau putih kekuningan berbentuk bulat panjang. Panjang telur antara 0.3 mm-0.8 mm dan lebar 0.2 mm dengan micropyle protruding yang tipis di bagian akhir anterior (CABI, 2007).
Telur akan menetas menjadi larva dua hari setelah diletakkan di dalam buah (Ditlin Hortikultura 2006).
Larva berwarna putih keruh kekuningan, berbentuk bulat panjang dan salah satu ujungnya runcing. Kepala berbentuk runcing, mempunyai alat pengait
Cell cup
dan bintik yang jelas. Larva instar ketiga berukuran sedang, dengan panjang 7.0 mm- 9.0 mm dan lebar 1.5-1.8 mm (White & Harris, 1994).
Puparium lalat buah berbentuk oval berwarna kuning kecoklatan dengan panjang ± 5 mm (Ditlin Hortikultura, 2006). Imago lalat buah umumnya memiliki ciri-ciri penting di kepala, toraks, sayap, dan abdomen. Kepala terdiri atas antena, mata, dan spot. Pada toraks terdapat dua bagian penting yakni skutum dan skutelum. Sayap mempunyai bentuk dan pola pembuluh yakni costa, radius, median, cubitus, anal, r-m dan dm-cu (pembuluh sayap melintang). Pada genus Bactrocera ruas-ruas abdomen terpisah dan genus Dacus ruas-ruas abdomen menyatu. Pada abdomen, Bactrocera, tergum I dan II menyatu, tergum III-V terpisah.
Pada Dacus, antara toraks dan abdomen mempunyai pinggang ramping (petiole) sehingga menyerupai tawon (Siwi et al., 2006). Lalat buah komplek B.
dorsalis memiliki membran sayap yang cerah, kecuali pada costal band (tidak mencapai R4+5); cell basal costa dan costa tidak berwarna dan tidak ada microtrichia. Skutum umumnya berwarna hitam dengan pita kuning di sisi lateral dan tidak memiliki pita kuning di bagian tengah skutum. Skutelum berwarna kuning kecuali pada bagian basal dengan pita hitam yang tipis. Abdomen dengan garis medial pada tergum III-V dan berwarna hitam di sisi lateral (CABI, 2007).
Abdomen umumnya mempunyai pita melintang dan pita membujur berwarna hitam atau berbentuk huruf T yang kadang-kadang tidak jelas (Lawson et al., 2003). Ujung abdomen lalat betina lebih runcing dan mempunyai alat peletak telur (ovipositor) yang cukup kuat untuk menembus kulit buah. Pada jantan, abdomen
lebih bulat dan pada tergum III di kedua sisi lateral abdomen terdapat pecten (Drew, 1989).
2.1.3 Bioekologi Lalat Buah
Lalat buah mengalami perkembangan sempurna atau dikenal dengan perkembangan holometabola. Perkembangan holometabola memiliki 4 fase metamorfosis yaitu: telur, larva, pupa, dan imago (Vijaysegaran & Drew, 2006).
Telur lalat buah diletakkan berkelompok 2-15 butir. Lalat buah betina dapat meletakkan telur 1- 40 butir/hari. Seekor lalat betina dapat meletakkan telur 100- 500 butir (Sodiq 1992 dalam Siwi, 2005). Menurut Vijaysegaran dan Drew (2006), satu ekor betina B. dorsalis dapat menghasilkan telur 1200 - 1500 butir.
Telur-telur diletakkan pada buah di tempat yang terlindung dan tidak terkena sinar matahari langsung serta pada buah-buah yang agak lunak dan permukaannya agak kasar (Ditlin Hortikultura, 2006).
Larva terdiri atas 3 instar. Larva hidup dan berkembang di dalam daging buah selama 6-9 hari. Pada instar ke tiga menjelang pupa, larva akan keluar dari dalam buah melalui lubang kecil. Setelah berada di permukaan kulit buah, larva akan melentingkan tubuh, menjatuhkan diri dan masuk ke dalam tanah. Di dalam tanah larva menjadi pupa (Djatmiadi & Djatnika, 2001).
Pupa pada awalnya berwarna putih, kemudian berubah menjadi kekuningan dan akhirnya menjadi coklat kemerahan. Masa pupa berkisar antara 4 - 10 hari (Ditlin Hortikultura, 2006). Pupa berada di dalam tanah atau pasir pada kedalaman 2-3cm di bawah permukaan tanah atau pasir. Setelah 6 -13 hari, pupa menjadi imago (Djatmiadi & Djatnika, 2001).
Siklus hidup lalat buah dari telur sampai imago di daerah tropis berlangsung lebih kurang 27 hari dapat dilihat pada Gambar 2.7. Lama hidup imago betina berkisar antara 23-27 hari dan imago jantan antara 13-15 hari. Imago betina setelah kopulasi akan meletakkan telur setelah 3-8 hari. Nisbah kelamin jantan berbanding dengan betina yakni 1:1 (Sodiq 1992 dalam Siwi, 2005). Lalat buah dewasa hidup bebas di alam dan bergerak secara aktif. Lalat betina sering dijumpai di sekitar tanaman buah-buahan dan sayuran pada pagi dan sore hari, sedangkan lalat buah jantan bergerak aktif dan memburu lalat buah betina untuk melakukan kopulasi (Siwi, 2005).
2.1.4 Gejala Serangan Lalat Buah
Lalat buah betina menyerang buah dengan memasukkan telur melalui ovipositornya ke dalam buah (Agarwal, 1984). Pemasukan ovipositor ke dalam buah menyebabkan adanya gejala tusukan pada buah belimbing pada Gambar 2.8 wsterlihat spot berwarna gelap cokelat kehitaman.
Telur kemudian menetas menjadi larva yang hidup, makan dan berkembang di dalam buah sehingga buah menjadi busuk berisi larva atau dikenal dengan belatung (Kalshoven, 1981). Sesudah telur menetas, larva membuat lubang di dalam buah sehingga mempermudah masuknya bakteri dan cendawan (Siwi et al., 2006). Lalat buah hidup secara simbiosis mutualisme dengan bakteri, sehingga ketika lalat buah meletakkan telur pada buah, bakteri akan terbawa dengan diikuti cendawan yang akhirnya menyebabkan busuk. Sesudah telur menetas, larva mengorek daging buah sambil mengeluarkan enzim perusak atau pencerna yang berfungsi melunakkan daging buah sehingga mudah diisap dan
dicerna. Enzim tersebut diketahui yang mempercepat pembusukan, selain bakteri pembusuk yang mempercepat aktivitas pembusukan buah. Bakteri tersebut hidup pada dinding ovari, tembolok, dan ileum lalat (Hill 1983; Ria, 1994). Buah yang terserang lalat buah dan busuk, akhirnya jatuh ke tanah.
Gambar 2.8 Gejala Serangan Lalat Buah
Serangan lalat buah pada buah yang terserang terdapat luka tusukan dalam ukuran kecil, seperti tertusuk jarum. Hal tersebut akan mengakibatkan terdapatnya spot hitam pada buah. Buah yang terserang menjadi busuk lunak dan menghitam seperti pada Gambar 2.9. Luka akibat tusukan menimbulkan infeksi sekunder berupa busuk buah, baik yang disebabkan oleh cendawan maupun bakteri. Buah yang terkena tusukan lalat buah ini akan rontok. Jika buah dibelah akan terlihat biji-biji berwarna hitam dan terdapat belatung yang merupakan larva lalat buah.
Gejala Tusukan Ovipositor Lalat
Buah
Gambar 2.9 Gejala Membusuknya Buah Akibat Serangan Lalat Buah.
2.1.5 Peranan Tanaman Inang dalam Menentukan Besarnya Populasi Lalat Buah
Ketersediaan buah dapat memoengaruhi fenologi dan kelimpahan lalat buah (Israely et al., 1997). Kelimpahan populasi lalat buah jantan dipengaruhi oleh tingkat kematangan buah sebagai contoh misalnya di Hanalei dan Kilauea sebelah timur Kanei, Hawaii tahun 1988-1989 dengan puncak kelimpahan jambu air terjadi bulan Mei dan September. Pada saat tersebut lalat buah oriental tidak ditemukan di perkebunan komersial buah jambu sebelum terjadi pematangan buah dan peningkatan lalat buah terjadi dengan meningkatnya kematangan buah (Stark et al., 1991 dalam Vargas et al., 1993). Hasil penelitian Aluja et al., (1996) juga menunjukkan bahwa 90% sampel buah manga dari perkebunan manga komersial terinfeksi oleh A.obliqua. Stark and Vargas (1992), Strark et al., (1991) dan Tan (1994) berpendapat fenologi tanaman inang merupakan penduga paling baik dalam memprediksi dinamika populasi lalat buah, Bactrocera dorsalis kompleks.
Jambu batu (Psidium guajava) merupakan tanaman inang utama B. dorsalis di Buah membusuk
beberapa bagian dunia dan puncak populasi B.dorsalis bertepatan dengan musim buah jambu.
2.1.6 Faktor yang Mempengaruhi Perkembangan Lalat Buah
Dinamika populasi lalat buah terjadi karena pengaruh kombinasi antara faktor lingkungan yang bekerja pada populasi dan karakteristik intrinsik spesies dan individu-individu (Celedonia et al., 1995 dalam Israely et al., 1997). Secara umum lalat buah terbagi menjadi dua kelompok sifat populasi yaitu lalat buah univoltine yang habitatnya di daerah temperate dan lalat buah multivoltine yang habitatnya di daerah tropis dan subtropics (Harris, 1993). Besarnya populasi lalat buah di lingkungan temperate diatur oleh suhu, sedangkan kelimpahan populasi lalat buah di daerah tropis diatur oleh curah hujan (Celedonio et al., 1995 dalam Israely et al., 1997). Sebagai contoh misalnya B.cucubitae Conquillet yang hidup di daerah tropis, kelimpahan populasinya dipengaruhi kelembaban, sedangkan Rhagoletis pomonella (Walsh) yang hidup di daerah temperate kelimpahan populasinya dikendalikan oleh suhu (Bateman, 1972 dalam Israely et al., 1997).
Contoh lain misalnya di India populasi lalat buah melon meningkat bila turun hujan cukup memadai dan menurun selama musim kemarau (Allwood, 1996).
Faktor-faktor lain yang mempengaruhi dinamika populasi adalah faktor suhu, kelembaban, cahaya, curah hujan, tanaman inang, dan musuh alami. Faktor iklim berpengaruh pada pemencaran, perkembangan, daya bertahan hidup, perilaku, reproduksi, dinamika populasi, dan peledakan hama (McPheron &
Steck, 1996). Menurut Messenger (1976 dalam Siwi, 2005), iklim berpengaruh
terhadap perilaku seperti aktifitas kawin dan peletakan telur yang mempengaruhi angka kelahiran, kematian, dan penyebaran serangga.
Curah hujan mempunyai pengaruh yang kuat terhadap kelimpahan buah inang dan populasi B.dorsalis dewasa (Tan dan Serit, 1994).. Kemunculan imago lalat buah dari pupa juga dipengaruhi oleh kelembaban tanah. Kelembaban tanah yang optimal bagi kehidupan pupa lalat buah antara 80-90% (Sodiq, 1993). Pada umumnya kepadatan populasi meningkat dengan curah hujan yang meningkat, akan tetapi melalui suatu studi diketahui bahwa terjadi ledakan pada kepadatan populasi B.dorsalis setelah badai topan. Hal tersebut menunjukkan bahwa iklim berperan sebagai faktor mortalitas yang tidak tergantung kepadatan (Williamson et al., 1985 ). Kepadatan populasi B.dorsalis cenderung tinggi selama musim hujan, dan peningkatan populasinya tidak harus berkorelasi dengan fenologi tanaman inang (Bagle & Prasad 1983). Walaupun demikian curah hujan tidak selalu berkorelasi secara linier dengan kelimpahan populasi lalat buah.
Kelimpahan lalat buah dengan curah hujan memiliki hubungan yang saling berkaitan, seperti lalat buah spesies Anastrepha oblique mempunyai hubungan yang tidak linier (Aluja et al., 1996).
Kelembaban yang rendah dapat menurunkan keperidian lalat buah dan meningkatkan mortalitas imago yang baru keluar dari pupa. Kelembaban udara yang terlalu tinggi (95-100%) dapat mengurangi laju peletakan telur (Bateman,1972). Semakin tinggi kelembaban udara maka lama perkembangan akan semakin panjang. Kelembaban optimum perkembangan lalat buah berkisar antara 70-80%. Lalat buah dapat hidup baik pada kelembaban antara 62-90%
(Landolt & Quilici 1996).
Intensitas cahaya dan lama penyinaran dapat mempengaruhi aktivitas lalat betina dalam perilaku makan, peletakan telur, dan kopulasi. Lalat aktif pada keadaan terang, yaitu pada siang hari dan kopulasi pada intensitas cahaya rendah.
Selain itu, lalat betina yang banyak mendapatkan sinar akan lebih cepat bertelur ( Siwi, 2005).
Suhu adalah faktor yang mempengaruhi laju perkembangan stadium muda lalat buah dan akan menentukan fluktuasi populasinya (Flecher, 1987). Pada daerah tropis yang tidak banyak mengalami fluktuasi suhu, fluktuasi populasi lalat buah secara nyata tetap terjadi. Populasi lebih besar terjadi selama musim kemarau daripada di musim hujan. Untuk lalat buah yang multivoltine, suhu di bawah 210C dapat menurunkan laju pertumbuhan lalat buah selama stadium muda. Produksi telur maksimum terjadi pada suhu 250C sampai dengan 300C (Allwood, 1996).
Bateman (1968) dalam Pritchard (1970) menyatakan bahwa faktor cuaca adalah determinan paling penting pada kelimpahan populasi Dacus tryoni. D.
tryoni betina lebih cepat perkembangan pematangan ovarynya pada suhu tinggi daripada pada suhu rendah, sebagai contoh misalnya pada suhu 150C persentase perkembangan per hari sebesar 2,94% sedangkan pada suhu 250C persentase perkembangan mencapai 17,95%, kemudian menurun dengan meningkatnya suhu yaitu menjadi 15,48% pada 300C. Menurut Bateman (1972), suhu berpengaruh terhadap perkembangan, keperidian, lama hidup, dan mortalitas Bactrocera spp.
Lalat buah umumnya dapat hidup dan berkembang pada suhu 10-30ºC. Pada suhu antara 25-30oC telur lalat buah dapat menetas dalam waktu yang singkat yaitu 30- 36 jam.
Lalat buah yang menyerang buah-buahan musiman, akan mempunyai dinamika populasi yang erat hubungannya dengan keberadaan buah. Lalat buah yang menyerang tanaman sayuran mempunyai dinamika populasi yang berbeda karena keberadaan inang tanaman sayuran ada sepanjang tahun. Berdasarkan hasil penelitian Muryati et al. (2005), B. carambolae dan B. papayae merupakan spesies lalat buah yang paling banyak ditemukan. Hal ini disebabkan tanaman inang kedua spesies tersebut tersedia sepanjang waktu. Inang tersebut antara lain jambu biji, jambu air, belimbing, manggis, nangka, pisang, dan cabe..
Tingkat kematangan buah berpengaruh terhadap kehidupan lalat buah.
Buah yang lebih matang lebih disukai oleh lalat buah untuk meletakkan telur daripada buah yang masih hijau. Tingkat kematangan buah sangat mempengaruhi populasi lalat buah. Jenis pakan yang banyak mengandung asam amino, vitamin, mineral, air, dan karbohidrat dapat memperpanjang umur serta meningkatkan keperidian lalat buah. Peletakan telur dipengaruhi oleh bentuk, warna, dan tekstur buah. Bagian buah yang ternaungi dan agak lunak merupakan tempat ideal untuk peletakan telur (Siwi, 2005).
Musuh alami adalah salah satu faktor penyebab kematian lalat buah.
Musuh alami dapat berupa parasitoid, predator, dan patogen. Di lapang dijumpai parasitoid famili Braconidae (Hymenoptera), yaitu Fopius spp. dan Biosteres spp.
Predator yang memangsa lalat buah antara lain semut, laba-laba, kumbang, dan cocopet. Patogen yang menyerang lalat buah diduga cendawan Mucor sp. (Siwi et al., 2006).
2.1.7 Peranan Parasitoid dalam Fluktuasi Populasi Lalat Buah
Telur, larva dan pupa lalat buah diserang oleh sejumlah parasit Hymenoptera yang merupakan mayoritas musuh alami lalat. Famili Braconidae merupakan parasitoid mayoritas dengan enam belas spesies. Terutama terdiri dari opiines, tujuh spesies yang telah ditemukan dari Malaysia (van den Bosch &
Haramoto, 1951; Christenson & Foote, 1960; Clausen, 1972; Deulucci, 1976;
Wharton & Gilstrap, 1983; Ooi, 1984; Vijaysegaran, 1984; Rohani, 1986;
Serit et al, 1986;.. Udayagiri, 1987; Wharton, 1989; Palacio, 1991; Ramadhan et al, 1995). Spesies yang diamati tingkat parasitasinya pada spesies Bactrocera dorsalis pada buah belimbing di kebun dan di desa-desa di Malaysia adalah;
Fobius (Sinonim = Fopius) arisanus (Sonan), Diachasmimorpha longicaudatus (Ashmead), Psytallia (Sinonim = Fopius) fletcheri (Silvestri), Psytallia (Sinonim
= Fopius) incisi (Silvestri), Fopius vandenboschi (Fullaway), Fopius skinneri (Fullaway) (Ooi, 1984; Vijaysegaran, 1984; Rohani, 1986; Serit et al, 1986;. Serit 1987; Palacio et al, 1992, Ibrahim,dkk.,1994). Sementara, satu-satunya musuh alami yang menyerang Bactrocera umbrosa adalah Pilinothrix sp. (Hymenoptera:
Cynipidae) (Yunus & Ho, 1980).
Larva B.dorsalis yang terparasit oleh parasitoid Famili Braconidae dapat mencapai 57% dan parasitasi oleh parasitoid Famili Euphelmidae pada B.oleae dapat mencapai 80 sampai dengan 95% (Malau, 1968; Delrio, 1978; Delrio dan Gavalloro, 1977; Delio dan Prota, 1976 dalam Flecher, 1987). Di Kamerun, diperoleh sejumlah besar parasitoid (Fopius sp. dan Fopius sp.) pada buah kopi dengan derajat parasitasi pada pupa lalat buah berkisar antara 10 sampai 56%
dengan rata-rata 35% (Garry et al., 1986). Di Yogyakarta didapatkan 33,9%
puparium B. carambolae yang menginfestasi buah belimbing terparasit oleh B.vandenboschi (Soesilohadi, 1995). Beberapa parasitoid seperti Strepsiptera menyerang lalat buah dewasa, tetapi tidak berpengaruh pada populasi lalat buah.
Komposisi jenis dan efektivitas spesies parasitoid tertentu dari spesies opiine bisa bervariasi tergantung pada wilayah dan jenis buah yang di serang, ukuran dan kematangan mempengaruhi tingkat parasitisasi larva lalat buah (Daratan et al, 1950;. Van den Bosch & Haramoto 1953, Hinckly, 1965;
Gonzalez, 1975; Wharton et al, 1981, Nishida et al, 1985; Harris & Lee, 1986; Wong & Ramadhan, 1995). Tingkat parasitisasi terbesar oleh Diachasmimorpha longicaudatus telah ditemukan dari buah-buahan kecil seperti buah kopi, kopi (Harris et al., 1986), loquat, Ertobtrya japonica (Lindl.), dan buah persik Prunus persica L. (Wong et al. , 1984; Wong & Ramadhan, 1987) dibandingkan dari buah jeruk besar (Wharton dkk, 1981;. Harris et al, 1986 dan 1988;. Harris & Bautista, 1996).
Opiine kompleks parasitoid Bactrocera dorsalis di belimbing bervariasi setiap habitat dan dari tempat ke tempat. Dalam studi lapangan terpisah pada komposisi parasitoid B. dorsalis di kebun belimbing yang berbeda di Serdang, Selangor, Ooi (1984) mencatat Diachasmimorpha longicaudatus, Fopius vandenboschi dan Fobius insici. Dari tiga spesies, B. vandenboschi adalah parasitoid dominan, dibandingkan P. incisi dan D. longicaudatus. Parasitisme oleh ketiga spesies tersebut berkisar 15,1-56,8% dengan rata-rata 28%
(Vijaysegaran, 1984).
Penurunan kelimpahan lalat buah di alam sering dikendalikan oleh empat parasitoid dari subfamili opiinae yaitu: F. arisanus; B. vandenboschi; D.
longicaudatus; B. skinneri dalam buah belimbing di Penang, Malaysia Barat (Serit et al, 1986; Serit, 1987.). Berdasarkan kelimpahan imago parasitoid yang muncul dari pupa sampel buah-buahan, empat spesies parasitoid dikaitkan dengan B.
dorsalis di kebun belimbing dari Universitas Putra Malaysia (UPM) di Puchong, Selangor. Parasitoid didominasi oleh B. persulcatus. Tingkat parasitisasi oleh masing-masing spesies adalah B. persulcatus, 46.53%; F. arisanus, 32.82%; D.
longicaudatus, 15,69%; P. fletcheri, 4,95%. Bersama-sama, parasitoid ini menyebabkan parasitasi keseluruhan rata-rata 36,96% (Palacio, 1991).
Fopius arisanus adalah satu-satunya parasitoid telur-larva dari spesies Opiinae (Wharton & Gilstrap, 1983). Lalat buah pada fase telur-larva akan dibunuh oleh F. arisanus dan F. arisanus tahap pupa dan parasitoid dewasa akan muncul. Biosteres vandenboschi merupakan parasitoid larva instar pertama Bactrocera dorsalis. Mulanya setelah terparasit larva dapat berkembang secara normal tetapi akhirnya dibunuh pada tahap kepompong. Parasitoid B.
vandenboschi sebagai kontrol biologis lalat buah memiliki kemampuan untuk memparasitasi dan dikenal sebagai pengendali tujuh spesies yang berbeda dari hama tephritid (Wharton & Gilstrap, 1983) dan preferensi B. vandenboschi pada lalat buah sangat cepat (instar pertama dam instar kedua) yang terjadi dekat permukaan buah (van den Bosch & Haramoto, 1953;. Ramadhan dkk, 1995).
Diachasmimorpha longicaudatus Ashmead adalah endoparasitoid larva- pupa soliter dari sejumlah spesies lalat buah tephritid ekonomis penting (Clausen et al, 1965; Greany et al, 1976). Parasitasi pada instar larva kedua dan ketiga, biasanya terjadi pada buah yang hampir membusuk. Penemuan inang oleh imago
betina dari D. longicaudatus melibatkan produk fermentasi yang dihasilkan dari buah yang membusuk (Greany et al., 1977).
Psytallia fletcheri awalnya ditemukan pada Bactrocera cucurbitae di India (Silvestri, 1916, Pruthi, 1937). Ini menjadi parasitoid yang paling penting dari di Hawaii (Fullaway, 1920, Swezey, 1928). Keberadaannya di belimbing di Bactrocera dorsalis di Malaysia pertama kali dilaporkan oleh Vijaysegaran (1984)..
Vargas et al. (1993) menyatakan bahwa terdapat korelasi positif antara Fopius arisanus, parasitoid dominan dengan jumlah individu lalat buah yang mengindikasikan adanya hubungan tergantung kepadatan. Seperti misalnya laju parasitasi B.arisanus pada Ceratitis capitata dan Bactrocera dorsalis bervariasi pada habitat-habitat. Hal tersebut menunjukkan bahwa distribusi, kelimpahan dan kopulasi dan reproduksi ada di bawah pengaruh kompleks faktor fisiologis dan lingkungan (Nishida et al., 1985 dalam Harris dan Okamoto, 1991). Kualitas larva lalat buah yang bervariasi dari minggu ke minggu mengakibatkan fluktuasi populasi parasitoid yang nyata (Messing et al., 1993). Parasitoid yang menginfestasi telurnya pada larva inang instar ketiga menghasilkan generasi parasitoid yang berkualitas tinggi (Wong dan Ramadhan, 1992 dalam Messing et al, 1993). Besarnya populasi parasitoid tergantung kepadatan besarnya populasi lalat buah. Sebagai contoh misalnya penelitian yang dilakukan oleh Vargas et al., (1993) menyimpulkan bahwa kelimpahan B.arisanus secara jelas dideterminasi oleh kepadatan populasi lalat buah baik di habitat tumbuhan liar maupun di habitat tanaman budidaya.
Bautista dan Harris (1996) menyatakan, bahwa parasitoid akan tertarik pada buah inang yang menjadi preferensinya tanpa menghiraukan ada atau tidaknya telur atau larva lalat buah di dalamnya. Tanaman mempengaruhi kecocokan serangga inang melalui bagian tanaman yang sesuai, secara langsung akan mempengaruhi juga dinamika populasi parasitoid (Vet, 1999). Parasitoid soliter mampu mendeteksi adanya larva lalat buah yang telah mengandung dan tidak mengandung telur parasitoid spesies yang sama oleh karena adanya perubahan “homocoel” jaringan inang. Wong dan Ramadhan (1987) menyatakan bahwa parasitoid betina tidak dapat mendeteksi larva lalat buah yang mati.
Usia dan kondisi larva lalat buah sangat berpengaruh pada persentase kemunculan imago dan seks rasio parasitoid. Larva lalat buah yang berukuran besar pada umumnya menghasilkan persentase kemunculan parasitoid (Diachasma longicaudata dan D.tyoni) yang lebih tinggi daripada parasitoid yang dihasilkan larva lalat buah yang berukuran lebih kecil. Larva inang (lalat buah) yang besar juga akan menghasilkan persentase individu betina parasitoid (D.
longicaudata dan D.tryoni) yang lebih tinggi daripada individu jantan kedua parasitoid tersebut (Messing et al., 1993). Parasitoid secara fakultatif mengubah seks rasio turunannya sebagai tanggapan atas perubahan lingkungan (Kirby and Spence, 1816 dalam Godfray, 1994).
Peningkatan proporsi parasitoid jantan dipicu oleh stimuli lingkungan yang “hidden additive genetik varience” di dalam populasi Ukuran inang, kualitas pakan, kerapatan inang mungkin mempunyai pengaruh kuat pada rasio seks parasitoid hymenoptera.
2.1.8 Persebaran Lalat Buah
Lalat buah sebagai hama telah diketahui sejak tahun 1920, dan telah dilaporkan menyerang mangga di Pulau Jawa. Pada tahun 1938, lalat buah juga dilaporkan menyerang cabai, jambu, belimbing dan sawo. Lalat buah di Indonesia bagian barat dilaporkan sudah menyebar B. albistrigata, B. carambolae, B.
cucurbitae, B. papayae, B. tau, B. umbrosa, dan D. longicornis yang merupakan hama penting (Orr 2002). Menurut Vijaysegaran dan Drew (2006), B. albistri gata, B. carambolae, B. cucurbitae, B. occipitalis, B. papayae, B. philippinensis, dan B. umbrosa, adalah spesies yang sudah menyebar luas di Asia Tenggara dengan populasi sangat tinggi.
Menurut White dan Hancock (1997), daerah sebar lalat buah sudah hampir terdapat di seluruh belahan dunia. Daerah sebarannya antara lain: Australia (P.
Chrismas), Vanuatu, Indonesia (Sumatera, Jawa, Sulawesi, Sumbawa, Lombok, Maluku, Flores, Kalimantan), Malaysia, Singapore, Brunei, Taiwan, Hong Kong, Thailand, Laos, Vietnam, India (P. Andaman), Sri Lanka, Myanmar, China, Pulau Bagian Selatan Jepang, Indian Oceania, Afrika, Timur Tengah, Eropa, Guiana Perancis, Surinam, Amerika Utara, California, Laut pasifik, dan Palau.
Pertamakali dilakukan penelitian pada tahun 1985 oleh Hardy dan petugas karantina tumbuhan, ditemukan ± 66 spesies lalat buah (Dacus spp.) di Indonesia.
Periode 1992-1994, survei lalat buah dilakukan oleh Pusat Karantina Pertanian, ditemukan ± 47 spesies dari 66 spesies yang pernah ditemukan. Dari spesies yang telah ditemukan 20 diantaranya termasuk dalam grup Bactrocera dorsalis complex (Drew, 1994).
2.2 Pengaruh Tanaman Inang Terhadap Perilaku Serangga
Serangga dalam menentukan pilihan terhadap tanaman inang sangat dipengaruhi oleh banyak faktor terutama faktor nutrisi yang terkandung dalam tanaman inang tersebut. Tanaman mengandung 13 nutrisi mineral elemen yang sangat berfungsi untuk pertumbuhannya. Nutrisi tersebut dapat digolongkan ke dalam dua kelompok besar, yaitu makronutrisi dan mikronutrisi. Makronutrisi terdiri dari Nitrogen (N), Phosphor (p), Kalsium (Ca), Magnesium (Mg) dan Sulfur (S), sedangkan yang termasuk dalam mikronutrisi adalah Besi (Fe), Tembaga (Cu), Zeng (Zn), Boron (B), Molebdenum (Mo) dan Klorin (Cl) (Motavalli et. al. 2005). Nutrisi yang terkandung pada tanaman selain dibutuhkan dalam pertumbuhan dan perkembangan tanaman, juga sangat dibutuhkan oleh serangga untuk perkembangan hidupnya.
Sifat atraktan tanaman inang terhadap serangga sangat dipengaruhi oleh kandungan nutrisi tanaman inang yang sangat menentukan jumlah protein di dalam tanaman inang.konsentrasi protein juga sangat ditentukan oleh tipe tanaman, umur dan kandungan nutrisi tanah. Daun tanaman merupakan bagian tanaman yang sangat disukai oleh serangga karena memiliki nutrisi makanan paling baik dibandingkan dengan bagian tanaman lainnya, salah satu nutrisi tanaman yang utama dibutuhkan oleh serangga adalah Nittrogen. Nitrogen dalam bentuk protein dan asam amino sangat penting untuk pertumbuhan, perkembangan dan reproduksi serangga. Tanaman yang banyak mengandung asam amino dapat meningkatkan kemampuan reproduksinya. Nutrisi tanaman paling banyak terdapat pada jaringan tanaman yang lebih muda dibandingkan dengan jaringan tanaman yang sudah tua. Bunga, buah dan daun tanaman mengandung 1-5% atau lebih
Nitrogen, sedangkan pada batang pembuluh floem mengandung 0,5% Nitrogen dan xylem hanya 0,1% Nitrogen (Cloyd, 2005).
Chapman (1971) mengemukakan bahwa makanan sangat berperan terhadap perkembangbiakan serangga terutama terhadap keperidian serangga betina. Tobing (1996) menyatakan bahwa menurunnya kondisi nutrisi tanaman dengan bertambahnya umur berkaitan dengan perubahan-perubahan dalam komposisi asam-asam amino pada tanaman. Hal tersebut di atas menunjukkan bahwa kandungan nutrisi sangat menentukan preferensi serangga terhadap tanaman inang, baik untuk makanan maupun meletakkan telur.
Menurut Doutt (1959) terdapat empat tahapan yang harus dilewati agar parasitoid berhasil memarasit inangnya yaitu: 1) penemuan habitat inang, 2) penemuan inang, 3) penerimaan inang dan 4) kesesuaian inang. Penemuan inang terutama oleh parasitoid dipandu oleh rangsangan kimia yang berasal dari senyawa-senyawa volatile. Rangsangan tersebut daoat berupa bau yang berasal dari makanan atau tanaman yang terluka atau yang rusak, organisme yang berasosiasi dengan inang atau inang itu sendiri. Tanaman merupakan syarat utama karena tanaman mempunyai peran yang dominan dalam mendukung suatu habitat yang khas, akibatnya ssuatu parasitoid kadang-kadang tertarik pada tanaman tertentu meskipun di tempat tersebut tidak terdapat inang. Parasitoid kadang- kadang juga memarasit inang yang terdapat pada jenis tanaman tertentu dan tidak pada jenis tanaman yang lain (Vinson, 1981).
Penemuan inang oleh parasitoid dipandu oleh rangsangan fisik dan kimia.
Rangsangan fisik yang berperan terutama suara dan gerakan. Rangsangan kimia dapat dibagi menjadi 2 kelompok. Pertama, rangsangan kimia yang dapat diterima
dari jarak jauh misalnya bau inang. Rangsangan yang diterima memungkinkan parasitoid untuk melokalisasi areal pencarian inang. Kedua, rangsangan kimia yang dapat dideteksi hanya dari jarak dekat yaitu setelah terjadi kontak fisik.
Rangsangan ini biasanya berasal dari senyawa-senyawa padat atau cair misalnya kotoran inang, sekresi dari kelenjar labium inang, produk inang lain dan bekas parasitoid lain. Adanya rangsangan ini memungkinkan terjadinya kontak antara parasitoid dengan inangnya yang dicirikan oleh perilaku pengujian oleh parasitoid berupa pergerakan memutar dengan cepat dan perubahan kecepatan pergerakan.
Faktor lain yang ikut menentukan penemuan inang adalah pengalaman dan perilaku orientasi parasitoid.
Penerimaan inang atau pengenalan inang adalah proses diterima atau ditolak inang untuk peletakan telur setelah terjadi kontak (Arthur, 1981). Proses tersebut dibagi dalam empat fase yaitu : 1) kontak dan pemeriksaan, 2) penusukan dengan ovipositor, 3) pemasukan ovipositor dan 4) peletakan telur. Keempat fase tersebut harus lengkap dan berurutan sehingga bila terjadi hambatan pada salah satu fase, proses dimulai kembali dari awal. Penerimaan inang juga dipandu terutama oleh rangsangan fisik dan kimia selain itu, pengalaman parsitoid sebelumnya termasuk tempat perkembangan parsitoid juga berpengaruh pada proses penerimaan inang. Rangsangan fisik yang berperan adalah kondisi fisik inangnya seperti ukuran, bentuk, tekstur atau bentuk permukaan, warna dan kandungan air. Rangsangan lainnya adalah pergerakan inang misalnya kegiatan makan inang dan perkembangan embrio dalam telur. Rangsangan kimia dapat berasal dari senyawa-senyawa yang terdapat di luar dan di dalam tubuh inang yang dapat dideteksi dengan antenna, tarsi atau ovipositor. Senyawa-senyawa