.
KEANEKARAGAMAN BAKTERI PELARUT FOSFTA DARI KAWASAN MANGROVE
WONOREJO DENGAN PENDEKATAN TAKSONOMI NUMERIK FENETIK
Enny Zulaika1 dan Dyan Novita Rositawati1 1Institut Teknologi Sepuluh Nopember, Surabaya *Email: enny@bio.its.ac.id; Telp/fax: 0315963857
________________________________________________________________________________________________
ABSTRAK
Kawasan mangrove Wonorejo merupakan lahan basah di pantai timur Surabaya. Umumnya lahan mangrove merupakan habitat dari berbagai mikroorganisme dekomposer termasuk bakteri sehingga kaya akan bahan organik dari hasil dekomposisinya. Keanekaragaman bakteri dapat dikaji dengan pendekatan taksonomi numerik fenetik. Beberapa bakteri tanah dapat melarutkan fosfat, dimana fosfat terlarut merupakan salah satu parameter bahwa bakteri pelarut fosfat dapat digunakan sebagai bahan baku biofertilizer. Tujuan penelitian adalah mengetahui kenekaragaman bakteri pelarut fosfat dari kawasan mangrove Wonorejo Surabaya dengan pendekatan taksonomi numerik fenetik. Isolasi bakteri dilakukan dengan metode komposit. Seleksi bakteri pelarut fosfat menggunakan media selektif Pykovskaya’s agar. Karakterisasi isolat terdiri dari morfologi koloni dan sel, uji fisiologi dan reaksi biokimia. Pendekatan taksonomi numerik fenetik menggunakan indeks similaritas dengan algoritma UPGMA dan konstruksi dendogram. Isolat terseleksi yang dapat melarutkan fosfat sebanyak 8 isolat, berdasarkan konstruksi dendogram terbentuk 3 klaster yaitu klaster A, B, dan C dengan indeks similaritas 59 - 90%.
Kata Kunci : pelarut-fosfat, similaritas, taksonomi-numerik, Wonorejo
PENDAHULUAN
Kawasan mangrove merupakan suatu ekosistem peralihan yang kompleks, terletak di daerah pasang surut, mempunyai interaksi antara tanaman, hewan dan mikroorganisme. Pohon-pohon mangrove menghasilkan sejumlah seresah seperti daun, bunga, dan ranting-ranting sehingga merupakan habitat yang ideal bagi banyak detritivor termasuk mikroorganisme pendegradasi seresah seperti jamur dan bakteri (Vanmathi & Sivakumar, 2013). Kemampuan bakteri mendegradasi seresah mangrove akan menghasilkan molekul sederhana yang ikut berperanan dalam siklus biogeokimia, di antaranya siklus karbon, nitrogen, fosfor dan sebagainya (Mendes & Tsai, 2014). Lingkungan di ekosistem mangrove sangat bervariasi, mempunyai salinitas yang fluktuatif, perubahan temperatur yang relatif cepat, kadar oksigen yang ekstrim rendah, hal tersebut menjadikan kondisi lingkungan mangrove mempunyai keanekaragaman organisme termasuk bakteria yang relatif tinggi. Umumnya mikroorganisme di kawasan ekosistem mangrove memiliki bermacam-macam potensi dan peranan penting terhadap perbaikan lingkungan, disamping ikut berperanan dalam siklus biogeokimia (Mendes & Tsai, 2014).
Keanekaragaman bakteri dapat ditinjau dari morfologi, fisiologi, genetika maupun molekularnya. Beberapa bakteri dengan morfologi yang sama dapat memiliki fisiologi atau reaksi biokimia yang berbeda dan sebaliknya. Keanekaragaman dapat dikaji melalui pendekatan taksonomi yaitu taksonomi numerik fenetik. Menurut Sahoo dan Dhal (2009) beberapa genus bakteri yang berasal dari ekosistem mangrove mempunyai potensi dapat melarutkan fosfat, genus tersebut adalah Bacillus, Azotobacter dan
Pseudomonas. Di kawasan mangrove Wonorejo yang memproduksi serasah sampai 4,5 ton/ha/tahun
(Sopana dkk., 2013), merupakan kawasan yang kaya akan bahan organik. Apakah di kawasan mangrove tersebut juga akan didapatkan isolat bakteri yang dapat melarutkan fosfat dengan suatu pendekatan taksonomi numerik fenetik.
.
METODE PENELITIAN Isolasi
Pengambilan tanah sebagai sumber inokulum dilakukan dengan metode komposit. Satu gram sampel tanah dimasukkan ke dalam 9 ml aquadest steril selanjutnya dilakukan pengenceran sampai 10-3. Dengan metode cawan tuang, sebanyak 100 µL diinokulasikan ke dalam medium agar Pykovskaya dengan komposisi glukosa 10 g; Ca3(PO4)2 5 g; (NH4)2SO4 0,5 g; NaCl 0,2 g; MgSO4.7H2O 0,1 g; KCl 0,2 g; ekstrak yeast 0,5 g; MnSO4.H2O 0,002 g; FeSO4.7H2O 0,002 g, dan akuades sampai dengan 1000 ml (Zulaika dkk, 2015). Diinkubasi pada suhu ruang selama 24 jam, koloni yang tumbuh diindikasikan sebagai bakteri pelarut fosfat endogenik dari kawasan mangrove Wonorejo Surabaya.
Purifikasi
Koloni yang tumbuh dimurnikan dengan metode 16 gores pada medium nutrien agar, diinkubasi pada suhu ruang ± 24 jam, jika bentuk koloni sudah seragam dianggap koloni sudah murni. Validasi kemurniannya dilakukan dengan melihat bentuk sel menggunakan pewarnaan sederhana methylene blue yang diamati di bawah mikroskop (Harley & Prescott, 2002). Jika belum diperoleh bentuk sel yang seragam maka dilakukan pemurnian ulang dengan metode 16 gores sampai diperoleh bentuk sel yang seragam.
Seleksi Isolat
Isolat yang telah murni selanjutnya diseleksi kemampuannya melarutkan fosfat secara kualitatif. Metode yang digunakan adalah inokulasi titik pada medium Pikovskaya agar. Kultur diinkubasi pada suhu ruang diamati setiap 24 jam. Koloni yang tumbuh dan membentuk zona bening di sekitar koloni merupakan isolat yang mampu melarurkan fosfat. Koloni yang dipilih adalah koloni yang mempunyai zona bening relatif lebar dibanding isolat lainnya (Zulaika dkk, 2015).
Karakterisasi Fenotipik
Data fenotipik, respon fisiologis dan reaksi biokimiawi dari hasil karakterisasi dikonversikan dengan nilai positif (+) atau negatif (-). Data ditabulasikan ke dalam matrik n x t dan dimasukkan ke dalam PFE atau
Programer File Editor (Moore et al., 2010), Pengolahan data
Matrik n x t diedit dengan PFE dan danalisis dengan MVSP. Untuk Operational Taxonomical Unit (OTU), setiap isolat dikode dengan A, B, C dan seterusnya, untuk karakter diberi kode AA, AB, AC dan seterusnya sesuai dengan banyaknya karakter yang digunakan. Penghitungan nilai similaritas antar isolat digunakan Simple Matching Coeficient (SSM). Pengelompokan dilakukan dengan algoritma Unweighted
Paired Group Methode with Arithmatic Averages (UPGMA) dalam bentuk dendogram (Sneath & Sokal, 1973
dalam Zulaika et al., 2013). Dendogram yang didapatkan kemudian diedit sesuai dengan skala yang tepat sehingga dihasilkan dendogram yang representatif sesuai dengan nilai masing-masing titik (Backeljau et al., 1996).
HASIL DAN PEMBAHASAN Isolat pelarut fosfat
Isolat yang mampu melarutkan fosfat sebanyak 8, ditandai dengan terbentuknya zona pelarutan fosfat di sekitar koloni. Isolat tersebut adalah isolat BW 3, BW 5, BW 6, BW 8, BW 10, BW 10a, BW 10 b, dan BW 11. Koloni dan zona pelarutan fosfat dapat dilihat pada Gambar 1.
.
Gambar 1. Isolat dengan zona pelarutan fosfat
Medium Pikovskaya padat berwarna putih keruh karena mengandung fosfat terikat yaitu Ca3(PO4)2. Fosfat yang terlepas dalam medium akan ditunjukkan dengan terbentuknya zona bening yaitu zona pelarutan fosfat (Rahardjo et al., 2007). Zona pelarutan fosfat yang semakin lebar dan semakin jernih secara kualitas menunjukkan kelarutan fosfat yang semakin tinggi. Menurut Khiari & Parent (2005), bakteri pelarut fosfat dapat melarutkan fosfat karena menghasilkan asam organik. Asam organik dihasilkan melalui proses metabolisme glukosa dalam siklus asam trikarboksilat (TCA) yang merupakan kelanjutan dari reaksi glikolisis. Menurut Chen et al. (2006), asam organik mampu berikatan dengan ion Ca dari Ca3(PO4)2 dan membebaskan H2PO4 sehingga membentuk area yang berwarna jernih dan mengakibatkan fosfat terikat menjadi bentuk yang tersedia.
Konstruksi Dendogram
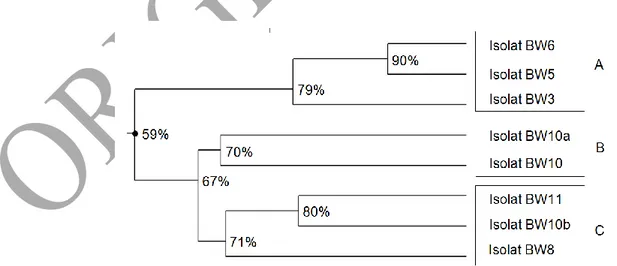
Isolat bakteri pelarut fosfat yang terseleksi, selanjutnya dikarakterisai bentuk koloni dan selnya, tipe pertumbuhan koloni pada media, sifat Gram, keberadaan spora, kista dan kapsul, kebutuhan oksigen, metabolisme karbohidrat, resistensi terhadap kadar garam dan antibiotik, kemampuan melarutkan fosfat, semuanya berjumlah 74 karakter. Konstruksi dendogram terhadap 8 isolat menghasilkan 3 kelompok isolat berdasarkan karakter di atas yaitu A, B, dan C dengan indeks similaritas antara 59 – 90 % (Gambar 2).
Gambar 2. Dendogram yang menunjukkan similaritas di antara 8 isolat bakteri pelarut fosfat
(angka menunjukkan tingkat similaritas)
Nilai koefisien korelasi fenetik pada konstruksi dendogram adalah 90%. Menurut Sembiring (2011), jika nilai koefisien korelasi fenetik ≥ 70% maka diasumsikan dendogram yang terbentuk semakin mewakili matriks similaritas yang menjadi dasar konstruksinya.
.
Berdasarkan Gambar 1, kelompok A beranggotakan 3 isolat yaitu BW 3, BW5, dan BW 6 dengan indeks similaritas 79-90%, kelompok B beranggotakan BW 10 dan BW 10a dengan indeks similaritas 70% dan kelompok C beranggotakan BW 3, BW 8, dan BW 11 dengan indeks similaritas 71-80%. Menurut Goodfellow & O´Donnell (1993 ) dalam Zulaika et al., (2013), apabila nilai similaritas isolat satu dengan isolat lain ≥70% maka berdasarkan konsep takso spesies isolat tersebut dapat dimasukkan ke dalam spesies yang sama. Namun untuk menentukkan spesies yang sama dengan strain sama atau berbeda diperlukan uji karakterisasi molekular dengan marka 16S rRNA (Woose, 1987 dalam Zulaika et al., 2013).
Isolat pada kelompok A mempunyai karakter mampu melarutkan fosfat, dapat mendegradasi senyawa organik karbohidrat dan lipid, mampu menggunakan senyawa organik seperti asam sitrat sebagai sumber energi, mampu tumbuh pada kadar NaCl 3%, dan resisten terhadap kloramfenikol dan tetrasiklin 30 μg. Isolat pada kelompok B mempunyai karakter mampu melarutkan fosfat, dapat mendegradasi protein seperti gelatin dan kasein, mampu menggunakan senyawa organik sebagai sumber energi, mampu tumbuh pada kadar NaCl 6,5%, resisten terhadap kloramfenikol dan ampisilin 30 μg. Sedangkan isolat pada mempunyai karakter mampu melarutkan fosfat, dapat menghidrolisis karbohidrat dan protein, resisten terhadap antibiotik ampisilin, kloramfenikol, dan tetrasiklin 30 μg.
KESIMPULAN
Delapan isolat yang terseleksi mampu melarutkan fosfat, mendegradasi senyawa organik karbohidrat, lipid dan protein, serta resisten terhadap salinitas tinggi dan beberapa antibiotik.
UCAPAN TERIMAKASIH
Penelitian ini didukung dengan dana BOPTN Institut Teknologi Sepuluh Nopember Surabaya, nomor kontrak: 01711/IT2.11/PN.08/2016
DAFTAR PUSTAKA
Backeljau, T. & Winnepenninckx, B. 1996. 18S rRNA alignments derived from different secondary structure
models can produce alternative phylogenies. J. Zool. Syst. Evol. Res., 34: 135-143.
Chen Y.P., Rekha P.D., Arunshen A.B., Lai W.A. & Young C.C. 2006. Phosphate solubilizing bacteria from
subtropical soil and their tri-calcium phosphate solubilizing abilities. Appl. Soil Ecol. 34: 33-41.
Harley, J.P. & Prescott L.M. 2002. Laboratory Exercises in Microbiology. Fifth Edition. USA : The McGraw−Hill Companies.
Khiari, L. & Parent,L.E. 2005. Phosphorous Transformation In Acid-Textured Soils Treated With Dry Swine
Manure. Canadian Journal of Soil Sciene, 85: 75-87.
Mendes, L.W. & Tsai, S.M. 2014. Variations of Bacterial Community Structure and Composition in Mangrove
Sediment at Different Depths in Southeastern Brazil. Journal Diversity, 6: 827-843.
Moore, E.R.B., Mihaylova S.A, Vandamme, P. & Krichevsky M.I. 2010. Microbial Systematics and
Taxonomy: Relevance for a Microbial commons. Lenie Dijkshoorn Research in Microbiology, 161: 430-438.
Raharjo, B., Suprihadi, A. & Agustina D.K. 2007. Pelarutan Fosfat Anorganik oleh Kultur Campur Jamur
Pelarut Fosfat Secara In Vitro. Jurnal Sains & Matematika (JSM), 15 (2): 12-18.
Sahoo, K. & Dhal, N.K. 2009. Potential microbial diversity in mangrove ecosystem. A review. Indian journal of marine sciences, 38(2): 249-256.
Sembiring, L. 2011. Sistemtika Mikrobia. Yogyakarta: Laboratorium Mikrobiologi, Universitas Gajah Mada. Sopana, A.B., Widyaleksono, T. & Soedarti, T. 2013. Produktivitas Serasah Mangrove di Kawasan
Wonorejo Pantai Timur Surabaya. Jurnal Ilmiah Biologi, 1 (1); 6-10.
Vanmathi, S.K. & Sivakumar, T. 2013. Isolation, identification and Characterization of Fusarium species
.
Zulaika, E. 2013. Eksplorasi Bakteri Resisten Merkuri (BRM) Endogenik Kalimas Surabaya yang Berpotensi
Sebagai Kandidatus Agensia Bioremediasi Pencemar Merkuri. Disertasi. Fakultas Sains & Teknologi.
Universitas Airlangga. Surabaya.
Zulaika, E., Shovitri, M. & Alami, H. 2015. Konsorsium Azotobacter Sebagai Agensia Bioremediasi Lahan
Tercemar Merkuri Sekaligus Sebagai Bahan Baku Biofertilizer Yang Ramah Lingkungan, LPPM ITS,