STUDI PEWARISAN SIFAT TOLERANSI ALUMINIUM
TANAMAN SORGUM MANIS [Sorghum bicolor (L.) Moench]
ISNAINI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2010
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis dengan judul Studi Pewarisan
Sifat Toleransi Aluminium Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench] adalah karya saya dengan arahan dari komisi pembimbing dan belum
diajukan dalam bentuk karya apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, Februari 2010
ISNAINI NIM A 253070051
ABSTRACT
ISNAINI. Study of Aluminum Tolerance Inheritance of Sweet Sorghum
[Sorghum bicolor (L.) Moench]. Under direction of
TRIKOESOEMANINGTYAS and DESTA WIRNAS.
Sweet sorghum [Sorghum bicolor (L.) Moench] is actually not Indonesian origin but it has a big potential to be grown and cultivated in this country as the solution to food and energy crisis owing to its wide adaptability and other advantages. Al toxicity is one of limiting factors in agriculture development in Indonesia. Further research on sorghum breeding is needed especially to search for genotypes that can be grown and cultivated in acid soil with Al toxicity. The successes in the sorghum breeding program to obtain adaptable varieties in Al stress is determined by selecting the appropriate breeding method. The objective of this research was to study inheritance of Al tolerance of sweet sorghum by analysis of F2 distribution, the genetic components and heritability of the four generation of sorghum to determine the effective and efficient method in breeding program for sorghum tolerance to Al. The four populations in this research were UPCA S1 (P1, susceptible parent) and Numbu (P2, tolerant parent), F1 and F1 reciprocal and F2. This research was also to develop selection criteria for Al tolerance in sorghum breeding and selection based on yield character for food and bio-ethanol. The crosses of parental, F1, F1R and F2 generation were established in University Farm of IPB and UPTD Tenjo from June 2008 to June 2009. The study of Al tolerance inheritance in nutrient culture was conducted in green house of University Farm of IPB from July-August 2009. The study of Al tolerance inheritance in field was conducted in UPTD Tenjo from July-November 2009. Selection was applied to 600 genotypes of F2 generation. Based on reciprocal analysis, all of characters are controlled by nuclear genes and their inheritance in not affected by maternal cytoplasm. The distribution of F2 genotype is indicating continuous with skewnesses which indicate that all characters are polygenics. All characters of Al tolerance in seedling stage i.e. root length, root and shoot dry weight were controlled by additive gene action with contribution by complementary epistasis gene except shoot length that is controlled by additive gene action with duplicate epistasis gene. Plant height in field experiment is controlled by additive gene action. Plant weight, total of biomass weight, ear length and total of grain weight are controlled by additive gene action with complementary epistasis gene. In F2 generation, there are transgresive segregants. All characters had broad sense heritability from medium to high. Based on heritability value, pedigree selection is the most effective selection method in breeding program for sorghum tolerance to Al. Multi-characters selection is more effective in F2 selection both for food and bio-ethanol.
Keywords: Sorghum, inheritance, quantitative traits, Al tolerance, differential selection
RINGKASAN
ISNAINI. Studi Pewarisan Sifat Toleransi Aluminium Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench]. Dibimbing oleh TRIKOESOEMANINGTYAS dan DESTA WIRNAS.
Sorgum manis [Sorghum bicolor (L.) Moench] merupakan salah satu jenis tanaman yang sangat potensial dalam pemecahan masalah krisis pangan dan energi. Sorgum manis ideal untuk dikembangkan di Indonesia karena tanaman ini mempunyai kesesuaian yang tinggi untuk dikembangkan di lahan kering. Namun lahan kering di Indonesia didominasi oleh tanah yang bereaksi masam. Karena itulah perlu dilakukan program pemuliaan untuk memperoleh varietas yang mampu beradaptasi terhadap cekaman Al ditentukan oleh pemilihan metode pemuliaan yang tepat sehingga kegiatan pemuliaan dapat menjadi efektif dan efisien. Sampai sejauh ini informasi mengenai pewarisan toleransi keracunan Al pada tanaman sorgum manis masih sangat sedikit. Tujuan penelitian ini adalah untuk memperoleh informasi mengenai pewarisan sifat toleransi Al pada tanaman sorgum manis. Tujuan lainnya adalah memperoleh karakter untuk seleksi pemuliaan sorgum yang toleran Al serta memperoleh genotipe-genotipe toleran Al melalui seleksi pada generasi F2 di tanah masam. Percobaan dimulai dengan pembentukan populasi melalui persilangan antara tetua peka cekaman Al (UPCA S1) dan tetua toleran cekaman Al (Numbu) sejak bulan Juni 2008-Juli 2009 di KP Cikabayang University Farm (UF) IPB dan UPTD Lahan Kering Tenjo, Dinas Pertanian dan Kehutanan Kabupaten Bogor. Studi pewarisan sifat toleransi Al pada karakter stadia bibit pada kondisi tercekam Al dengan menggunakan metode kultur hara, dilakukan rumah kaca KP Cikabayan, UF IPB sejak bulan Juli 2009-Agustus 2009. Studi pewarisan karakter agronomi dan hasil pada kondisi tercekam Al di lapang dilaksanakan di UPTD Tenjo sejak bulan Juli 2009-November 2009. Seleksi genotipe sorgum toleran Al dilakukan pada populasi F2 yang ditanam pada percobaan lapang. Hasil analisa uji-t pada populasi F1 dan F1R diketahui bahwa pada kondisi cekaman Al, gen yang mengendalikan karakter fase bibit, karakter agronomi dan karakter hasil terdapat pada gen-gen inti dan pewarisannya tidak dipengaruhi tetua betina. Analisa sebaran frekuensi genotipe F2 menunjukkan bahwa semua karakter pengendali toleransi tanaman sorgum terhadap cekaman Al bersifat poligenik. Semua karakter toleransi Al yang berhubungan dengan stadia bibit yaitu panjang akar, bobot kering akar dan bobot kering tajuk dikendalikan aksi gen aditif dengan pengaruh gen epistasis komplementer kecuali pada karakter panjang tajuk yang memiliki aksi gen aditif dengan pengaruh gen epistasis duplikasi. Karakter agronomi dan hasil hanya karakter tinggi tanaman yang dikendalikan oleh aksi gen aditif. Karakter bobot batang, bobot total biomassa, panjang malai dan bobot biji per tanaman terdapat aksi gen aditif dengan pengaruh aksi gen epistasis komplementer. Hasil analisa sebaran frekuensi genotipe F2 juga menunjukkan terdapat keragaman yang besar pada populasi F2 yang merupakan zuriat hasil persilangan dari tetua genotipe toleran Al dan genotipe peka Al dan terdapat terdapat segregan transgresif pada semua karakter-karakter yang diuji dengan nilai melebihi nilai kedua tetua. Nilai
heritabilitas arti luas untuk semua karakter toleransi Al yang berhubungan dengan pertumbuhan akar maupun karakter agronomi cekaman Al tergolong sedang hingga tinggi sehingga metode seleksi yang dapat digunakan dalam seleksi galur sorgum dengan toleransi terhadap Al dapat menggunakan metode pedigree sebagai metode seleksi yang paling efektif. Seleksi pada populasi F2 untuk keperluan pangan menghasilkan nilai diferensial seleksi yang lebih tinggi jika seleksi dilakukan berdasarkan bobot biji per tanaman dan tinggi tanaman dibandingkan jika hanya berdasarkan bobot biji per tanaman. Seleksi pada populasi F2 sebagai bahan bioetanol menghasilkan nilai diferensial yang lebih tinggi apabila seleksi dilakukan berdasarkan kedua karakter bobot biji per tanaman dan bobot batang segar secara bersamaan dibandingkan dengan seleksi yang dilakukan dengan menggunakan karakter tunggal.
Kata Kunci: Sorgum, pewarisan karakter kuantitatif, toleransi Al, diferensial seleksi
© Hak Cipta Milik IPB, tahun 2010
Hak Cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan sebagian pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh Karya tulis dalam bentuk apa pun tanpa izin IPB
STUDI PEWARISAN SIFAT TOLERANSI ALUMINIUM
TANAMAN SORGUM MANIS [Sorghum bicolor (L.) Moench]
ISNAINI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2010
Judul Tesis : Studi Pewarisan Sifat Toleransi Aluminium Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench]
Nama : ISNAINI
NRP : A 253070051
Disetujui Komisi Pembimbing
Dr. Ir. Trikoesoemaningtyas, MSc. Dr. Desta Wirnas, SP. MSi.
Ketua Anggota
Diketahui
Ketua Mayor Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, MSc. Prof. Dr. Ir. Khairil A. Notodiputro, MS.
PRAKATA
Puji syukur penulis panjatkan ke hadirat Allah SWT atas segala karunia-Nya sehingga penelitian dan penulisan tesis dengan judul Studi Pewarisan Sifat Toleransi Aluminium Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench] ini dapat diselesaikan dengan baik. Tesis ini merupakan tugas akhir sebagai salah satu syarat untuk memperoleh gelar Master Sains di Sekolah Pascasarjana Institut Pertanian Bogor. Penelitian dan penulisan tesis ini dapat diselesaikan berkat bantuan dari Direktorat Jenderal Pendidikan Tinggi, Departemen Pendidikan Nasional Republik Indonesia melalui beasiswa BPPS tahun 2007-2009 dan Hibah Penelitian Tim Pascasarjana-HPTP tahun 2008-2010.
Penulis mengucapkan terima kasih yang tulus dan penghargaan yang tidak terhingga kepada:
1. Tim BPPS, Dirjen Pendidikan Tinggi, Departemen Pendidikan Nasional RI yang telah memberikan beasiswa kepada penulis untuk mengikuti program Master di IPB.
2. Dr. Ir. Trikoesoemaningtyas, MSc. selaku Ketua Komisi Pembimbing atas bimbingan dan perhatian selama penulis menempuh pendidikan di IPB dan selama pelaksanaan penelitian serta penulisan tesis ini.
3. Dr. Desta Wirnas, SP. MSi. selaku Pembimbing Tesis Kedua yang telah memberikan bimbingan dan pengarahan selama kegiatan penelitian dan penulisan tesis.
4. Prof. Dr. Ir. Sriani Sujiprihati, MS. selaku Kepala Bagian Laboratorium Genetika dan Pemuliaan Tanaman, Departemen Agronomi dan Hortikultura IPB, yang telah memberikan kesempatan yang sangat berharga dan rekomendasi kepada penulis untuk mendaftar di Sekolah Pascasarjana IPB. 5. Prof. Dr. Ir. Bambang S Purwoko, MSc., Prof. Dr. Ir. Supiandi Sabiham,
MAgr., Prof. Dr. Ir. MA Chozin, MAgr. dan Dr. Ir. Sobir, MSi. yang telah berkenan memberikan izin dan rekomendasi kepada penulis untuk mendaftar di Sekolah Pascasarjana IPB.
6. Dr. Ir. Yudiwanti WE Kusumo, MSi. sebagai penguji luar komisi pada saat ujian akhir tesis atas saran-saran untuk perbaikan tesis ini.
7. Kepala dan staf UPTD Lahan Kering Tenjo, Dinas Pertanian dan Kehutanan Kabupaten Bogor.
8. Dosen-dosen Departemen Agronomi dan Hortikultura IPB, yang telah memberikan ilmu dan pengetahuan tiada batas kepada penulis selama menempuh pendidikan di IPB.
9. Ayahanda Asmawi Bakrie dan Ibunda Asmarawati MA Bakrie, Abang Hendri Afrizal dan Dang Nur Asmi Hayati, SPdI, Abang Syafrizal, SH. dan Dedek Aidil Syah Putra atas semua doa, dukungan dan curahan kasih selama penulis menyelesaikan pendidikan.
10. Sahabat tersayang, Fifin N Nisya, SP. MSi., (semangat, Sir! biar kita lanjut ke project berikutnya), Genta Atmaja, SP. (atas sponsorship perbanyakan tesis), Novy Anggraini, SP. MM., Effi Noverya, SPi., Melinda T Wulan, SP., I Gst Ayu Dwi Putri Mayasari, SP., Rahdini Safitri, SSi. dan Yenni Rahmawati, SE. (my bfs forever!) atas semua kebersamaan dan dukungan.
11. Rekan-rekan satu tim HPTP (Ir. Sungkono, MP., Ir. Karlin Agustina, MSi., Sumiyati, SP. dan Rahmansyah Darmawan, SP.), rekan-rekan PBT 2007 (Heni Safitri, SP. MSi., Amin Nur, SP. MSi., Siti Noorohmah, SP. MSI., Nurwanita E Putri, SP. MSi., Yussi Arisandi, SP. MSI., Alfin Widiastuti, SP. MSi., Rokhana Faizah, SP. MSi. dan Hairin Dalimunthe, SP. MSi.) dan Ayunda Dian Novita, SP. atas semua kebersamaan dan bantuan.
12. Seluruh pihak yang tidak dapat penulis sebut satu per satu yang banyak membantu menyelesaikan studi dan penelitian penulis.
Akhir kata, penulis berharap semoga karya ilmiah ini dapat memperkaya keilmuan dan bermanfaat bagi semua.
Bogor, Februari 2010
RIWAYAT HIDUP
Penulis dilahirkan di Kota Bengkulu pada tanggal 17 Januari 1984 dari Bapak Asmawi Bakrie BA dan Ibu Asmarawati MA Bakrie. Penulis merupakan putri ketiga dari empat bersaudara.
Penulis menempuh pendidikan dasar di SD Negeri 20 Kota Bengkulu pada tahun 1990 -1996, dilanjutkan dengan SLTP Negeri 2 Kota Bengkulu (1996-1999) dan dilanjutkan ke SMU Negeri 2 Kota Bengkulu (1999-2002). Tahun 2002 penulis diterima di Program Studi Pemuliaan Tanaman dan Teknologi Benih, Departemen Agronomi dan Hortikultura Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI) dan lulus pada tahun 2007.
Penulis berkesempatan melanjutkan program master pada Mayor Pemuliaan dan Bioteknologi Tanaman, Sekolah Pascasarjana IPB pada tahun 2007 atas Beasiswa Pendidikan Pascasarjana dari Direktorat Pendidikan Tinggi Departemen Pendidikan Nasional Republik Indonesia. Karya ilmiah yang merupakan bagian dari tesis penulis telah dipresentasikan dalam Simposium dan Kongres Nasional VI Perhimpunan Ilmu Pemuliaan Indonesia (PERIPI) pada bulan November 2009 dengan Judul Genetic Variation, Heritability and Gene Action of Sorghum [Sorghum bicolor (L.) Moench] in Nutrient Solution.
DAFTAR ISI
Halaman
DAFTAR TABEL ……….……… xv
DAFTAR GAMBAR ……….……… xvii
DAFTAR LAMPIRAN ……….……… xix
PENDAHULUAN ……….……… 1
Latar Belakang ……….………. 1
Tujuan ……….……….…………. 4
Hipotesis ……… 4
TINJAUAN PUSTAKA ……… 6
Sorgum Manis [Sorghum bicolor (L.) Moench] ………... 6
Mekanisme Toleransi terhadap Aluminium ………. 8
Pemuliaan bagi Lingkungan Bercekaman ……… 11
Arah Program Pemuliaan Tanaman Sorgum ……… 13
Metode untuk Mempelajari Pola Pewarisan Sifat ……… 22
Pewarisan Toleransi terhadap Aluminium ……… 26
BAHAN DAN METODE ………. 28
Bahan Tanaman ……… 28
Waktu dan Tempat ……… 28
Pewarisan Karakter Toleransi Al pada Stadia Bibit ………. 28
Pewarisan Karakter Agronomi dan Hasil pada Kondisi Tercekam Al ……… 29
Seleksi Genotipe F2 Sorgum Toleran Cekaman Al ……….. 30
Analisis Data ………. 30
HASIL DAN PEMBAHASAN ………. 33
Pewarisan Karakter Toleransi Al pada Stadia Bibit …………... 34
Pengaruh Tetua Betina terhadap Karakter Toleransi Al pada Stadia Bibit ……….. 37
Sebaran Frekuensi Genotipe Generasi F2 Karakter Toleransi Al pada Stadia Bibit ………. 38
Pendugaan Komponen Ragam dan Nilai Heritabilitas Arti Luas Karakter Toleransi Al pada Stadia Bibit ………. 42
Pewarisan Karakter Agronomi dan Hasil pada Kondisi Tercekam Al ……… 45
Pengaruh Tetua Betina Karakter Agronomi dan Hasil ………….. 48
Sebaran Frekuensi Genotipe Generasi F2 Karakter Agronomi dan Hasil ……… 48 Pendugaan Komponen Ragam dan Nilai Heritabilitas Arti Luas Karakter Agronomi dan Hasil ………. 53
DAFTAR TABEL
Seleksi Genotipe F2 Sorgum Toleran Cekaman Al ……….. 55
KESIMPULAN DAN SARAN ………. 59
Kesimpulan ………... 59
Saran ………. 59
DAFTAR PUSTAKA ……… 60
Halaman
1 Rata-rata Tetua P1 (UPCA S1), P2 (Numbu), Generasi F1 dan F2
Hasil Persilangan Sorgum Varietas UPSA-S1 dan Numbu pada
Cekaman Aluminium di Kultur Hara ………. 35
2 Pendugaan Pengaruh Tetua Betina Hasil Persilangan Sorgum Varietas
UPCA-S1 dan Numbu pada Cekaman Aluminium di Kultur Hara …... 38
3 Nilai Skewness, Aksi Gen, Nilai Kurtosis Genotipe F2 Hasil
Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada Cekaman
Aluminium di Kultur Hara ………. 42
4 Nilai Pendugaan Komponen Ragam dan Heritabilitas Arti Luas Hasil
Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada Cekaman
Aluminium di Kultur Hara ………. 44
5 Rata-rata tetua P1 (UPCA S1), P2 (Numbu), F1 dan F2 Hasil
Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada Cekaman
Aluminium di Lapangan ………. 47
6 Pendugaan Pengaruh Tetua Betina Hasil Persilangan Sorgum Varietas
UPCA-S1 dan Numbu pada Cekaman Aluminium di Lapangan ……... 48
7 Nilai Skewness, Aksi Gen, Nilai Kurtosis Genotipe F2 Hasil
Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada Cekaman
Aluminium di Lapangan ………. 52
8 Nilai Pendugaan Komponen Ragam dan Heritabilitas Arti Luas Hasil
Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada Cekaman Al di Lapangan ……… 54
9 Kemajuan Seleksi Berdasarkan Karakter Bobot Biji per Tanaman
Hasil Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada
Cekaman Aluminium di Lapangan ……… 56 10 Kemajuan Seleksi Berdasarkan Karakter Bobot Biji per Tanaman
Hasil dan Tinggi Tanaman Persilangan Sorgum Varietas UPCA-S1
dan Numbu pada Cekaman Aluminium di Lapang an ………... 56 11 Kemajuan Seleksi Berdasarkan Karakter Bobot Batang Hasil
Persilangan Sorgum Varietas UPCA-S1 dan Numbu pada Cekaman Al di Lapangan ……… 57
12 Kemajuan Seleksi Berdasarkan Karakter Bobot Biji per Tanaman dan Bobot Batang Hasil Persilangan Sorgum Varietas UPCA-S1 dan
DAFTAR GAMBAR
1 Bagan Alir Penelitian Studi Pewarisan Toleransi Al Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench] ……….. 5
2 Penampilan UPCA S1 dan Numbu (kiri) dan F1/F1 Resiprokal Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu (kanan) pada
Cekaman Al di Kultur Hara ………... 36
3 Penampilan Generasi F2 Hasil Persilangan Sorgum Varietas UPCA S1
dan Numbu pada Cekaman Al di Kultur Hara ………... 36
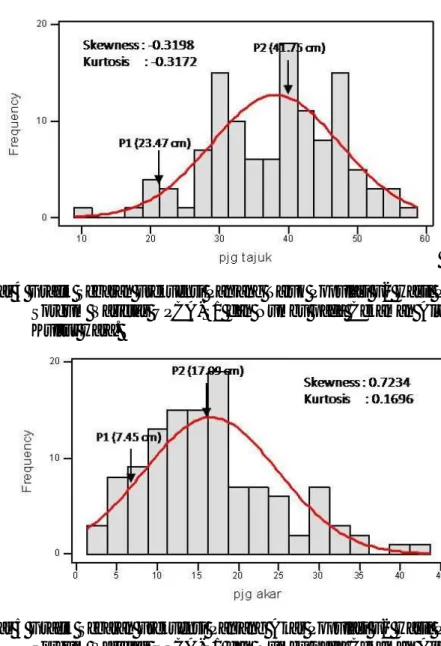
4 Grafik Sebaran Frekuensi Panjang Tajuk Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman
Aluminium di Kultur Hara ………. 40
5 Grafik Sebaran Frekuensi Panjang Akar Populasi F2 Hasil Persilangan
Sorgum Varietas UPCA S1 dan Numbu pada Cekaman Aluminium di Kultur Hara ………. 40
6 Grafik Sebaran Frekuensi Bobot Kering Tajuk Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman
Aluminium di Kultur Hara ………. 41
7 Grafik Sebaran Frekuensi Bobot Kering Akar Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman
Aluminium di Kultur Hara ………. 41
8 Penampilan Tanaman dan Malai UPCA S1 (P1) dan Numbu (P2) di
Lapangan ……… 46
9 Penampilan Tanaman dan Malai F1 dan F1 Resiprokal Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu di Lapangan …… 46
10 Penampilan Tanaman dan Malai F2 Hasil Persilangan Sorgum
Varietas UPCA S1 dan Numbu di Lapangan ………. 46
11 Grafik Sebaran Frekuensi Tinggi Tanaman Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman Al di Lapangan ……… 50 12 Grafik Sebaran Frekuensi Bobot Batang Populasi F2 Hasil Persilangan
Sorgum Varietas UPCA S1 dan Numbu pada Cekaman Al di
Lapangan ……… 50
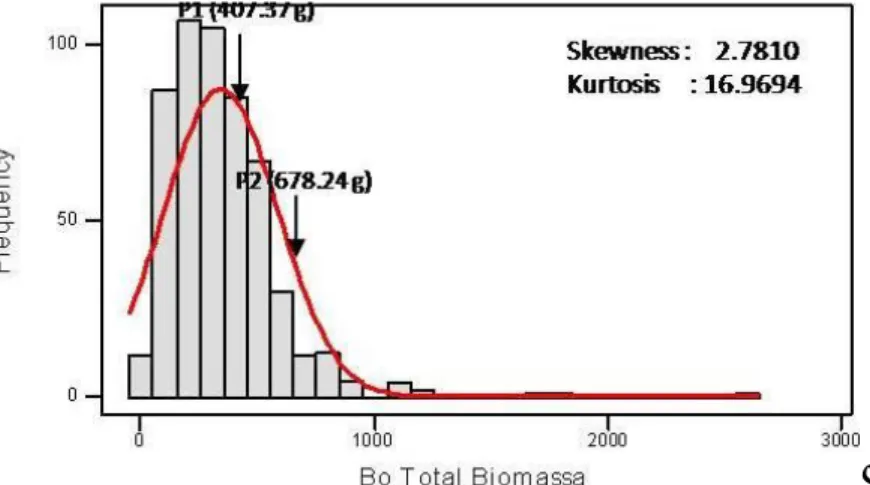
13 Grafik Sebaran Frekuensi Bobot Biomassa Total Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman Al di Lapangan ……… 51
14 Grafik Sebaran Frekuensi Panjang Malai Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman
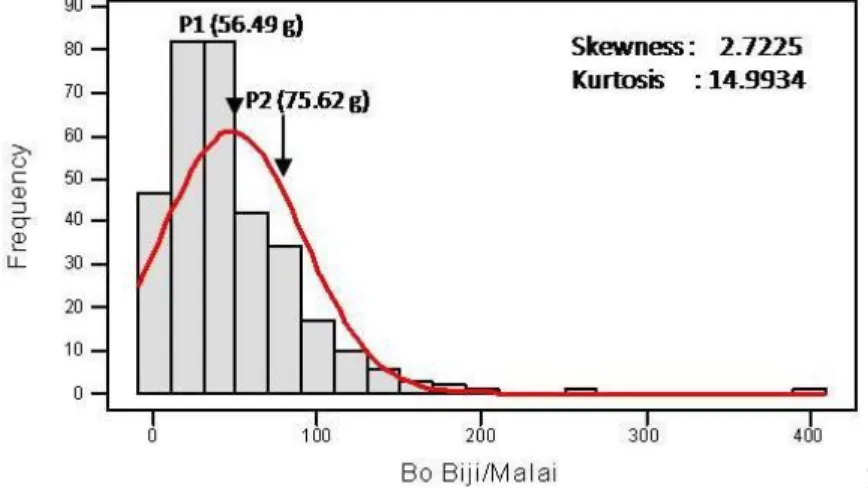
Aluminium di Lapangan ………. 51 15 Grafik Sebaran Frekuensi Bobot Biji per Tanaman Populasi F2 Hasil
Persilangan Sorgum Varietas UPCA S1 dan Numbu pada Cekaman
Aluminium di Lapangan ………. 52
Halaman
1 Komposisi Larutan Hara Steinberg yang telah dimodifikasi (Ohki
1987) ………... 70
2 Daftar Hasil Analisis Contoh Tanah Lapang UPTD Lahan Kering
Tenjo ……….. 71
3 Kriteria Kondisi Lahan Kering Masam Tenjo, Jasinga ………. 72
4 Deskripsi Varietas Numbu ………. 73
5 Deskripsi Varietas UPCA S1 ………. 74
6 Daftar Genotipe Terpilih Hasil Seleksi Berdasarkan Karakter Bobot
Biji per Tanaman Hasil dan Tinggi Tanaman Persilangan Sorgum Manis Varietas UPCA-S1 dan Numbu pada Cekaman Aluminium di Lapangan ……… 75
7 Daftar Genotipe Terpilih Hasil Seleksi Berdasarkan Karakter Bobot
Biji per Tanaman dan Bobot Batang Hasil Persilangan Sorgum Manis Varietas UPCA-S1 dan Numbu pada Cekaman Aluminium di
PENDAHULUAN
Latar Belakang
Sorgum manis [Sorghum bicolor (L.) Moench] merupakan salah satu tanaman pangan utama dunia. Hal ini ditunjukkan oleh data mengenai luas areal tanam, produksi dan kegunaan tanaman ini di dunia yang menempati peringkat ke-lima setelah gandum, padi, jagung dan barley (Murty et al. 1994). Tanaman sorgum merupakan sumber pangan utama di Afrika yang dikonsumsi lebih dari 300 juta penduduknya (Rajvanshi dan Nimbkar 2001). Pemanfaatan sorgum lainnya adalah produk olahannya berupa roti, bubur, bahan minuman (bir dan sirup) serta gula atau jiggery (ICRISAT 1996).
Tanaman sorgum manis merupakan salah satu jenis tanaman yang sangat potensial dalam pemecahan masalah krisis pangan dan energi. Berkaitan dengan program diversifikasi pangan di Indonesia, tanaman sorgum sangat potensial untuk dijadikan sebagai bahan pangan utama sumber karbohidrat karena memiliki kandungan nutrisi yang tinggi (Yudiarto 2006). Beberapa kandungan nutrisi yang dimiliki sorgum melebihi kandungan yang dimiliki penghasil karbohidrat utama (beras) yaitu protein, lemak, kalsium, besi, fosfor dan vitamin B-1 (Departemen Kesehatan RI 1992).
Batang dan biji sorgum dapat dikonversi menjadi bioetanol melalui proses fermentasi (Reddy dan Dar 2007). Sorgum memenuhi tiga syarat utama yang sangat diperlukan untuk diproduksi menjadi bahan bakar non-fosil yaitu tidak berkompetisi dengan tanaman pangan, produktivitasnya tinggi dan biaya produksinya rendah (Medco Energi 2007). Produktivitas sorgum sebagai bahan baku bioetanol lebih tinggi dibandingkan tanaman yang umum digunakan sebagai bahan baku bioetanol di Indonesia seperti tebu, ubi kayu, jagung dan gula bit (Medco Energi 2007).
Sorgum memiliki daya adaptasi agroekologi yang luas, tahan terhadap kekeringan, produksi tinggi, kebutuhan input lebih sedikit serta lebih tahan terhadap hama dan penyakit dibanding tanaman pangan lain (Hoeman 2007). Sorgum dapat tumbuh di dataran rendah hingga dataran tinggi mencapai 1500 m
di atas permukaan laut dan dapat ditanam di daerah tropis atau subtropis. Tanaman sorgum memiliki toleransi tinggi terhadap kekeringan dan kondisi iklim yang berbeda-beda (FAO 2002; Hoeman 2007). Selain itu tanaman sorgum memiliki toleransi terhadap cekaman abiotik dan dapat dibudidayakan dengan hasil yang cukup baik.
Sorgum manis ideal untuk dikembangkan di Indonesia karena tanaman ini mempunyai kesesuaian yang tinggi untuk dikembangkan di lahan kering. Lahan kering di Indonesia mencapai 144 juta hektar. Di antara luas lahan kering yang tersedia, baru sekitar 55.6 juta hektar atau sekitar 29.4% yang telah digunakan sebagai lahan pertanian (BPS 2001). Berdasarkan bentuk wilayah (topografi) sekitar 31.5 juta ha merupakan lahan kering dengan topografi yang datar berombak (kemiringan lereng < 8 %) dan sesuai untuk dibangun perkebunan sorgum. Dengan demikian lahan kering merupakan salah satu sumberdaya lahan yang mempunyai potensi besar untuk pembangunan pertanian, baik tanaman pangan, hortikultura, perkebunan maupun peternakan. Kendala yang dihadapi dari potensi lahan kering di Indonesia adalah sebesar 99.5 juta hektar (69.1%) dari total lahan kering tersebut merupakan tanah yang bereaksi masam (Hidayat dan Mulyani 2002).
Lahan kering masam dicirikan dengan pH < 5.0 dan kejenuhan basa < 50% (Hidayat dan Mulyani 2002). Lahan kering masam juga ditandai oleh
tingginya konsentrasi Aluminium (Al), terutama Al3+, yaitu bentuk Al yang
dianggap paling beracun bagi tanaman (Marschner 1995). Konsentrasi Al pada larutan tanah mineral berkisar di bawah 1 mg/l (±37 µM) pada pH ≥ 5.5 dan akan
meningkat dengan penurunan pH tanah. Konsentrasi Al3+ pada taraf tersebut dapat
dengan cepat menghambat pertumbuhan akar (Carver and Ownby 1995). Lahan bertanah masam juga mengalami defisiensi P akibat terikatnya P oleh Al yang menyebabkan P menjadi tidak tersedia bagi tanaman. Toksisitas Al pada tanaman serealia dapat menurunkan hasil antara 28-63% dari kapasitas optimumnya (Sierra et al. 2005). Aktivitas Al menyebabkan pertumbuhan akar sorgum terhambat hingga 30% untuk tetua toleran dan mencapai 45% pada populasi bersegregasi sorgum yang digunakan (Caniato et al. 2007).
Menurut Marschner (1995), terdapat dua model pendekatan yang dapat digunakan untuk mengatasi tanah marjinal yaitu dengan pendekatan bermasukan tinggi melalui penerapan agroteknologi seperti pengapuran dan pemupukan, serta dengan pendekatan bermasukan rendah melalui program pemuliaan tanaman yang diarahkan untuk mendapatkan varietas yang adaptif. Bellon (2001) menyatakan bahwa penggunaan varietas toleran Al merupakan pilihan terbaik dalam mengatasi permasalahan keracunan Al karena pengapuran terlalu mahal dan tidak efektif dalam mengatasi kemasaman tanah pada lapisan tanah yang lebih dalam. Keberhasilan program pemuliaan untuk memperoleh varietas yang mampu beradaptasi terhadap cekaman Al ditentukan oleh pemilihan metode pemuliaan yang tepat sehingga kegiatan pemuliaan dapat menjadi efektif dan efisien. Dalam upaya tersebut perlu diketahui informasi mengenai kendali genetik karakter-karakter yang akan diperbaiki (Roy 2000; Chahal and Gosal 2003). Sampai sejauh ini informasi mengenai pewarisan toleransi keracunan Al pada tanaman sorgum manis masih sangat sedikit.
Program pemuliaan sorgum toleran Al diawali dengan pembentukan populasi dasar dengan variasi genetik yang tinggi sebagai bahan pemuliaan. Populasi dasar yang memiliki variasi genetik tinggi akan memberikan respon yang baik terhadap seleksi karena akan memberikan peluang besar untuk mendapatkan genotipe dengan gabungan sifat-sifat yang diinginkan. Kegiatan selanjutnya adalah menyeleksi galur-galur/varietas-varietas dari koleksi yang dimiliki untuk memperoleh galur/varietas sorgum yang toleran Al. Seleksi pada sorgum akan menunjukkan kemajuan genetik yang tinggi jika sifat yang dilibatkan dalam seleksi mempunyai heritabilitas yang tinggi. Jika nilai heritabilitas tinggi, sebagian besar variasi fenotipe disebabkan oleh variasi genetik, maka seleksi akan memperoleh kemajuan genetik (Bernando 2002). Seleksi terhadap sifat yang mempunyai nilai heritabilitas tinggi dapat dilakukan pada generasi awal, sedangkan untuk sifat yang menunjukkan nilai heritabilitas rendah, seleksi dilakukan pada generasi akhir (Zen 1995).
Tujuan Penelitian
Tujuan penelitian ini adalah untuk
1. Memperoleh informasi mengenai pewarisan sifat toleransi Al pada tanaman sorgum manis.
2. Memperoleh karakter untuk seleksi sorgum yang toleran Al.
3. Memperoleh genotipe-genotipe toleran Al melalui seleksi pada generasi F2 di tanah masam.
Hipotesis Penelitian
Hipotesis yang digunakan dalam penelitian ini antara lain
1. Karakter-karakter pengendali toleransi Al pada sorgum manis baik stadia bibit maupun karakter agronomi dan hasil dikendalikan oleh gen-gen yang terdapat di inti.
2. Karakter-karakter pengendali toleransi Al pada sorgum manis baik stadia bibit maupun karakter agronomi dan hasil bersifat poligenik.
3. Karakter-karakter pengendali toleransi Al pada sorgum manis baik stadia bibit maupun karakter agronomi dan hasil dikendalikan oleh aksi gen aditif.
4. Terdapat karakter yang dapat dijadikan sebagai kriteria seleksi pemuliaan sorgum yang toleran Al.
Gambar 1 Bagan Alir Penelitian Studi Pewarisan Toleransi Al Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench].
Plasma Nutfah
Seleksi Adaptasi Tanah Masam Dept. AGH IPB
Seleksi Produktivitas Bioethanol B2TP-BPPT
Galur Toleran
Hibridisasi / Persilangan
Studi Pewarisan Sifat Toleransi Al di
Lapangan
Studi Pewarisan Sifat Toleransi Al di Kultur
Hara
Analisis Data
Informasi Kendali Gen dan Parameter
Genetik Pewarisan Toleransi Al pada
Sorgum
Galur Peka
Seleksi Individu pada Generasi F2 (segregan) Genotipe F2 Toleran Al Selfing F1/F1R F2 Karakter Seleksi pemuliaan sorgum yang toleran Al
TINJAUAN PUSTAKA
Sorgum Manis [Sorghum bicolor (L.) Moench]
Sorgum manis [Sorghum bicolor (L.) Moench] merupakan tanaman dari famili Gramineae (Poaceae), subfamili Panicoideae dan genus Andropogon (Rukmana dan Oesman 2001). Daerah asal penyebaran tanaman sorgum baik spesies liar maupun spesies budidaya terbesar ditemukan di Afrika (Grubben dan Partohardjomo 1996) untuk selanjutnya tumbuh dan berkembang di berbagai daerah seperti India, Nigeria, Argentina, Meksiko dan Sudan (ICRISAT 1996).
Keunggulan tanaman sorgum adalah sangat efisien dalam penggunaan air. Hal ini disebabkan karena sorgum memiliki sistem perakaran yang halus dan pertumbuhan akar agak dalam sehingga memungkinkan penyerapan air yang cukup intensif (Rismunandar 1989). Sistem perakaran sorgum terdiri dari akar-akar seminal (akar-akar-akar-akar primer) pada dasar buku pertama pangkal batang, akar-akar koronal (akar-akar pada pangkal batang yang tumbuh ke arah atas) serta akar-akar udara. Tanaman sorgum dapat membentuk perakaran sekunder berukuran dua kali lipat dari akar sekunder jagung (Rukmana dan Oesman 2001). Doggett (1970) menyimpulkan bahwa pada endodermis akar sorgum terdapat endapan silika yang berfungsi mencegah kerusakan akar pada kondisi kekeringan.
Batang tanaman sorgum berbentuk silinder, beruas-ruas dan berbuku-buku (ICRISAT 1996). Setiap ruas memiliki alur yang letaknya berselang-seling. Batang sorgum memiliki sel-sel parenkim atau seludang pembuluh yang diselubungi oleh sebuah lapisan keras. Beberapa varietas sorgum dapat membentuk cabang dan memiliki anakan (Rukmana dan Oesman 2001). Menurut Martin (1970) banyaknya anakan yang berkembang tergantung faktor genetik, jarak tanam, kelembaban tanah, fotoperiodisme, vigor tanaman dan waktu tanam. Ukuran diameter batang bervariasi antara 0.5 sampai 5 cm. Begitu juga dengan tinggi tanaman bervariasi dari 0.5 sampai 4 m (Murty et al. 1994). Tinggi tanaman dipengaruhi oleh jumlah buku, panjang ruas batang, panjang tangkai malai, panjang malai dan faktor genetik (Doggett 1970). Perpanjangan buku pada
tanaman sorgum dikendalikan oleh empat lokus gen Dwarf yaitu Dw1, Dw2, Dw3
dan Dw4(House 1985).
Daun tanamam sorgum terdiri dari helai daun dan pelepah daun dengan panjang helai daun mencapai 30-135 cm dan lebar daun maksimum 13-15 cm. Menurut Rismunandar (1989) daun sorgum dilapisi sejenis lilin yang agak tebal berwarna putih, berfungsi untuk menahan atau mengurangi penguapan air dari dalam tanaman sehingga toleran terhadap kekeringan.
Bunga sorgum tersusun dalam bentuk malai dengan banyak bunga pada setiap malai sekitar 1500-4000 bunga. Bunga sorgum akan mekar teratur dari cabang malai paling atas hingga ujung malai paling bawah. Malai sorgum memiliki tangkai yang tegak atau melengkung, berukuran pendek atau panjang dan bentuk malai dari agak kompak sampai terbuka, oval, kerucut, ramping panjang atau piramida (Murty et al. 1994).
Biji sorgum berbentuk kariopsis atau karnel yaitu buah berbiji tunggal dengan kulit buah (pericarp) yang bersatu dengan kulit biji. Warna, ukuran dan bentuk biji sorgum beragam. Kulit biji dapat berwarna putih, krem, kekuningan, merah atau coklat. Biji sorgum dapat berbentuk bola, bentuk seperti buah pear dan gepeng pada salah satu bagiannya. Bobot 100 biji sorgum berkisar 0.75-7.5 gram (Murty et al. 1994). Waktu yang diperlukan biji sorgum untuk mencapai berat kering maksimal tergantung pada kondisi pertumbuhan, biasanya 25-55 hari setelah antesis dengan kadar air 25-35% (Doggett 1970). Menurut Rismunandar (1989) malai sorgum dapat dipanen rata-rata setelah tanaman berumur 90-120 hari.
Tanaman sorgum mampu tumbuh di daerah tropis maupun subtropis mulai dari dataran rendah hingga dataran tinggi sampai ketinggian 1500 m di atas permukaan laut (Rismunandar 1989). Kondisi yang optimum untuk tanaman
sorgum adalah daerah bersuhu 20-30oC, kelembaban rendah dan curah hujan
375-425 mm selama tanaman masih muda hingga mencapai umur 4-5 minggu. Sorgum dapat tumbuh di hampir setiap jenis tanah.
Sorgum merupakan tanaman C4 yaitu spesies tanaman yang menghasilkan
dalam proses metabolisme. Tanaman jenis ini memanfaatkan kedua sel mesofil
dan sel seludang berkas untuk menambat CO2 sehingga jenis tanaman C4 menjadi
sangat efisien dalam fotosintesis. Produk yang dihasilkan sel mesofil berupa asam malat dan asam aspartat dengan cepat ditransfer ke sel seludang berkas dan asam
tersebut mengalami dekarboksilasi dengan melepaskan CO2 yang selanjutnya
ditambat oleh Rubisco untuk diubah menjadi 3-PGA (asam fosfogliserat). Tanaman C4 juga memiliki sel seludang berkas yang lebih tebal dibandingkan dengan sel seludang pada tanaman C3 sehingga mengandung lebih banyak kloroplas, mitokondria dan organel penting lainnya dalam proses fotosintesis (Salisbury dan Ross 1992; Taiz dan Zeiger 2002). Tanaman C4 juga mampu berfotosintesis lebih cepat pada penyinaran tinggi dan suhu panas sehingga mampu menghasilkan biomassa yang lebih banyak dibandingkan tanaman C3 (Salisbury dan Ross 1992).
Pada tanaman sorgum, selain memiliki mekanisme fotosintesis yang efisien juga memiliki mekanisme fisiologi lainnya antara lain permukaan daun yang dilapisi lilin sehingga dapat mengurangi laju transpirasi dan mempunyai sistem perakaran yang ekstensif (House 1985). Hal ini membuat produktivitas biomassa sorgum lebih tinggi dibandingkan tanaman C4 sejenis yaitu jagung (Hoeman 2007). Penelitian Borrel et al. (2005) menemukan bahwa tanaman sorgum efisien dalam penggunaan radiasi dan transpirasi karena memiliki gen pengendali stay-green sejak fase pengisian biji yang berhubungan dengan kandungan nitrogen daun spesifik. Fenomena ini mampu memperlambat proses senescen pada daun sorgum sehingga mampu mempertahankan batang dan daun tetap hijau meskipun pasokan air sangat terbatas (Seetharama dan Mahalakshmi 2006; Borrel et al. 2006).
Mekanisme Toleransi terhadap Aluminium
Lahan kering masam dicirikan dengan pH < 5.0 dan kejenuhan basa < 50% (Hidayat dan Mulyani 2002). Tanah masam juga ditandai oleh tingginya
konsentrasi Aluminium (Al), terutama Al3+, yaitu bentuk Al yang dianggap
morfologi, fisiologi dan ekspresi gen tanaman. Cekaman Al menyebabkan terganggunya penyerapan hara tanaman (Adam et al. 1999). Meningkatnya konsentrasi Al terlarut menyebabkan penyerapan unsur-unsur hara berkurang sehingga pada akhirnya mengakibatkan terjadinya defisiensi hara antara lain Ca, P, K Mg dan hara mikro seperti seng, tembaga dan molybdenum. Terganggunya penyerapan secara langsung disebabkan kerusakan membran sel akar. Akumulasi Al dapat menyebabkan kebocoran membran, mengurangi kandungan K dalam jaringan ujung akar dan merusak viabilitas protoplasma karena Al dan membran plasma akar membentuk ikatan polimer sehingga terjadi kerusakan pada membran dan kebocoran K dari sel akar (Matsumoto et al. 1992).
Kerusakan akar akibat cekaman Al juga disebabkan terjadinya penebalan ujung akar dan akar cabang. Penelitian Caniato et al. (2007) memperlihatkan bahwa aktivitas Al menyebabkan pertumbuhan akar sorgum terhambat hingga 30% untuk tetua toleran dan mencapai 45% pada populasi bersegregasi sorgum yang digunakan. Penelitian Yamamoto (1992) mendapatkan hasil bahwa toksisitas Al pada tembakau menyebabkan tanaman kekurangan hara dan juga mengubah struktur dan fungsi membran plasma serta menghalangi pembelahan sel pada ujung-ujung akar.
Kelarutan Al yang tinggi secara tidak langsung berpengaruh terhadap proses-proses fisiologi dan metabolisme tanaman. Terganggunya penyerapan hara menyebabkan ketersediaan unsur hara menurun sehingga pertumbuhan tajuk tanaman menjadi tertekan (Marschner 1995). Polle dan Konzak (1990) menjelaskan bahwa kerusakan akar oleh Al menyebabkan terganggunya hara dan
meningkatkan kepekaan terhadap kekeringan sehingga mempengaruhi
pertumbuhan dan produktivitas tanaman.
Mekanisme toleransi terhadap Al dapat dikelompokkan menjadi mekanisme eksternal dan mekanisme internal (Taylor 1991; Sopandie 2006). Tanaman yang memiliki mekanisme eksternal mampu mencegah Al masuk ke dalam simplas dan bagian metabolik yang sensitif. Mekanisme ini dapat dicapai antara lain dengan imobilisasi Al pada dinding sel dan selektivitas plasma membran terhadap Al, induksi pH di rizosfir atau apoplas akar, sekresi senyawa
organik pengkelat Al. Penelitian Pineros dan Kochian (2001) pada pewarnaan akar dengan hematoxylin menunjukkan bahwa jagung yang toleran Al mampu melakukan akumulasi Al pada lapisan sel bagian luar tudung akar sebagai upaya penahanan Al masuk ke dalam jaringan.
Peningkatan pH rizosfer pada larutan/media merupakan salah satu indikator toleransi tanaman terhadap cekaman Al. Beberapa penyebab penurunan pH pada zone perakaran: 1) Pelepasan ion H akibat absorpsi kation > anion, 2)
Pelepasan dan hidrolisis CO2, 3) Pelepasan ion-ion H dari gugus karboksil asam
poligalakturonat dan sisa-sisa asam pektat dan 4) Ekskresi proton-proton dari mikroorganisme-mikroorganisme yang berhubungan dengan akar. Tanaman yang memiliki mekanisme toleransi dengan mengubah pH di daerah perakaran akan meningkat pH larutan hara sehingga tanaman mampu menurunkan kelarutan dan toksisitas Al (Sasaki et al. 1997). Percobaan pada metode kultur hara menunjukkan bahwa genotipe toleran pada tanaman jagung, gandum, barley dan padi mengalami peningkatan pH larutan serta terjadi penurunan kelarutan dan toksisitas Al (Caniato et al. 2007; Furukawa et al. 2007). Menurut Hayes (1990), peningkatan pH rizosfer akan meningkatkan ketersediaan unsur hara seperti P dan menurunkan ketersediaan Zn, Cu, Fe, Mn dan Al. Penelitian Delhaize et al. (1995) pada tanaman kedelai menunjukkan bahwa genotipe toleran mampu meningkatkan pH rizosfer dua kali lipat dibandingkan dengan genotipe peka. Kenaikan pH akan mengurangi kelarutan Al, toksisitas dan melepaskan ikatan dengan P.
Tanaman toleran mampu menghasilkan asam organik yang dapat mengkelat Al dan mencegah pengikatan Al-P dalam akar (Delhaize et al. 1993; Furukawa et al. 2007). Ujung akar tanaman yang toleran Al selain mengeksudasi asam organik juga mampu mengeksudasi fosfat organik. Hal ini merupakan proses alami hara P dan bagian dari keseimbangan hara P dalam tanaman (Pellet et al. 1996). Detoksifikasi Al oleh fosfat karena terbentuknya ikatan kompleks Al-P (Delhaize et al. 1993). Tanaman jagung dan gandum toleran mampu mengeksudasi fosfat organik lebih banyak dibandingkan tanaman peka (Pellet et
al. 1995 dan 1996). Mekanisme eksternal menyebabkan kandungan Al dalam jaringan menjadi rendah.
Tanaman dengan mekanisme internal memiliki daya toleransi untuk mengakumulasi Al dalam sel sehingga kandungan Al dalam jaringan tinggi. Mekanisme resistesi internal dicapai dengan pengkelatan Al oleh asam organik dalam sitosol, kompartementasi Al di vakuola, memproduksi protein pengikat Al serta produksi dan peningkatan enzim yang tahan Al.
Senyawa organik pada tanaman toleran Al mampu melakukan kompleks atau bahkan kelat (menjepit) Al sehingga dapat mengurangi kelarutan Al (Hayes dan Swift 1990; Tan 1993). Senyawa organik pada tanaman peka Al tidak efektif melakukan kompleks atau mengkelat ion logam, salah satunya disebabkan karena jumlah yang dihasilkan tidak mampu untuk menetralkan atau mengusir Al (Sopandie et al. 2003; Kasim et al. 2001).
Anion organik pada barley mengaktifkan Al-effluks dari akar dan berkorelasi dengan toleransi Al (Furukawa et al. 2007). Caniato et al. (2007) menyatakan bahwa mekanisme fisiologi toleransi Al pada sorgum berdasarkan pengkelatan Al di rizosfer oleh malat yang dilepas dari apikal akar dan mencegah logam mencapai situs sensitif di dalam akar. Tanaman teh merupakan salah satu tanaman yang menghasilkan asam organik/polifenol yang dapat menawarkan racun Al dengan cara mengkelatnya (Matsumoto dan Sasaki 2002). Aluminium berikatan dengan catechin di dalam daun muda dan pucuk, sedangkan pada daun tua ditemukan adanya kompleks aluminium-asam fenolik dan asam alumonium-asam organik.
Pemuliaan bagi Lingkungan Bercekaman
Lingkungan bercekaman adalah lingkungan sub optimum bagi pertumbuhan dan produksi tanaman. Lingkungan sub optimum merupakan lingkungan yang beragam dan tidak sama tingkat cekamannya. Tujuan pemuliaan untuk perbaikan produktivitas tanaman pada lingkungan bercekaman ditentukan oleh tingkat cekaman pada lingkungan target (Sopandie et al. 2004; Sopandie 2006).
Perbaikan produktivitas tanaman di lahan bercekaman dapat dilakukan melalui perbaikan potensi hasil untuk mendapatkan varietas berdaya hasil tinggi dan perbaikan daya adaptasi tanaman untuk mendapatkan varietas yang toleran (Accevedo dan Fererres 1993). Menurut Baker (1993), produktivitas tanaman pada kondisi bercekaman ditentukan oleh potensi hasil tanaman serta daya
adaptasi tanaman terhadap cekaman. Upaya untuk meningkatkan produktivitas tanaman pada kondisi cekaman
ringan atau sedang dilakukan melalui perbaikan potensi hasil dan pembentukan idiotype breeding (Romagosa dan Fox 1993). Perbaikan potensi hasil dapat dilakukan melalui perbaikan kapasitas fotosintesis dan respirasi untuk meningkatkan biomassa tanaman dan perbaikan sink capacity dan partisi fotosintat (Accevedo dan Fererres 1993). Upaya pembentukan idiotype breeding dilakukan dengan mengembangkan kombinasi karakter yang mendukung fotosintesis, pertumbuhan dan produksi tanaman (Sopandie 2006). Melalui pembentukan idiotype breeding, pemulia akan memperoleh gambaran yang jelas dalam menyeleksi karakter tanaman yang mendukung peningkatan potensi hasil (Romagosa dan Fox 1993; Sopandie 2006).
Upaya untuk meningkatkan produktivitas tanaman pada kondisi cekaman berat dilakukan melalui peningkatan adaptasi tanaman (Romagosa dan Fox 1993). Pada lingkungan dengan cekaman berat terdapat interaksi genotipe dengan lingkungan baik interaksi yang bersifat kuantitatif maupun interaksi yang bersifat kualitatif. Interaksi genotipe dengan lingkungan yang bersifat kuantitatif tidak menyebabkan perubahan rangking genotipe. Genotipe yang unggul pada satu lingkungan tetap unggul pada lingkungan yang berbeda. Interaksi genotipe lingkungan yang bersifat kualitatif merupakan kendala dalam pemuliaan bagi lingkungan bercekaman berat karena mengakibatkan perubahan rangking genotipe (Ceccareli 1996). Genotipe berdaya hasil tinggi pada satu lingkungan bisa mengalami penurunan hasil yang cukup tajam pada lingkungan berbeda akibat pengaruh cekaman (Romagosa dan Fox 1993; Roy 2000; Bernardo 2002; Chahal dan Gosal 2003).
Seleksi pada lingkungan bercekaman harus dilakukan di lingkungan target sehingga dapat memaksimalkan ekspresi gen-gen yang mengendalikan daya hasil maupun daya adaptasi tanaman terhadap cekaman (Ceccareli et al. 2007). Seleksi pada kondisi bercekaman dapat dilakukan berdasarkan fenotipe, marka molekular dan gabungan antara fenotipe dan marka molekuler (Bernardo 2002). Seleksi berdasarkan fenotipe menjadi sulit karena nilai heritabilitas yang rendah dan adanya interaksi antara genotipe dan lingkungan yang bersifat kualitatif. Untuk meningkatkan efisiensi seleksi maka seleksi dapat dilakukan menggunakan marka molekuler. Marka molekuler yang terpaut dengan QTL atau yang mengendalikan daya hasil pada kondisi bercekaman serta QTL yang mengendalikan toleransi terhadap cekaman merupakan salah satu marka yang dapat dijadikan alat bantu seleksi (Forster et al. 2000; Hussain 2006).
Arah Program Pemuliaan Tanaman Sorgum
Tanaman sorgum manis merupakan salah satu jenis tanaman yang sangat potensial dalam pemecahan masalah krisis pangan dan energi. Kebutuhan terhadap tanaman ini terus meningkat (Grassi 2001). Konsekuensinya adalah perbaikan hasil dan kualitas sangat perlu dilakukan. Secara umum, tanaman sorgum manis yang diinginkan adalah tanaman sorgum yang memiliki indeks panen yang tinggi, produktivitas yang tinggi, stabilitas hasil, resisten terhadap penyakit dan memiliki toleransi terhadap cekaman abiotik seperti tidak sensitif terhadap photoperiodisme, kelembaban tinggi, toleran kekeringan dan lahan masam (Baenziger 2006).
Ketersediaan bahan bakar tidak terbarukan (berbasis fosil) saat ini semakin terbatas menyebabkan kebutuhan akan sumber energi yang terbarukan (biofuel) menjadi penting. Penggunaan bioetanol dapat mengurangi emisi gas rumah kaca sampai 12% (Reddy dan Dar 2007). Dari tahun ke tahun kebutuhan terhadap bioetanol semakin meningkat. Sorgum merupakan salah satu tanaman yang dapat menjadi bahan baku industri bioetanol karena batang dan juga bijinya dapat dikonversi menjadi bioetanol melalui proses fermentasi (Yudiarto 2006; Reddy dan Dar 2007). Selain produktivitasnya yang tinggi dan biaya produksi yang
rendah, tanaman sorgum tidak berkompetisi dengan tanaman pangan. Saat ini produsen bioetanol yang menggunakan sorgum masih didominasi oleh Amerika Serikat, Cina, Afrika Selatan dan India (Dirjen Tanaman Pangan 2007). Sorgum memenuhi persyaratan sebagai bahan baku bioetanol antara lain dapat tumbuh dalam berbagai agroekologi, lebih tahan penyakit dan memerlukan input produksi yang relatif sedikit jika dibandingkan dengan tanaman penghasil bioetanol lainnya.
Rasmusson (1987) menjelaskan bahwa ideotipe merupakan sifat yang diharapkan dapat ditingkatkan potensi genetik hasilnya. Pemuliaan ideotipe didefinisikan sebagai metode yang digunakan dalam pemuliaan untuk meningkatkan potensi genetik karakter yang diinginkan dengan memodifikasi karakter tersebut secara spesifik (Roy 2000). Arah pemuliaan sorgum untuk produksi bioetanol diarahkan pada perbaikan karakter-karakter produksi bioetanol yaitu karakter malai dan batang.
Tanaman sorgum memiliki efisiensi fotosintesis yang tinggi yaitu 2.5% dengan efisiensi fotosintesis maksimum pada beberapa jam tertentu dalam siklus harian mencapai 27% (Grassi 2005). Perbaikan efisiensi fotosintesis diharapkan dapat meningkatkan produksi biomassa sorgum sehingga produktivitas bioetanol juga akan meningkat. Alkohol diperoleh dari nira bagian batang sorgum. Kualitas nira sorgum manis setara dengan nira tebu, kecuali kandungan amilum dan asam akonitat yang relatif tinggi (Sirappa 2003). Kandungan amilum yang tinggi tersebut merupakan salah satu masalah dalam proses kristalisasi nira sorgum sehingga gula yang dihasilkan berbentuk cair. Untuk mengatasi masalah tersebut, pengembang ideotipe dapat diarahkan pada penurunan kandungan amilum dari kadar awal.
Bioetanol juga diperoleh dari bagian malai. Sirappa (2003) menjelaskan bahwa biji sorgum mengandung 65-71% pati yang dapat dihidrolisis menjadi gula sederhana. Gula sederhana yang diperoleh dari biji sorgum selanjutnya dapat difermentasi untuk menghasilkan alkohol. Menurut Somani dan Pandrangi (1993) dalam Sumarno dan Karsono (1996), setiap ton biji sorgum dapat menghasilkan
384 liter bioalkohol. Seleksi berdasarkan bobot biji per tanaman diharapkan akan diperoleh perbaikan produksi malai surgum.
Kelebihan lain adalah manfaat ganda tanaman sorgum yang dapat digunakan dalam memenuhi kebutuhan pangan dan pakan. Sorgum manis merupakan salah satu tanaman pangan dunia dengan luas areal tanam, produksi dan kegunaan tanaman ini di dunia yang menempati peringkat ke-lima setelah gandum, padi, jagung dan barley (Murty et al. 1994). Tanaman sorgum merupakan sumber pangan utama di Afrika yang dikonsumsi lebih dari 300 juta penduduknya (Rajvanshi dan Nimbkar 2001). Pemanfaatan sorgum lainnya adalah produk olahannya berupa roti, bubur, bahan minuman (bir dan sirup) serta gula atau jiggery (ICRISAT 1996).
Berkaitan dengan program diversifikasi pangan di Indonesia, tanaman sorgum sangat potensial untuk dijadikan sebagai bahan pangan utama sumber karbohidrat karena memiliki kandungan nutrisi yang tinggi (Yudiarto 2006) Beberapa kandungan nutrisi yang dimiliki sorgum melebihi kandungan yang dimiliki penghasil karbohidrat utama (beras) yaitu protein, lemak, kalsium, besi, fosfor dan vitamin B-1 (Sirappa 2003).
Kandungan tannin pada beberapa jenis sorgum cukup tinggi (0,40−3,60%), sehingga hasil olahannya kurang enak. Sumarno dan Karsono (1996) menyarankan untuk mengatasi masalah ini melalui teknologi pengolahan kulit dan lapisan testa dengan pengikisan (penyosohan). Pendekatan melalui program pemuliaan dapat dilakukan melalui perakitan sorgum dengan kandungan tannin rendah. Keberadaan tannin pada sorgum sulit terdeteksi dan tidak tergantung pada warna biji (House 1985).
Sorgum memenuhi syarat gizi dan faktor biofisik untuk dijadikan jalan keluar dalam krisis pangan dan dalam rangka perbaikan gizi masyarakat, sehingga pengembangan varietas sorgum dengan level vitamin, mineral dan protein tinggi harus dilakukan. Sorgum memiliki kandungan glutin sorgum sangat rendah (Graybosch 1992) sehingga sorgum dikembangkan sebagai bahan pangan premium untuk keperluan diet pada penderita diabetes dan diet pada penderita alergi glutin (Hoeman 2008).
Pemanfaatan sorgum sebagai pakan ternak yaitu biji sorgum untuk bahan campuran ransum pakan ternak unggas, sedangkan batang dan daun sorgum (stover) untuk ternak ruminansia (Hoeman 2007). Kandungan lemak sorgum yang relatif tinggi dapat dimanfaatkan untuk meningkatkan bobot ternak. Dengan demikian, pemilihan ideotipe dapat diarahkan pada perakitan genotipe yang memiliki kandungan lemak tinggi. Penggunaan biji sorgum dalam ransum pakan ternak bersifat suplemen (substitusi) terhadap jagung karena nilai nutrisinya tidak berbeda dengan jagung, namun karena kandungan tannin yang cukup tinggi membuat rasa pakan biji sorgum menjadi pahit (Sirappa 2003). Menurut Koentjoko (1996), kandungan tanin dalam ransum di atas 0,50% dapat menekan pertumbuhan ayam, dan apabila mencapai 2% akan menyebabkan kematian (Rayudu et al. 1970) sehingga salah satu ideotipe yang dapat dikembangkan dalam program pemuliaan sorgum untuk keperluan pakan adalah menurunkan kadar tannin pada lapisan aleuron biji.
Potensi daun sorgum manis sekitar 14−16% dari bobot segar batang atau sekitar 3 ton daun segar/ ha dari total produksi 20 ton/ha. Soebarinoto dan Hermanto (1996) melaporkan bahwa setiap hektar tanaman sorgum dapat menghasilkan jerami 2,62 + 0,53 ton bahan kering. Konsumsi rata-rata setiap ekor sapi adalah 15 kg daun segar/hari (Direktorat Jenderal Perkebunan 1996). Dengan demikian pengembangan sorgum sebagai tanaman pakan ternak (forage sorghum) dapat dilakukan dengan meningkatkan potensi hijauan sorgum. Borrel et al. (2006) menjelaskan kondisi stay green pada tanaman sorgum dipengaruhi oleh efisiensi penggunaan radiasi dan transpirasi yang tinggi.
Penelitian sorgum selain mengarah pada perbaikan sifat agronomi dan kualitas sebagai sumber pangan, pakan dan bahan baku bioetanol, juga diarahkan pada peningkatan produktivitas antara lain dengan perakitan sorgum yang memiliki daya adaptasi luas dengan produktivitas tinggi namun memerlukan input relatif rendah, peningkatan ketahanan terhadap hama dan penyakit tanaman serta toleransi terhadap kondisi marjinal (kekeringan, salinitas dan lahan masam). Arah pemuliaan sorgum dalam peningkatan komponen hasil dan produktivitas dapat dilakukan dengan memanfaatkan galur-galur murni dan hibrida (Institute of
Agricultural Sciences at Xinxian-Shanxi 1972). Hasil panen pada hibrida sorgum berkorelasi dengan berat malai. Poelman dan Sleper (1996) menerangkan bahwa program pemuliaan sorgum untuk daya hasil tinggi memerlukan faktor-faktor yang mempengaruhi pertumbuhan tanaman seperti umur tanam, sensitifitas terhadap fotoperiode, resistensi terhadap kerebahan dan resistensi terhadap cekaman (biotik maupun abiotik).
Karakter agronomi seperti tinggi tanaman, jumlah daun dan produksi biji dapat digunakan sebagai indikator tahan kekeringan. Grassi et al. 2004 menjelaskan bahwa daya adaptasi sorgum terhadap kekeringan berhubungan dengan mekanisme efisiensi penggunaan air. Penelitian yang dilakukan Di Fonzo et al. (1999) diketahui bahwa luas area hijau daun pada fase generatif meningkatkan produksi biji. Genotipe dengan kemampuan stay-green pada tanaman pangan seperti jagung, sorgum dan gandum, lebih aktif berfotosintesis selama masa pengisian biji. Hal serupa juga dijelaskan oleh van Oosterom et al. (1996), bahwa karakter stay-green merupakan komponen toleransi terhadap kekeringan pada fase akhir pembungaan sorgum. Sorgum dengan fenotipe stay-green mengakumulasi lebih banyak gula pada batang baik selama maupun setelah fase pengisian biji. Karakter stay-green merupakan karakter penting dalam perakitan varietas sorgum manis toleran kekeringan dan juga dapat digunakan dalam aplikasi pemuliaan lainnya.
Penurunan produktivitas tanaman pada tanah masam terutama dialami oleh jenis tanaman semusim (annual plant) dengan sistem perakaran yang dangkal (Kochian et al. 2004). Pada kondisi tercekam kemasaman, pertumbuhan tajuk akan terhambat yang ditunjukkan oleh pertumbuhan yang kerdil (Marschner 1995). Hal ini disebabkan oleh terhambatnya akses air dan nutrisi karena pertumbuhan akar yang terhambat. Sistem perakaran merupakan salah satu karakter yang dapat digunakan tanaman untuk mengurangi toksisitas Al dan meningkatkan kemampuan tanaman dalam memanfaatkan unsur hara.
Strategi untuk meningkatkan produktivitas tanaman di lingkungan bercekaman adalah melalui program pemuliaan tanaman yang didukung oleh pemahaman tentang aspek fisiologi adaptasi tanaman (Sopandie 2006).
Pengembangan sorgum yang memiliki toleransi terhadap Al diperlukan upaya mengembangkan varietas-varietas sorgum yang beradaptasi pada kondisi agroekologi lahan bercekaman Al. Varietas sorgum toleran Al telah dikembangkan di beberapa negara (Kochian et al. 2004), tetapi di Indonesia saat ini belum dikembangkan, karenanya pengembangan varietas sorgum toleran tanah masam sangat diperlukan. Pemahaman tentang mekanisme adaptasi dan kendali genetiknya dapat membantu meningkatkan efektivitas seleksi galur-galur sorgum toleran Al.
Indonesia bukan merupakan daerah origin sorgum, namun tanaman sorgum di Indonesia sebenarnya sudah sejak lama dikenal. Perkembangan sorgum di Indonesia tidak sebaik padi dan jagung karena masih sedikit daerah yang memanfaatkan tanaman sorgum sebagai bahan pangan. Budidaya, penelitian dan pengembangan tanaman sorgum di Indonesia masih sangat terbatas (Hoeman 2007). Salah satu penyebab keterbatasan tersebut adalah tidak tersedianya benih varietas unggul sorgum. Hal inilah yang menyebabkan keragaman genetik yang tersedia masih sangat terbatas. Peningkatan keragaman genetik sorgum dapat dilakukan melalui introduksi, hibridisasi, mutasi dan bioteknologi maupun kombinasi antara metode-metode tersebut.
Varietas sorgum yang terdapat di Indonesia sebagian besar merupakan introduksi dari International Crop Research Institut for Semi-Arid Tropics (ICRISAT) dan dari beberapa negara seperti India, Thailand dan Cina. Setelah melalui proses pengujian adaptasi dan daya hasil selama beberapa generasi, beberapa varietas introduksi tersebut telah dilepas oleh Departemen Pertanian Republik Indonesia sebagai varietas sorgum unggul nasional antara lain UPCA, Keris, Mandau, Higari, Numbu, Kawali, Badik, Gadam dan Sangkur (Dirjen Bina Produksi Tanaman Pangan 2003).
Teknik hibridisasi atau persilangan buatan dapat menimbulkan keragaman baru melalui rekombinasi yang terbentuk dari alela-alela yang berasal dari tetua-tetua persilangan. Persilangan dapat menghasilkan keragaman baru yang tidak ditemukan pada genotipe kedua tetuanya karena adanya rekombinasi (Baenziger 2006). Setelah melalui segregasi akan terbentuk populasi yang mempunyai
keragaman genetik lebih tinggi. Dengan hibridisasi, keragaman yang dibentuk dapat diarahkan sesuai dengan sasaran program pemuliaan, berupa gabungan karakter-karakter unggul yang ada di tetua-tetua persilangan.
Persilangan buatan dapat melibatkan semua bentuk genotipe, varietas dari suatu spesies yang sama atau antar spesies yang berbeda. Berdasarkan pengelompokan genotipe ini persilangan buatan dapat dikelompokkan menjadi (1) Intervarietal yaitu persilangan yang melibatkan tetua-tetua berupa kultivar dari spesies yang sama (Harlan dan de Wet 1971), (2) Interspesifik yaitu persilangan yang melibatkan tetua yang berasal dari dua spesies yang berbeda dari genus yang sama (Harlan dan de Wet 1971), (3) Intergenerik yaitu persilangan yang melibatkan tetua-tetua yang berasal dari spesies dari genus yang berbeda (Greene dan Morris 2001) dan (4) Introgressi yaitu persilangan yang melibatkan tetua-tetua yang berasal dari spesies yang berbeda dengan tujuan untuk memindahkan satu atau beberapa gen saja dari tetua spesies liar ke spesies budidaya (Gepts 2002). Tipe persilangan yang paling umum dilakukan adalah tipe persilangan intervarietal, karena rekombinasi gen dapat terjadi lebih mudah dan tanpa hambatan reproduksi. Persilangan yang melibatkan spesies tanaman yang berbeda mempunyai kendala pada hambatan reproduksi sehingga memerlukan teknik khusus.
Pemilihan tetua sangat penting diperhatikan dalam hibridisasi karena menentukan keberhasilan dari tujuan persilangan yang diinginkan. Tetua yang digunakan dalam persilangan harus membawa karakter unggul yang diinginkan (Sutjahjo et al. 2005). Selain itu, salah satu atau kedua tetua memiliki adaptasi dan penampilan agronomis yang baik dan juga antara tetua mempunyai jarak kekerabatan yang jauh sehingga dapat menghasilkan keragaman genetik yang tinggi pada turunannya.
Sumber keragaman untuk seleksi tetua persilangan dapat diperoleh dari koleksi plasma nutfah yang ada atau jika tidak ada dapat diintroduksikan dari wilayah lain. Sutjahjo et al. (2005) menjelaskan bahwa plasma nutfah yang menjadi sumber keragaman bagi seleksi tetua dapat terdiri dari varietas komersial, galur-galur elit hasil pemuliaan (breeding lines), varietas lokal atau landrace dan
spesies liar. Varietas komersial adalah varietas yang telah ditanam luas dan diterima baik oleh petani. Galur-galur elit hasil pemuliaan (breeding lines) adalah yaitu galur-galur terpilih dengan sifat-sifat unggul tetapi belum dilepas sebagai varietas. Varietas lokal atau landrace yaitu kultivar hasil seleksi petani yang mempunyai keunggulan dalam adaptasi terhadap cekaman lingkungan tertentu seperti kekeringan, tanah masam, salinitas atau tanah gambut sulfat masam, sedangkan spesies liar adalah spesies bukan budidaya namun mempunyai sifat-sifat yang diinginkan dan akan digunakan dalam persilangan dengan spesies budidaya.
Jika keragaman dari karakter yang dikehendaki tidak dapat ditemukan dalam koleksi plasma nutfah, maka keragaman dapat diperoleh melalui mutasi induksi (Ahloowalia et al. 2004). Aisyah (2006) menjelaskan bahwa mutasi adalah perubahan materi genetik yang terjadi secara mendadak dan bersifat permanen akibat perlakuan radiasi atau bahan kimia (mutagen fisik atau kimia). Keragaman yang ditimbulkan oleh mutasi tidak dapat diduga arahnya (van Harten 1998). Peningkatan keragaman lainnya dapat dilakukan dengan melalui pendekatan bioteknologi antara lain embryo rescue (Comeau et al. 1992), manipulasi kromosom sitoplasma (Fedak 1999; Jauhar dan Chibbar 1999) atau manipulasi kromosom molekular dengan transformasi genetik (Zhong 2001).
Kegiatan selanjutnya adalah proses seleksi dan pengujian selama beberapa generasi hingga diperoleh galur-galur harapan dengan karakter yang diinginkan. Sorgum termasuk kelompok tanaman yang melakukan penyerbukan sendiri dengan persentase menyerbuk silang sebesar 6% (Poehlman dan Sleper 1996). Sasaran yang hendak dicapai pada program pemuliaan tanaman menyerbuk sendiri adalah sifat unggul dan tanaman homozigot (Poespodarsono 1988). Metode seleksi yang digunakan untuk tanaman menyerbuk sendiri seperti sorgum dapat dilakukan pada populasi yang heterogen dan pada populasi bersegregasi (Trikoesoemaningtyas, bahan kuliah 2007). Seleksi pada populasi heterogen dilakukan dengan metode seleksi massa dan seleksi galur murni, sedangkan pada populasi bersegregasi, seleksi dapat dilakukan dengan metode adalah pedigree, bulk, silang balik, single seed descent dan double haploid.
Metode pedigree banyak digunakan untuk seleksi pada karakter kualitatif atau karakter kuantitatif yang mempunyai nilai heritabilitas yang tergolong tinggi sehingga seleksi dapat dilakukan pada generasi awal (Moreno-Gonzales dan Cubero 1993; Roy 2000; Chahal dan Gosal, 2003). Metode pedigree merupakan metode yang paling sering digunakan dalam pemuliaan sorgum (House 1985). Kelebihan metode pedigree antara lain hanya keturunan dari tanaman unggul saja yang dilanjutkan, menghemat lahan karena jumlah tanaman tiap generasi semakin sedikit dan silsilah dari galur diketahui (Sutjahjo et al. 2006). Namun pada metode ini banyak genotipe akan terbuang pada saat masih bersegregasi, sedangkan genotipe tersebut mungkin akan mempunyai fenotipe yang baik pada generasi lanjut setelah seluruh gen-gen aditif terfiksasi dan juga pencatatan yang dilakukan setiap generasi memerlukan banyak tenaga dan ketelitian yang tinggi dalam mencatat dan menyimpan data silsilah.
Metode silang balik (back cross) diterapkan dengan tujuan memasukkan satu atau dua karakter pada varietas atau genotipe yang sudah mempunyai daya adaptasi atau karakter agronomi yang sudah baik (Stoskopf et al. 1993; Roy 2000; Chahal and Gosal 2003; Sutjahjo et al. 2006). Metode silang balik bertujuan untuk memperbaiki kultivar yang sudah mempunyai karakter agronomi dan adaptasi yang baik, tetapi kekurangan satu atau beberapa karakter saja, misalnya pemuliaan untuk memperbaiki resistensi terhadap penyakit dari varietas unggul komersial yang diadopsi luas. Tujuan dari silang balik berulang dengan tetua recurrent adalah untuk meningkatkan proporsi gen tetua recurrent. Metode silang balik dilaksanakan dengan menyilangkan kembali F1 dengan tetua yang mempunyai sifat agronomi baik sebagai tetua berulang (recurrent parent) untuk beberapa generasi. Untuk itu metode silang balik memerlukan tetua recurrent dengan sifat agronomi baik. Umumnya tetua recurrent merupakan varietas unggul komersial yang diadopsi secara luas oleh petani. Metode silang balik hanya memperbaiki satu sifat tetapi tidak dapat meningkatkan potensial hasil dari varietas yang ada.
Metode bulk dan single seed descent umum digunakan untuk seleksi pada karakter kuantitatif atau karakter yang mempunyai nilai heritabilitas yang
tergolong rendah atau sedang sehingga seleksi dilakukan pada generasi lanjut yaitu F5 atau F6. Prinsip kedua metode ini adalah upaya menangani populasi bersegregasi selama beberapa generasi secara bersama-sama sampai mencapai tingkat homozigositas yang diinginkan sebelum melakukan seleksi terhadap individu tanaman. Generasi F1 sampai F4 pekerjaan tidak terlalu berat, karena
pada generasi tersebut tidak dilakukan seleksi. Dalam metode single seed descent hanya satu benih yang diambil secara acak dari setiap tanaman untuk ditanam pada generasi berikutnya (Stoskopf et al. 1993; Roy 2000; Chahal dan Gosal 2003; Sutjahjo et al. 2006).
Metode untuk Mempelajari Pola Pewarisan Sifat
Karakter-karakter yang diekspresikan tanaman dibedakan atas karakter kualitatif dan karakter kuantitatif. Karakter kualitatif seperti warna dan bentuk bunga, bentuk dan warna biji, penampilannya sangat sedikit dipengaruhi oleh faktor lingkungan. Karakter-karakter ini akan mempunyai penampilan yang tetap pada berbagai lingkungan yang berbeda (Stoskopf 1993). Hal ini dapat terjadi karena karakter-karakter ini dikendalikan oleh gen-gen yang mempunyai pengaruh yang sangat besar terhadap penampilan/fenotipe (gen mayor), sehingga pengaruh lingkungan terhadap karakter tersebut kecil. Karakter kualitatif memiliki keragaman yang dapat dengan mudah dikelaskan dan pewarisan karakternya mengikuti hukum pewarisan sifat Mendel (Roy 2000) yaitu menurut Hukum Segregasi, di mana alel-alel dari pasangan gen bersegregasi (berpisah) satu dengan lainnya ke dalam gamet dan Hukum Perpaduan Bebas yaitu pada waktu pembentukan gamet, salah satu pasangan gen berpadu secara bebas dengan pasangan gen lainnya. Fenotipe dari karakter kualitatif dapat dikelaskan dengan jelas.
Keragaman karakter kualitatif dapat dibedakan berdasarkan aksi gen yang mengendalikan karakter tersebut (Stoskopf 1993; Chahal dan Gosal 2002). Karakter kualitatif dapat dikendalikan oleh satu gen yang mempunyai aksi dominan, over dominan atau kodominan, yang merupakan bentuk interaksi alela dalam lokus/gen yang sama. Dominansi adalah bentuk interaksi antar alel yang
![Gambar 1 Bagan Alir Penelitian Studi Pewarisan Toleransi Al Tanaman Sorgum Manis [Sorghum bicolor (L.) Moench]](https://thumb-ap.123doks.com/thumbv2/123dok/4584063.3340032/24.918.121.772.134.1064/gambar-penelitian-pewarisan-toleransi-tanaman-sorgum-sorghum-bicolor.webp)