Summary Beech seedlings (Fagus sylvatica L.) were grown in various combinations of three photosynthetic photon flux densities (PPFD, 0.7, 7.3 or 14.5 mol m−2 day−1) for two years in a controlled environmental chamber. Dry mass of leaves, stem and roots, leaf area and number of leaves, and unit leaf rate were affected by both previous-year and current-year PPFD. Number of shoots and length of the main shoot were affected by previous-year PPFD but not by current-year PPFD. Number of leaves per shoot did not change with PPFD, whereas leaf dry mass/leaf area ratio was mainly affected by current-year PPFD. During the first 10 days that newly emerged seedlings were grown at a PPFD of 0.7 or 14.5 mol m−2 day−1, transpiration rate per unit leaf area declined. Thereafter, tran-spiration increased to a constant new rate. Trantran-spiration rate per seedling was closely related to leaf area but the relationship changed with time. In two-year-old seedlings grown at various PPFD combinations of 0.7, 7.3 and 14.5 mol m−2 day−1 during Years 1 and 2, leaf area and transpiration rate per seedling were closely correlated at Weeks 7 and 11 after bud burst. Weak correlations were found between root dry mass and transpira-tion rate per seedling. During Year 2, transpiratranspira-tion rate per leaf area was higher at a particular PPFD in seedlings grown at a previous-year PPFD of 0.7 mol m−2 day−1 than in seedlings grown at a previous-year PPFD of 14.5 mol m−2 day−1. After transfer of two-year-old seedlings at the end of the experiment to a new PPFD (7.3 or 14.5 mol m−2 day−1) for one day, transpiration rates per leaf area, measured at the new PPFD, were correlated with leaf area and root dry mass, irrespective of former PPFD treatment.
Keywords: beech, irradiance, seedling morphology, shading.
Introduction
During natural regeneration, growth of beech seedlings may be limited by the availability of light because, although seedlings grow at irradiances as low as 1--2% of the light above the canopy, maximum dry matter production is only reached under unshaded conditions (Watt 1923, Suner and Röhrig 1980, Madsen 1994). The morphology of beech seedlings, including numbers of leaves, leaf area, branching and length of the main shoot, is also affected by the light conditions (Burschel and
Huss 1964, Burschel and Schmaltz 1965). Seedlings in the understory may experience variation in irradiance as a result of changes in foliage in the upper canopy. In addition, the seed-lings may be shaded by fast-growing ground vegetation. The influence of shelterwood and surrounding vegetation on the light environment may affect not only growth and morphology but also transpiration. According to Stickan and Zhang (1992), transpiration in beech is influenced more by PPFD than by vapor pressure deficit. A relationship between transpiration and dry matter production (water use efficiency) can be deter-mined for a seedling; however, it varies with environmental conditions (Davies and Pereira 1992). Furthermore, in young Douglas-fir seedlings, transpiration rates are influenced not only by current environmental conditions but also by previous growing conditions (Unterscheutz et al. 1974).
The aim of this work was to: (i) investigate the growth, morphology and transpiration of beech (Fagus sylvatica L.) seedlings at various PPFDs including a change in PPFD be-tween seasons; and (ii) establish the relationship bebe-tween transpiration and growth in young beech seedlings under vari-ous light conditions and at varivari-ous developmental stages of the seedlings.
Materials and methods
Plant material
Seeds of Fagus sylvatica were collected from trees of unknown origin at the end of September 1989, in Alnarp, southern Sweden. All seeds were vernalized at 4 °C for 5 weeks and then dried at 21 °C until their water content was about 5%. The dried seeds were stored in plastic bags at 4 °C. Eight weeks later, the seeds were soaked in tap water for 3 h and placed under a plastic cover at 5 °C for 5 weeks. The seeds were redried to about 5% water content and stored at 4 °C. After 8 weeks, the seeds were mixed with moist perlite and germi-nated at 4 °C. Seeds that germinated within 3 weeks (i.e., when about 5 mm of the root was visible) were planted in 10-cm-di-ameter plastic pots containing a 3/1 (v/v) mix of fertilized compost soil and perlite. The pots were keptat 10°C. When the shoot emerged from the soil, the pots were transferred to the various experimental conditions. The pots were irrigated
Influence of photosynthetic photon flux density on growth and
transpiration in seedlings of
Fagus sylvatica
N. T. WELANDER
1and B. OTTOSSON
21
Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, Box 49, S-230 53 Alnarp, Sweden
2 Swedish University of Agricultural Sciences, Department of Horticultural Science, Box 55, S-230 53 Alnarp, Sweden
Received August 28, 1995
alternately with demineralized water and a commercial nutri-ent solution (diluted to give N 15.9, P 2.2, K 8.3, Mg 1.7, S 1.1, Fe 0.031, Mn 0.013, Cu 0.011, Zn 0.0053, B 0.016, Mo 0.00058, Co 0.00035 mM in the final solution) throughout the experiment. Plants were irrigated once a week in the 0.7 mol m−2 day−1 PPFD treatment, and plants in 7.3 and 14.5 mol m−2 day−1 were irrigated twice a week. In a preliminary experi-ment, we ascertained that the effects of the light treatments on plant growth and development were not limited by the avail-ability of nutrients.
Experiment 1
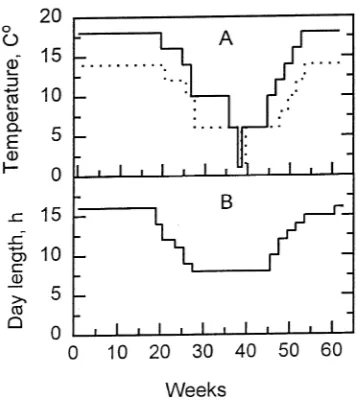
Newly emerged beech seedlings in pots were placed in a controlled environment chamber providing a 16-h photoperiod and a relative air humidity of about 80% throughout the day (day/night; 1.69/1.25 kPa). Twenty seedlings were used for each PPFD treatment. The PPFD treatments applied were 0.7, 7.3 or 14.5 mol m−2 day−1 corresponding to about 2, 21 and 43% relative light (total PPFD per day) under field conditions in southern Sweden. Light was supplied from cool-white fluo-rescent lamps (Sylvania 215 W). The various irradiances were attained by regulating the distance between the seedlings and the lamps, or by using shade cloth. Irradiance (PPFD) was measured with a quantum meter (LI-189, Li-Cor, Inc., Lincoln, NE) at the top of the plants. The various irradiances were measured and adjusted every second week. The variation in PPFD among plants was about 10% more or less than the given values. Irradiance was increased in stages during the first two hours of the day, and was decreased in a similar way at the end of the day. Peak irradiances during the middle 12 h of the photoperiod were 15, 150 and 300 µmol m−2 s−1, for the 0.7, 7.3 or 14.5 mol m−2 day−1 PPFD treatments, respectively. The temperature was 14 ± 0.2 °C during the night and rose continu-ously to 18 ± 0.2 °C during the first hour of light. A similar decrease in temperature occurred at the end of the day. After 9 weeks, the light and the temperature conditions were changed according to Figure 1 to induce dormancy. Twenty-one weeks later, a second growing period was induced by gradually in-creasing day length and temperature (Figure 1). At the start of the second growing period, seedlings from the 0.7 mol m−2 day−1 PPFD treatment were redistributed among the 0.7, 7.3 and 14.5 mol m−2 day−1 PPFD treatments. Similarly, plants from the 7.3 and 14.5 mol m−2 day−1 PPFD treatments were also redistributed among all of the PPFD treatments. The experiment was ended after 62.5 weeks.
At the end of the experiment, dry mass of stem, leaves and root were determined for each seedling. In addition, the num-bers of shoots and leaves, leaf area, seedling height, and length of the main shoot were recorded. The ratio between increase in dry mass per seedling and leaf area (unit leaf rate; ULR), which was calculated as a measure of the productivity of the leaves, was determined for the second year by subtracting the dry mass of one-year-old seedlings, determined from a sample taken 14 weeks after planting in the first year. The dry mass of the sample did not include the dry mass of the leaves because no leaves were present at the onset of growth in the second year.
Leaf dry mass/leaf area per seedling was calculated, and leaf fresh mass per area was also determined in some treatments.
Experiment 2
Newly emerged beech seedlings in pots were grown at 0.7 and 14.5 mol m−2 day−1 (PPFD) in a 16-h photoperiod. Light, temperature and air humidity conditions were the same as described for the first 8-week period in Experiment 1. During the period of leaf expansion, transpiration rates were recorded over 24-h periods. When the recordings started, the first true leaves were about 8 and 20 mm long in the 0.7 and 14.5 mol m−2 day−1 PPFD treatments, respectively. Transpiration rate was measured by weighing the seedlings and pots to the nearest 10 mg. During the measurement period, each pot was placed in a polythene bag that was sealed around the stem of the seedling. At the start of the measurements, seedlings were rewatered to approximately the same mass. Between measure-ments the plastic bag was removed to ensure aeration of the roots. At the beginning and end of each measurement period, leaf area per seedling was determined by drawing the outline of the leaves and determining the area within the outline with a digital polar planimeter (HAFF No. 330, Gebr. HAFF GmbH., Pfronten, Germany). Daily transpiration rate, includ-ing transpiration in the dark, was expressed per unit leaf area. In addition, transpiration for the entire 56-day measuring pe-riod was calculated for each seedling as the sum of the daily transpiration rate of the measuring days and the interpolated values for those days when no measurements were made. Ten seedlings were used for each PPFD treatment.
Experiment 3
after bud burst. Transpiration rate was measured during three consecutive 24-h cycles. When the second measurement pe-riod was performed a month later (Week 11), no new shoot formation had occurred. Thus, all transpiration measurements were performed on shoots that had been initiated in the pre-vious year. During the second period (Week 11), the irradiance was changed daily (see Figure 7). The procedures for transpi-ration measurements and leaf area determination were the same as described for Experiment 2. After the second period of transpiration measurements, dry mass of leaves, stem and roots were determined. Correlations between increase in dry mass and total transpiration per seedling over the second grow-ing season were determined based on integrated values of transpiration rate per unit leaf area between Weeks 0, 7 and 11 and integrated leaf area between Weeks 0, 2, 7 and 11 weeks. Diurnal courses of transpiration were recorded in seedlings in the 0.7, 7.3 and 14.5 mol m−2 day−1 PPFD treatments to assess the proportion of transpiration during the light and dark peri-ods. Measurements were performed on a balance connected to a computer that stored the hourly values.
Statistic analysis
Analysis of variance was carried out using the SAS statistical analysis software (SAS Institute Inc. 1988). A two-tailed t-test (Cochran and Cox 1957) was used to test the difference be-tween means.
Results
Experiment 1
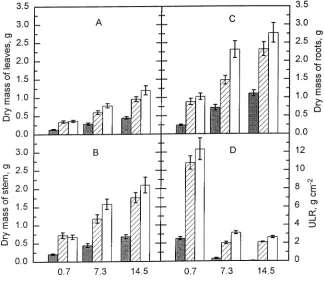
Both previous-year and current-year PPFD influenced leaf, stem and root dry mass at the end of the second growing season (P ≤ 0.0001), and there was an interaction between the PPFDs of the two seasons (P ≤ 0.02) (Figure 2A--C). When related to previous-year PPFD, dry mass of all plant parts increased with increasing PPFD. In the current season, dry mass of leaves, stem and roots were lower in seedlings grown at 0.7 mol m−2 day−1 than in seedlings grown at 7.3 or 14.5 mol m−2 day−1 (P ≤ 0.001). In two-year-old seedlings, ULR increased with increasing current-year PPFD (P ≤ 0.001) (Figure 2D), whereas it decreased with increasing previous-year PPFD.
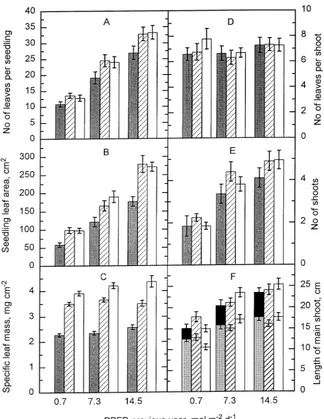
Number of leaves and leaf area per plant increased with increasing PPFD, but there was no interaction between the PPFDs of the two seasons(Figures 3A and 3B). Leaf area per seedling was closely correlated with number of leaves (r = 0.97), but less closely correlated with mean area per leaf (r = 0.71) (data not shown). The leaf dry mass/leaf area ratio increased with increasing current-year PPFD (P ≤ 0.001), whereas previous-year PPFD had no consistent effect (Fig-ure 3C). Leaf fresh mass per leaf area increased in a similar way as leaf dry mass with increasing current-year PPFD (data not shown).
Number of leaves per shoot was not affected by PPFD in either growing seasons (Figure 3D). Number of shoots per seedling was affected more by previous-year PPFD
Figure 2. Dry mass of (A) leaves, (B) stem, (C) roots, and (D) unit leaf rate (ULR) in two-year-old beech seed-lings exposed to various PPFD treat-ments during the previous and current year. Previous-year PPFDs are indicated on the x-axis and the current-year PPFDs are indicated by the shading in the columns: (j) 0.7,
(striped square) 7.3, and (h) 14.5
(P ≤ 0.0001) than by current-year PPFD (P ≤ 0.05). Fewer shoots developed in seedlings exposed to a previous-year PPFD of 0.7 mol m−2 day−1 than in seedlings exposed to previous-year PPFDs of 7.3 and 14.5 mol m−2 day−1. Current-year PPFD had no consistent effect on number of shoots per seedling (Figure 3E). Current-year and previous-year PPFD treatments had similar effects on seedling height and length of the main shoot as on number of shoots per seedling (P ≤ 0.01) (Figure 3F). Seedling height was less than the length of the main shoot as the main shoot was oblique or horizontally orientated at all PPFDs.
Experiment 2
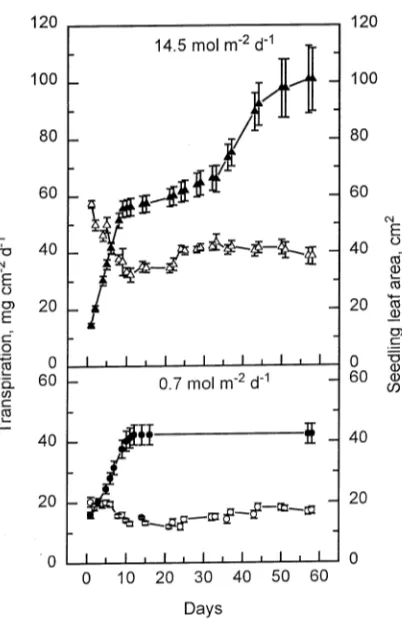
During the first 10 days that newly emerged seedlings were exposed to PPFDs of 0.7 or 14.5 mol m−2 day−1, transpiration rates per leaf area declined and then remained constant for a further 10 days (Figure 4). Thereafter, transpiration increased
to a new rate. During the 10-day period of decline in transpi-ration rate, expansion growth of the first two true leaves ended. At this time, the total leaf area per seedling comprised the cotyledons and these first two true leaves. About 20 days later, new leaves started to expand in seedlings in the 14.5 mol m−2 day−1 PPFD treatment. This expansion period started later than the increase in transpiration rate. In the 0.7 mol m−2 day−1 PPFD treatment, no leaves were formed after the first true leaves had expanded.
Close correlations were found between integrated transpira-tion rate and dry mass per seedling (Figure 5, r = 0.98). In the 14.5 mol m−2 day−1 PPFD treatment, there was a close corre-lation between leaf area per seedling and transpiration rate over a 24-h period, both during the first 10 days (r = 0.88) and between Days 25 and 60 (r = 0.87) (Figure 6) after seedling emergence. However, there was a change in the relationship between the two periods as indicated by the difference in the
slope of the regression line (P ≤ 0.001). A similar correlation between leaf area and transpiration rate was also observed in the 0.7 mol m−2 day−1 PPFD treatment (r = 0.85 for Days 1--10 and r = 0.66 for Days 25--60); however, the relationship (slope
of line) differed between the 0.7 and 14.5 mol m−2 day−1 PPFD treatments (P ≤ 0.001).
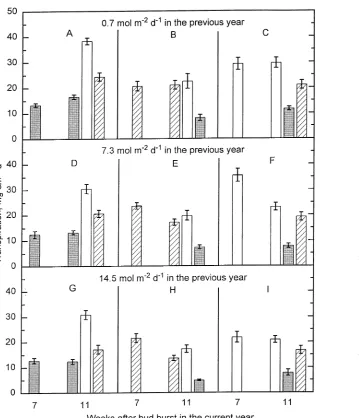
Experiment 3
Seven weeks after bud burst, transpiration rate per unit leaf area (measured at the same PPFD as the current-year PPFD treatment of the seedling) was compared in seedlings from all nine combinations of previous-year and current-year PPFD treatments. Transpiration rate increased with current-year PPFD (P ≤ 0.001) in seedlings from previous-year PPFD treatments of 0.7 and 7.3 mol m−2 day−1 (Figure 7A--C and D--F, 7 weeks). In seedlings from the previous-year PPFD treatment of 14.5 mol m−2 day−1, transpiration rates of seed-lings exposed to a current-year PPFD of 7.3 or 14.5 mol m−2 day−1 were higher than transpiration rates of seedlings exposed to a current-year PPFD of 0.7 mol m−2 day−1 (P ≤ 0.001) (Figure 7G--I). A close correlation was found between seedling leaf area and transpiration rate per seedling (not shown in figure) irrespective of previous-year PPFD (r = 0.90, 0.94 and 0.87 for 0.7, 7.3 and 14.5 mol m−2 day−1).
Eleven weeks after bud burst, rates of transpiration per unit leaf area were measured, first at the PPFD under which cur-rent-year leaf expansion took place, and second after a one-day transfer to a contrasting PPFD. Before the one-day transfer, rates of transpiration were similar at Weeks 7 and 11, in all seedlings exposed to a previous-year PPFD of 0.7 mol m−2 day−1 (Figure 7A--C), whereas in seedlings exposed to a pre-vious-year PPFD of 7.3 mol m−2 day−1, transpiration rates declined between Weeks 7 and 11 in the current-year 7.3 or Figure 4. Effects of PPFD on transpiration rate (open symbols) and
leaf area (closed symbols) during the first 56 days after beech seedling emergence. Standard errors of the mean are depicted by the vertical bars, n = 10.
Figure 5. Correlation between the sum of transpiration over 56 days and dry mass in beech seedlings grown at (s) 0.7 or (n) 14.5 mol m−2
day−1. Each symbol represents an individual seedling.
Figure 6. Correlation between seedling leaf area and transpiration rate per seedling during Days 1--10 (closed symbols) and Days 25--56 (open symbols) in seedlings grown at 0.7 (d and s) or 14.5 mol m−2
14.5 mol m−2 day−1 PPFD treatments (P ≤ 0.01) (Figure 7E--F). A similar decline was seen in the current-year 7.3 mol m−2 day−1 PPFD treatment for seedlings exposed to a previous-year PPFD of 14.5 mol m−2 day−1 (Figure 7G--H).
In seedlings from all nine combinations of previous-year and current-year PPFD treatments, the rate of transpiration per leaf area showed similar changes in relation to current-year PPFD treatment. However, the overall rate of transpiration recorded at a PPFD of 0.7 mol m−2 day−1 was higher in seedlings grown in current-year PPFD of 0.7 mol m−2 day−1 (Figure 7A, D and G, shaded bars, Week 11) than in seedlings grown in 7.3 and 14.5 mol m−2 day−1 (P ≤ 0.01) (Figure 7B, E, H and C, F, I, shaded bars, Week 11). In addition, the overall rate of transpiration was highest in seedlings exposed to a previous-year PPFD of 0.7 mol m−2 day−1 (Figure 7A--C, Week 11) and lowest in seedlings exposed to a previous-year PPFD of 14.5 mol m−2 day−1 (Figure 7G--I) (P ≤ 0.01).
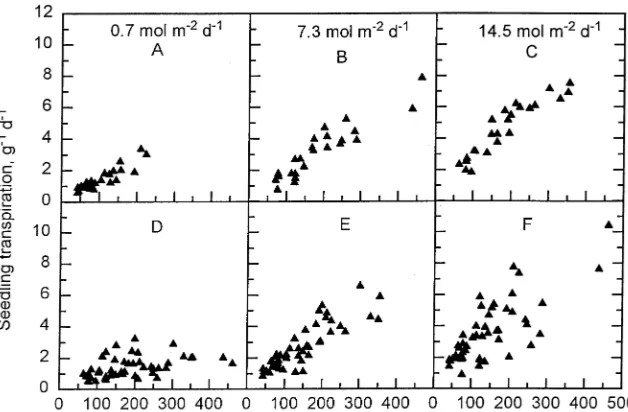
Close correlations were seen between leaf area and transpi-ration rate per seedling on the first day of Week 11 after bud burst in seedlings exposed to current-year PPFDs of 0.7 (r = 0.90), 7.3 (r = 0.92) and 14.5 mol m−2 day−1 (r = 0.93) (Figure 8A--C). Close correlations were also found between root dry mass and transpiration rate for seedlings exposed to current-year PPFDs of 0.7 (r = 0.80), 7.3 (r = 0.82) and 14.5 mol m−2 day−1 (r = 0.79)(data not shown).
When PPFD was changed daily (one-day PPFD treatment), the correlations between transpiration rate per seeedling and leaf area decreased on Days 2 and 3 compared to Day 1. Measurements at the one-day PPFD of 0.7 mol m−2 day−1 showed a weak correlation between transpiration rate and leaf area (r = 0.45) (Figure 8D, Week 11), but close correlations were found between leaf area and transpiration rate per seed-ling measured at a one-day PPFD of 7.3 (r = 0.76) or 14.5 mol m−2 day−1 (r = 0.86) (Figures 8E and 8F), irrespective of Figure 7. Influence of various combinations of PPFD on transpiration rate per unit leaf area at Weeks 7 and 11 after bud burst in two-year-old beech seedlings. The first col-umn represents the median of three consecutive days of measurements at Week 7. The following three columns rep-resent three consecutive sin-gle-day measurements at Week 11, when the PPFD was changed daily. The pre-vious-year PPFD was: (A--C) 0.7, (D--F) 7.3, and (G--I) 14.5 mol m−2 day−1. Current-year PPFD was (A, D and G) 0.7, (B,E and H) 7.3, and (C, F and I) 14.5 mol m−2 day−1. The PPFD on the day of measurement is indicated by the shading: stippled = 0.7; crosshatched = 7.3; and open = 14.5 mol m−2 day−1. Verti-cal bars represent ± SE;
previous-year PPFD or current-year PPFD during the leaf expansion period. Weak correlations were found for root dry mass and transpiration (r = 0.36, 0.79 and 0.76) when meas-ured at the one-day PPFDs of 0.7, 7.3 and 14.5 mol m−2 day−1 on Days 2 and 3 of Week 11 (data not shown).
Discussion
Current growth, morphology and transpiration in two-year-old beech seedlings were all affected by both previous- and cur-rent-year light conditions. For example, beech seedlings trans-ferred from a low previous-year PPFD to a higher current-year PPFD showed a higher ULR than seedlings maintained at the higher PPFD for both years, indicating that ULR was affected by both previous-year and current-year PPFD. In oak seedlings subjected to a similar transfer (Ziegenhagen and Kausch 1995) a decrease in productivity was found, showing that the re-sponse to a year to year change in light environment may vary among species.
The influence by PPFD on seedling morphology showed various patterns. Both leaf dry mass/leaf area and leaf fresh mass/leaf area were only affected by current-year PPFD. In contrast, Goulet and Bellefleur (1986) found that, in Fagus grandifolia trees, the light conditions during both bud initia-tion in the previous year and leaf outgrowth in the current year affected leaf fresh weight per area.
Leaf area was predetermined by the PPFD prevailing in the year of leaf initiation and final area was modified by the PPFD prevailing when the leaves expanded in the following year. Several morphological parameters appear to determine the leaf area per seedling. First, the area depends on the number of leaves as indicated by the close correlation between these parameters. Second, neither previous-year nor current-year PPFD influenced the number of leaves per shoot, indicating
that the number of leaves per beech seedling is determined by the number of shoots. In contrast, Hansen (1959) found that buds developed in full light contained more leaf initials than buds developed in the shade. Because the range of PPFDs we employed was less than in the study of Hansen (1959), it is possible that the difference between the highest and lowest PPFDs in our study was not large enough to affect the rate of leaf initiation significantly. Moreover, in our study, fewer leaves developed in plants at the lowest current-year PPFD than in plants at the highest current-year PPFD even when seedlings had been exposed to the same previous-year PPFD, indicating that either some leaf initials from the previous year did not develop, or new leaves were initiated during shoot elongation in the current year. The number of shoots that developed in the current season was also determined more by the PPFD of the previous year when the shoot buds were initiated than by current-year PPFD.
Transpiration rate per unit leaf area was influenced by PPFD directly during the transpiration period, and indirectly by the PPFD when the leaves were expanding, and during the year when the leaves were initiated. Although transpiration rate per unit leaf area increased with increasing PPFD, the effect was less marked as the leaves became older. The decrease with leaf age was not the result of reduced light caused by increased self-shading because no new leaves were formed during this period. Federer (1976) reported that leaf age appears to influ-ence the effect of light on transpiration in various deciduous tree species.
the developing leaf rather than on leaf developmental stage. In Year 2, seedlings exposed to high previous-year PPFDs showed lower transpiration rates per unit leaf area than seed-lings exposed to low previous-year PPFDs, indicating that some light effects are long lasting. In the year of seedling emergence, integrated transpiration, based on transpiration measurements at close intervals, was closely correlated to dry mass after a 56-day period of growth under conditions of constant irradiance. However, the correlation was less close when integrated transpiration was based on only two measur-ing times in the year and when PPFD was changed between the two growing seasons, indicating that accurate estimation of water use efficiency under natural conditions where environ-mental conditions fluctuate requires repeated measurements of transpiration at close intervals.
We have demonstrated that previous environmental condi-tions need to be taken into account when studying the influences of current environmental parameters on growth, morphology and transpiration in beech seedlings. These findings have par-ticular relevance for comparative studies of light effects on current growth and for the study of nursery-grown plant mate-rial under field conditions.
Acknowledgment
We thank Dr. Olof Hellgren for valuable comments on the manuscript.
References
Burschel, P. and J. Huss. 1964. Die Reaktion von Buchensämlingen auf Beschattung. Forstarchiv. 35:225--233.
Burschel, P. and J. Schmaltz. 1965. Die Bedeutung des Lichtes für die Entwicklung junger Buchen. Allg. Forst- u. Jagdztg. 136:193--210. Cochran, W.G. and G. M. Cox. 1957. Experimental design. John Wiley
and Sons, Inc., NY, pp 95--102.
Davies, W. J. and J.S. Pereira. 1992. Seedling growth and water use efficiency. In Crop Photosynthesis: Spatial and Temporal Determi-nants. Eds. N.R. Baker and H. Thomas. Elsevier Science Publishers b.v., The Netherlands, pp 213--233.
Federer, C.A. 1976. Differing diffusive resistance and leaf develop-ment may cause differing transpiration among hardwoods in spring. For. Sci. 22:359--364.
Goulet, F. and P. Bellefleur. 1986. Leaf morphology plasticity in response to light environment in deciduous tree species and its implication on forest succession. Can. J. For. Res. 16: 1192--1195. Hansen, H.C. 1959. Einfluss des Lichtes auf die Bildung von
Licht-und Schattenblättern der Buche, Fagus silvatica. Physiol. Plant. 12:545--550.
Madsen, P. 1994. Growth and survival of Fagus sylvatica seedlings in relation to light intensity and soil water content. Scand. J. For. Res. 9:316--322.
Stickan, W. and X. Zhang. 1992. Seasonal changes in CO2 and H2O
gas exchange of young European beech (Fagus sylvatica L.). Trees 6:96--102.
Suner, A. and E. Röhrig. 1980. Die Entwicklung der Buchennaturver-jungung in Abhängigkeit von der Auflichtung des Altbestandes. Forstarchiv. 51:145--149.
Turner, N.C. and G.H. Heichel. 1977. Stomatal development and seasonal changes in diffusive resistance of primary and regrowth foliage of red oak (Quercus rubra L.) and red maple (Acer ru-brum L.). New Phytol. 78:71--81.
Unterscheutz, P., W.F. Reutz, R.R. Geppert and W.K. Ferrell. 1974. The effect of age, pre-conditioning, and water stress on transpira-tion rates of Douglas-fir (Pseudotsuga menziesii) seedlings of sev-eral ecotypes. Physiol. Plant. 32:214--221.
Watt, A.S. 1923. On the ecology of British beechwoods with special reference to their regeneration. J. Ecol. 11:1--48.